 2025, Vol. 45
2025, Vol. 45文章信息
- 罗健梅, 阿布都热合曼·哈力克, 段越帆, 姚凯旋, 姚磊, 唐华, 布威阿依谢姆·吐合提
- LUO Jianmei, Abudureheman·Halike, DUAN Yuefan, YAO Kaixuan, YAO Lei, TANG Hua, Buweiayixiemu·Tuheti
- 艾比湖流域植被NPP时空演变特征及其驱动因素
- Spatiotemporal dynamics and driving factors of net primary productivity in the Ebinur Lake Basin
- 生态学报. 2025, 45(1): 182-196
- Acta Ecologica Sinica. 2025, 45(1): 182-196
- http://dx.doi.org/10.20103/j.stxb.202401240224
-
文章历史
- 收稿日期: 2024-01-24
- 网络出版日期: 2024-09-23

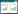




2. 新疆大学绿洲生态重点实验室, 乌鲁木齐 830017;
3. 新疆大学智慧城市与环境建模自治区普通高校重点实验室, 乌鲁木齐 830017
2. Xinjiang Key Laboratory of Oasis Ecology, Xinjiang University, Urumqi 830017, China;
3. Key Laboratory of Smart City and Environment Modelling of Higher Education Institute, Xinjiang University, Urumqi 830017, China
植被净初级生产力(Net Primary Productivity, NPP)是指绿色植物在单位时间、单位面积上由光合作用产生的有机物质的净积累量[1]。植被NPP是植被自身生长状况、生命活动、固碳能力的重要表征, 同时也是评价生态系统质量状况、功能状况的重要指标[2], 对研究陆地生态系统可持续发展具有重要的意义。
植被NPP测定方法主要有传统测量法和模型估算法, 传统实地测量虽然精度较高, 但只适用于小范围研究, 模型估算可以实现大范围动态监测[3—4]。Lieth等建立了第一个生态系统NPP遥感估算模型(Miami模型)[5], 发展至今, 形成了许多植被NPP估算模型, 主要有气候生产力模型、过程模型和光能利用率模型[6]。CASA(Carnegie-Ames-Stanford Approach)模型是目前估算植被NPP的主流模型之一, CASA模型输入参数少, 遥感数据丰富且易获取, 在长时间和大尺度上的有效性和准确性已得到验证[7—8]。
近年来, 国内外学者对植被NPP开展了大量的研究, 这些研究集中在NPP估算、时空演变规律及驱动因素分析等方面[9—10], 涵盖了全球[11]、国家[12]、省域[13]及流域[14]等不同尺度。植被NPP受多种因素的影响, 对气候变化的响应尤为敏感, 在大多数研究中将植被指数变化归因于气候变化[15]。自然因素对植被NPP空间分布具有重要影响, 在不同海拔、坡度、坡向以及地形起伏条件下, 水热条件和土壤养分存在差异, 进而影响植被的生长和分布[16]。此外, 人类活动也对植被NPP变化带来不同程度的影响, 森林砍伐、过渡放牧以及城市扩张等造成植被NPP下降, 2000—2010年城市扩张导致全球陆地NPP每年损失22.4 TgC[17];而实施植树造林、退耕还林等生态工程项目, 可有效提升生态系统植被生产力[18], 退耕还林计划和造林工程的实施促进了中国的绿化趋势[19]。
艾比湖流域是新亚欧大陆桥国土安全的重要屏障, 也是丝绸之路经济带核心区建设的战略支点[20]。流域具有典型的干旱区环境特征, 受气候变化的影响和人类活动的干扰, 流域生态环境脆弱, 保障其生态环境健康稳定至关重要[21]。目前, 关于艾比湖流域植被NPP的研究尚少, 朱旻[22]和赵鹏[23]的研究侧重于NPP时空变化及其对气候因子的响应, 前人更多聚焦于艾比湖流域NDVI时空变化及其驱动因子研究。孙倩[24]、赵坤成[25]、任立清[26]等探究了不同时段艾比湖流域NDVI的时空变化特征, 揭示了气象因子、地形、植被、土壤以及土地利用类型等因子对艾比湖流域NDVI的影响。虽然NDVI和NPP都是评价植被和生态系统的重要指标, 但二者侧重点有所不同, NDVI主要反映植被的覆盖程度和生长状态, 而NPP能衡量植物在一定时间内通过光合作用固定的碳量[27]。艾比湖流域地形、植被类型、土壤类型等自然因子和人为因子及其交互作用对植被NPP的影响, 以及找出各因子中最适合植被生长的范围或类型等问题有待进一步探究。鉴于此, 本文基于CASA模型模拟艾比湖流域2001—2020年的植被NPP, 分析植被NPP时空分异特征, 探究自然因素及人类活动对植被NPP的影响, 找出植被生长的最佳条件, 以期为艾比湖流域可持续发展提供参考依据。
1 研究区概况艾比湖流域(79°89′—85°38′E, 43°38′—45°52′N)位于新疆西北部, 深居大陆腹地, 海拔高度为183—4874 m, 流域面积为5.03×104 km2(图 1)。三面环山阻隔外界气流, 流域内降水稀少、日照充足, 蒸发量大, 多年平均降水量为100—200 mm, 多年平均蒸发量为1500—2000 mm[28]。艾比湖流域地貌类型复杂, 植被类型以荒漠、草原和草甸为主, 土壤类型主要为钙层土、漠土、高山土和干旱土, 草地和未利用地是流域内主要的土地利用类型[26]。艾比湖流域是亚欧大陆重要的交通枢纽, 也是北疆重要的粮食基地、棉花基地和畜牧业基地。

|
| 图 1 研究区概况图 Fig. 1 An overview of the study area |
本研究利用的NDVI数据为2001—2020年MOD13Q1数据集, 空间分辨率为250 m, 时间分辨率为16 d, 采用最大值合成法(Maximum Value Composite, MVC)生成月最大NDVI数据;月太阳辐射数据采用TerraClimate数据集, 植被类型数据采用MCD12Q1产品, MOD17A3 NPP数据用于CASA模型精度验证, 以上数据均来自Google Earth Engine(https://earthengine.google.com/)平台。月平均气温和月降水数据来自国家青藏高原科学数据中心(https://data.tpdc.ac.cn/), 空间分辨率0.0083333°(约1 km), 高程数据分辨率为30 m, 来源于地理空间数据云(https://www.gscloud.cn/), 坡度和坡向从高程中提取。土壤类型数据采用1∶100万中华人民共和国土壤数据;GDP和人口密度数据分辨率为1 km, 土地利用数据分辨率为30 m, 均来在中国科学院资源环境科学与数据中心(https://www.resdc.cn/)。将所有数据投影坐标系转换为WGS_1984_UTM_Zone_44N, 空间分辨率重采样为500 m。
2.2 研究方法 2.2.1 NPP估算CASA模型是目前广泛运用于植被NPP估算的光能利用率模型, 本文采用朱文泉等[29]改进的CASA模型, 公式为:
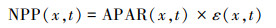
|
(1) |
式中, APAR(x, t)表示像元x在t月植被吸收的光合有效辐射, ε(x, t)表示像元x在t月植被的实际光能利用率。
2.2.2 热点分析基于全局Moran′s I指数判断植被NPP在空间上是否存在集聚特征, 公式为[30]:
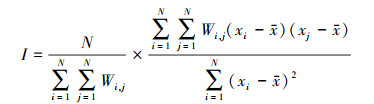
|
(2) |
式中, N为像元数量;Wi, j为标准化的空间权重矩阵, xi和xj为像元i和像元j的NPP值, x为NPP均值, I大于0为正相关, 空间分布具有集聚特征, I为0表示随机分布。
采用Getis-Ord Gi*热点分析探究植被NPP局部空间自相关特征, 公式为:
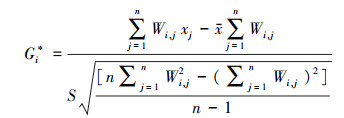
|
(3) |
式中, n为像元总数, Wi, j为标准化的空间权重矩阵, xi和xj表示像元i和像元j的NPP值,x为NPP均值, S为标准差。Gi*为统计值, 值越大表示热点集聚程度越高。
2.2.3 趋势分析Theil-Sen Median方法属于非参数统计方法, 能够排除时间序列中异常值干扰, 适合长时间序列数据的趋势分析。公式为[31]:
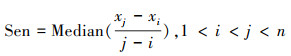
|
(4) |
式中, Sen表示变化斜率, xi和xj表示第i年和第j年的变量值。
为了进一步分析变化趋势的显著性, 采用Mann-Kendall显著性检验。给定显著性水平α, |Z|>Z(1-α)/2时, 时间序列数据存在显著性变化趋势。当|Z|大于1.65、1.96和2.58时, 分别表示通过置信度为90%、95%、99%显著性检验。
2.2.4 变异系数变异系数(Coefficient of Variation, CV)主要用于检验数据在时间尺度上的波动情况, 变异系数越小表示稳定性越高[32]。公式为:
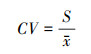
|
(5) |
式中, CV为变异系数;S表示标准差;x表示平均值。将变异系数划分为高稳定(CV≤0.1)、稳定(0.1<CV≤0.2)、不稳定(0.2<CV≤0.3)、高波动(CV>0.3)4个等级。
2.2.5 地理探测器地理探测器是探测空间分异性并揭示其背后驱动力的一组统计学方法[33]。传统地理探测器存在主观性和离散化不佳的问题, 因此, 本研究采用参数最优地理探测器(Optimal Parameters-based Geographical Detector, OPGD), 借助R语言完成因子探测、交互作用探测及风险探测[34]。因子探测主要通过因子解释力的大小来衡量单个自变量对因变量空间分异的影响程度, 交互作用探测用于分析两个自变量作用力叠加后, 对因变量的作用力是否增强, 风险探测用于寻找植被NPP值高的区域。q值计算公式为:
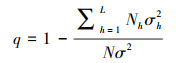
|
(6) |
式中, q为因子解释力, 范围为[0, 1], 值越大解释力越强;h表示对自变量和因变量的分区数目, N和Nh表示整个研究区和分区后h区内的样本数和;σ2、σh2表示整个研究区和分区后h区内的NPP方差。
3 结果 3.1 模型精度验证由于缺乏实测数据, 本文利用MOD17A3数据验证CASA模型模拟结果的准确性, 大量学者已经证明这种验证法具有一定的可靠性[35—36], MOD17A3产品数据已在全球不同地区研究中得到广泛的验证和应用[37—38]。基于CASA模型模拟的NPP值与MOD17A3 NPP数据创建1 km×1 km的渔网, 提取渔网中心点像元值, 剔除异常值和空值, 对剩余34938组数据进行相关性分析(图 2), R2为0.79(P<0.01), 相关性较强, 总体而言模拟结果可靠。

|
| 图 2 模拟结果精度验证 Fig. 2 Accuracy verification of simulation results NPP:净初级生产力Net Primary Productivity;CASA:卡内基-艾姆斯-斯坦福方法Carnegie-Ames-Stanford Approach |
艾比湖流域20年的植被NPP均值介于0—802.26 gC m-2 a-1之间, 整体呈四周高中间低的空间分布格局(图 3)。NPP值大于500 gC m-2 a-1的区域占研究区总面积的3.78%, 分布在西部及南部的山区;NPP值在200—500 gC m-2 a-1之间的区域占39.98%, 主要分布在河流沿岸的绿洲;NPP值小于200 gC m-2 a-1的区域占56.24%, 广泛分布在研究区中部。全局Moran′s I指数为0.85(P<0.01), 表明艾比湖流域植被NPP分布具有显著的空间自相关特征, 进一步对植被NPP的冷热点区域进行识别, 揭示植被NPP的空间集聚规律。空间集聚类型以99%置信水平为主, 其中, 热点区域和冷点区域占比均为36.33%。NPP高值区在空间上聚集形成热点区, 分布在研究区西部、南部及北部河流上游的山区以及河流沿岸的绿洲, NPP低值区在空间上聚集形成冷点区, 主要分布在研究区中部的未利用地。

|
| 图 3 艾比湖流域植被NPP多年平均值和冷热点区空间分布 Fig. 3 Spatial distribution of the average NPP and cold hot spots in the Ebinur Lake Basin |
2001—2020年艾比湖流域植被NPP年际变化整体呈波动增加趋势(图 4), 增长率为1.83 gC m-2 a-1(R2=0.21, P<0.05), 多年平均值为199.67 gC m-2 a-1, 最小值为163.46 gC m-2 a-1, 出现在2001年, 最大值为257.27 gC m-2 a-1, 出现在2016年。2015—2016年NPP增幅最大, 变化幅度为72.48 gC m-2 a-1, 2016—2017年NPP降幅最大, 为55.25 gC m-2 a-1。

|
| 图 4 2001—2020年艾比湖流域植被NPP年际变化 Fig. 4 Interannual change of NPP in the Ebinur Lake Basin from 2001 to 2020 |
2001—2020年艾比湖流域植被NPP呈增加趋势的面积占61.79%, 表明大部分地区植被处于生长或恢复状态(图 5)。其中, 极显著增加(P<0.01)和显著增加(P<0.05)的区域占20.44%, 主要分布在河流沿岸的耕地区域;26.32%的区域呈下降趋势, 极显著减少(P<0.01)和显著减少(P<0.05)的区域零星分布在建设用地扩张区;11.89%的区域植被NPP几乎没有发生变化, 主要分布在西部及南部的山区。植被NPP变异系数范围在0.01—4.12之间, 平均值为0.33(图 6)。高稳定区(CV≤0.1)和稳定区(0.1<CV≤0.2)面积占32.02%, 主要分布在山区及河流沿岸的绿洲, 不稳定区(0.2<CV≤0.3)主要分布在绿洲外围, 45.18%的区域变异系数大于0.3, 植被NPP波动变化较大。

|
| 图 5 2001—2020年艾比湖流域植被NPP空间变化趋势及显著性检验 Fig. 5 Spatial variation trend and significance test of NPP in the Ebinur Lake Basin from 2001 to 2020 |

|
| 图 6 艾比湖流域植被NPP变异系数空间分布 Fig. 6 Spatial distribution of variation coefficient in the Ebinur Lake Basin |
气候变化直接影响植被光合、呼吸和蒸腾等代谢过程, 进而影响植被NPP变化。如表 1和图 7所示, 植被NPP与气象因子整体呈正相关, 植被NPP与气温偏相关系数为0.17, 呈正相关的区域占75.23%, 其中, 18.23%的区域呈极显著正相关与显著正相关, 主要分布在西部及南部的耕地和草地区域;24.77%的区域呈负相关, 极显著负相关和显著负相关的区域占0.74%, 分布在东北部植被稀疏区域。植被NPP与降水偏相关系数为0.38, 92.77%的区域呈正相关, 其中, 极显著正相关与显著正相关的区域占53.99%;呈负相关的区域分布在东部建设用地和未利用地区域。植被NPP与太阳辐射偏相关系数为0.14, 呈正相关的区域占67.25%, 其中, 21.39%的区域呈极显著正相关与显著正相关, 主要分布在耕地区域;32.75%的区域呈负相关, 其中, 极显著负相关和显著负相关的区域占2.74%, 零散分布在研究区北部和东部。
| 相关性 Correlation |
气温偏相关 Partial correlation of temperature |
降水偏相关 Partial correlation of precipitation |
太阳辐射偏相关 Partial correlation of solar radiation |
| 极显著正相关(P<0.01) Highly significant positive correlation(HSPC) |
6.90 | 30.07 | 11.41 |
| 显著正相关(P<0.05) Significant positive correlation(SPC) |
11.33 | 23.92 | 9.98 |
| 不显著正相关 No significant positive correlation(NSPC) |
57.00 | 38.78 | 45.86 |
| 不显著负相关 No significant negative correlation(NSNC) |
24.03 | 6.93 | 30.01 |
| 显著负相关(P<0.05) Significant negative correlation(SNC) |
0.64 | 0.25 | 2.18 |
| 极显著负相关(P<0.01) Highly significant negative correlation(HSNC) |
0.10 | 0.05 | 0.56 |

|
| 图 7 植被NPP与气象因子的偏相关分析及显著性检验 Fig. 7 Partial correlation coefficients and significance test of NPP with meteorological factors HSPC:极显著正相关(P<0.01);SPC:显著正相关(P<0.05);NSPC:不显著正相关;NSNC:不显著负相关;SNC:显著负相关(P<0.05);HSNC:极显著负相关(P<0.01) |
不同地形条件水热分配存在差异, 影响植被NPP的积累和空间分布。本研究以50 m高程和2°坡度为间隔, 分析植被NPP随高程和坡度的变化特征, 标准差表示变化幅度。坡向对植被NPP的影响不明显, 随海拔升高, NPP值呈先增加后减少的变化趋势(图 8)。海拔低于1000 m的区域主要分布天然草地和栽培植被, NPP值在34.11—203.97 gC m-2 a-1之间, 海拔在2300—2500 m之间NPP值最大, 海拔超过2500 m, 气温下降甚至有冰雪覆盖, 环境条件不适合大多数植被生长, NPP值不断降低。随坡度增加, NPP值总体呈先上升后下降的变化趋势。坡度小于6°的区域NPP均值为172.62 gC m-2 a-1, 主要分布耕地、草地和未利用地, 坡度6°—25°的区域主要分布草地和灌丛, 随坡度增加植被NPP小幅上升;坡度大于25°, 土壤侵蚀增强, 保水保肥能力下降, 植被NPP呈下降趋势。

|
| 图 8 植被NPP均值随高程和坡度的变化 Fig. 8 The Variation of NPP mean values with elevation and slope |
不同植被生理特征存在差异, 积累NPP的能力不同, 不同植被类型NPP均值大小为:混交林>阔叶林>针叶林>栽培植被>草甸>灌丛>草原>荒漠(图 9)。森林具有较高的生物多样性和复杂的生态系统结构, 固碳能力较强, 森林NPP值最高, 研究区森林覆盖面积较小;栽培植被在科学的人为管理下, 年际变化较稳定;草原是主要的植被类型, 承担了研究区近90%的固碳量。不同土壤类型供给植被的土壤肥力和营养物质不同, 进而影响植被生长状况和NPP空间分异, 不同土壤类型NPP值大小为:半淋溶土>人为土>半水成土>高山土>钙层土>水成土>盐碱土>干旱土>漠土>初育土。半淋溶土拥有较好的土壤肥力和水分保持能力, 适合森林、灌丛生长, NPP值最高;研究区36.97%的面积分布高山土和钙层土, 高山土有机质积累量大, 钙层土养分供给能力较强, NPP值较高;研究区中部和东部分布盐碱土、干旱土、漠土、初育土, 保水保肥能力较差, 易受到侵蚀和人为破坏, 不利于植被生长。

|
| 图 9 不同植被类型和不同土壤类型的NPP均值 Fig. 9 NPP mean values of different vegetation types and different soil types |
人类活动对植被NPP具有双重影响, 一方面, 为了满足人口增长和经济发展的需求, 城市建设、土地开发等人类活动会导致植被NPP下降;另一方面, 通过实施生态工程项目, 如植树造林、三北防护林工程等, 可以提升生态系统植被NPP。人类活动改变土地利用方式, 影响植物的生长和分布, 进而影响植被NPP变化。土地利用变化是人类活动影响自然环境的主要方式, 也是人类活动强度在空间上的直观体现[39]。如表 2所示, 本文通过土地利用变化造成NPP总量变化来表示人类活动对植被NPP的影响。
| 2001 | 2010 | |||||
| 耕地 | 林地 | 草地 | 水域 | 建设用地 | 未利用地 | |
| 耕地Cropland | — | 0.008 | 1.17 | -0.03 | -0.13 | -1.59 |
| 林地Forest | 2.99 | — | 0.26 | -0.18 | -0.06 | -0.02 |
| 草地Grassland | 124.17 | 1.83 | — | -0.29 | -1.12 | -2.61 |
| 水域Water area | 0.23 | 0.05 | 0.37 | — | 0.06 | 0.25 |
| 建设用地Construction land | 0.01 | 0.0006 | 0.002 | 0.0002 | — | 0.0002 |
| 未利用地Unused land | 52.64 | 0.24 | 1.34 | 0.42 | 0.15 | — |
| 2010 | 2020 | |||||
| 耕地 | 林地 | 草地 | 水域 | 建设用地 | 未利用地 | |
| 耕地Cropland | — | 1.32 | 12.87 | 0.63 | -1.98 | -0.78 |
| 林地Forest | 11.32 | — | 56.25 | 0.59 | -0.007 | -1.44 |
| 草地Grassland | 234.32 | 13.51 | — | 0.95 | 2.52 | 34.29 |
| 水域Water area | 2.02 | 0.26 | 6.39 | — | 0.15 | -7.51 |
| 建设用地Construction land | 2.32 | -0.01 | 0.35 | 0.15 | — | 0.11 |
| 未利用地Unused land | 120.13 | 0.52 | 94.69 | -1.58 | 3.16 | — |
2001—2010年、2010—2020年土地利用类型发生变化的面积分别占总面积的2.62%和24.04%, 整体表现为未利用地、林地和水域面积减少, 耕地、草地和建设用地面积增加(图 10)。艾比湖流域主要土地利用类型为草地和未利用地, 2001—2010年草地面积减少729.95 km2, 造成草地NPP减少1.19×108 kgC, 其中96.27%的面积转化为耕地;2010—2020年草地变化复杂, 有3815.99 km2草地转出, 同时5968.28 km2其他土地类型转为草地, 导致草地NPP变化1.51×108 kgC。2001—2010年未利用地面积减少401.94 km2, 主要转为耕地和水域;2010—2020年有5160.45 km2未利用地转出, 同时2509.96 km2其他土地类型转为未利用地, 面积一共减少2650.49 km2, 导致NPP总量增加1.92×108 kgC, 其中677.99 km2转为耕地, 耕地NPP增加1.20×108 kgC, 4221.96 km2转变为草地, 草地NPP增加9.47×107 kgC。

|
| 图 10 2001—2020年土地利用类型转移桑基图 Fig. 10 Sankey map of land use type transfer from 2001 to 2020 |
2001—2010年、2010—2020年耕地面积分别增加952.79 km2和2206.74 km2, 耕地NPP分别增加1.81×108 kgC和3.58×108 kgC, 主要由草地和未利用地转变而来。2001—2010年林地面积变化较小, 2010—2020年林地面积减少1284.21 km2, 导致NPP损失5.11×107 kgC, 其中82.96%的面积转化为草地。2010—2020年水域面积缩减742.04 km2, 导致NPP损失5.66×105 kgC。建设用地面积占比最小, 2001—2010年建设用地面积变化较小, 2010—2020年建设用地面积增加317.69 km2, 主要占用耕地和草地, 导致NPP变化9.19×105 kgC。
3.4 驱动因子定量探测不同因子对植被NPP的影响程度具有差异(图 11), 驱动因子q均值大小为:土地利用类型(0.453)>植被类型(0.406)>土壤类型(0.350)>年降水量(0.348)>高程(0.330)>年太阳辐射(0.251)>年均气温(0.237)>人口密度(0.235)>GDP(0.196)>坡度(0.106)>坡向(0.009)。因子交互作用下的q值明显高于单因子q值, 不同因子的影响不是独立的, 也不是简单叠加, 表现为双变量增强或非线性增强(图 12)。不同年份q值不同, 2001年、2010年和2020年植被类型和降水交互作用最强, 2005年植被类型和高程交互作用最强, 2015年土地利用类型和降水交互作用最强。气象因子之间交互作用后变化不明显, GDP、坡度和坡向的影响较小, 与其他因子交互后影响强度有所增加。

|
| 图 11 艾比湖流域植被NPP影响因子探测q值 Fig. 11 The q-statistics of driving factors influencing NPP′s spatial pattern in the Ebinur Lake Basin |

|
| 图 12 艾比湖流域植被NPP影响因子交互作用探测 Fig. 12 The interactive effect between influencing factors and NPP in the Ebinur Lake Basin |
风险探测用于找出各影响因子中植被NPP值最大的范围或类型, 确定最适合植被生长的条件。气象因子中年均气温、年降水量和年太阳辐射分别在-1.80—1.62 ℃、353—407 mm和4850—5210 MJ/m2范围内, 植被NPP值最大, 主要分布在研究区西部、北部及南部的山区;东南坡向的植被NPP值略高, 高程和坡度最适范围分别为2110—2610 m和17.30—64.70°;土壤类型中半淋溶土NPP值最高, 植被类型中混交林的NPP值最高;人均GDP在150—1.08×104元、人口密度为21—34人/km2的区域, 人类活动强度适合植被NPP积累。
4 讨论 4.1 CASA模型适用性本研究结果与其他学者的研究结果进一步对比验证(表 3), 在对应时段NPP值相对大小关系和变化趋势一致, 不同学者估算的NPP数值存在差异, 这与不同模型算法、输入数据不同以及研究时空范围不完全重叠有关。本研究基于CASA模型估算的NPP值高于MOD17A3 NPP和C-Fix模型模拟值[22], 博尔塔拉精河流域位于本研究区中部, 植被NPP时空分布与本研究结果一致[23], MOD17A3产品低估了灌丛、草地、荒漠的生产力[40], 艾比湖流域草地和荒漠面积分布较广, 本研究突出了CASA模型在艾比湖流域研究中应用的优势。天山北坡的范围包含了本研究区[41], 天山北坡森林覆盖面积广, 森林类型复杂, 林地NPP值明显高于本研究结果。
| 研究时段 Study period |
研究区域 Study area |
模型/产品 Model/Product |
平均值 mean values |
耕地 Cropland |
林地 Forest |
草地 Grassland |
未利用地 Unused land |
| 2001—2020 | 艾比湖流域(本文) | CASA模型 | 163.46—257.27 | 372.16 | 366.48 | 212.54 | 56.92 |
| 2001—2020 | 艾比湖流域(本文) | MOD17A3 | 118.13—175.17 | 191.82 | 260.93 | 169.67 | 48.78 |
| 2005、2012、2020 | 艾比湖流域[22] | C-Fix模型 | 290.81—338.56 | 286.86—332.17 | 105.99—115.09 | ||
| 2000—2015 | 博尔塔拉精河流域[23] | MOD17A3 | 182.00—248.60 | 254.10 | 321.10 | 131.70 | |
| 2000—2014 | 西北干旱区[40] | CASA模型 | 191.63 | 415.60 | 196.50 | 252.20 | 51.10 |
| 2004—2016 | 天山北坡[41] | CASA模型 | 110.68—172.06 | 385.43 | 554.72 | 165.56 | 31.58 |
| CASA:卡内基-艾姆斯-斯坦福方法Carnegie-Ames-Stanford Approach | |||||||
艾比湖流域西部及南部的山区降水和冰川融水提供水分, 水热条件适宜植被生长, 并且远离人类活动;河流沿岸的绿洲水源充足以及科学的人为管理, 促进栽培植物和草地NPP积累, 所以山区和河流沿岸的绿洲植被NPP值较高, 并且植被NPP波动变化小。耕地分布区NPP呈显著增加趋势, 可能与农业技术水平提高、灌溉技术改进, 农作物单位面积产量提高有关[42]。绿洲外围的荒漠地带受盐渍化、荒漠化的威胁以及人类活动的影响, 生态系统稳定性较差, 植被NPP值较低, 变异程度较大。
气候变化对植被NPP的影响存在区域差异, 本研究中植被NPP与降水相关性较强, 这与前人的研究结果相符[26], 西北干旱区植被主要受降水因素制约[43]。艾比湖流域降水量呈增加趋势[20], 降水增多一定程度上能促进植被生长, 但研究区植被生长并不完全取决于降水, 还依赖冰雪融水补给, 部分地区河水灌溉弥补了降水不足。研究区日照充足, 气温升高能促进海拔较高山区冰川融化, 同时蒸发量增大[44], 太阳辐射适量增加能促进栽培植被光合作用, 但在植被稀疏区域, 太阳辐射增强反而对植被生长产生一定的抑制作用。
研究时段内艾比湖流域植被NPP总量增加是土地利用类型变化和各土地类型自身NPP增加共同作用的结果。研究区未利用地面积持续缩减, 草地面积增加, 与近年来实施了一系列生态工程有关, 2007年国务院批准建立艾比湖湿地国家级自然保护区, 还先后实施了退耕还林(草)工程、山水林田湖草沙生态修复工程和草原生态保护补助奖励政策等[45]。经济发展和城镇化导致耕地和建设用地扩张, 一方面, 良好的政策能提高管理能力, 人工灌溉和施肥改善作物生长;另一方面, 人类活动超过一定阈值, 会破坏植被生长环境[46]。
4.3 不足与展望本研究采用的遥感数据空间分辨率较低, 未来研究应采用高分辨率数据结合实地考察资料来提高结果精度;近年来极端气候事件频发, 对植被NPP的影响不容忽视, 极端气候事件对植被NPP的影响在未来有待深入探究。
5 结论(1) 2001—2020年艾比湖流域植被NPP多年平均值为199.67 gC m-2 a-1, 整体呈波动上升趋势, 增长率为1.83 gC m-2 a-1;空间分异明显, 整体呈四周高中间低的分布格局。61.79%的区域植被NPP呈增加趋势, 植被处于生长或恢复状态, 河流沿岸的绿洲呈显著增加趋势。
(2) 植被NPP与气象因子呈正相关关系, 与降水的相关性最强;随高程、坡度增加, 植被NPP呈先增后减的变化趋势, 坡向对植被NPP的影响不明显。不同植被类型和土壤类型NPP值差异明显, 土地利用类型变化对植被NPP具有双重影响。
(3) 地理探测器结果表明, 土地利用类型和植被类型是影响植被NPP空间分异的主要因子, 因子交互作用表现为双变量增强或非线性增强, 植被类型、土地利用类型与气象因子、高程之间的交互作用较强, 基于风险探测确定出最适合植被生长的范围或类型。
研究结果可为决策者提供数据支撑和理论依据, 以期为统筹制定艾比湖流域可持续发展方案提供参考。艾比湖流域草地面积分布广, 对气候变化和人类活动响应敏感, 应继续落实退耕还草工程、科学放牧, 持续开展天然林保护修护等工作;未利用地面积逐年缩减, 在已有成效的基础上应继续开展沙地植被修复, 控制荒漠扩张;应严格落实永久基本农田保护任务, 依托科技力量发展绿洲生态农业, 城镇集约绿色发展。同时必须兼顾干旱区水资源合理分配, 合理有效地保护流域生态环境, 实现经济、社会和生态环境可持续发展。
| [1] |
Field C B, Behrenfeld M J, Randerson J T, Falkowski P. Primary production of the biosphere: integrating terrestrial and oceanic components. Science, 1998, 281(5374): 237-240. DOI:10.1126/science.281.5374.237 |
| [2] |
Teng M J, Zeng L X, Hu W J, Wang P C, Yan Z G, He W, Zhang Y, Huang Z L, Xiao W F. The impacts of climate changes and human activities on net primary productivity vary across an ecotone zone in Northwest China. Science of the Total Environment, 2020, 714: 136691. DOI:10.1016/j.scitotenv.2020.136691 |
| [3] |
Goetz S J, Prince S D, Goward S N, Thawley M M, Small J. Satellite remote sensing of primary production: an improved production efficiency modeling approach. Ecological Modelling, 1999, 122(3): 239-255. DOI:10.1016/S0304-3800(99)00140-4 |
| [4] |
Li C H, Sun H, Wu X D, Han H Y. An approach for improving soil water content for modeling net primary production on the Qinghai-Tibetan Plateau using Biome-BGC model. Catena, 2020, 184: 104253. DOI:10.1016/j.catena.2019.104253 |
| [5] |
Lieth H. Primary production: terrestrial ecosystems. Human Ecology, 1973, 1(4): 303-332. DOI:10.1007/BF01536729 |
| [6] |
朱文泉, 陈云浩, 徐丹, 李京. 陆地植被净初级生产力计算模型研究进展. 生态学杂志, 2005, 24(3): 296-300. DOI:10.3321/j.issn:1000-4890.2005.03.014 |
| [7] |
Potter C S, Randerson J T, Field C B, Matson P A, Vitousek P M, Mooney H A, Klooster S A. Terrestrial ecosystem production: A process model based on global satellite and surface data. Global Biogeochemical Cycles, 1993, 7(4): 811-841. DOI:10.1029/93GB02725 |
| [8] |
Guo X M, Tong S Q, Ren J Y, Ying H, Bao Y H. Dynamics of vegetation net primary productivity and its response to drought in the Mongolian Plateau. Atmosphere, 2021, 12(12): 1587. DOI:10.3390/atmos12121587 |
| [9] |
石智宇, 王雅婷, 赵清, 张连蓬, 朱长明. 2001-2020年中国植被净初级生产力时空变化及其驱动机制分析. 生态环境学报, 2022, 31(11): 2111-2123. |
| [10] |
单振东, 刘顿, 骆汉, 刘建伟, 张丽梅, 魏宇航. 气候变化背景下人类活动对承德接坝区植被净初级生产力的影响. 环境科学, 2023, 44(11): 6215-6225. |
| [11] |
Dang C Y, Shao Z F, Huang X, Zhuang Q W, Cheng G, Qian J X. Global vegetation productivity increased in response to COVID-19 restrictions. Geo-spatial Information Science, 2024, 1-14. |
| [12] |
Shi S H, Zhu L P, Luo Z H, Qiu H. Quantitative analysis of the contributions of climatic and anthropogenic factors to the variation in net primary productivity, China. Remote Sensing, 2023, 15(3): 789. DOI:10.3390/rs15030789 |
| [13] |
方浩玲, 程先富, 秦丽. 安徽省植被净初级生产力估算——基于改进的CASA模型. 生态学报, 2024, 44(4): 1601-1612. DOI:10.20103/j.stxb.202303270590 |
| [14] |
何颖, 马松梅, 张林, 张云玲, 贺凌云. 玛纳斯河流域NPP时空变化及其生物多样性维护功能重要性评价. 生态学报, 2023, 43(11): 4664-4673. DOI:10.5846/stxb202201190178 |
| [15] |
Higgins S I, Conradi T, Muhoko E. Shifts in vegetation activity of terrestrial ecosystems attributable to climate trends. Nature Geoscience, 2023, 16(2): 147-153. DOI:10.1038/s41561-022-01114-x |
| [16] |
Wang G J, Peng W F, Zhang L D, Zhang J. Quantifying the impacts of natural and human factors on changes in NPP using an optimal parameters-based geographical detector. Ecological Indicators, 2023, 155: 111018. DOI:10.1016/j.ecolind.2023.111018 |
| [17] |
Liu X P, Pei F S, Wen Y Y, Li X, Wang S J, Wu C J, Cai Y L, Wu J G, Chen J, Feng K S, Liu J G, Hubacek K, Davis S J, Yuan W P, Yu L, Liu Z. Global urban expansion offsets climate-driven increases in terrestrial net primary productivity. Nature Communications, 2019, 10(1): 5558. DOI:10.1038/s41467-019-13462-1 |
| [18] |
Yang H F, Zhong X N, Deng S Q, Xu H. Assessment of the impact of LUCC on NPP and its influencing factors in the Yangtze River Basin, China. Catena, 2021, 206: 105542. DOI:10.1016/j.catena.2021.105542 |
| [19] |
茆杨, 蒋勇军, 张彩云, 乔伊娜, 吕同汝, 邱菊. 近20年来西南地区植被净初级生产力时空变化与影响因素及其对生态工程响应. 生态学报, 2022, 42(7): 2878-2890. DOI:10.5846/stxb202101260270 |
| [20] |
Yao C L, Wang Y J, Yang G, Xia B F, Tong Y P, Yao J Q, Chen H H. Spatial and temporal variation in vegetation response to runoff in the Ebinur Lake Basin. Forests, 2023, 14(9): 1699. DOI:10.3390/f14091699 |
| [21] |
Tang H, Halike A, Yao K X, Wei Q Q, Yao L, Tuheti B, Luo J M, Duan Y F. Ecosystem service valuation and multi-scenario simulation in the Ebinur Lake Basin using a coupled GMOP-PLUS model. Scientific Reports, 2024, 14(1): 5071. DOI:10.1038/s41598-024-55763-6 |
| [22] |
朱旻, 王让会, 吕雅, 宗连玲, 江涤非. 艾比湖流域生态系统NPP对气候变化及景观动态的响应. 广东农业科学, 2014, 41(10): 176-180, 237. DOI:10.3969/j.issn.1004-874X.2014.10.040 |
| [23] |
赵鹏, 于瑞德, 陈桃, 郭艳飞, 吴燕良. 新疆博尔塔拉-精河流域植被净初级生产力变化及其与主要气候因子的关系. 植物资源与环境学报, 2019, 28(1): 79-84. DOI:10.3969/j.issn.1674-7895.2019.01.10 |
| [24] |
孙倩, 张敏, 曾永兵, 齐梦钰, 陈献震. 艾比湖流域降水和风速对植被NDVI时空变化的影响. 西南农业学报, 2018, 31(11): 2407-2412. |
| [25] |
赵坤成, 张永福, 朱小强, 顾刘阳, 郭家新. 2000-2014年艾比湖NDVI变化及其与降水、温度响应关系. 广东农业科学, 2017, 44(9): 82-88, 2. |
| [26] |
任立清. 艾比湖流域植被时空变化及驱动力分析. 干旱区地理, 2022, 45(2): 467-477. |
| [27] |
Liu Y X, Liu S L, Sun Y X, Li M Q, An Y, Shi F N. Spatial differentiation of the NPP and NDVI and its influencing factors vary with grassland type on the Qinghai-Tibet Plateau. Environmental Monitoring and Assessment, 2021, 193(1): 48. DOI:10.1007/s10661-020-08824-y |
| [28] |
Wei Q Q, Abudureheman M, Halike A, Yao K X, Yao L, Tang H, Tuheti B. Temporal and spatial variation analysis of habitat quality on the PLUS-InVEST model for Ebinur Lake Basin, China. Ecological Indicators, 2022, 145: 109632. DOI:10.1016/j.ecolind.2022.109632 |
| [29] |
朱文泉, 潘耀忠, 张锦水. 中国陆地植被净初级生产力遥感估算. 植物生态学报, 2007, 31(3): 413-424. DOI:10.3321/j.issn:1005-264X.2007.03.010 |
| [30] |
王川, 王丽莎, 张勇勇, 赵文智, 冯相艳. 2000-2020年祁连山植被净初级生产力时空变化及其驱动因素. 生态学报, 2023, 43(23): 9710-9720. DOI:10.20103/j.stxb.202208272444 |
| [31] |
Sen P K. Estimates of the regression coefficient based on Kendall's tau. Journal of the American Statistical Association, 1968, 63(324): 1379-1389. DOI:10.1080/01621459.1968.10480934 |
| [32] |
刘亮, 关靖云, 穆晨, 韩万强, 乔雪丽, 郑江华. 2008-2018年伊犁河流域植被净初级生产力时空分异特征. 生态学报, 2022, 42(12): 4861-4871. DOI:10.5846/stxb202104251084 |
| [33] |
王劲峰, 徐成东. 地理探测器: 原理与展望. 地理学报, 2017, 72(1): 116-134. |
| [34] |
Song Y Z, Wang J F, Ge Y, Xu C D. An optimal parameters-based geographical detector model enhances geographic characteristics of explanatory variables for spatial heterogeneity analysis: cases with different types of spatial data. GIScience & Remote Sensing, 2020, 57(5): 593-610. |
| [35] |
Liu L, Guan J Y, Zheng J H, Wang Y D, Han W Q, Liu Y J. Cumulative effects of drought have an impact on net primary productivity stability in Central Asian grasslands. Journal of environmental management, 2023, 344: 118734. DOI:10.1016/j.jenvman.2023.118734 |
| [36] |
王菲, 曹永强, 周姝含, 范帅邦, 姜雪梅. 黄河流域生态功能区植被碳汇估算及其气候影响要素. 生态学报, 2023, 43(6): 2501-2514. DOI:10.5846/stxb202203110591 |
| [37] |
Gulbeyaz O, Bond-Lamberty B, Akyurek Z, West T O. A new approach to evaluate the MODIS annual NPP product (MOD17A3) using forest field data from Turkey. International Journal of Remote Sensing, 2018, 39(8): 2560-2578. DOI:10.1080/01431161.2018.1430913 |
| [38] |
De Leeuw J, Rizayeva A, Namazov E, Bayramov E, Marshall M T, Etzold J, Neudert R. Application of the MODIS MOD 17 Net Primary Production product in grassland carrying capacity assessment. International Journal of Applied Earth Observation and Geoinformation, 2019, 78: 66-76. DOI:10.1016/j.jag.2018.09.014 |
| [39] |
Deng S Y, Beale C M, Platts P J, Thomas C D. Human modification of land cover alters net primary productivity, species richness and their relationship. Global Ecology and Biogeography, 2024, 33(3): 385-399. DOI:10.1111/geb.13795 |
| [40] |
焦伟, 陈亚宁, 李稚. 西北干旱区植被净初级生产力的遥感估算及时空差异原因. 生态学杂志, 2017, 36(1): 181-189. |
| [41] |
高军. 天山北坡植被净初级生产力时空分布特征及影响因素分析[D]. 石河子: 石河子大学, 2019.
|
| [42] |
李金珂, 杨玉婷, 张会茹, 黄铝文, 高义民. 秦巴山区近15年植被NPP时空演变特征及自然与人为因子解析. 生态学报, 2019, 39(22): 8504-8515. DOI:10.5846/stxb201807231575 |
| [43] |
贾俊鹤, 刘会玉, 林振山. 中国西北地区植被NPP多时间尺度变化及其对气候变化的响应. 生态学报, 2019, 39(14): 5058-5069. DOI:10.5846/stxb201808241810 |
| [44] |
谭娇, 丁建丽, 陈文倩, 董煜, 张钧泳, 唐梦迎. 近54年艾比湖绿洲太阳辐射变化特征. 气象科技, 2018, 46(3): 532-538. |
| [45] |
陈宸, 许玉凤. 天山北坡经济带稳定草地净初级生产力时空变化特征及其驱动力. 草业科学, 2023, 40(5): 1154-1167. |
| [46] |
Wu K, Zhou C F, Zhang Y X, Xu Y. Long-term spatiotemporal variation of net primary productivity and its correlation with the urbanization: a case study in Hubei Province, China. Frontiers in Environmental Science, 2022, 9: 656. |