 2024, Vol. 44
2024, Vol. 44文章信息
- 秦雨珂, 程珂珂, 杨波, 郑惠娜, 蔡中华, 肖宝华, 周进
- QIN Yuke, CHENG Keke, YANG Bo, ZHENG Huina, CAI Zhonghua, XIAO Baohua, ZHOU Jin
- 珊瑚共生体碳代谢特征研究进展
- Characterization of carbon metabolism in coral symbiont
- 生态学报. 2024, 44(9): 3561-3574
- Acta Ecologica Sinica. 2024, 44(9): 3561-3574
- http://dx.doi.org/10.20103/j.stxb.202307201558
-
文章历史
- 收稿日期: 2023-07-20
- 网络出版日期: 2024-02-01





2. 广东海洋大学深圳研究院, 深圳 518120
2. Shenzhen Institute of Guangdong Ocean University, Shenzhen 518120, China
珊瑚礁是地球上生产力和生物多样性最高的生态系统之一, 初级生产力达300—5000 g C m-2 a-1[1]。该生态系统维持高生产力的原因, 主要得益于“珊瑚-虫黄藻-微生物”高效的共生体系。复杂的共生体通过光合、呼吸、摄食和钙化等理化过程驱动着高效的碳循环。虫黄藻可以通过光合作用固定二氧化碳(CO2), 并合成有机物为珊瑚提供能量基质; 珊瑚不仅为虫黄藻提供栖息地和光合作用所需原料, 还可以从捕食消化的浮游生物中获得维生素、微量元素和其它必需物质供给藻类使用, 同时产生的代谢废物也可通过藻类转化为氨基酸等物质再次供珊瑚循环利用。对于微生物而言, 它们通常作为底盘生产力和活跃的物质代谢中介, 细菌\真菌-古菌等作为初级生产力的生产者和代谢者, 在虫黄藻和珊瑚宿主之间充当物质循环和元素转化的桥梁和引擎[2—3], 维持珊瑚礁生态系统的高生产力、高生物多样性和高代谢效率。
除了生态价值, 对于礁体本身, 它既是地质历史上石灰岩的主要物质来源, 也是现代海洋中最重要的储碳媒介。珊瑚礁体主要成分是碳酸钙, 珊瑚虫生物体成分主要是有机碳。珊瑚虫死亡后被埋藏的礁体可直接转换成石灰岩, 永久封存。定量分析结果表明, 珊瑚礁在全球尺度上每年可固定9亿t碳[4]。然而, 珊瑚共生体碳代谢具有复杂的过程和多样的形式, 这使得共生体中的碳流和碳通量较难核算。与红树林、海草床、盐沼等生态系统相比, 珊瑚礁还未被纳入“蓝碳”生态系统[5], 主要原因在于珊瑚礁碳释放和碳吸收的差值尚存争议, 碳源和碳汇的结论还有待厘清。产生这一矛盾有多种原因, 最主要的原因在于共生体有机碳代谢和无机碳矿化之间的对立。有机碳代谢包括光合作用和呼吸作用, 无机碳矿化包括碳酸钙的沉淀和碳酸钙的溶解; 这两对过程均有碳释放和碳吸收, 因而形成一对矛盾。同时, 不同礁区由于物理、化学、生物过程的复杂性, 也使得碳通量和碳收支的核算很难定量测定[6—7]。此外, 珊瑚健康状态、环境条件、人为干扰程度以及时空边界等其它因素均会对源汇差异产生影响[8]。一些作者认为珊瑚礁生态系统在净碳收支上表现为碳源[6]; 而近年来越来越多的证据显示, 从长时间的尺度看, 珊瑚礁生态系统最终可能表现为碳汇[9]。石拓等总结了共生体通过光合作用、摄食、钙化等生理生化过程驱动珊瑚生态系统高效的有机碳和无机碳循环, 指出珊瑚礁是减缓气候变化的潜在蓝色碳汇[10]。然而, 珊瑚礁碳源汇属性的判断还需要更多的数据, 梳理共生体的碳代谢过程显得尤为重要。
为此, 考虑到共生体碳代谢过程的重要性与复杂性, 本文尝试总结了珊瑚共生体碳循环的研究进展, 包括有机碳的迁移、无机碳的转化; 呼吸作用-光合作用-钙化作用下的源汇特征; 共生体中细菌-虫黄藻-病毒对碳循环的贡献; 以及珊瑚礁共生系统源汇悖论的由来和受控因素等; 最后对可行的增汇模式进行了探讨。论文目的旨在梳理共生体碳代谢特征, 解析珊瑚礁生态系统的源汇悖论, 为珊瑚礁生态系统进入海洋蓝碳俱乐部提供科学依据, 也为珊瑚的保育和增汇策略提供新视角和框架补充。
1 珊瑚共生体的碳组成 1.1 有机碳的构成与迁移共生体的有机碳主要来源于虫黄藻的光合作用和珊瑚宿主的异养摄食等, 包括稳定-不稳定碳库或者活性-惰性碳库; 依据原理和性质差异大致能通过物理C、化学C和生物C的方式进行区分。
物理C物理C主要是以珊瑚密度和粒径大小作为判别依据, 该理论认为有机碳的稳定性主要受团聚体的物理保护和矿物吸附过程的影响, 并强调有机碳存在状态及空间分布与其受保护程度具有较强的相关性。由于物理分离方法破坏性小, 更能表征珊瑚有机碳的功能和形态, 因此是目前有机碳分组的主流方法, 其包括以密度、粒径和团聚体性质而进行的细分。在密度分组中, 将珊瑚在重液(1.6—2.0 g/cm3)中离心, 浮在液体表面的组分为轻有机碳, 沉淀部分为重有机碳, 前者主要由分解一半的微生物残体和大块的动植物残体组成, 是介于新鲜有机质和腐殖质间的中间碳库, 这类碳分解程度较低, 稳定性较差, 易被微生物分解利用; 而后者主要是指与矿物结合的有机碳, 分解程度较高, 稳定性较强[11—12]。若以粒径大小为依据, 将>0.053 mm的颗粒作为颗粒有机碳, 将<0.053 mm的颗粒作为矿物结合有机碳[13—14]。两者的主要区别在于其来源成分和形成途径。其中, 颗粒有机碳主要通过藻源途径形成, 含有更多藻类来源的碳; 而矿物结合有机碳主要以菌源途径为主, 细菌碳占比较高[15]。在三种以团聚体为依据进行的分组中, 根据珊瑚团聚体大小, 以250 μm为界划分为大团聚体(>250 μm)和微团聚体(<250 μm)。其中大团聚体主要由有机胶结剂组成, 微团聚体主要由有机-矿质复合体组成[13]。
化学C化学分组是采取传统的化学方法将不同珊瑚碳组分按照它的化学性质进行区分。这些性质可在一定程度上反映有机碳在珊瑚中的化学稳定性。目前化学分组主要是利用珊瑚有机碳在各种提取剂(如水、缓冲液、稀酸或氧化水溶液等)中的溶解性、水解性和氧化性进行。化学C主要包括可溶性有机碳、酸水解有机碳和易氧化有机碳三类。在一类中泛指珊瑚中所有可溶解的有机碳。可溶性有机碳主要来源于珊瑚粘液、共附生生物体和分泌物等, 是珊瑚微生物分解的活性底物, 也是珊瑚有机碳最为活跃、迁移性最大的碳组分[16]。二类酸水解有机碳是指被酸(盐酸或硫酸)水解后的活性有机碳, 易被水解的主要是蛋白质、核酸和多糖, 而未能水解的为惰性有机碳, 如木质素、脂肪、蜡、树脂和软木脂等。依据该方法可将有机碳划分为活性和惰性有机碳, 进而可计算惰性指数, 衡量惰性有机碳库大小[17]。三类易氧化的有机碳是指: 通过333 mmol/L高锰酸钾的氧化作用将有机碳分为活性碳和非活性碳两个组分, 并计算碳库指数、活性指数和碳管理指数[18]。一般来说, 易被高锰酸钾氧化的成分主要有简单的糖类、氨基酸、胺基和氨基糖等物质。除了上述三种分类法, 随着化学表征技术手段(红外光谱、核磁共振和气相/液相色谱、质谱)的发展, 可以对珊瑚有机碳的成分及化学结构进行定性和定量分析。目前以13C核磁共振技术的应用最为常见, 由于其对样品破坏性小、处理简单、能够定量分子信息等优点被广泛用于珊瑚有机碳化学组成的研究[19]。其根据化学结构可将珊瑚有机碳定性分为四大类: 烷基碳、烷氧碳、芳香碳和羧基羰基碳[20]。
生物C生物分组往往是根据被微生物分解利用的程度来衡量珊瑚有机碳的稳定性, 将可被微生物分解利用的有机碳定义为活性有机碳(或微生物量碳), 主要通过测定珊瑚微生物生物量、可矿化碳或生物标志物来进行评估[21]。在一种微生物生物量的测定中, 珊瑚经氯仿熏蒸处理后, 被杀死的微生物残体等易分解有机质被释放到珊瑚中, 导致珊瑚中的可提取碳总量增加, 测定浸提液中总碳的含量可以计算微生物量碳。二种可矿化碳的测定中, 可矿化碳又称生物可降解碳, 主要通过微生物分解有机质后, 测定CO2释放量或专性呼吸速率qCO2(每单位微生物产生的CO2量)来量化可矿化碳量。同时, 利用珊瑚矿化速率和累积矿化量可以评估珊瑚微生物活性。三种生物C的衡量是针对生物标志物的测定, 常用的生物标志物种类包括脂类、糖类、木质素酚、氨基酸和氨基糖等, 其中木质素酚被定义为珊瑚中藻源碳的代表成分, 而氨基糖被认为是菌源碳的代表成分[22]。
上述物理C、化学C和生物C是对共生体有机碳的分组, 而对它们运动的描述常用到不同的迁移方式。目前有机碳的迁移方式主要有两种—垂直沉降和水平输送。在一种垂直沉降中, 海水中有机碳以颗粒态(Particulate organic carbon, POC)和溶解态(Dissolved organic carbon, DOC)的形式存在。在珊瑚礁区中的POC主要由微生物、藻类、浮游生物以及分泌物等构成, 这类物质的沉降及浮游生物的行为构成了POC的垂直迁移。DOC主要由珊瑚代谢产物和胞外聚合物(Extracellular Polymeric Substances, EPS)组成, 温跃层-盐跃层等不同水层的垂直混合带来了DOC的垂直运动。远海环境中由于浮游生物的活动以及有机物的氧化降解, 导致有机碳的浓度一般会随深度递减。有研究表明泻湖沉积物中有机碳的含量仅为周围生物体中碳含量的1%, 这表明泻湖中有机碳在下沉过程中被高效利用, 具有十分明显的垂直分布特征[23]。相比于远海和大洋环境, 浅水区由于受航运、生物活动以及陆源水流的扰动, 其POC和DOC没有明显的垂直分布差异[24]。除了垂直沉降, 水平输送也是有机碳迁移的重要方式。珊瑚礁生态系统物种丰富, 多样性和生产力远高于外周水体, 有相当比例的碳由共生体向外输出[25]。Zheng等通过沉积物捕捉器比较了沉积物的碳素含量, 发现有机碳的分布在共生体最高、泻湖次之、外周水体最低, 具有明显的圈层扩散特征[26]。Hata等对有机碳的水平输送能力进行了估算, 发现共生体向外周水平扩散的DOC和POC通量分别为(33±3) mmol C m-2 d-1和(6±1) mmol C m-2 d-1[25]。而从全球规模来看, 水平输出的总有机碳量约为15 ×1012 g Ct a-1, 占总净生产力的四分之三[27]。
1.2 无机碳的构成与转化共生体中无机碳的赋存状态主要为CO2、H2CO3、HCO3-、CO32-和CaCO3, 由矿化过程进行转化, 如图 1所示。气态CO2通过海水界面与水结合形成H2CO3分子, 碳酸通过两级电离释放出H+、HCO3-和CO32-, 游离的CO32-与Ca2+结合形成碳酸钙沉淀[28]。大多数珊瑚礁区正在以稳定的速率沉积碳酸钙。珊瑚及虫黄藻对碳酸钙的生成有很强的促进作用, 大部分矿化的无机碳沉降在珊瑚礁区, 只有少量被水流带到外周环境。健康的珊瑚礁生态系统中, 海水中二氧化碳分压(pCO2)基本保持稳定, 碳酸钙以相对恒定的速率沉积, 构成海洋中重要的碳酸盐沉降区。因此, 全球珊瑚礁面积虽小却贡献了碳酸钙产率的32%—43%[29]。

|
| 图 1 珊瑚礁共生体中无机碳的形成与转化过程 Fig. 1 Formation and transformation of inorganic carbon in coral reef symbiont |
珊瑚共生体中的碳来源主要有三个, 一是来源于海水中的DIC, 二是来源于虫黄藻的光合产物, 三是来源于被珊瑚摄取的浮游生物。在珊瑚共生体的碳循环过程中, 最重要的两名角色是虫黄藻和珊瑚宿主, 主要的碳循环过程包括有机碳循环和无机碳循环。有机碳循环主要是指光合作用和呼吸作用; 无机碳循环主要是指钙化作用(在珊瑚研究中也被叫做矿化作用)和反向钙化作用(即钙化作用的逆向)[30]。珊瑚礁生态系统中的虫黄藻通过光合作用利用无机碳(Dissolved inorganic carbon, DIC), 产生有机物并释放O2供珊瑚呼吸。其中大部分有机物被珊瑚排出体外, 但又很快被珊瑚作为食物来源捕获, 再次进入循环。与此同时, 珊瑚呼吸产生的CO2和虫黄藻的内生CO2与水中的Ca2+发生作用合成钙质珊瑚骨骼[31]。共生微生物常常充当代谢中介, 加快碳循环速度[3]。通常, 在健康的珊瑚礁系统中, 共生体中有机碳和无机碳的高效循环支撑了珊瑚礁的高生产力[32]; 而在珊瑚礁系统健康状态受损时, 其群落的碳循环过程受到显著干扰, 并改变碳的流向[10]。因此, 珊瑚钙化、虫黄藻光合作用和微生物代谢共同决定碳的代谢轨迹。
近年来, 微传感器技术、叶绿素荧光技术和离子质谱技术的迅速发展为共生系统的碳循环研究带来了微观层面的证据, 并取得了许多新的进展。有学者采用纳米二次离子质谱技术(NanoSIMS)分析了共生体中的碳循环, 以亚细胞分辨率跟踪和定量分析了组织中的碳素和稳定同位素的分布; 在群落和组织水平上重新审视了共生密度对珊瑚共生体的生理和营养影响; 同时精确地揭示了共生体中碳素的循环和能量传递[33—34], 这些结果证实了碳的代谢存在源与汇的交织, 而且提出了精确认识碳的代谢过程需要3D的空间意识。此外, 同位素技术也大放异彩, 例如Chen等利用氧同位素(δ18O)和碳同位素(δ13C), 示踪了我国台湾省南部核电站附近海域水温对珊瑚生长的影响, 结果表明温度越高, 珊瑚δ18O越低, 而δ13C越高[35]。Deng等利用δ13C示踪了南海北部的珊瑚礁体, 显示在近百年尺度, 光照强度改变了虫黄藻的光合作用[36]。Xu等通过δ13C示踪技术反演了南海珊瑚礁碳酸盐的长期变化, 证明在足够长的时间序列上珊瑚对碳的代谢过程中, 吸收大于释放[37]。
3 共生体成员的碳代谢特征 3.1 原核微生物(细菌与病毒)共生体中原核生物的组成具有高度多样性, 主导微生物包含α变形杆菌Alphaproteobacteria (如Rhodobacterales和Sphingomonadales), γ变形杆菌Gammaproteobacteria(如Alteromonadales、Pseudomonadales和Vibrionales), β-变形菌以及拟杆菌等[38]。有研究表明, 珊瑚共生体的微生物组成是影响珊瑚健康的关键因素[39]。微生物群落组装方式包括扩散限制、生态漂移和同质选择等, 在不同的生境中常常使用不同的方式, 以适应多变的环境, 因此微生物群落的组成是动态的[40]。相比于物种组成, 微生物在功能上差异更为多样, 在营养丰富的上升流区, 微生物群落中存在丰富的有机质代谢基因; 而在相对贫瘠的营养环境中, 微生物则具有更多参与光合作用的基因[38]。此外, 与开放海域相比, 共生体微生物的新陈代谢速率也明显更高。放射性胸腺嘧啶实验证明大堡礁区的珊瑚共生菌的倍增时间为3 h, 而开放海域的倍增时间为4 d[41]。共生体的这种高代谢率归因于宿主释放的有机碳被细菌吸收[42], 同时这种高效率对生态系统的碳流通也存在较大影响[43]。高代谢率通常与低生长效率相关, 这是一种速率-产量的平衡。细菌在不同的共生关系中代谢碳素(如DOC)时, 也常常使用不同的代谢途径。如在与藻类共生的关系中, 微生物会优先利用低效率的中心碳代谢途径, 如Entner-Doudoroff途径和戊糖磷酸途径; 而在珊瑚中则更偏向于利用Embden-Meyerhof-Parnas途径[44]。微生物这些代谢路径的差异主要取决于珊瑚和藻类释放的有机质不同, 其中珊瑚渗出液主要以脂类和蛋白质为主, 而藻类渗出液以不稳定的碳水化合物为主[45]。此外, EMP途径代谢速率要远远高于中心代谢途径, 因而提升了对碳的周转速率。这一过程涉及珊瑚宿主、异养微生物和多种代谢中间物的参与, 并受有氧和无氧环境的影响(图 2)。

对于一些深海的冷水珊瑚, 如石珊瑚和深水软珊瑚, 细菌碳源是重要的类别[47—48]。多数共生细菌都具有参与深水珊瑚碳循环的能力, DNA稳定同位素探测了海底火山口的一些深水珊瑚, 发现这些珊瑚依靠与其共生的化能细菌及其产物为碳源[49—50]。Middelburg等也表明深水石珊瑚的食物来源可能与共生的化能自养微生物有关[51]。一般情况下, 化能自养微生物寄生在珊瑚的粘膜层或者体外, 这些微生物不会借助光合作用的方式获取碳源, 而是通过氧化一些简单的无机化合物, 例如同化二氧化碳来获取化学能[51]。除此之外, 稳定同位素的数据表明, 许多珊瑚群落获取食物依靠的是化学合成反应, 但不同种类的珊瑚摄食策略存在差异。有研究表明Paramuricea sp.与C. delta两种珊瑚的摄食与硫氧化家族细菌SUP05有关, 该家族细菌通过重组(形成不同的C加工策略)帮助珊瑚利用冷源渗漏碳, 以适应周围环境[52]。该研究表明这些深海珊瑚可能会改变某些特定的微生物丰度, 以获取碳素。
除细菌外, 共生体中病毒也具有重要的作用(图 3)。在病毒裂解作用下细胞内容物作为溶解有机质释放, 病毒发挥着分流作用, 增加了有机质的循环, 减少了微生物生物量碳向食物网中的较高营养级的转移, 同时增加了上层食物网的初级生产力[53—54]。这种病毒裂解对细菌群落的控制被称为杀死赢家(Kill the Winner, KtW) 策略[55]。在波利尼西亚Moorea岛相对原生态的珊瑚礁中, 通过病毒裂解每天会清除24%—367%的细菌量, 每天每升水释放1.0—62 μg有机碳[56]。同时, 在这一区域还发现了一种有别于KtW策略的搭便车“Piggyback-the-Winner”理论[57], 即溶源性病毒不付成本而坐享收益。有学者总结裂解是富营养条件下的主要策略, 而溶源是在寡营养条件下的应对方式[58—59]。波利尼西亚的营养含量很低, 因而更偏向于搭便车理论。对2110个完整细菌基因组中噬菌体的分析表明, 细菌生长速度与溶源性之间存在正相关关系, 进一步印证了搭便车理论[60]。这种行为减缓了珊瑚礁有机碳流的代谢速率, 但是却提高了利用效率。

虫黄藻(Zooxanthellae)是棕色单细胞的甲藻, 有着不同的形态特征[62], 且大多都是自养生物, 其光合作用是珊瑚共生体中碳代谢不可缺少的一环。虫黄藻给珊瑚宿主提供其自身绝大部分的光合作用产物, 如糖类、氨基酸、氧气等, 这些产物用于满足珊瑚的生长需要。而珊瑚给虫黄藻提供N、P、CO2等代谢产物, 这些产物可以满足虫黄藻的生长和光合作用。对C素的代谢中, 主要受光合磷酸化和卡尔文循环中的影响(图 4)。健康状态下CO2来源顺利, 卡尔文循环正常, 光合磷酸化行使, 碳素得以有效固定; 而当CO2来源受阻时, 则卡尔文循环中断, 光合磷酸化受阻, 有机碳不能有效合成。这一对过程调控着碳的走向。

一方面, 虫黄藻吸收珊瑚释放的CO2和环境中的CO2, 在光照条件下生成葡萄糖和O2; 有机碳一部分成为珊瑚和虫黄藻呼吸作用的原料, 一部分直接排放到环境中再被珊瑚的触手捕捉[31]。另一方面, 虫黄藻利用自身生成的部分有机碳和环境中的O2进行呼吸作用产生CO2和水, 作为光合作用的原料。虫黄藻呼吸作用产生的CO2也能被珊瑚用来与环境中的Ca2+作用合成宿主的钙质骨骼[31]。具体的过程为CO2与水反应生成HCO3-和H+, HCO3-再与Ca2+反应生成CaCO3和CO2(Ca2++2HCO3-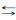

|
| 图 5 珊瑚礁碳酸盐体系及矿化过程 Fig. 5 Carbonate system and mineralization process of coral reef |

Kelton等人应用贝叶斯混合模型和稳定碳同位素比值分析, 量化了红海食物网中从初级生产者到珊瑚礁鱼类的碳流; 证实了虫黄藻在C源层级上的重要份额[65]。Christine等人应用氨基酸稳定同位素分析进一步强化了虫黄藻在共生体中营养交换、碳周转和抵御环境压力下的重要作用[66]。除此之外, 还有研究表明, 虫黄藻密度影响共生体的碳吸收和转运模式, 面对无害的POC环境, 有机碳的积累与珊瑚体内的虫黄藻密度呈正相关[67]; 而面对有害POC颗粒(如微塑料), 有机碳的积累则与珊瑚体内的虫黄藻密度呈负相关[68]。因此, 虫黄藻的健康、数量, 它参与的光合作用、呼吸作用与钙化作用共同影响着共生体系统中的碳源-碳汇属性。
在环境参数对虫黄藻碳代谢的影响上, 温度和光照两个参数被较多关注。Jones等人利用脉冲调幅(PAM) 叶绿素荧光和光呼吸测定法, 探讨了热应激对造礁珊瑚中虫黄藻光合作用的影响[69]。笔者近期针对深圳大鹏湾的数据表明, 光照强度或浊度影响虫黄藻的光合效率, 浊度越高会影响光的射入, 降低藻类的光合效率(2024, 未发表)。近年来得益于建模技术的发展, 评估环境参数对虫黄藻C代谢的影响更多依赖模型[70]。目前主流的数值模型包括三类, 一种是珊瑚—虫黄藻内循环模型, 该模型在细胞层面上参数化水质、营养盐、光照、温度、pH等, 评估虫黄藻对无机碳的获取和有机碳的制造过程。二种是珊瑚微碳循环模型, 该模型将虫黄藻光合作用所积累的能量和珊瑚钙化消耗的能量关联起来, 耦合环境参数共同判别虫黄藻的碳周转及CaCO3的沉降作用。三种为综合耦合模型, 结合水体生源要素, 海气界面处的气体交换, 以及珊瑚生理生化过程带来的DIC、DIN和DOM浓度的变化, 建立耦合反馈模型。这类模型的应用, 可以将共生藻光合呼吸、珊瑚虫有机体生化过程、以及主要的环境因子联系起来, 探讨全球变化背景下共生关系对单一或联合环境参数变化的响应机制。
4 珊瑚礁生态系统“源-汇(source-sink)”解析 4.1 “源-汇”悖论分析珊瑚礁的“源-汇”属性目前主要定义为三种, 即净源、净汇或源-汇间转变[71]。这主要源于不同珊瑚共生体中复杂的“理-化-生”过程, 使得碳通量和碳收支的核算不易确定。共生体的碳通量受多种因素的影响, 主要归咎于共生体有机碳代谢和无机碳矿化。共生体群落的光合作用能够降低水体的pCO2, 而呼吸作用能够增加水体pCO2。此外, 碳酸钙的合成与溶解导致水体CO2的释放与消耗。因此, 有机碳代谢和CaCO3沉淀与溶解的平衡控制着珊瑚礁生态系统的源与汇[72]。
尽管早有报道礁区可能是潜在的碳汇, 但是由于钙化过程会产生CO2(Ca2++ 2HCO3-
面对源-汇的现状, 需要梳理几个问题。首先, 从造礁珊瑚自身来看, 大气CO2浓度升高可能会有利于虫黄藻的光合作用和初级生产力; 但伴随的酸化又会抑制珊瑚的钙化作用, 导致其碳汇属性被削弱。这一点在印度洋—太平洋多个珊瑚礁生态系统中均有过报道[8]。其次, 生态系统之间的相互作用对于碳交换有着重要影响。在红树林、海草床和珊瑚礁的连续生态系统中, 虫黄藻可固定大量来自海草床和红树林的溶解无机碳, 并将其转化为有机碳。此外, 珊瑚释放的CO2可以被大型海草、海藻等初级生产者再次利用, 因此连续生态系统整体上表现出较强的碳汇性质, 这一点在“源-汇”计算中经常被忽略[76]。最后, 珊瑚礁的不同状态也影响着其“源-汇”状态, 而珊瑚礁的不同状态由珊瑚群落的生理活动状态决定。一般情况下, 光照、温度和环境中的二氧化碳浓度会影响珊瑚群落的健康状态和生理活动[77—78]。
目前, 一些技术逐渐在“碳-汇”悖论的分析中大放异彩, 其中以模型法为代表, 模型法是评估与核算碳源碳汇问题的好方法。在陆生系统, Jia等人应用模型法分析了青藏高原水体在过去20年里的碳源碳汇属性, 通过输入历史CO2交换通量、速率、pCO2等, 证实高原湖泊总体上是碳汇, 且具有西南部较高、东北部较低的地理分布性[79]。该研究的思路和方法为珊瑚礁源汇属性的判断提供了参照。的确, 这方面已经有了一些积极的尝试, 其中最具代表性的为共生体耦合模型[80]。该模型在细胞尺度上描述珊瑚光合、呼吸和有机碳转运, 在生物体尺度上描述了珊瑚不同组分(即腔肠体、钙化流体)中的离子转运及钙化作用, 并考虑了与周围海水的物质交换, 最后综合概述了影响珊瑚共生关系碳收支的主要过程。该模型指出对于共生体来说, 光合作用产生的葡萄糖是主要的碳源, 有机碳向宿主的转运是其主要的碳汇。对于珊瑚来说从共生体转运的大部分有机碳会以粘液渗漏的形式丢失。此外, 该耦合模型还应用于不同大气CO2分压下的情景, 结果显示在较高的大气CO2分压下, 珊瑚钙化率下降, 光合速率增加, 珊瑚体内的碳酸盐体系也会发生相应变化[80]。总的来说, 该模型以一种耦合的方式模拟珊瑚共生系统, 不仅突出了共生体碳代谢特征这一至关重要的环节, 而且系统涉及了珊瑚碳源汇过程, 为全面剖析珊瑚共生体系统碳代谢归属提供了重要参照。
4.2 “源-汇”效应的受控因子光照、温度和pCO2等是影响珊瑚群落生理活动(呼吸作用、光合作用和钙化作用)的主要因素[77—78], 可以左右珊瑚礁生态系统碳的“源-汇”属性。以光照为例, 适宜的光强有利于共生藻的光合作用, 但是强光会带来光损伤抑制光合作用, 导致与宿主脱离进而发生白化或死亡[81]。除了光照外, 温度也是珊瑚的敏感因子, 当环境温度<28℃时, 珊瑚的钙化作用与光合作用均随温度的上升而增强; 当温度>28℃时, 珊瑚的钙化速率与光合效率均呈现显著下降的趋势[82], 因而高温会损害碳汇效应。此外, CO2分压也影响其源汇走向, 海水CO2含量的增加(海洋酸化)使得海水碳酸盐(CO32-)浓度降低, 珊瑚钙化效率随之下降[83]。Gattuso等首次阐明了珊瑚群落初级生产力、呼吸率以及钙化率对海水CO2分压升高的响应, 表明pCO2升高对珊瑚共生藻的光合作用产生明显影响, 并导致CaCO3饱和度显著降低[84]。Langdon等进一步对珊瑚群落净钙化速率进行了研究, 证实随着pCO2的持续升高, 海水中CO32-浓度下降30%, 珊瑚钙化效率降低14%—40%[85]。张远辉等根据碳酸盐的热力学模式预测, 从工业革命至2100年南沙海域的CaCO3饱和度将下降43%左右, 从而可能引起珊瑚礁钙化速率回落33%[86]。
除了环境参数, 珊瑚的营养利用方式、健康状态、与周边生态系统的连通性都会影响源汇结局。造礁珊瑚是混合营养生物, 其自养和异养这两种生活方式间的转换对珊瑚礁生态系统的碳“源-汇”属性会产生影响, 甚至可能会决定其属性[87]。当珊瑚礁生态系统健康时, 它整体上表现为大气CO2的净汇; 但当珊瑚礁生态系统健康受到胁迫时, 它则表现为大气CO2的净源[10]。严宏强等指出珊瑚健康受损可能导致级联效应, 如生物多样性下降、食物网结构简化、营养级缩短以及空间结构多样性衰退等严重后果; 这可能会引发从碳汇到碳源的转变, 导致原本固定在各营养层级生物体内的有机碳被释放, 从而削弱珊瑚礁生态系统的储碳总量[28]。
5 总结与展望共生体碳代谢的研究在有机碳的水平-垂直输送、无机碳的转化和C代谢过程动力学等方面取得了长足进展。然而目前对珊瑚礁生态系统碳“源-汇”的研究仍较为有限, 我们未能全面地认识共生体细胞、个体、群落多尺度上的碳循环过程及调控机制, 为了进一步明确珊瑚礁生态系统蓝碳储库的身份, 未来值得关注的研究方向包括:
(1) 珊瑚共生体增汇模式 在提升珊瑚礁生态系统碳汇能力的途径中, 如果重点关注珊瑚共生体的成员, 可以从两条路径进行尝试, 一是通过调节外界因素, 如人工上升流方法, 二是关注珊瑚群落的健康状态, 如珊瑚资源修复策略。目前广西涠洲岛海域、广东徐闻、福建东山、三亚鹿回头以及深圳大鹏湾海域等主要示范区和保护地正在实施有性繁殖相关的方法, 期望在不久的将来能看到更多新品种, 抗逆品系以及新修复资源。
(2) 新型技术的应用 近年来一些新技术在珊瑚碳代谢的研究中崭露头角。如纳米液相色谱-串联质谱, Motone等人首次采用该技术对珊瑚幼虫进行了蛋白质组学分析, 结果表明可以精准了解水螅体功能蛋白的变化情况, 帮助计算C通量[88]。此外, 纳米二次离子质谱技术(NanoSIMS)也是重要的方法, Krueger等采用该方法描绘了珊瑚共生体之间能量传递的规律[33—34], 对C流动力学的明晰提供了便利。上述纳米尺度的新方法对加深珊瑚共生体C代谢的理解具有重要参考价值。
(3) 拓展时间尺度(十年或百年跨度)和空间尺度(生态连通性), 全方位、多层次评估珊瑚共生体系统碳通量的变化以及固碳、储碳特征珊瑚碳代谢过程的复杂性和源汇身份的不确定性, 不易通过短期的试验和有限的数据获得, 亦不可作为孤立单一的生态系统来对待, 而应在时空尺度上纵深考虑。一是评估长时序范围的净汇潜力; 二是评估与其它生态系统(红树林、海草床、盐沼、浮游植物)的耦合性, 探索多生态系统互作下的综合C汇效果, 为珊瑚礁进入蓝碳俱乐部获取科学筹码。
(4) 资源修复新策略 积极探索基于营养流利用、上升流方式改善珊瑚生境的方法, 营造更为健康、更具环境缓冲能力的生态系统[89]。拓展从无性繁殖到有性繁殖的增殖保育模式, 通过基因编辑技术筛选抗逆基因、选育抗逆品系, 获取工程珊瑚株, 为珊瑚礁资源修复和增汇潜力提供科技支撑[90]。
(5) 建立珊瑚碳汇示范区 珊瑚健康状态是影响珊瑚碳汇结局的关键性因素, 珊瑚的健康受到胁迫时, 会吸收来自生态系统以外的有机碳, 共生体被动经历从自养到异养的转变, 同时从生态效应上也从健康珊瑚礁主导的碳“汇”系统转向由退化珊瑚礁主导的碳“源”系统。由此看来, 保护珊瑚礁的健康可以大大增加珊瑚礁的碳负排放能力, 防止珊瑚礁退化。该行为不仅能保护珊瑚礁, 而且可以规范治理珊瑚礁的方式, 探寻更科学更高效地增加珊瑚负碳排放措施[91]。
(6) 探索珊瑚碳汇交易类型 为了加大海洋生态系统对碳汇的贡献, 可以引入市场机制, 重视碳汇交易市场[92—93]。我国有着丰富的海洋资源, 三大海岸带生态系统以及海洋牧场的案例为海洋碳汇交易市场提供了有力基础[94]。但是目前以珊瑚礁生态系统为代表的碳交易类型、和市场机制还相当薄弱。为此我们应该鼓励吸引社科和经济领域的专家积极参与, 支持企业和相关机构进行有益尝试。以政策环为推力、利益环为引力、责任环为压力建立体系, 为珊瑚礁生态系统碳汇交易市场的发展做出积极尝试[95]。
| [1] |
Hatcher B G. Coral reef primary productivity: a beggar's banquet. Trends in Ecology & Evolution, 1988, 3(5): 106-111. |
| [2] |
王涛, 程珂珂, 蔡中华, 周进. 珊瑚共生体中"细菌-虫黄藻-宿主"三角关系的通讯交流. 应用生态学报, 2022, 33(9): 2572-2584. |
| [3] |
Zhou J, Jin H, Cai Z H. A review of the role and function of microbes in coral reef ecosystem. Chinese Journal of Applied Ecology, 2014, 25(3): 919-930. |
| [4] |
Kinsey D W, Hopley D. The significance of coral reefs as global carbon sinks-response to Greenhouse. Global and Planetary Change, 1991, 3(4): 363-377. DOI:10.1016/0921-8181(91)90117-F |
| [5] |
Macreadie P I, Anton A, Raven J A, Beaumont N, Connolly R M, Friess D A, Kelleway J J, Kennedy H, Kuwae T, Lavery P S, Lovelock C E, Smale D A, Apostolaki E T, Atwood T B, Baldock J, Bianchi T S, Chmura G L, Eyre B D, Fourqurean J W, Hall-Spencer J M, Huxham M, Hendriks I E, Krause-Jensen D, Laffoley D, Luisetti T, Marbà N, Masque P, McGlathery K J, Megonigal J P, Murdiyarso D, Russell B D, Santos R, Serrano O, Silliman B R, Watanabe K, Duarte C M. The future of Blue Carbon science. Nature Communications, 2019, 10: 3998. DOI:10.1038/s41467-019-11693-w |
| [6] |
Ware J R, Smith S V, Reaka-Kudla M L. Coral reefs: sources or sinks of atmospheric CO2?. Coral Reefs, 1992, 11(3): 127-130. DOI:10.1007/BF00255465 |
| [7] |
Chisholm J R, Barnes D J. Anomalies in coral reef community metabolism and their potential importance in the reef CO2 source-sink debate. Proceedings of the National Academy of Sciences of the United States of America, 1998, 95(11): 6566-6569. |
| [8] |
Mayer B, Rixen T, Pohlmann T. The spatial and temporal variability of air-sea CO2 fluxes and the effect of net coral reef calcification in the Indonesian Seas: a numerical sensitivity study. Frontiers in Marine Science, 2018, 5: 116. DOI:10.3389/fmars.2018.00116 |
| [9] |
Zhang S Q, Bai Y, He X Q, Yu S J, Song Z G, Gong F, Zhu Q K, Pan D L. The carbon sink of the Coral Sea, the world's second largest marginal sea, weakened during 2006-2018. Science of the Total Environment, 2023, 872: 162219. DOI:10.1016/j.scitotenv.2023.162219 |
| [10] |
石拓, 郑新庆, 张涵, 王啟芳, 钟昕. 珊瑚礁: 减缓气候变化的潜在蓝色碳汇. 中国科学院院刊, 2021, 36(3): 270-278. |
| [11] |
杨丽霞, 潘剑君. 土壤活性有机碳库测定方法研究进展. 土壤通报, 2004, 35(4): 502-506. |
| [12] |
梁贻仓. 不同农田管理措施下土壤有机碳及其组分研究进展. 安徽农业科学, 2013, 41(24): 9964-9966. |
| [13] |
张国, 曹志平, 胡婵娟. 土壤有机碳分组方法及其在农田生态系统研究中的应用. 应用生态学报, 2011, 22(7): 1921-1930. |
| [14] |
佟小刚, 徐明岗, 张文菊, 卢昌艾. 长期施肥对红壤和潮土颗粒有机碳含量与分布的影响. 中国农业科学, 2008, 41(11): 3664-3671. |
| [15] |
梁爱珍, 张晓平, 杨学明, 申艳, 时秀焕, 范如芹, 方华军. 黑土颗粒态有机碳与矿物结合态有机碳的变化研究. 土壤学报, 2010, 47(1): 153-158. |
| [16] |
Grottoli A G, Dalcin Martins P, Wilkins M J, Johnston M D, Warner M E, Cai W J, Melman T F, Hoadley K D, Pettay D T, Levas S, Schoepf V. Coral physiology and microbiome dynamics under combined warming and ocean acidification. PLoS One, 2018, 13(1): e0191156. DOI:10.1371/journal.pone.0191156 |
| [17] |
张焕军, 郁红艳, 丁维新. 土壤碳水化合物的转化与累积研究进展. 土壤学报, 2013, 50(6): 1200-1206. |
| [18] |
郑红. 土壤活性有机碳的研究进展. 中国林副特产, 2011(6): 90-94. |
| [19] |
Lu S Q, Li X W, Li S W, Cui Z, Guo Y W, Han G Y. Sinuhirtins A and B, two uncommon norhumulene-type terpenoids from the South China Sea soft coral Sinularia hirta. Tetrahedron Letters, 2019, 60(51): 151308. DOI:10.1016/j.tetlet.2019.151308 |
| [20] |
李娜, 盛明, 尤孟阳, 韩晓增. 应用13C核磁共振技术研究土壤有机质化学结构进展. 土壤学报, 2019, 56(4): 796-812. |
| [21] |
张丽敏, 徐明岗, 娄翼来, 王小利, 李忠芳. 土壤有机碳分组方法概述. 中国土壤与肥料, 2014(4): 1-6. |
| [22] |
冯晓娟, 王依云, 刘婷, 贾娟, 戴国华, 马田, 刘宗广. 生物标志物及其在生态系统研究中的应用. 植物生态学报, 2020, 44(4): 384-394. |
| [23] |
Song J, Diao W, Li F, Cui Z, Guo Y, Han G. Carbon cycling in Nansha coral reef ecosystem, South China Sea. Oceanologia et Limnologia Sinica, 2003(06): 586-592. |
| [24] |
Reiswig H M. Partial carbon and energy budgets of the bacteriosponge verohgia fistularis (Porifera: Demospongiae) in Barbados. Marine Ecology, 1981, 2(4): 273-293. DOI:10.1111/j.1439-0485.1981.tb00271.x |
| [25] |
Hata H, Kudo S, Yamano H, Kurano N, Kayanne H. Organic carbon flux in Shiraho coral reef (Ishigaki Island, Japan). Marine Ecology Progress Series, 2002, 232: 129-140. DOI:10.3354/meps232129 |
| [26] |
Zheng P, Guo W, Hu M, Yang Y, Wu L, Wang H. Distribution and diurnal changes of organic carbon in Zhubi reef ecosystem Nansha Islands. Marine Science Bulletin, 2004, 23(2): 13-18. |
| [27] |
Crossland C J, Hatcher B G, Smith S V. Role of coral reefs in global ocean production. Coral Reefs, 1991, 10(2): 55-64. DOI:10.1007/BF00571824 |
| [28] |
严宏强, 余克服, 谭烨辉. 珊瑚礁区碳循环研究进展. 生态学报, 2009, 29(11): 6207-6215. |
| [29] |
Suzuki A, Kawahata H. Reef water CO2 system and carbon production of coral reefs: topographic control of system-level performance. Global environmental change in the ocean and on land, 2004, 229-248. |
| [30] |
黄晖, 袁翔城, 宋严, 李颖心, 周伟华, 龙爱民. 瑚礁生态系统固碳过程及储碳机制研究进展. 热带海洋学报, 2023, 1-10. |
| [31] |
Rädecker N, Pogoreutz C, Ziegler M, Ashok A, Barreto M M, Chaidez V, Grupstra C G B, Ng Y M, Perna G, Aranda M, Voolstra C R. Assessing the effects of iron enrichment across holobiont compartments reveals reduced microbial nitrogen fixation in the Red Sea coral Pocillopora verrucosa. Ecology and Evolution, 2017, 7(16): 6614-6621. DOI:10.1002/ece3.3293 |
| [32] |
Wang J, Douglas A E. Nitrogen recycling or nitrogen conservation in an alga-invertebrate symbiosis?. Journal of Experimental Biology, 1998, 201(16): 2445-2453. DOI:10.1242/jeb.201.16.2445 |
| [33] |
Krueger T, Horwitz N, Bodin J, Giovani M E, Escrig S, Fine M, Meibom A. Intracellular competition for nitrogen controls dinoflagellate population density in corals. Proceedings of the Royal Society B: Biological Sciences, 2020, 287(1922): 20200049. DOI:10.1098/rspb.2020.0049 |
| [34] |
Loussert-Fonta C, Toullec G, Paraecattil A A, Jeangros Q, Krueger T, Escrig S, Meibom A. Correlation of fluorescence microscopy, electron microscopy, and NanoSIMS stable isotope imaging on a single tissue section. Communications Biology, 2020, 3: 362. DOI:10.1038/s42003-020-1095-x |
| [35] |
Chen C T, Wang C H, Soong K Y, Wang B J. Water temperature records from corals near the nuclear power plant in southern Taiwan. Science in China Series D: Earth Sciences, 2001, 44(4): 356-362. DOI:10.1007/BF02907106 |
| [36] |
Deng W F, Chen X F, Wei G J, Zeng T, Zhao J X. Decoupling of coral skeletal δ13C and solar irradiance over the past millennium caused by the oceanic Suess effect. Paleoceanography, 2017, 32(2): 161-171. DOI:10.1002/2016PA003049 |
| [37] |
Xu S D, Yu K F, Fan T L, Jiang W, Wang R, Zhang Y, Yue Y F, Wang S P. Coral reef carbonate δ13C records from the northern South China Sea: a useful proxy for seawater δ13Cand the carbon cycle over the past 1.8 Ma. Global and Planetary Change, 2019, 182: 103003. DOI:10.1016/j.gloplacha.2019.103003 |
| [38] |
Kelly L W, Williams G J, Barott K L, Carlson C A, Dinsdale E A, Edwards R A, Haas A F, Haynes M, Lim Y W, McDole T, Nelson C E, Sala E, Sandin S A, Smith J E, Vermeij M J A, Youle M, Rohwer F. Local genomic adaptation of coral reef-associated microbiomes to gradients of natural variability and anthropogenic stressors. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(28): 10227-10232. |
| [39] |
MacKnight N J, Cobleigh K, Lasseigne D, Chaves-Fonnegra A, Gutting A, Dimos B, Antoine J, Fuess L, Ricci C, Butler C, Muller E M, Mydlarz L D, Brandt M. Microbial dysbiosis reflects disease resistance in diverse coral species. Communications Biology, 2021, 4: 679. DOI:10.1038/s42003-021-02163-5 |
| [40] |
Liu J M, Wang X L, Liu J A, Liu X Y, Zhang X H, Liu J W. Comparison of assembly process and co-occurrence pattern between planktonic and benthic microbial communities in the Bohai Sea. Frontiers in Microbiology, 2022, 13: 1003623. DOI:10.3389/fmicb.2022.1003623 |
| [41] |
Moriarty D J W, Pollard P C, Hunt W G. Temporal and spatial variation in bacterial production in the water column over a coral reef. Marine Biology, 1985, 85(3): 285-292. DOI:10.1007/BF00393249 |
| [42] |
Ferrier-Pagès C, Leclercq N, Jaubert J, Pelegrí S P. Enhancement of pico- and nanoplankton growth by coral exudates. Aquatic Microbial Ecology, 2000, 21: 203-209. DOI:10.3354/ame021203 |
| [43] |
Lipson D A. The complex relationship between microbial growth rate and yield and its implications for ecosystem processes. Frontiers in Microbiology, 2015, 6: 615. |
| [44] |
Haas A F, Fairoz M F M, Kelly L W, Nelson C E, Dinsdale E A, Edwards R A, Giles S, Hatay M, Hisakawa N, Knowles B, Lim Y W, Maughan H, Pantos O, Roach T N F, Sanchez S E, Silveira C B, Sandin S, Smith J E, Rohwer F. Global microbialization of coral reefs. Nature Microbiology, 2016, 1: 16042. DOI:10.1038/nmicrobiol.2016.42 |
| [45] |
Nelson C E, Goldberg S J, Wegley Kelly L, Haas A F, Smith J E, Rohwer F, Carlson C A. Coral and macroalgal exudates vary in neutral sugar composition and differentially enrich reef bacterioplankton lineages. The ISME Journal, 2013, 7(5): 962-979. DOI:10.1038/ismej.2012.161 |
| [46] |
Bourne D G, Dennis P G, Uthicke S, Soo R M, Tyson G W, Webster N. Coral reef invertebrate microbiomes correlate with the presence of photosymbionts. The ISME Journal, 2013, 7(7): 1452-1458. DOI:10.1038/ismej.2012.172 |
| [47] |
Mueller C E, Larsson A I, Veuger B, Middelburg J J, van Oevelen D. Opportunistic feeding on various organic food sources by the cold-water coral Lophelia pertusa. Biogeosciences Discussions, 2013, 10(7): 11375-11403. |
| [48] |
Elias-Piera F, Rossi S, Gili J M, Orejas C. Trophic ecology of seven Antarctic Gorgonian species. Marine Ecology Progress Series, 2013, 477: 93-106. DOI:10.3354/meps10152 |
| [49] |
Jensen S, Neufeld J D, Birkeland N K, Hovland M, Murrell J C. Methane assimilation and trophic interactions with marine Methylomicrobium in deep-water coral reef sediment off the coast of Norway. FEMS Microbiology Ecology, 2008, 66(2): 320-330. DOI:10.1111/j.1574-6941.2008.00575.x |
| [50] |
Rincón-Tomás B, Duda J P, Somoza L, González F J, Schneider D, Medialdea T, Santofimia E, López-Pamo E, Madureira P, Hoppert M, Reitner J. Cold-water corals and hydrocarbon-rich seepage in Pompeia Province (Gulf of Cádiz)-living on the edge. Biogeosciences, 2019, 16(7): 1607-1627. DOI:10.5194/bg-16-1607-2019 |
| [51] |
Middelburg J J, Mueller C E, Veuger B, Larsson A I, Form A, van Oevelen D. Discovery of symbiotic nitrogen fixation and chemoautotrophy in cold-water corals. Scientific Reports, 2016, 5: 17962. |
| [52] |
Osman E O, Vohsen S A, Girard F, Cruz R, Glickman O, Bullock L M, Anderson K E, Weinnig A M, Cordes E E, Fisher C R, Baums I B. Capacity of deep-sea corals to obtain nutrition from cold seeps aligned with microbiome reorganization. Global Change Biology, 2023, 29(1): 189-205. DOI:10.1111/gcb.16447 |
| [53] |
Suttle C A. Viruses in the sea. Nature, 2005, 437(7057): 356-361. DOI:10.1038/nature04160 |
| [54] |
Suttle C A. Marine viruses-major players in the global ecosystem. Nature Reviews Microbiology, 2007, 5(10): 801-812. DOI:10.1038/nrmicro1750 |
| [55] |
Thingstad T F. Elements of a theory for the mechanisms controlling abundance, diversity, and biogeochemical role of lytic bacterial viruses in aquatic systems. Limnology and Oceanography, 2000, 45(6): 1320-1328. DOI:10.4319/lo.2000.45.6.1320 |
| [56] |
Payet J P, McMinds R, Burkepile D E, Vega Thurber R L. Unprecedented evidence for high viral abundance and lytic activity in coral reef waters of the South Pacific Ocean. Frontiers in Microbiology, 2014, 5: 493. |
| [57] |
Knowles B, Silveira C B, Bailey B A, Barott K, Cantu V A, Cobián-Güemes A G, Coutinho F H, Dinsdale E A, Felts B, Furby K A, George E E, Green K T, Gregoracci G B, Haas A F, Haggerty J M, Hester E R, Hisakawa N, Kelly L W, Lim Y W, Little M, Luque A, McDole-Somera T, McNair K, de Oliveira L S, Quistad S D, Robinett N L, Sala E, Salamon P, Sanchez S E, Sandin S, Silva G G Z, Smith J, Sullivan C, Thompson C, Vermeij M J A, Youle M, Young C, Zgliczynski B, Brainard R, Edwards R A, Nulton J, Thompson F, Rohwer F. Lytic to temperate switching of viral communities. Nature, 2016, 531(7595): 466-470. DOI:10.1038/nature17193 |
| [58] |
Liang X L, Zhang Y Y, Wommack K E, Wilhelm S W, DeBruyn J M, Sherfy A C, Zhuang J, Radosevich M. Lysogenic reproductive strategies of viral communities vary with soil depth and are correlated with bacterial diversity. Soil Biology and Biochemistry, 2020, 144: 107767. DOI:10.1016/j.soilbio.2020.107767 |
| [59] |
Voigt E, Rall B C, Chatzinotas A, Brose U, Rosenbaum B. Phage strategies facilitate bacterial coexistence under environmental variability. PeerJ, 2021, 9: e12194. DOI:10.7717/peerj.12194 |
| [60] |
Touchon M, Bernheim A, Rocha E P. Genetic and life-history traits associated with the distribution of prophages in bacteria. The ISME Journal, 2016, 10(11): 2744-2754. DOI:10.1038/ismej.2016.47 |
| [61] |
Uta Passow, Craig A. The biological pump in a high CO2 world. Marine Ecology Progress Series, 2012, 470: 249-271. DOI:10.3354/meps09985 |
| [62] |
覃良云, 许勇前, 陈金妮, 牛天祎, 余克服, 梁甲元. 造礁石珊瑚共生虫黄藻离体培养方法的优化. 微生物学报, 2023, 63(4): 1658-1671. |
| [63] |
Wooldridge S A, Done T J. Improved water quality can ameliorate effects of climate change on corals. Ecological Applications, 2009, 19(6): 1492-1499. DOI:10.1890/08-0963.1 |
| [64] |
Silveira C B, Cavalcanti G S, Walter J M, Silva-Lima A W, Dinsdale E A, Bourne D G, Thompson C C, Thompson F L. Microbial processes driving coral reef organic carbon flow. FEMS Microbiology Reviews, 2017, 41(4): 575-595. DOI:10.1093/femsre/fux018 |
| [65] |
McMahon K W, Thorrold S R, Houghton L A, Berumen M L. Tracing carbon flow through coral reef food webs using a compound-specific stable isotope approach. Oecologia, 2016, 180(3): 809-821. DOI:10.1007/s00442-015-3475-3 |
| [66] |
Christine F, Stephane M, Renaud G, Jonathan C, Eli S, Dan T. Tracing the trophic plasticity of the coral-dinoflagellate symbiosis using amino acid compound-specific stable isotope analysis. Microorganisms, 2021, 9(1): 182. DOI:10.3390/microorganisms9010182 |
| [67] |
McIlroy S, TerHorst C, Teece M, Coffroth M. Nutrient dynamics in coral symbiosis depend on both the relative and absolute abundance of Symbiodiniaceae species. Microbiome, 2022, 10: 1921. |
| [68] |
Zhou Z, Tang J A, Cao X C, Wu C L, Cai W Q, Lin S J. High heterotrophic plasticity of massive coral Porites pukoensis contributes to its tolerance to bioaccumulated microplastics. Environmental Science & Technology, 2023, 57(8): 3391-3401. |
| [69] |
Jones R J, Hoegh-Guldberg O, Larkum A W D, Schreiber U. Temperature-induced bleaching of corals begins with impairment of the CO2 fixation mechanism in zooxanthellae. Plant, Cell and Environment, 1998, 21(12): 1219-1230. DOI:10.1046/j.1365-3040.1998.00345.x |
| [70] |
Xu Y, Zhang J, Huang H, Yuan X C, Zhang J X, Ge J Z. Coral symbiosis carbon flow: a numerical model study spanning cellular to ecosystem levels. Frontiers in Marine Science, 2022, 9: 749921. DOI:10.3389/fmars.2022.749921 |
| [71] |
Cotovicz L C, Chielle R, Marins R V. Air-sea CO2 flux in an equatorial continental shelf dominated by coral reefs (Southwestern Atlantic Ocean). Continental Shelf Research, 2020, 204: 104175. DOI:10.1016/j.csr.2020.104175 |
| [72] |
Suzuki A, Kawahata H. Carbon budget of coral reef systems: an overview of observations in fringing reefs, barrier reefs and atolls in the Indo-Pacific regions. Tellus Series B-Chemical And Physical Meteorology, 2003, 55(2): 428-444. DOI:10.3402/tellusb.v55i2.16761 |
| [73] |
Kayanne H, Suzuki A, Saito H. Diurnal changes in the partial pressure of carbon dioxide in coral reef water. Science, 1995, 269(5221): 214-216. DOI:10.1126/science.269.5221.214 |
| [74] |
Fagan K E, MacKenzie F T. Air-sea CO2 exchange in a subtropical estuarine-coral reef system, Kaneohe Bay, Oahu, Hawaii. Marine Chemistry, 2007, 106(1/2): 174-191. |
| [75] |
Lønborg C, Calleja M L, Fabricius K E, Smith J N, Achterberg E P. The Great Barrier Reef: a source of CO2 to the atmosphere. Marine Chemistry, 2019, 210: 24-33. DOI:10.1016/j.marchem.2019.02.003 |
| [76] |
Akhand A, Watanabe K, Chanda A, Tokoro T, Chakraborty K, Moki H, Tanaya T, Ghosh J, Kuwae T. Lateral carbon fluxes and CO2 evasion from a subtropical mangrove-seagrass-coral continuum. Science of the Total Environment, 2021, 752: 142190. DOI:10.1016/j.scitotenv.2020.142190 |
| [77] |
Hughes T P, Huang H, Young M A L. The wicked problem of China's disappearing coral reefs. Conservation Biology, 2013, 27(2): 261-269. DOI:10.1111/j.1523-1739.2012.01957.x |
| [78] |
Burmester E M, Breef-Pilz A, Lawrence N F, Kaufman L, Finnerty J R, Rotjan R D. The impact of autotrophic versus heterotrophic nutritional pathways on colony health and wound recovery in corals. Ecology and Evolution, 2018, 8(22): 10805-10816. DOI:10.1002/ece3.4531 |
| [79] |
Jia J J, Sun K, Lü S D, Li M X, Wang Y F, Yu G R, Gao Y. Determining whether Qinghai-Tibet Plateau waterbodies have acted like carbon sinks or sources over the past 20 years. Science Bulletin, 2022, 67(22): 2345-2357. DOI:10.1016/j.scib.2022.10.023 |
| [80] |
Xu H, Shi G, Liao B, Chen G, Dong S, Qiu L, Li Q, Luo J, Shen J, Zeng X, Wang Y, Su D, He J, Dong G, Yan G, Wang Y, Zhou X, Hu X, Luo Q, Chen Z, Tao M, Na Q, Wang Q, Wang Y, Shen J, Ma X, Fu H, Wu H, Ma Y, Chen S. Coral-coral reefs in China seas: the biodiversity characteristics of coral-coral reefs in the central South China Sea. Journal of Palaeogeography, 2021, 23(4): 771-788. |
| [81] |
江志坚, 黄小平. 珊瑚虫-虫黄藻共生系统碳循环研究的若干进展. 海洋科学进展, 2009, 27(1): 112-120. |
| [82] |
Howe S A, Marshall A T. Temperature effects on calcification rate and skeletal deposition in the temperate coral, Plesiastrea versipora (Lamarck). Journal of Experimental Marine Biology and Ecology, 2002, 275(1): 63-81. DOI:10.1016/S0022-0981(02)00213-7 |
| [83] |
李言达, 易亮. 全球变暖和海洋酸化背景下珊瑚礁生态响应的研究进展. 海洋地质与第四纪地质, 2021, 41(1): 33-41. |
| [84] |
Gattuso J P, Allemand D, Frankignoulle M. Photosynthesis and calcification at cellular, organismal and community levels in coral reefs: a review on interactions and control by carbonate chemistry. Integrative and Comparative Biology, 1999, 39(1): 160-183. |
| [85] |
Langdon C, Takahashi T, Sweeney C, Chipman D, Goddard J, Marubini F, Aceves H, Barnett H, Atkinson M J. Effect of calcium carbonate saturation state on the calcification rate of an experimental coral reef. Global Biogeochemical Cycles, 2000, 14(2): 639-654. DOI:10.1029/1999GB001195 |
| [86] |
张远辉, 陈立奇. 南沙珊瑚礁对大气CO2含量上升的响应. 台湾海峡, 2006, 25(1): 68-76. |
| [87] |
Conti-Jerpe I E, Thompson P D, Wong C W M, Oliveira N L, Duprey N N, Moynihan M A, Baker D M. Trophic strategy and bleaching resistance in reef-building corals. Science Advances, 2020, 6(15): eaaz5443. DOI:10.1126/sciadv.aaz5443 |
| [88] |
Motone K, Takagi T, Aburaya S, Aoki W, Miura N, Minakuchi H, Takeyama H, Nagasaki Y, Shinzato C, Ueda M. Protection of coral larvae from thermally induced oxidative stress by redox nanoparticles. Marine Biotechnology, 2018, 20(4): 542-548. DOI:10.1007/s10126-018-9825-5 |
| [89] |
Radice Veronica Z, Ove H, Brian F, Fox Michael D, Dove Sophie G. Upwelling as the major source of nitrogen for shallow and deep reef-building corals across an oceanic atoll system. Functional Ecology, 2019, 33(6): 1120-1134. DOI:10.1111/1365-2435.13314 |
| [90] |
Barshis D J, Ladner J T, Oliver T A, Seneca F O, Traylor-Knowles N, Palumbi S R. Genomic basis for coral resilience to climate change. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(4): 1387-1392. |
| [91] |
王丽荣, 于红兵, 李翠田, 孙妮. 海洋生态系统修复研究进展. 应用海洋学学报, 2018, 37(3): 435-446. |
| [92] |
Zong J F, Chen L A, Sun L A, Guo H T. Analysis of domestic and overseas carbon trading market. E3S Web of Conferences, 2018, 53: 03006. DOI:10.1051/e3sconf/20185303006 |
| [93] |
谢素美, 罗伍丽, 贺义雄, 黄华梅, 李春林. 中国海洋碳汇交易市场构建. 科技导报, 2021, 39(24): 84-95. |
| [94] |
何磊, 叶思源, 赵广明, 谢柳娟, 裴绍峰, 丁喜桂, 杨士雄, Hans Brix, Edward A.Laws.. 海岸带滨海湿地蓝碳管理的研究进展. 中国地质, 2023, 50(3): 777-794. |
| [95] |
曹云梦, 吴婧. "双碳"目标下我国海洋碳汇交易的发展机制研究. 中国环境管理, 2022, 14(4): 44-51. |