 2024, Vol. 44
2024, Vol. 44文章信息
- 邓楚可, 黄青峰, 李平, 夏静怡, 夏继刚
- DENG Chuke, HUANG Qingfeng, LI Ping, XIA Jingyi, XIA Jigang
- 不同生活史阶段秦岭细鳞鲑与同域物种拉氏鱥爆发游泳能力比较研究
- Comparative studies on burst swimming performance of Brachymystax tsinlingensis and sympatric Phoxinus lagowskii in different life history stages
- 生态学报. 2024, 44(9): 3999-4008
- Acta Ecologica Sinica. 2024, 44(9): 3999-4008
- http://dx.doi.org/10.20103/j.stxb.202307061451
-
文章历史
- 收稿日期: 2023-07-06
- 网络出版日期: 2024-02-01





2. 进化生理与行为学实验室, 重庆市动物生物学重点实验室, 重庆师范大学生命科学学院, 重庆 401331;
3. 中国电建集团西北勘测设计研究院有限公司, 西安 710065
2. Laboratory of Evolutionary Physiology and Behavior, Chongqing Key Laboratory of Animal Biology, College of Life Sciences, Chongqing Normal University, Chongqing 401331, China;
3. Power-China Northwest Engineering Corporation Limited, Xi'an 710065, China
游泳运动是鱼类集群、快速追击捕食或躲避敌害、生殖洄游、穿越激流与堤坝等生态过程实现的主要方式[1—2], 为其生存、生长及繁衍提供了基本保证, 具有重要的生态学意义。因此, 游泳能力也被看作是表征鱼类生存适合度的关键组分。感应流速(Inductive velocity)、快速启动游泳(Fast-start swimming)、临界游泳速度(Critical swimming speed, Ucrit)和爆发游泳速度(Burst swimming speed, Uburst)等是评估鱼类游泳性能的几个关键指标[3]。通常认为鱼类的长距离生殖与索饵洄游能力主要与有氧运动能力(例如Ucrit)有关; 而鱼类完成捕获猎物与逃避猎食者、穿越激流寻觅适宜生境等主要与无氧运动能力(例如Uburst)有关[4—5]。水利水电开发等涉水工程建设往往造成山区溪流连通性降低, 导致鱼类生境片段化、破碎化等一系列负面生态问题[6—8]。栖息地破坏对溪流鱼类(尤其是洄游性鱼类)的生存繁衍构成了严重威胁, 同时也对鱼类爆发游泳能力提出了更高要求, 有关溪流生态系统关键鱼类爆发游泳能力及其种内种间差异的研究是开展种群保护、栖息地管理与生态修复的基础与前提[9—10]。
秦岭细鳞鲑(Brachymystax tsinlingensis)隶属于鲑形目(Salmoniformes)、鲑科(Salmonidae)、细鳞鲑属(Brachymystax), 俗称梅花鱼, 为我国珍稀特有种、国家Ⅱ级重点保护野生动物。该物种是第四纪冰川残留种、北半球鲑科鱼类中分布在最南端的两物种之一, 为典型的陆封型冷水性山麓鱼类, 主要分布在秦岭海拔1000 m以上的山涧溪流中, 分布区涉及长江流域汉江水系(湑水河、酉水河、子午河等)以及黄河流域渭河水系(石头河、千河、黑河等)[11—16]。近年来, 由于全球气候变化以及人类活动干扰造成生存环境恶化, 秦岭细鳞鲑的分布范围呈现碎片化趋势、野生种群数量锐减[12]。调查研究发现, 秦岭细鳞鲑生境特殊, 水文地理环境复杂, 多大高差地貌与湍急激流, 常需要进行多次重复爆发游泳运动以穿越激流甚至越过堤坝阻隔, 因此爆发游泳能力及其可持续性对该物种生存至关重要。然而, 对该物种的研究集中于资源调查、物种鉴定、驯养繁殖、种群遗传和热生物学等方面[13—14], 对其不同生活史阶段该游泳性能(尤其爆发游泳能力)至今尚无相关研究。
秦岭细鳞鲑栖息地鱼类群落结构简单, 同域分布鱼类主要为拉氏鱥(Phoxinus lagowskii)和少量鳅类。拉氏鱥隶属于鲤形目(Cypriniformes)、鲤科(Cyprinidae)、鱥属(Phoxinus), 为小型野生经济鱼类, 杂食性, 主要以水生昆虫、软体动物、浮游动物及水生植物为食; 分布范围较广, 常成群生活于水流较急、清澈、溶氧量高、温度低的山区河沟与小溪环境[17—18]。秦岭细鳞鲑发育较慢, 通常三龄性成熟, 而拉氏鱥小型成体、二龄即可性成熟, 二者在生境选择上具有一定的相似性, 但其栖息地重叠程度依生活史阶段不同而有较大差异[18—19]。一般认为, 秦岭细鳞鲑处于溪流生态系统食物链顶端、是顶级捕食者, 拉氏鱥是秦岭细鳞鲑主要的饵料生物之一。然而, 作为同域分布物种, 二者的种间关系又在很大程度上受到生活史动态变化和栖息地重叠程度的影响; 特别是, 尽管拉氏鱥处于被捕食者地位, 但其可能对秦岭细鳞鲑胚胎造成捕食压力、且对早期生活史阶段秦岭细鳞鲑幼体在食物资源和生存空间等多方面构成竞争。鉴于此, 揭示两物种不同生活史阶段游泳能力的动态变化及其种间差异就成为迫切值得探究的科学问题。在种间水平上, 作为捕食关系紧密、栖息环境与生存需求相异的同域物种, 环境选择压力是否驱动二者进化出不同的爆发游泳性能?在种内水平上, 不同生活史阶段秦岭细鳞鲑生态习性不同、栖息环境存在较大差异, 而各生活史阶段拉氏鱥栖息环境相对趋于交叠, 生活史阶段效应是否驱动两物种爆发游泳性能表现出不同的种内变异模式?
为探究秦岭细鳞鲑及其同域物种拉氏鱥爆发游泳能力的种内种间差异, 本研究采用自制的仿生态鱼类爆发游泳能力测定装置, 分别测定了不同生活史阶段两种实验鱼的相对和绝对Uburst及其可持续性, 以期为秦岭溪流生态系统生物多样性保护以及未来秦岭细鳞鲑栖息地生态修复提供理论基础。
1 材料与方法 1.1 实验动物实验鱼采集自陕西陇县秦岭细鳞鲑国家级自然保护区科研基地, 共采集到健康活泼的秦岭细鳞鲑63尾、拉氏鱥64尾, 二者皆分布于蒲峪河流域(渭河水系千河一级支流)。根据两种实验鱼个体大小及其体长特征[18—19], 将采集到的实验鱼划分成不同的生活史阶段, 具体规格见表 1。将实验鱼依据种类及其生活史阶段分列成不同的实验组, 分别置于圆形自净化循环控温水槽(直径2 m, 水深1 m)中驯养适应48 h。驯养用水为实验鱼自然栖息地经过沉淀净化处理后的河水, 实验期间实时监测水槽内水温与溶氧的变化, 将水温控制在(17±0.5)℃, 水体溶氧>7 mg/L, 采用自然光照。
| 种类 Species |
生活史阶段 Life history stage |
体长/cm Body length |
体重/g Body weight |
样本量 n |
| 秦岭细鳞鲑 | 成体 | 18.79±0.284 | 108.25±1.301 | 33 |
| Brachymystax tsinlingensis | 亚成体 | 13.27±0.367 | 38.54±0.848 | 16 |
| 幼体 | 7.08±0.122 | 4.17±0.092 | 14 | |
| 拉氏鱥 | 成体 | 9.62±0.146 | 13.54±0.145 | 45 |
| Phoxinus lagowskii | 亚成体 | 6.89±0.166 | 4.98±0.087 | 19 |
实验所用的鱼类仿生态游泳能力测定装置由本研究团队自行研发。该装置主体为一长直型泳道(图 1), 并专设有适应区、隐蔽区和开阔区, 装置底部模拟自然河道铺设有碎石。泳道起点处(隐蔽区1)上方加盖有遮光板, 并配备可上下拉伸的活动式闸门以及强光刺激装置; 开阔区规格为5 m×0.25 m×1 m; 泳道终点另设置遮光隐蔽区(隐蔽区2)。泳道旁平行设置5 m钢直尺, 以实时监测实验鱼位移变动; 开阔区上方架设有与电脑连接的摄像头(Logitech Webcam Pro 920), 摄像头距离水面2 m。装置中水深10 cm, 由循环控温系统将水温控制在(17±0.5)℃, 水体溶氧>7 mg/L, 泳道末端提供微水流(流速约3 cm/s)。

|
| 图 1 仿生态鱼类爆发游泳能力测定装置 Fig. 1 Ecologically simulated apparatus for the determination of fish burst swimming ability |
将单尾实验鱼置入鱼类仿生态游泳能力测定装置的适应区(图 1)并使其适应15 min, 之后开始爆发游泳速度测定。测定时, 快速揭开隐蔽区1上方的遮光板, 开启强光刺激装置并用玻璃棒对实验鱼进行追捕刺激, 诱导实验鱼产生爆发游泳。测定期间全程录像, 为验证实验鱼爆发游泳速度测定的可持续性, 对每尾实验鱼连续两次进行测定, 两次测定的时间间隔为10 min。
采用视频处理软件Adobe Premiere Pro对录制视频进行解析。实验录制视频为30帧/s, 按帧数播放并定位每帧画面中实验鱼的位置, 以每3帧(即0.1 s)为单位提取实验鱼爆发运动的距离参数, 选取距离最长的一段用于计算其绝对和相对爆发游泳速度。
绝对爆发游泳速度(Uburst)计算公式如下:
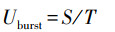
|
式中: Uburst为绝对爆发游泳速度(cm/s), S是实验鱼瞬时爆发游泳冲刺的距离(cm), T是实验鱼爆发游泳逃逸的时间(s)。
相对爆发游泳速度(rUburst)计算公式如下:
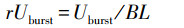
|
式中, rUburst(Relative burst swimming speed)为相对爆发游泳速度(BL/s), Uburst为绝对爆发游泳速度(cm/s), BL为实验鱼体长(cm)。
1.4 数据处理采用SPSS软件(SPSS 20.0 Inc., USA)对数据进行统计分析。首先检验数据是否服从正态分布(Shapiro-Wilks检验)及方差齐性。通过计算组内相关系数(Intraclass correlation coefficient, ICC)评估实验鱼爆发游泳速度(Uburst和rUburst)的可持续性; 采用双因素方差分析(Two-way ANOVA)检验物种和生活史阶段对Uburst和rUburst的影响, 采用单因素方差分析(One-way ANOVA)和Tukey′s HSD多重比较检验物种间以及种内不同生活史阶段的差异。数据均以平均值±标准误(Mean±SE)表示, 显著性水平设为P < 0.05。
2 结果与分析 2.1 爆发游泳能力测定的可持续性重复测定对不同生活史阶段秦岭细鳞鲑的Uburst和rUburst均无显著影响(P>0.05, 表 2, 图 2);重复测定对不同生活史阶段拉氏鱥的Uburst和rUburst均影响显著, 导致其Uburst和rUburst均显著低于首次测定(P < 0.05, 表 2, 图 3)。
| 种类 Species |
生活史阶段 Life history stage |
Uburst | rUburst |
| 秦岭细鳞鲑 | 成体 | F=0.035, P=0.852 | F=0.014, P=0.970 |
| Brachymystax tsinlingensis | 亚成体 | F=0.170, P=0.683 | F=0.402, P=0.531 |
| 幼体 | F=0.142, P=0.709 | F=0.091, P=0.765 | |
| 拉氏鱥 | 成体 | F=4.457, P=0.038* | F=4.556, P=0.036* |
| Phoxinus lagowskii | 亚成体 | F=4.226, P=0.047* | F=4.965, P=0.032* |
| Uburst: 爆发游泳速度Burst swimming speed; rUburst: 相对爆发游泳速度Relative burst swimming speed | |||

|
| 图 2 秦岭细鳞鲑爆发游泳速度测定的可持续性 Fig. 2 Repeatability of burst swimming speed measurements in Brachymystax tsinlingensis Uburst: 爆发游泳速度Burst swimming speed; rUburst: 相对爆发游泳速度Relative burst swimming speed |

|
| 图 3 拉氏鱥爆发游泳速度测定的可持续性 Fig. 3 Repeatability of burst swimming speed measurements in Phoxinus lagowskii Uburst: 爆发游泳速度Burst swimming speed; rUburst: 相对爆发游泳速度Relative burst swimming speed |
对不同生活史阶段两种实验鱼Uburst两次连续测定组内相关系数ICC的分析结果见表 3。经检验, 所有参数两次连续测定值均显著相关(P < 0.001)且ICC系数均大于0.75, 表明不同生活史阶段两种实验鱼游泳能力均具有可持续性(图 2, 图 3)。
| 种类 Species |
生活史阶段 Life history stage |
因变量 Variables |
组内相关系数ICC Intraclass correlation coefficient |
95% CI |
| 秦岭细鳞鲑 | 成体 | Uburst | 0.965(P < 0.001) | 0.930—0.983 |
| Brachymystax tsinlingensis | rUburst | 0.967(P < 0.001) | 0.929—0.985 | |
| 亚成体 | Uburst | 0.973(P < 0.001) | 0.924—0.991 | |
| rUburst | 0.932(P < 0.001) | 0.805—0.976 | ||
| 幼体 | Uburst | 0.970(P < 0.001) | 0.907—0.990 | |
| rUburst | 0.970(P < 0.001) | 0.906—0.990 | ||
| 拉氏鱥 | 成体 | Uburst | 0.891(P < 0.001) | 0.802—0.940 |
| Phoxinus lagowskii | rUburst | 0.882(P < 0.001) | 0.786—0.935 | |
| 亚成体 | Uburst | 0.867(P < 0.001) | 0.777—0.914 | |
| rUburst | 0.822(P < 0.001) | 0.759—0.893 |
物种、生活史阶段及其交互作用对实验鱼Uburst和rUburst均影响显著(P < 0.05, 表 4)。
| 测定 Measurement |
因变量 Variables |
物种 Species |
生活史阶段 Life history stage |
交互作用 Interaction effect |
| 首次测定 | Uburst | F=411.359, P < 0.001 | F=114.020, P < 0.001 | F=9.381, P=0.003 |
| First measurement | rUburst | F=15.121, P < 0.001 | F=60.146, P < 0.001 | F=7.031, P=0.009 |
| 重复测定 | Uburst | F=471.279, P < 0.001 | F=106.547, P < 0.001 | F=8.731, P=0.004 |
| Repeat measurement | rUburst | F=10.607, P < 0.001 | F=56.651, P < 0.001 | F=4.600, P=0.034 |
对于Uburst, 无论首次还是重复测定, 在相同的生活史阶段种间Uburst均差异显著(P < 0.05, 表 5), 表现为: 秦岭细鳞鲑>拉氏鱥(图 4);种内不同生活史阶段Uburst均差异显著(P < 0.05, 表 6), 均表现为: 成体>亚成体>幼体(图 4)。
| 测定Measurement | Uburst | rUburst | |||||
| 拉氏鱥Phoxinus lagowskii | 拉氏鱥Phoxinus lagowskii | ||||||
| 成体 | 亚成体 | 成体 | 亚成体 | ||||
| 首次测定First measurement | 秦岭细鳞鲑Brachymystax tsinlingensis | 成体 | F=322.172, P < 0.001 | F=217.784, P < 0.001 | F=1.019, P=0.316 | F=68.913, P < 0.001 | |
| 亚成体 | F=157.803, P < 0.001 | F=235.380, P < 0.001 | F=8.155, P=0.006 | F=9.378, P=0.003 | |||
| 幼体 | F=0.452, P=0.504 | F=4.031, P=0.052 | F=48.584, P < 0.001 | F=0.193, P=0.663 | |||
| 重复测定Repeat measurement | 秦岭细鳞鲑Brachymystax tsinlingensis | 成体 | F=353.618, P < 0.001 | F=239.172, P < 0.001 | F=1.227, P=0.272 | F=37.972, P < 0.001 | |
| 亚成体 | F=186.462, P < 0.001 | F=288.799, P < 0.001 | F=11.550, P=0.002 | F=4.147, P=0.051 | |||
| 幼体 | F=1.239, P=0.270 | F=5.565, P=0.021 | F=66.975, P < 0.001 | F=6.436, P=0.016 | |||

|
| 图 4 首次和重复测定下实验鱼绝对爆发游泳速度的种内种间差异(平均值±标准误) Fig. 4 Intraspecific and interspecific differences in absolute burst swimming speed of the experimental fish under the first and repeated measurements Uburst: 爆发游泳速度Burst swimming speed; a-c, x-y字母不同分别表示不同生活史阶段秦岭细鳞鲑和拉氏鱥种内Uburst差异显著(P < 0.05); *、**、***表示特定生活史阶段物种间Uburst差异显著性水平为P < 0.05、P < 0.01、P < 0.001 |
| 测定 Measurement |
因变量 Variables |
秦岭细鳞鲑 Brachymystax tsinlingensis |
拉氏鱥 Phoxinus lagowskii |
| 首次测定 | Uburst | F=79.759, P < 0.001 | F=8.242, P=0.006 |
| First measurement | rUburst | F=33.724, P < 0.001 | F=60.184, P < 0.001 |
| 重复测定 | Uburst | F=77.115, P < 0.001 | F=5.566, P=0.021 |
| Repeat measurement | rUburst | F=36.531, P < 0.001 | F=47.773, P < 0.001 |
对于rUburst, 无论首次还是重复测定, 成体rUburst种间均无显著差异(P>0.05, 表 5), 首次测定亚成体rUburst种间差异显著(P < 0.05)而重复测定后种间无显著差异(P>0.05, 表 5);种内不同生活史阶段rUburst均差异显著(P < 0.05, 表 6), 表现为: 成体 < 亚成体 < 幼体(图 5)。

|
| 图 5 首次和重复测定下实验鱼相对爆发游泳速度的种内种间差异(平均值±标准误) Fig. 5 Intraspecific and interspecific differences in relative burst swimming speed of the experimental fish under the first and repeated measurements rUburst: 相对爆发游泳速度Relative burst swimming speed; a-c, x-y字母不同分别表示不同生活史阶段秦岭细鳞鲑和拉氏鱥种内rUburst差异显著(P < 0.05); *、**、***表示特定生活史阶段物种间rUburst差异显著性水平为P < 0.05、P < 0.01、P < 0.001 |
目前, 对鱼类游泳能力的测定尚无统一的标准化实验装置。对Ucrit测定国际上通常采用“流速递增法”[20], 但由于爆发游泳运动的非稳定特性, 采用该方法衡量鱼类爆发游泳能力难度较大[21], 具体测定中研究者大都采用匀加速游泳速度(Constant acceleration test speed, Ucat)表示或替代实验鱼的Uburst[22—23]。Ucat测定方法为将实验鱼置入游泳管、适应后以某一恒定的加速度持续增加水流速度直至鱼体到达力竭状态, 该方法的优点是测试环境稳定、测试结果具有良好的可重复性, 其缺点是Ucat完全依赖于持续的水流刺激诱导、缺乏足够的生态关联。本研究发明了一种仿生态鱼类爆发游泳能力测定装置, 该装置最大程度地模拟实验鱼野外生境, 通过突发性的刺激诱导实验鱼自主完成爆发逃逸过程。本研究发现, 不同组别鱼类两次测定结果的ICC系数均大于0.75(表 3), 表明该测定方法具有较好的可靠性与可重复性[24]。
重复游泳表现可用于评估暴露于应激源后鱼类生理性能的恢复率, 被认为是比单一运动测试更敏感的生态功能指标[25]。Killen等[26]研究表明, 饱足摄食组欧洲鲈(Dicentrarchus labrax)重复游泳表现明显高于饥饿组和低摄食水平组; Wagner等[27]发现改变饲料脂肪酸组成会降低大西洋鲑(Salmo salar)在重复测定中的游泳表现, 推测这可能与食物脂肪成分影响能量产生有关。本研究中, 不同生活史阶段秦岭细鳞鲑Uburst和rUburst的首次测定和重复测定结果均无显著差异, 而拉氏鱥成体和亚成体重复测定结果均显著低于首次测定(表 2), 表明与拉氏鱥相比、秦岭细鳞鲑无氧代谢恢复能力更强, 推测Uburst恢复力的种间差异可能与两物种异化的生境选择与环境适应方式有关(野外生境下秦岭细鳞鲑需要反复地穿越激流甚至向上跃起穿过叠水区寻找产卵场或其它适宜生境, 而拉氏鱥对此并无强烈生态需求); 研究结果与彭敏锐等[28]对秦岭细鳞鲑步法转换速度(Gait transition speed)以及Ucat的测定结果相近。
3.2 两种实验鱼Uburst的种间差异自然选择压力塑造鱼类游泳能力的进化方向, 生存于不同环境(例如急流vs缓流生境)的鱼类游泳能力存在明显差异[29], 但同域分布物种游泳能力是否存在趋同适应尚无定论[30]。Raffini等[31]研究发现位于同域火山口湖的阿氏双冠丽鱼(Amphilophus astorquii)与札里双冠丽鱼(A. zaliosus)的急加速运动能力(Sprint swimming speed)无显著差异, 但偏好开阔区的札里双冠丽鱼Ucrit高于喜底栖的阿氏双冠丽鱼、且Ucrit提高与较高的氧需求有关, 异质栖息地中的氧供给差异可能在同域物种适应性分化中起重要作用; Myrick等[32]考察了不同温度下同域分布物种臼齿鱼(Mylopharodon conocephalus)、细尾美拟鲴(Lavinia exilicauda)和丰满叶唇鱼(Ptychocheilus grandis)的游泳能力, 发现它们具有相近的游泳速度、且以类似的方式对温度变化作出反应, 这可能是由于相似环境选择压力造成的趋同进化所致; 而Yamada等[33]研究结果则表明, 6 ℃时花羔红点鲑(Salvelinus malma)的游泳能力强于同域分布的白点鲑(Salvelinus leucomaenis)、但在12 ℃时游泳能力无种间差异, 结合两者不同温度下优势地位发生改变的背景, 推测游泳能力受季节性温度变化的分化可能介导种间竞争结果, 从而影响二者共存; 此外, 对同域分布且生态位重叠的食蚊鱼(Gambusia affinis)和唐鱼(Tanichthys albonubes)的研究发现, 长期栖息于流动性更大水环境中的唐鱼Ucat和Ucrit均高于生存于水体流动较小生境中的食蚊鱼, 提示微生境选择分化可能驱动同域物种游泳能力表型的种间差异[34]。本研究中, 总体上秦岭细鳞鲑Uburst高于拉氏鱥, 两物种Uburst差异显著、不存在趋同适应。除遗传因素外, 我们推测生态习性不同可能是该同域物种爆发游泳能力出现差异的主要原因。一方面, 秦岭细鳞鲑生殖与索饵洄游强烈、经常需要穿越各种复杂激流生境、为洄游性鱼类, 而拉氏鱥则相对趋于定居性、生存范围变动不大; 另一方面, 秦岭细鳞鲑为肉食性鱼类且以拉氏鱥为主要饵料生物、更高的Uburst有利于提高捕食成功率, 而拉氏鱥为杂食性鱼类、相较而言对爆发游泳能力的依赖不强。值得注意的是, 尽管秦岭细鳞鲑成体和亚成体Uburst均高于拉氏鱥(无论是其成体还是亚成体), 在首次测定中秦岭细鳞鲑幼体与拉氏鱥成体和亚成体Uburst均无显著差异, 其相关种间互作机制有待于进一步研究。
3.3 两种实验鱼Uburst的种内变异(生活史阶段效应)物种生态性能通常随生活史发育历程而动态变化[18, 35—36], 因此对溪流鱼类游泳能力的评估应特别关注生活史阶段效应。Kopf等[37]测定了澳大利亚突吻麦鳕鲈(Maccullochella macquariensis)、虫纹麦鳕鲈(Maccullochella peelii)、金鲈(Macquaria ambigua)等6种土著淡水鱼类早期生活史阶段Ucrit和疲劳游泳速度(Prolonged swimming speed), 发现Ucrit随幼鱼的生长发育而增加, 且相比于体长、大小、年龄等因素, 发育阶段与游泳能力的相关性更高; 类似地, 高泉等[38]发现, 6月龄斑马鱼(Danio rerio)普通成体与30月龄衰老期个体相比, 6月龄个体的Uburst、游泳距离和持续时间等均显著高于衰老期个体; 然而, Swanson等[39]发现, 生活史阶段效应对越洋公鱼(Hypomesus transpacificus)Ucrit及其游泳耐力均无显著影响, 作者推测该结果可能与实验鱼体型较小、随生活史阶段变化不明显有关。本研究中, 生活史阶段效应对秦岭细鳞鲑和拉氏鱥Uburst、rUburst均影响显著; 两物种爆发游泳能力种内变异模式相近, Uburst均随发育年龄增加而增加、呈现出“成体>亚成体>幼体”模式, rUburst均随发育年龄增加而减小、呈现出“成体 < 亚成体 < 幼体”模式。这种变化趋势与相关文献中对斑重唇鱼(Diptychus maculatus Steindachner)、青鱼(Mylopharyngodon piceus)和斯氏高原鳅(Triplophysa stoliczkae)等鱼类测定结果相一致[40]。伴随发育历程, Uburst增加可能与机体能量储备、肌肉增长和效率增加等有关, 而rUburst减小则可能是机体表面积增大引起游泳时水阻非线性增加等原因所致[41—42]。
综上, 本实验研制了一种仿生态鱼类爆发游泳能力测定装置, 通过对秦岭细鳞鲑及其同域物种拉氏鱥爆发游泳能力的研究表明该方法可靠性较高; 相较于拉氏鱥, 秦岭细鳞鲑爆发游泳能力具有更强的恢复力; 秦岭细鳞鲑和拉氏鱥爆发游泳能力种内种间差异显著, 二者在种间水平上不存在趋同适应、在种内水平上均表现出显著的生活史阶段效应。研究结果提示, 未来鱼类游泳能力关联的鱼道设计或涉水工程评价中, 应综合考虑生活史阶段效应和不同物种的生态习性及其种间差异。
| [1] |
付世建, 曹振东, 曾令清, 夏继刚, 李秀明. 鱼类游泳运动: 策略与适应性进化. 北京: 科学出版社, 2014, 1-10. |
| [2] |
Merino G E, Piedrahita R H, Conklin D E. Effect of water velocity on the growth of California halibut (Paralichthys californicus) juveniles. Aquaculture, 2007, 271(1-4): 206-215. DOI:10.1016/j.aquaculture.2007.06.038 |
| [3] |
石小涛, 陈求稳, 刘德富, 黄应平, 庄平, 陈志强, 王博, 路波. 胭脂鱼幼鱼的临界游泳速度. 水生生物学报, 2012, 36(1): 133-136. |
| [4] |
Wysujack K, Greenberg L A, Bergman E, Olsson I C. The role of the environment in partial migration: food availability affects the adoption of a migratory tactic in brown trout Salmo trutta. Ecology of Freshwater Fish, 2009, 18(1): 52-59. DOI:10.1111/j.1600-0633.2008.00322.x |
| [5] |
Jones D R. Anaerobic exercise in teleost fish. Canadian Journal of Zoology, 1982, 60(5): 1131-1134. DOI:10.1139/z82-157 |
| [6] |
Barbarossa V, Schmitt R J P, Huijbregts M A J, Zarfl C, King H, Schipper A M. Impacts of current and future large dams on the geographic range connectivity of freshwater fish worldwide. Proceedings of the National Academy of Sciences, 2020, 117(7): 3648-3655. DOI:10.1073/pnas.1912776117 |
| [7] |
Katopodis C, Williams J G. The development of fish passage research in a historical context. Ecological Engineering, 2012, 48: 8-18. DOI:10.1016/j.ecoleng.2011.07.004 |
| [8] |
Laborde A, González A, Sanhueza C, Arriagada P, Wilkes M, Habit E, Link O. Hydropower development, riverine connectivity, and non-sport fish species: criteria for hydraulic design of fishways. River Research and Applications, 2016, 32(9): 1949-1957. DOI:10.1002/rra.3040 |
| [9] |
蔡露, 金瑶, 潘磊, 赵萍, 贺达, 侯轶群, 陈小娟, 张鹏. 过鱼设施设计中的鱼类行为研究与问题. 生态学杂志, 2018, 37(11): 3458-3466. |
| [10] |
Hinch S G, Standen E M, Healey M C, Farrell A P. Swimming patterns and behaviour of upriver-migrating adult pink (Oncorhynchus gorbuscha) and sockeye (O. nerka) salmon as assessed by EMG telemetry in the Fraser River, British Columbia, Canada. In: Thorstad EB, Fleming IA, Næsje TF (eds). Aquatic Telemetry. Dordrecht: Springer, 2002: 147-160.
|
| [11] |
Xia J G, Peng M R, Huang Y, Elvidge C K. Acute warming in winter eliminates chemical alarm responses in threatened Qinling lenok Brachymystax lenok tsinlingensis. Science of the Total Environment, 2021, 764: 142807. DOI:10.1016/j.scitotenv.2020.142807 |
| [12] |
李平, 齐喜荣, 夏继刚, 曹谡杰, 沈红保, 刘其根. 秦岭细鳞鲑野生种群与人工养殖种群肌肉营养成分的差异. 生态学杂志, 2023, 42(1): 140-145. |
| [13] |
夏静怡, 郑雪丽, 李平, 夏继刚. 秦岭细鳞鲑(Brachymystax tsinlingensis)与细鳞鲑(Brachymystax lenok)当年幼鱼几何形态学特征的比较. 生态学杂志, 2023, 42(4): 905-910. |
| [14] |
Xia J G, Ma Y J, Fu C, Fu S J, Cooke S J. Effects of temperature acclimation on the critical thermal limits and swimming performance of Brachymystax lenok tsinlingensis: a threatened fish in Qinling Mountain region of China. Ecological Research, 2017, 32(1): 61-70. DOI:10.1007/s11284-016-1418-z |
| [15] |
夏继刚, 陈梅, 肖静, 付世建. 秦岭细鳞鲑代谢及低氧耐受能力对温度驯化的响应. 水生生物学报, 2017, 41(1): 201-205. |
| [16] |
Zhao Y H, Zhang C G. Threatened fishes of the world: Brachymystax lenok tsinlingensis Li, 1966 (Salmonidae). Environmental Biology of Fishes, 2009, 86(1): 11-12. DOI:10.1007/s10641-008-9337-7 |
| [17] |
Sakai H, Ito Y, Shedko S V, Safronov S N, Frolov S V, Chereshnev I A, Jeon S R, Goto A. Phylogenetic and taxonomic relationships of northern Far Eastern phoxinin minnows, Phoxinus and Rhynchocypris (Pisces, Cyprinidae), as inferred from allozyme and mitochondrial 16S rRNA sequence analyses. Zoological Science, 2006, 23(4): 323-331. DOI:10.2108/zsj.23.323 |
| [18] |
黄青峰, 邓楚可, 夏静怡, 严红娟, 夏继刚. 秦岭细鳞鲑与同域物种拉氏鱥几何形态学特征: 生活史阶段效应及种间差异. 生态学杂志, 2023, 42, Online early. https://link.cnki.net/urlid/21.1148.Q.20230902.1204.002.
|
| [19] |
薛超, 危起伟, 孙庆亮, 吴金明, 李罗新. 秦岭细鳞鲑的年龄与生长. 中国水产科学, 2013, 20(4): 743-749. |
| [20] |
Brett J R. The respiratory metabolism and swimming performance of young sockeye salmon. Journal of the Fisheries Research Board of Canada, 1964, 21(5): 1183-1226. DOI:10.1139/f64-103 |
| [21] |
Kern P, Cramp R L, Gordos M A, Watson J R, Franklin C E. Measuring Ucrit and endurance: equipment choice influences estimates of fish swimming performance. Journal of Fish Biology, 2018, 92(1): 237-247. DOI:10.1111/jfb.13514 |
| [22] |
Hammer C. Fatigue and exercise tests with fish. Comparative Biochemistry and Physiology Part A: Physiology, 1995, 112(1): 1-20. DOI:10.1016/0300-9629(95)00060-K |
| [23] |
Beamish F W H. Swimming capacity. Fish Physiology. New York: Academic Press, 1978: 101-187.
|
| [24] |
Fleiss J L. The equivalence of weighted kappa and the intraclass correlation coefficient as measures of reliability. Educational and Psychological Measurement, 1973, 33(3): 613-619. DOI:10.1177/001316447303300309 |
| [25] |
Jain K E, Birtwell I K, Farrell A P. Repeat swimming performance of mature sockeye salmon following a brief recovery period: a proposed measure of fish health and water quality. Canadian Journal of Zoology, 1998, 76(8): 1488-1496. DOI:10.1139/z98-079 |
| [26] |
Killen S S, Marras S, McKenzie D J. Fast growers sprint slower: effects of food deprivation and re-feeding on sprint swimming performance in individual juvenile European sea bass. Journal of Experimental Biology, 2014, 217(6): 859-865. |
| [27] |
Wagner G N, Balfry S K, Higgs D A, Lall S P, Farrell A P. Dietary fatty acid composition affects the repeat swimming performance of Atlantic salmon in seawater. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2004, 137(3): 567-576. |
| [28] |
彭敏锐, 郑雪丽, 李平, 李秀明, 夏继刚. 温度和重复测定对秦岭细鳞鲑快速启动反应、游泳性能及力竭后代谢特征的影响. 生态学报, 2021, 41(6): 2505-2514. |
| [29] |
Taylor E B, McPhail J D. Variation in burst and prolonged swimming performance among British Columbia populations of coho salmon, Oncorhynchus kisutch. Canadian Journal of Fisheries and Aquatic Sciences, 1985, 42(12): 2029-2033. DOI:10.1139/f85-250 |
| [30] |
Blake R W. Functional design and burst-and-coast swimming in fishes. Canadian Journal of Zoology, 1983, 61(11): 2491-2494. DOI:10.1139/z83-330 |
| [31] |
Raffini F, Schneider R F, Franchini P, Kautt A F, Meyer A. Diving into divergence: differentiation in swimming performances, physiology and gene expression between locally-adapted sympatric cichlid fishes. Molecular Ecology, 2020, 29(7): 1219-1234. DOI:10.1111/mec.15304 |
| [32] |
Myrick C A, Cech J J. Swimming performances of four California stream fishes: temperature effects. Environmental Biology of Fishes, 2000, 58(3): 289-295. DOI:10.1023/A:1007649931414 |
| [33] |
Yamada T, Koizumi I, Urabe H, Nakamura F. Temperature-dependent swimming performance differs by species: implications for condition-specific competition between stream salmonids. Zoological Science, 2020, 37(5): 429-433. |
| [34] |
李江涛, 林小涛, 周晨辉, 曾鹏, 许忠能, 孙军. 饥饿对食蚊鱼和唐鱼幼鱼能量物质消耗及游泳能力的影响. 应用生态学报, 2016, 27(1): 282-290. |
| [35] |
付成, 肖玲韬, 赵昕雨, 黄恩梅, 徐佳佳, 王亚, 付世建. 不同捕食者暴露对鲫幼鱼行为、运动和形态特征的影响. 生态学报, 2021, 41(11): 4436-4446. |
| [36] |
Imbert H, Labonne J, Rigaud C, Lambert P. Resident and migratory tactics in freshwater European eels are size-dependent. Freshwater Biology, 2010, 55(7): 1483-1493. |
| [37] |
Kopf S M, Humphries P, Watts R J. Ontogeny of critical and prolonged swimming performance for the larvae of six Australian freshwater fish species. Journal of Fish Biology, 2014, 84(6): 1820-1841. |
| [38] |
高泉, 吕为群. 衰老对斑马鱼呼吸代谢和游泳行为的影响. 上海海洋大学学报, 2019, 28(2): 200-210. |
| [39] |
Swanson C, Young P S, Cech J J. Swimming performance of delta smelt: maximum performance, and behavioral and kinematic limitations on swimming at submaximal velocities. Journal of Experimental Biology, 1998, 201(3): 333-345. |
| [40] |
de Andrade e Santos H, de Faria Viana E M, Santos Pompeu P, Martinez C B. Optimal swim speeds by respirometer: an analysis of three neotropical species. Neotropical Ichthyology, 2012, 10(4): 805-811. |
| [41] |
Shi X T, Kynard B, Liu D F, Qiao Y, Chen Q W. Development of fish passage in China. Fisheries, 2015, 40(4): 161-169. |
| [42] |
Peake S, McKinley R S. A re-evaluation of swimming performance in juvenile salmonids relative to downstream migration. Canadian Journal of Fisheries and Aquatic Sciences, 1998, 55(3): 682-687. |