 2024, Vol. 44
2024, Vol. 44文章信息
- 杨红义, 韩瑞
- YANG Hongyi, HAN Rui
- 基于Vortex模型分析金沙江水电开发对圆口铜鱼种群生存力的影响
- Effect of the Jinsha River hydropower development on population viability of Coreius guichenoti based on the Vortex model
- 生态学报. 2024, 44(9): 3984-3998
- Acta Ecologica Sinica. 2024, 44(9): 3984-3998
- http://dx.doi.org/10.20103/j.stxb.202305221076
-
文章历史
- 收稿日期: 2023-05-22
- 网络出版日期: 2024-02-01

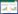




水电开发改变了河流的自然形态[1—2], 影响了河流的水文情势和水温节律[3—4], 阻隔了鱼类洄游通道[5], 缩减了流水生境[4, 6], 使得鱼类资源大幅下降[7], 甚至濒临崩溃[8]。水电开发后, 河流由自然流动状态转变为河湖相交替的形态, 生境破碎化严重[9], 特有鱼类种群数量衰减[10], 物种灭绝风险增加[11], 巴西的Capivara大坝运行后伊氏啮脂鲤(Bryconamericus iheringii)、条纹溪脂鲤(Characidium zebra)等30种当地特有鱼类灭绝[12]; 水电开发导致的水温“滞热”效应缩短了特有鱼类适宜产卵的水温时期, 影响了鱼类繁殖, 如湄公河水电开发引起的水温变化导致光唇裂腹鱼(Schizothorax lissolabiatus)产卵减少、种群数量锐减[13]; 大坝阻隔了鱼类的洄游通道, 如葛洲坝、三峡等工程建设运行后, 中华鲟(Acipencer sinensis)等特有鱼类种群数量持续下降、灭绝概率增加[14]、种群生存力降低[15]; 金沙江水电开发导致流水生境缩减[16], 部分河段无法满足漂流性鱼卵的孵化流速条件[17], 使得圆口铜鱼(Coreius guichenoti)、长鳍吻鮈(Rhinogobio ventralis)等特有鱼类早期资源量补充不足[18—20], 种群数量和优势地位下降[21—22]。
为评估物种种群变化, 种群模型逐步发展[23]。Malthus[24]考虑了单一种群的繁殖能力, 首次提出了种群指数增长模型; Verhurst[25]在此基础上考虑了环境容纳量对种群增长的限制, 提出了Logistic种群增长模型; Lotka[26]考虑了物种的种间竞争, 提出了Lotka-Voltera模型; Lewis[27]考虑了物种不同年龄的繁殖和存活率差异, 建立了矩阵模型, 模拟了年龄分布对种群的影响, 已经成为种群模拟的重要内容[23]。随着物种灭绝的加剧以及学者们对物种生态学的理解深入, 以种群模型为基础的种群生存力分析(PVA, Population Viability Analysis)模型得到了发展[28], 如死亡过程模型、矩阵模型、基于个体的模拟模型、种群随机模型[29—31]等, 其中基于个体模型(IBM, Individual-Based Model)的Vortex模型被广泛采用[32]。传统PVA模型以考虑单随机性为主[33], Vortex模型考虑了环境、遗传、灾害和统计等多种随机性对种群的影响, 并引入了物种特征、人类活动等, 可较好的模拟种群动态对环境变化的响应, 如王伟营等[34]采用Vortex模型模拟了在自然灾害和人类活动影响下, 云南碧塔海地区中甸叶须鱼(Ptychobarbus chungtienensis)种群数量变化, 干旱是影响该物种种群生存力的主要因素。Vortex已经成为分析关键物种种群生存力的重要方法[35]。
圆口铜鱼是长江上游特有鱼类和重要经济鱼类[36], 也是长江上游珍稀特有鱼类国家自然保护区的重要指示性物种[37]和优先保护对象之一[38—39]。近年来, 圆口铜鱼资源量急剧下降, 物种生存面临巨大威胁, 2016年被列入极度濒危等级[40], 其资源保护和恢复极为迫切[41]。目前研究主要围绕圆口铜鱼的繁殖特征[42]、人工驯养[43]、遗传分化[37]、产卵场分布[44]、种群资源量[45—46]及生态行为[47]等方面, 对其种群生存力的研究较少。
本研究基于1981—2021年金沙江圆口铜鱼资源调查数据, 采用Vortex模型构建了金沙江中下游干流大规模水电开发前、后两个时期圆口铜鱼PVA模型, 考虑了水电开发下, 水温、流速等关键因子变化对圆口铜鱼种群的影响, 对比分析了水电开发不同时期以及鱼类繁殖、人类活动等不同情景下圆口铜鱼种群数量及生存力变化规律, 为水电开发下特有鱼类种群资源保护及恢复提供参考。
1 研究区域介绍与数据收集 1.1 研究区域金沙江流域是我国水能开发的“富矿”和“西电东送”的战略要地, 也是我国典型的生态脆弱区, 峡谷急流的特殊地貌决定了其水能利用方式以巨型水电站群基地化开发为主[48]。随着梯级高坝大库的建设和运行, 金沙江许多江段的鱼类资源衰退, 其中, 圆口铜鱼资源严重下降[49]。近年来, 圆口铜鱼性成熟个体只在金沙江中下游有发现[50], 产卵场也主要分布于此[51], 研究该区域水电开发对圆口铜鱼的影响具有重要意义。
金沙江中下游已规划12个梯级水电工程, 部分水电工程建设、运行时间见表 1, 位置分布见图 1。由表 1可知: 21世纪初期, 金沙江尚未开展大规模水电开发, 2010年以后, 半数以上的水电工程已建设并逐步开始运行。基于水电开发进程, 以2010年为数据分析的时间节点, 划分并建立金沙江中下游大规模水电开发前、后圆口铜鱼PVA模型。
| 水电工程 Hydropower projects |
开始建设时间 Construction time |
运行时间 Operation time |
水电工程 Hydropower projects |
开始建设时间 Construction time |
运行时间 Operation time |
|
| 龙盘 | 已规划 | 鲁地拉 | 2009年1月 | 2013年6月 | ||
| 两家人 | 已规划 | 观音岩 | 2011年1月 | 2014年10月 | ||
| 梨园 | 2009年9月 | 2013年11月 | 乌东德 | 2015年12月 | 2020年6月 | |
| 阿海 | 2008年9月 | 2011年11月 | 白鹤滩 | 2013年8月 | 2021年6月 | |
| 皎平渡 | 2005年9月 | 2010年11月 | 溪洛渡 | 2007年4月 | 2014年6月 | |
| 龙开口 | 2009年1月 | 2012年11月 | 向家坝 | 2006年11月 | 2012年7月 |

|
| 图 1 金沙江中下游水电工程分布图 Fig. 1 The distribution map of hydropower projects in the middle and lower reaches of the Jinsha River |
圆口铜鱼隶属鲤形目(Cypriniformes)、鮈亚科(Gobioninae)、铜鱼属(Coreius), 是典型的河道洄游型鱼类, 喜流水生境, 产漂流性卵, 成熟亲鱼上溯洄游到金沙江中下游完成繁殖[36]。近年来, 圆口铜鱼出现种群年龄结构失衡[52]、个体性腺发育不成熟[53]、产卵孵化率降低[54]、资源量[55]及丰度降低[56]等现象, 物种生存面临威胁。圆口铜鱼种群变化与过度捕捞、水电开发等人类活动密切相关, 过度捕捞导致种群数量大幅降低, 出现大龄个体减少, 种群低龄化等现象[53]; 大坝低温水下泄改变了下游的水温节律, 圆口铜鱼产卵临界水温阈值的时间滞后, 对其产卵繁殖产生不利影响[10]; 大坝建设后, 库区江段流速减缓, 可满足圆口铜鱼漂流孵化的江段减少, 产卵孵化率降低和早期资源补充不足导致其种群数量下降[21], 资源丰度降低, 圆口铜鱼资源保护及恢复工作迫在眉睫[54—55]。
1.3 数据收集本研究收集了1981—2021年圆口铜鱼调查结果, 对圆口铜鱼资源量、繁殖特征、产卵场分布等进行了系统分析, 对攀枝花江段圆口铜鱼种群结构、资源动态进行了重点分析, 估算并确定了金沙江水电开发不同时期, 圆口铜鱼种群关键特征参数, 基本满足PVA模型构建的数据需求。基础数据时间涵盖了金沙江水电工程开发前和运行后两个阶段, 空间涵盖了金沙江攀枝花、巧家、皎平渡、水富等圆口铜鱼繁殖活动区域, 基础数据情况见表 2。
| 编号No | 调查时间Survey time | 调查区域Survey areas | 调查内容Survey contents | 数据来源Data sources |
| 1 | 1981—1986年 | 葛洲坝下宜昌江段 | 铜鱼及圆口铜鱼卵苗数量产卵场分布、亲鱼数量 | 刘乐和等[36] |
| 2 | 1983—1984年 | 葛洲坝上武陵江段 | 铜鱼及圆口铜鱼卵苗数量产卵场分布、亲鱼数量 | |
| 3 | 1984年 | 金沙江下游柏溪 | 铜鱼及圆口铜鱼卵苗数量产卵场分布、亲鱼数量 | |
| 4 | 1985—1986年 | 金沙江下游屏山 | 铜鱼及圆口铜鱼卵苗数量产卵场分布、亲鱼数量 | |
| 5 | 2000—2004年 | 长江上游干流万州江段 | 铜鱼及圆口铜鱼种群数量 | 严莉[56] |
| 6 | 2000—2005年每年2—4月 | 长江上游宜宾、巴南和万州江段 | 铜鱼及圆口铜鱼种群数量 | 段辛斌等[53] |
| 7 | 2007—2008年2012—2014年 | 金沙江下游巧家江段 | 圆口铜鱼等产漂流性卵鱼类卵苗数量及产卵场分布 | 肖琼[57] |
| 8 | 2007—2009年每年5—7月、9—11月 | 长江上游江津和宜宾江段 | 圆口铜鱼种群数量及分布 | 熊飞等[58] |
| 9 | 2008年、2011—2013年每年5—7月 | 金沙江下游宜宾江段 | 圆口铜鱼卵苗数量及产卵场分布 | 高少波等[46] |
| 10 | 2013年6、10月 | 金沙江中游攀枝花江段 | 圆口铜鱼种群数量及分布 | 邵科等[20] |
| 11 | 2017—2018年每年5—7月 | 金沙江下游巧家江段 | 圆口铜鱼等产漂流性卵鱼类卵苗数量及产卵场分布 | 周湖海等[44] |
| 12 | 2018—2021年 | 雅砻江金河 | 圆口铜鱼种群数量 | 熊美华等[51] |
| 13 | 2019年 | 金沙江绥江江段 | 圆口铜鱼个体分布 | |
| 14 | 2019年 | 金沙江永善江段 | 圆口铜鱼个体分布 | |
| 15 | 2019年 | 攀枝花雅砻江口 | 圆口铜鱼个体分布 | |
| 16 | 2019—2021年 | 金沙江下游宜宾江段 | 圆口铜鱼个体分布 | |
| 17 | 2019—2020年 | 金沙江下游巧家江段 | 圆口铜鱼个体分布 | |
| 18 | 2019—2021年 | 金沙江下游东川渡口 | 圆口铜鱼个体分布 | |
| 19 | 2019—2020年 | 金沙江下游皎平渡江段 | 圆口铜鱼个体分布 | |
| 20 | 2020—2021年 | 金沙江中游攀枝花江段 | 圆口铜鱼个体分布 |
Vortex模型通过模拟种群中个体的基本特征和行为(如存活、交配、繁殖、死亡等)预测整个种群的动态(如数量、分布和灭绝风险等), 模型中引入了环境容纳量、年龄结构、繁殖体制、出生性比等参数, 并同时考虑了环境、灾害、遗传、统计等多重随机性对种群的影响[30], 已被广泛应用于评估物种的种群生存力。
Vortex模型以年龄矩阵为基础[35], Leslie年龄矩阵如下:

|
(1) |
式中, Fi、Si分别为从第i-1年龄到第i年龄生物个体的繁殖率和存活率, i=1、2、3……n, n为目标生物个体最大存活年龄, 未达到性成熟年龄的个体, 其繁殖率为0%;Si可通过公式(2)计算:
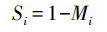
|
(2) |
式中, M为死亡率, 即某一单位时段内个体死亡概率, 与死亡个体数占种群数量的百分比相等, Mi为第i-1年龄到第i年龄生物个体的死亡率[59], 可通过公式(3)计算:
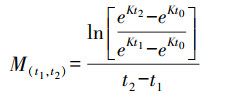
|
(3) |
式中, M(t1, t2)为时间t1到t2阶段生物死亡率, t1、t2为年龄组范围, 如M(0, 1)为0到1龄生物生长过程中的死亡率, 即M1; 当生物达到初次性成熟年龄后, 死亡率不再随年龄增长而变化, 即目标生物达到性成熟后, 死亡率为恒定值; K为该物种的生长系数; t0为该物种的理论生长起点年龄, 可根据von. Bertalanffy生长方程获得:
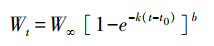
|
(4) |
式中, Wt为预测体长; W∞为极限体长; b为体长体重转化系数; Wt、W∞、b由大量个体数据统计获得。
Vortex模型中计算世代重叠的单物种种群数量Nt的公式如下:
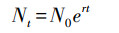
|
(5) |
式中, Nt为种群数量; N0为初始种群数量; t为模拟时间; r为种群内禀增长率, 其计算公式如下:
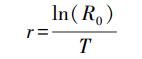
|
(6) |
式中, R0为净增长率, 即种群中个体平均繁殖后代数; T为首次繁殖时间; 计算公式如下:
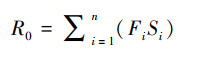
|
(7) |
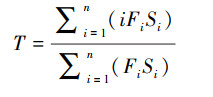
|
(8) |
式中, Fi、Si均通过年龄矩阵获得。
Vortex模型中模拟时间尺度与用于构建模型的原始数据时间长度相关, 原始数据时长应至少为模拟时长的10%—20%[60], 对于单世代时间较短且世代重叠的物种, 模拟时间通常为100a[14, 15, 34, 61—64]。Vortex模型可同时模拟多因素随机性对种群的影响, 模拟过程中, 采用对多次模拟结果进行统计分析的方法, 得出种群数量及生存力, 模拟次数通常为1000次[14, 15, 34]。
2.2 模型参数设定本研究采用Vortex 10.5.5版构建模型, 主要参数设置及依据如表 3所示:
| 参数 Parameters |
水电开发前 Before hydropower development(BD) |
水电开发后 After hydropower development(AD) |
参考依据 Evidence |
| 模拟次数Simulated times/次 | 1000 | 1000 | 模型内设置 |
| 模拟时间Simulated years/年 | 100a | 100a | 模型内设置 |
| 是否近交衰退Inbreeding depression | 无 | 无 | 模型内设置 |
| 灭绝定义Extinction definition | 单性[35] | 单性[35] | Lacy等[35] |
| 雌鱼初次性成熟年龄/龄Age of first offspring for females | 2[65] | 4[42] | 刘成汉等[65], 杨志等[42] |
| 最大繁殖年龄/龄Maximum age of reproduction | 8[37] | 8[37] | 李学梅等[37] |
| 亲鱼参与繁殖比例Parental fish participating in reproduction rate/% | 58[45] | 30[45, 66] | 程鹏[45], 王康等[66] |
| 成功产卵率Percentage of 1 brood/% | 60[67] | 60[67] | 鲁雪报等[67] |
| 出生性比Sex ratio at birth | 1∶1[45, 58] | 1∶1[45, 58] | 程鹏[45], 熊飞等[58] |
| 单次最大产卵数/粒Maximum number of progeny per brood | 1.379×105[42] | 1.379×105[42] | 杨志等[42] |
| 有效子代数/尾Effective subalgebra | 263 | 87 | 计算 |
| 不同年龄组死亡率Mortalities among different ages/% | 见表 4 | 见表 4 | 计算 |
| 灾害Catastrophes | 洪水 | 洪水 | 模型内设置 |
| 灾害发生频率Frequency of occurrence | 百年一遇 | 百年一遇 | 模型内设置 |
| 初始种群数量/尾Initial population quantity | 8万[58] | 5.6万[45] | 程鹏[45], 熊飞等[58] |
| 环境容纳量/尾Carrying capacity (K) | 15万 | 15万 | 计算 |
| 未来环境容纳量是否变化Future change in K | N | N | 模型内设置 |
| 渔业捕捞Fishery | Y | N | 模型内设置 |
| BD: 水电开发前Before hydropower development; AD: 水电开发后After hydropower development; N: 否, no; Y: 是, yes | |||
(1) 繁殖年龄
模型中与繁殖年龄相关的参数包括: 雌鱼初次性成熟年龄、最大繁殖年龄。圆口铜鱼初次性成熟年龄在水电开发前、后存在一定差异: 水电开发前, 雄鱼、雌鱼初次性成熟年龄均为2龄[65]; 水电开发后, 雄鱼、雌鱼初次性成熟年龄分别为4.44—7龄和4.32—6龄[42, 68]。基于多年野外调查渔获物中圆口铜鱼年龄结构分析, 其个体最大繁殖年龄为8龄[69]。
(2) 亲鱼参与繁殖比例
水电开发前, 圆口铜鱼为金沙江重要经济鱼类, 对其亲鱼参与繁殖比例的研究较少。水电开发对亲鱼参与繁殖产生了一定影响[51], 其中水温是亲鱼参与繁殖的关键影响因素[9], 因此, 本研究依据水电开发后的研究成果及水电开发造成产卵期水温变化的程度初步估算水电开发前亲鱼参与繁殖情况。
水电开发后, 金沙江攀枝花江段渔业捕捞圆口铜鱼亲鱼样本统计结果表明, 圆口铜鱼亲鱼参与繁殖比例为30%[42]。受水电开发对河道天然水温的影响, 适宜圆口铜鱼产卵的水温窗口期缩短为水电开发前的51%[66], 根据水电开发后亲鱼参与繁殖比例与产卵期缩短情况, 估算得出水电开发前亲鱼参与繁殖比例为58%。
(3) 有效子代数
有效子代数指雌鱼每次繁殖可存活的后代个数。圆口铜鱼的有效子代数受平均产卵量、受精率[69]、孵化率及24h内短期存活率[70]的综合影响, 依据调查及实验结果(平均产卵量20000粒、受精率70%、孵化率20.9%、短期存活率9%), 确定水电开发前圆口铜鱼平均有效子代数为263尾。金沙江中下游水电开发使得满足鱼卵孵化水流条件的江段仅占原来的33%[9], 依此估算水电开发后圆口铜鱼平均有效子代数为87尾。有效子代数计算公式如下:
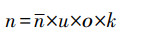
|
(9) |
式中, n为有效子代数; 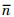
圆口铜鱼属匀速生长类型[68], 死亡率依据公式(3)计算获得, 根据长江上游渔获物数据拟合公式(4), 计算获得圆口铜鱼生长系数K和理论生长起点年龄t0分别为0.1096和-0.7788[46]。模型中, 水电开发前, 考虑过度捕捞影响, 对公式(3)获得的死亡率计算结果按照公式(10)进行修正:
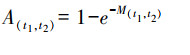
|
(10) |
式中, A为未考虑捕捞影响的修正死亡率[14]。
在修正死亡率基础上, 依据1998—2007年圆口铜鱼捕捞数据[68]估算过度捕捞死亡率进行叠加, 设定1—2龄捕捞死亡率为30%, 2龄以后各年龄段捕捞死亡率为10%。水电开发后, 为保护鱼类资源, 2017—2019年实施春季禁渔政策, 2020年后金沙江干流和主要支流实施十年全域禁捕政策, 未设置捕捞影响, 如表 4所示:
| 龄级 Age-class |
水电开发前死亡率/% Mortality of BD |
水电开发后死亡率/% Mortality of AD |
龄级 Age-class |
水电开发前死亡率/% Mortality of BD |
水电开发后死亡率/% Mortality of AD |
|
| 0—1 | 58.61 | 88.20 | 3—4 | 23.27 | 29.39 | |
| 1—2 | 69.54 | 50.32 | >4 | 23.27 | 21.35 | |
| 2—3 | 23.27 | 36.55 |
(1) 初始种群数量
水电开发前, 圆口铜鱼调查结果表明: 金沙江攀枝花至长江干流万州江段圆口铜鱼资源量约为160万尾[58, 71], 攀枝花江段约占总调查江段的5%, 故估算攀枝花江段圆口铜鱼种群数量约为8万尾; 水电开发后, 攀枝花江段渔业调查结果表明圆口铜鱼种群数量约为5.6万尾[20]。
(2) 环境容纳量
环境容纳量指特定环境可容纳目标物种种群数量的最大值, 获得其准确值较为困难[72], 通常可根据食物需求量和有效栖息地估算。本研究基于调查区域面积、初级生产力、目标物种食性等, 估算研究区域总体环境容纳量, 估算过程如下:
渔业调查流域面积约6.8×106m2[45], 以内陆淡水渔业的基本情况作为参考, 按照每平米可产生43.66g藻类, 杂食性鱼类每生长1g需消耗约40g藻类[73], 圆口铜鱼的平均体重约为50g[50, 74], 估算研究区域环境容纳量为15万尾。
2.2.4 自然灾害模型中自然灾害及其对鱼类种群影响程度参考同区域其它特有鱼类研究设置: 自然灾害按百年一遇洪水考虑, 灾害发生时圆口铜鱼存活率、繁殖率均为原参数的99%。
2.3 模型可靠性、误差及误差来源分析Vortex模型以岛屿生物地理学为理论起源, 以年龄矩阵模型为核心, 将描述个体迁移的IBM方法引入模型[75], 经多次模拟得出种群在多种确定性因素和随机性影响综合作用下的结果, 其模型结果比确定性的种群模型更为准确[35]。Brook等[76]采用大量种群生态学实测数据检验和证明了Vortex模型的准确性和预测能力, 为Vortex的广泛应用提供了支持。
Vortex模型模拟物种的生命过程, 模拟结果的准确性依赖于对目标物种生态学特征的理解以及所掌握数据的情况[77]。模拟误差主要来源包括: 数据质量、参数选择与设置等。本研究中, 圆口铜鱼是金沙江水电绿色开发的重要指示性物种, 其生态学特征研究基础较好; 野外观测技术的发展为Vortex模型获得高质量数据提供了保障, 本研究基础数据以野外调查结果为主, 数据覆盖了圆口铜鱼完整生活史; 原始数据参数化前, 对数据进行了分析、校正, 避免由于采样误差等造成的模拟误差, 尽量确保参数选择的合理性以及设置依据的充分性; 检验模型时, 将不同时期模拟结果与调查结果分别进行对比, 重点分析种群相对变化趋势的协调性, 为进一步提高模拟准确性提供依据。
2.4 模型模拟情景设定本研究构建了水电开发前、后两个时期圆口铜鱼PVA模型, 为明晰水电开发对圆口铜鱼种群生存力影响提供依据。为进一步识别水电开发后, 对圆口铜鱼种群生存力影响显著的参数, 本研究基于水电开发后的模型, 通过调整主要参数的取值, 即适当增加或减少原参数值, 设定了多个独立情景进行模拟, 调整参数包括: 幼鱼死亡率、亲鱼参与繁殖比例、有效子代数、灾害频率及单次最大产卵数等, 并对其中多个情景进行组合模拟, 为关键参数的甄别提供依据。同时, 结合金沙江圆口铜鱼增殖放流以补充幼鱼为主的特点[78—79], 模拟了持续补充幼鱼数量等情景, 为评估目前资源保护措施对种群生存力的影响提供依据。
3 结果 3.1 模拟结果与调查结果对比不同时期圆口铜鱼资源调查结果表明: 水电开发前, 研究区域2003—2007年圆口铜鱼资源量分别约为8—9.6万尾、7.9—9.5万尾、7.8—9.35万尾、7.25—8.7万尾、5.95—7.1万尾[71]; 水电开发后, 2013年、2017年圆口铜鱼资源量分别约为5.6万尾、4.4万尾[20]。水电开发前、后两个时期圆口铜鱼种群数量及其随时间变化趋势模拟结果如图 2所示, 与调查情况的对比结果表明: 水电开发前, 模拟结果2003—2007年圆口铜鱼资源量分别为9.44万尾、6.48万尾、8.23万尾、6.88万尾、7.8万尾; 水电开发前模型模拟结果受渔业捕捞影响, 资源量出现一定波动, 总体与调查资源量范围较为相近, 其种群数量变化趋势也较为一致; 水电开发后, 模拟结果2013年、2017年圆口铜鱼资源量分别5.6万尾、4.42万尾, 与调查结果种群数量相近, 种群下降趋势也较为一致。综上, 水电开发前、后的模拟结果基本可反映不同时期圆口铜鱼种群数量及变化趋势, 可作为其种群生存力分析的基础。

|
| 图 2 不同时期模拟结果与调查结果对比 Fig. 2 The comparison of simulation results and survey results in different periods |
| 时期 Period |
净增长率 Net growth rate |
首次繁殖时间/year First breeding time |
种群内禀增长率 Population growth rate |
灭绝概率/% Probability extinction |
种群数量 Population quantity |
平均灭绝时间/year Mean extinction time |
| 水电开发前(BD) | 12.8 | 3.34 | 0.085 | 1 | 48680.76 | — |
| 水电开发后(AD) | 0.60 | 5.60 | -0.087 | 36.1 | 198 | 83.7 |
| “—”表示此情景下无平均灭绝时间 | ||||||

|
| 图 3 不同时期圆口铜鱼种群数量变化趋势图 Fig. 3 Variation trend of C. guichenoti population in different periods BD: 水电开发前, before hydropower development; AD: 水电开发后, after hydropower development |
水电开发前, 圆口铜鱼受过度捕捞影响显著, 约10年内种群数量出现震荡下降, 该时间段种群内禀增长率为0.038;随后种群数量逐渐趋于稳定波动状态, 百年平均种群数量约为4.8万尾, 种群内禀增长率为0.085, 首次繁殖时间为3.34年, 平均灭绝概率为1%。水电开发后, 圆口铜鱼种群数量持续下降, 种群生存力明显减弱; 百年内, 每20年种群下降数量依次为1970尾/a、570尾/a、180尾/a、60尾/a、20尾/a; 种群数量下降的幅度逐渐减小, 依次为70.3%、20.4%、6.3%、2%、0.6%;百年平均种群数量为198尾, 种群内禀增长降低至-0.087, 首次繁殖时间延后至5.60a, 出现灭绝危机, 平均灭绝概率为36.1%, 平均灭绝时间为83.7年。
3.3 圆口铜鱼最小可存活种群数量估算物种最小可存活种群数量(MVP, Minimum Viable Population)指物种种群在当前环境下百年内灭绝概率小于5%时的最小种群数量[64], 对其资源保护具有重要的指导意义[77]。以水电开发后模型为基础, 通过增加圆口铜鱼初始种群数量至环境容纳量的方法估算MVP。
结果表明: 水电开发后, 圆口铜鱼种群内禀增长率随初始种群数量增加变化较小, 平均值-0.085±0.008, 百年内平均种群数量有一定上升, 灭绝概率总体呈下降趋势(图 4), 当初始种群数量达到环境容纳量时, 灭绝概率为27%;水电开发后, 该物种的繁殖及生存环境已受到显著影响, 初始种群数量的增加对种群内禀增长率几乎无影响, 对灭绝概率的影响较小, 目前环境下该物种不存在MVP, 即该物种已无法满足百年内种群灭绝概率小于5%的存活数量要求, 亟需采取相关措施提升其种群生存力, 保护物种延续。

|
| 图 4 圆口铜鱼种群灭绝概率随初始种群数量关系 Fig. 4 The relationship between C. guichenoti initial population quantity and probability of population extinction |
物种种群生存力主要受其生态学特征和环境变化的共同影响, 为识别影响圆口铜鱼种群生存力的关键参数, 本研究基于水电开发对物种和环境的影响特点, 通过调整水电开发后Vortex模型中相应参数取值, 设定了多种模拟情景, 如幼鱼死亡率(情景a、b)、亲鱼参与繁殖比例(情景c、d)、有效子代数(情景e, f)、灾害频率(情景g, h)、单次最大产卵数(情景i, j)、持续补充幼鱼数量(情景k)等多个参数取值变化, 及其中多个情景(情景c+e, d+f)的组合, 参数设置及情景模拟结果见图 5、表 6。

|
| 图 5 不同情景下圆口铜鱼种群数量变化 Fig. 5 Changes of population quantity of C.guichenoti in different scenarios a.b.c.d.等是按照字母顺序表示不同情景, c+e则表示在参数变化上同时发生c和e的变化 |
| 情景 Scenarios |
参数设置 Parameter setting |
净增长率 Net growth rate |
首次繁殖时间/a First breeding time |
种群内禀增长率 Population growth rate |
种群内禀增长率的变化率/% Rate of change r |
灭绝概率/% Probability extinction |
种群数量 Population quantity |
平均灭绝时间/a Mean extinction time |
| 基础Basic | 0.67 | 5.60 | -0.086 | 0 | 36.9 | 131 | 82.6 | |
| a. | 105%幼鱼死亡率* | 0.42 | 5.68 | -0.182 | -110 | 99.9 | 0 | 50.5 |
| b. | 95%幼鱼死亡率 | 0.93 | 5.54 | -0.019 | 119 | 0.0 | 19145 | — |
| c. | 110%亲鱼参与繁殖比例 | 0.74 | 5.58 | -0.068 | 16 | 17.6 | 763 | 87.4 |
| d. | 90%亲鱼参与繁殖比例 | 0.61 | 5.62 | -0.104 | -23 | 62.9 | 25 | 80.8 |
| e. | 110%有效子代数 | 0.74 | 5.58 | -0.068 | 16 | 17.7 | 797 | 85.5 |
| f. | 90%有效子代数 | 0.61 | 5.62 | -0.105 | -25 | 64.5 | 19 | 80.7 |
| g. | 十年一遇灾害频率 | 0.67 | 5.60 | -0.084 | 0 | 36.4 | 167 | 84.2 |
| h. | 五年一遇灾害频率 | 0.67 | 5.60 | -0.085 | 0 | 36.7 | 146 | 84.0 |
| i. | 110%单次最大产卵数 | 0.67 | 5.60 | -0.085 | 0 | 36.3 | 151 | 82.9 |
| j. | 90%单次最大产卵数 | 0.67 | 5.60 | -0.085 | 0 | 37.1 | 109 | 85.0 |
| k. | 每年补充1万尾幼鱼 | 0.67 | 5.60 | -0.0193 | 77.5 | 0 | 9334 | — |
| c+e. | 0.81 | 5.57 | -0.051 | 41 | 0.5 | 2643 | 84.8 | |
| d+f. | 0.55 | 5.63 | -0.128 | -48 | 86.5 | 26 | 76.7 | |
| a.b.c.d.等是按照字母顺序表示不同情景, c+e则表示在参数变化上同时发生c和e的变化, “*”表示在情景“基础”相应参数的基础上增加或减少一定比例, 如105%幼鱼死亡率=基础幼鱼死亡率×(100%+5%); “—”表示此情景下无平均灭绝时间 | ||||||||
模拟结果表明: 降低幼鱼死亡率、持续补充幼鱼数量(即增殖放流)均可明显提高圆口铜鱼种群生存力, 该两项参数的变化可直接改变圆口铜鱼种群数量变化趋势; 亲鱼参与繁殖比例和有效子代数可在一定程度上影响圆口铜鱼的种群生存力, 该两项参数与种群内禀增长率呈正相关关系, 若考虑同时提高亲鱼参与繁殖比例和有效子代数, 其叠加影响可将圆口铜鱼灭绝概率由36.9%降低至0.5%, 对种群生存力的提高显著。灾害频率和单次最大产卵量的变化对圆口铜鱼种群数量及生存力无明显影响。因此, 从降低圆口铜鱼幼鱼死亡率、持续补充幼鱼数量、提高亲鱼参与繁殖比例及有效子代数等角度开展资源保护措施研究, 对提高圆口铜鱼种群生存力具有积极意义。
3.5 提高圆口铜鱼种群生存力模拟结果本研究在水电开发后模型基础上, 模拟了降低幼鱼死亡率、持续补充幼鱼数量、提高亲鱼参与繁殖比例和有效子代数等单参数变化情况下的情景模拟, 结果如图 6至图 9所示。由图可知, 圆口铜鱼种群数量无下降趋势、种群生存力得以保障的阈值分别为: 幼鱼死亡率降低至80%;持续补充幼鱼数量增加至6万尾以上; 亲鱼参与繁殖比例提高约83%, 即参数增加至55%;有效子代数提高84%, 即参数增加至160尾。上述阈值情况下, 圆口铜鱼的种群生存力均可得到显著提高, 种群内禀增长率依次为: 0.0200、0.0012、0.0216、0.0252, 种群灭绝概率均小于0.1%, 种群不存在灭绝风险。该模拟结果可为初步评估资源保护措施对圆口铜鱼种群生存力影响提供依据。

|
| 图 6 不同幼鱼死亡率下圆口铜鱼种群数量变化 Fig. 6 Population quantity of C. guichenoti under different juvenile fish mortalities |

|
| 图 7 不同持续补充幼鱼数量下圆口铜鱼种群数量变化 Fig. 7 Population quantity of C. guichenoti under different larve releasing quantities |

|
| 图 8 不同亲鱼参与繁殖比例下圆口铜鱼种群数量 Fig. 8 Population quantity of C. guichenoti with different parental fish participating in reproduction rate |

|
| 图 9 不同有效子代数下圆口铜鱼种群数量 Fig. 9 Population quantity of C. guichenoti with different effective subalgebra |
水电开发后, 圆口铜鱼幼鱼死亡率、亲鱼参与繁殖比例、有效子代数等对其种群生存力均存在一定影响。其中: 幼鱼死亡率对种群的影响程度显著, 这与秀丽高原鳅的模拟结论一致[64], 幼鱼通常体质相对较弱、环境适应性较差, 较易受到流水生境面积、初级生产力单位密度等影响[52]; 亲鱼参与繁殖比例也可影响圆口铜鱼的种群生存力, 该参数通常受水温节律改变影响, 坝下水温发生了改变, 部分江段因无法达到圆口铜鱼的产卵水温要求, 导致亲鱼无法产卵[9]; 有效子代数是决定圆口铜鱼早期资源补充数量的重要影响因子, 水电开发后水流流速降低, 可满足圆口铜鱼卵孵化的江段减少, 导致鱼卵孵化率降低[17]。
水电开发引起的大坝阻隔和流量脉冲变化对圆口铜鱼种群存在一定影响[21]。大坝阻隔降低上溯亲鱼洄游, 进一步降低亲鱼参与繁殖的比例, 如在乌东德坝下东川渡口观测到圆口铜鱼亲鱼, 但未观察到产卵现象[51], 同时, 大坝阻隔也影响了幼鱼下行, 对幼鱼死亡率、有效子代数均有影响; 流量脉冲次数降级、强度减弱, 对亲鱼产卵存在不利影响, 如在金沙江一期工程运行后, 圆口铜鱼的产卵高峰次数减少, 高峰产卵量降低, 直接降低了早期资源量[46]。本研究中暂未考虑上述变化对圆口铜鱼的种群生存力的负面影响, 目前模拟结果偏于保守。
4.2 圆口铜鱼资源保护措施初步分析 4.2.1 增殖放流目前, 在长江上游保护区和金沙江下游等江段的圆口铜鱼放流任务为70万尾[78], 其中溪洛渡、向家坝水电站为50万尾, 乌东德、白鹤滩水电站为20万尾[79]。圆口铜鱼增殖放流规模均超过了本研究中增殖放流(即持续补充幼鱼数量)的临界阈值, 该项措施的持续开展可以在一定程度上保护资源, 但本研究主要考虑了圆口铜鱼繁殖生理参数的变化, 以及水电开发引起的水温、流速变化对PVA模型参数的影响, 提出增殖放流临界阈值可能偏保守, 同时, 由于对增殖放流实际效果的评估报道较少[43], 这也影响了对增殖放流规模的研究和确定。
4.2.2 增设过鱼设施圆口铜鱼调查结果表明, 其产卵场主要集中在金沙江中游、下游[44], 金沙江梯级水电开发对圆口铜鱼亲鱼上溯已经产生了阻隔效应, 在水电站处增设有效的圆口铜鱼过鱼设施, 将坝下群体转运到坝上流水江段, 将提高其亲鱼参与繁殖的比例。目前, 金沙江下游的四级电站中, 仅乌东德水电站建成了集运鱼系统, 以圆口铜鱼为优先过鱼对象, 在一定程度上可能帮助部分鱼类完成上溯洄游[80], 提高圆口铜鱼亲鱼参与繁殖比例[81—82], 可结合本研究中亲鱼参与繁殖比例的临界阈值, 对过鱼效果进行持续监测及评估。
4.3 圆口铜鱼与其它特有鱼类种群生存力比较本研究将圆口铜鱼与其它特有鱼类的Vortex模型建立过程及估算结果进行了对比(表 7), 对圆口铜鱼种群生存力进行进一步讨论。
| 物种 Species |
有效子代数 Effective subalgebra |
幼龄个体死亡率/% 0—1age mortality rate |
种群内禀增长率 Population growth rate |
灭绝概率/% Probability extinction |
平均灭绝时间/a Mean extinction time |
| 长江江豚[61] Neophocaena asiaeorientalis asiaeorientalis |
1 | 15.0—30.0 | -0.073—-0.020 | 40—80 | 24—94 |
| 长江江豚[63] Neophocaena asiaeorientalis asiaeorientalis |
1 | 30.0—60.0 | -0.622—-0.023 | 35—91 | 68.4—91.5 |
| 白鱀豚[62] Lipotes vexillifer |
1 | 8.3—18 | -0.07 | 100 | 54 |
| 中华鲟[14] Acipencer sinensis |
16 | 80.0 | -0.264—-0.12 | 100 | 40—80 |
| 中甸叶须鱼[34] Ptychobarbus chungtienensis |
2000 | 80.0 | -0.094 | 77.1 | 43.8 |
| 秀丽高原鳅[64] Triplophysa venusta |
500 | 93.6 | -0.057 | 54.8 | 61.4 |
| 水电开发前圆口铜鱼 C. guichenoti BD |
263 | 58.6 | 0.085 | 1 | — |
| 水电开发后圆口铜鱼 C. guichenoti AD |
87 | 88.2 | -0.087 | 36.1 | 83.7 |
| “—”表示此情景下无平均灭绝时间 | |||||
长江江豚、白鱀豚、中华鲟、中甸叶须鱼、秀丽高原鳅等模型构建过程中, 基础数据来源与分布较为复杂; 参数设置中, 多项参数根据研究经验直接设置, 如死亡率基于年龄段统一赋值; 有效子代数基于受精率、孵化率设置。本研究中圆口铜鱼PVA模型构建: 基础数据主要来源于研究区域野外调查结果, 并根据圆口铜鱼生态学特征进行分析、筛选; 参数设置以基础数据、开发影响机制与公式估算相结合的方法为主, 如死亡率的设置考虑了自然死亡率及过度捕捞的影响等, 并基于野外调查数据中渔获物年龄结果进行计算检验, 并考虑了初次性成熟年龄延迟对死亡率的影响, 有效子代数的设置补充考虑了短期存活率的影响; 模拟过程中, 将本模型模拟结果与调查结果进行了初步对比, 并分析了不同时期产生种群数量趋势变化的原因, 为模型结果的相对准确提供了依据。
由表 7可知: 特有鱼类种群内禀增长率范围为-0.622—-0.02, 灭绝概率范围为24%—100%, 均存在不同程度的灭绝风险。导致上述各物种的生存力威胁的诱因不同, 长江江豚的种群数量较少, 繁殖能力较低, 有效子代数为1, 在生长过程中可能存在性比失衡的现象, 在饵料减少、生境破坏、性比失衡等多种因素的共同影响下长江江豚的灭绝概率较高; 白鱀豚的初始种群数量较少, 生境逐渐恶化, 死亡率提高, 在自然环境下将逐渐灭绝; 中华鲟由于初次性成熟较晚, 且环境容纳量随水电开发进程逐步减少, 直接制约了中华鲟的繁殖密度, 导致其灭绝概率较高; 中甸叶须鱼产卵量和繁殖参数均有利于其种群生存, 但其受栖息地高温、干旱等自然灾害和肤霉病、鳃霉病、烂鳃病等疾病影响, 导致种群生存力受到威胁, 出现灭绝风险; 秀丽高原鳅的种群生存力受死亡率和灾害程度制约, 其中灾害是导致其种群灭绝的关键因子; 圆口铜鱼和其他特有物种相比, 其初始种群数量较高, 有效子代数较多[52], 初次性成熟年龄早于长江江豚、中华鲟等, 属于种群恢复能力较强的物种[39], 主要栖息于金沙江中下游江段, 灾害对其影响程度相对较小, 因此, 圆口铜鱼与其它特有鱼类相比, 其种群生存力相对较高, 物种灭绝概率相对偏低。
4.4 圆口铜鱼PVA模型的准确性分析圆口铜鱼是金沙江的指示性物种[37], 较为丰富的野外调查和生态学研究积累为本研究的顺利开展奠定了基础。圆口铜鱼PVA模型构建中, 基础数据的分析和选取遵守采样方法与数据处理方法相同的原则, 当采样江段一致时, 对比不同采样时间下, 数据变化分布情况, 并与水电开发时间进行对照, 分析数据波动原因, 剔除异常数据, 尽量避免采样随机误差、保障数据质量。参数设置过程中, 结合圆口铜鱼生态学特征分析, 补充考虑了死亡率、短期存活率等因素, 进一步提高了模拟的准确性。Vortex模型的验证一直是研究领域的难点[64, 72], 多数研究忽略了模型验证, 以不同参数设置下的情景模拟结果对比为主, 本研究尝试性将模拟结果与调查结果进行初步对比, 在一定程度为模拟结果的准确性提供了基础, 但距严谨的模型验证还存在差距。此外, 由于缺少调查数据等客观原因, 水电开发前、后部分参数采用设置统一值的方法, 且本模型暂未考虑疾病、洄游通道阻隔等因素对圆口铜鱼种群生存力的影响, 均会对模拟结果的准确性存在影响, 可结合生境模型等耦合研发种群生存力模型, 更好的反映水电开发导致的环境变化对特有物种的影响。
5 结论本研究在圆口铜鱼长期研究成果及相关资源调查的基础上, 结合金沙江流域水电开发进程, 建立了水电开发前、后圆口铜鱼PVA模型, 主要考虑了水电开发下, 水温、流速变化对圆口铜鱼生态学特征的影响, 对比分析了上述模型中圆口铜鱼种群生存力变化响应规律。主要结论如下:
(1) 水电开发前, 圆口铜鱼受过度捕捞影响, 种群数量呈现震荡下降趋势, 随后种群数量逐渐趋于稳定波动状态; 种群内禀增长率为0.085, 百年内灭绝概率为1%;水电开发后, 圆口铜鱼种群数量持续下降, 种群内禀增长率为-0.087, 百年内灭绝概率为36.1%, 平均灭绝时间为83.7a。
(2) 水电开发后, 圆口铜鱼种群生存力受物种内在生理生态因素和外在环境因素变化的综合影响, 已无法满足百年内种群灭绝概率小于5%的最小可存活种群数量; 可通过增殖放流、增设过鱼设施等生态恢复措施, 从降低幼鱼死亡率、持续补充幼鱼数量、提高亲鱼参与繁殖比例及有效子代数等角度提高其种群生存力, 保护物种延续。
| [1] |
Bice C M, Gehrig S L, Zampatti B P, Nicol J M, Wilson P, Leigh S L, Marsland K. Flow-induced alterations to fish assemblages, habitat and fish-habitat associations in a regulated lowland river. Hydrobiologia, 2014, 722(1): 205-222. DOI:10.1007/s10750-013-1701-8 |
| [2] |
Anderson E P, Jenkins C N, Heilpern S, Maldonado-Ocampo J A, Carvajal-Vallejos F M, Encalada A C, Rivadeneira J F, Hidalgo M, Cañas C M, Ortega H, Salcedo N, Maldonado M, Tedesco P A. Fragmentation of Andes-to-amazon connectivity by hydropower dams. Science Advances, 2018, 4(1): eaao1642. DOI:10.1126/sciadv.aao1642 |
| [3] |
邓云, 李嘉, 李然, 李克锋. 水库调度对溪洛渡电站下游水温的影响. 四川大学学报: 工程科学版, 2006, 38(5): 65-69. DOI:10.3969/j.issn.1009-3087.2006.05.010 |
| [4] |
Cheng F, Li W, Castello L, Murphy B R, Xie S G. Potential effects of dam cascade on fish: lessons from the Yangtze River. Reviews in Fish Biology and Fisheries, 2015, 25(3): 569-585. DOI:10.1007/s11160-015-9395-9 |
| [5] |
曹文宣. 长江鱼类资源的现状与保护对策. 江西水产科技, 2011(2): 1-4. DOI:10.3969/j.issn.1006-3188.2011.02.001 |
| [6] |
Timpe K, Kaplan D. The changing hydrology of a dammed Amazon. Science Advances, 2017, 3(11): e1700611-e1700611. DOI:10.1126/sciadv.1700611 |
| [7] |
高少波, 唐会元, 乔晔, 杨志, 陈金生. 金沙江下游干流鱼类资源现状研究. 水生态学杂志, 2013, 34(1): 44-49. |
| [8] |
农业部长江流域渔业资源管理委员会, 世界自然基金会北京代表处, 2013年长江上游联合科考报告. 2013.
|
| [9] |
李婷, 唐磊, 王丽, 安磊, 王骏, 莫康乐, 陈求稳. 水电开发对鱼类种群分布及生态类型变化的影响——以溪洛渡至向家坝河段为例. 生态学报, 2020, 40(4): 1473-1485. |
| [10] |
Li T, Chen Q W, Zhang Q, Feng T, Zhang J Y, Lin Y Q, Yang P S, He S F, Zhang H. Transcriptomic analysis on the effects of altered water temperature regime on the fish ovariandevelopment of Coreius guichenoti under the impact of river damming. Biology, 2022, 11(12): 1829. DOI:10.3390/biology11121829 |
| [11] |
Jager H I, Chandler J A, Lepla K B, Van Winkle W. A theoretical study of river fragmentation by dams and its effects on white sturgeon populations. Environmental Biology of Fishes, 2001, 60(4): 347-361. DOI:10.1023/A:1011036127663 |
| [12] |
Orsi M L, Britton J R. Long-term changes in the fish assemblage of a neotropical hydroelectric reservoir. Journal of Fish Biology, 2014, 84(6): 1964-70. DOI:10.1111/jfb.12392 |
| [13] |
Ding C, Sun J, Huang M R, Bond N, Ding L Y, Tao J. Flow and thermal regimes altered by a dam caused failure of fish recruitment in the upper Mekong River. Freshwater Biology, 2023, 68(8): 1319-1329. DOI:10.1111/fwb.14105 |
| [14] |
Gao X, Brosse S, Chen Y B, Lek S, Chang J B. Effects of damming on population sustainability of Chinese sturgeon, Acipenser sinensis: evaluation of optimal conservation measures. Environmental Biology of Fishes, 2009, 86(2): 325-336. DOI:10.1007/s10641-009-9521-4 |
| [15] |
Jiao W J, Zhang P, Chang J B, Tao J P, Liao X L, Zhu B. Variation in the suitability of Chinese sturgeon spawning habitat after construction of dams on the Yangtze River. Journal of Applied Ichthyology, 2019, 35(3): 637-643. DOI:10.1111/jai.13914 |
| [16] |
陈大庆, 常剑波, 顾洪宾. 金沙江一期工程对保护区生态环境的影响与对策. 长江科学院院报, 2005, 22(2): 21-24. DOI:10.3969/j.issn.1001-5485.2005.02.006 |
| [17] |
曹文宣. 长江上游水电梯级开发的水域生态保护问题. 长江技术经济, 2017, 1(1): 25-30. |
| [18] |
王海秀, 尹正杰. 长江上游水文情势变化对保护区铜鱼产卵的影响. 人民长江, 2019, 50(12): 46-50, 79. |
| [19] |
赵承远, 王海龙, 张德选, 邓育林, 邱承皓, 郑欢, 常娟, 杨标. 龙开口水电站坝上坝下鱼类群落结构变化趋势. 云南水力发电, 2022, 38(1): 224-229. DOI:10.3969/j.issn.1006-3951.2022.01.050 |
| [20] |
邵科, 杨志, 唐会元, 阙延福, 李伟涛, 熊美华. 观音岩水电站蓄水前后金沙江攀枝花江段鱼类群落结构及变化特征. 长江流域资源与环境, 2020, 29(11): 2417-2426. |
| [21] |
胡兴坤, 阙延福, 李伟涛, 朱滨, 余丹, 陈志刚, 邵科. 金沙江二期工程蓄水前下游皎平渡江段产漂流性卵鱼类产卵场现状. 长江流域资源与环境, 2020, 29(10): 2200-2208. |
| [22] |
吴强, 段辛斌, 徐树英, 熊传喜, 陈大庆. 长江三峡库区蓄水后鱼类资源现状. 淡水渔业, 2007, 37(2): 70-75. |
| [23] |
何文珊, 陆健健, 张修峰, 译. 生态模型基础. 北京: 高等教育出版社, 2008.
|
| [24] |
英 马尔萨斯Malthus, Thomas Robert, 王惠惠. 人口原理. 西安: 陕西师范大学出版社, 2008.
|
| [25] |
Miner J R. Pierre-françois verhulst, the discoverer of the logistic curve. Human Biology, 1933, 5(4): 673-689. |
| [26] |
Fisher R A. The wave of advance of advantageous genes. Annals of Eugenics, 1937, 7(4): 355-369. DOI:10.1111/j.1469-1809.1937.tb02153.x |
| [27] |
Leslie P H. On the use of matrices in certain population mathematics. Biometrika, 1945, 33(3): 183-212. DOI:10.1093/biomet/33.3.183 |
| [28] |
Beissinger S R. Population viability analysis. Chicago: University of Chicago Press, 2002.
|
| [29] |
Richter-Dyn N, Goel N S. On the extinction of a colonizing species. Theoretical Population Biology, 1972, 3(4): 406-433. DOI:10.1016/0040-5809(72)90014-7 |
| [30] |
Akçakaya H R, Sjögren-Gulve P. Population viability analyses in conservation planning: an overview. Ecological Bulletins, 2000(48): 9-21. |
| [31] |
esit Akçakaya H R. Linking population-level risk assessment with landscape and habitat models. Science of the Total Environment, 2001, 274(1/2/3): 283-291. |
| [32] |
田瑜, 邬建国, 寇晓军, 王天明, Andrew T.SMITH, 葛剑平. 种群生存力分析(PVA)的方法与应用. 应用生态学报, 2011, 22(1): 257-267. |
| [33] |
Boyce M S. Population viability analysis. Annual Review of Ecology and Systematics, 1992, 23: 481-497. DOI:10.1146/annurev.es.23.110192.002405 |
| [34] |
王伟营, 赵婷怡, 蒋万胜, 陈小勇. 中甸叶须鱼种群生存力初步分析. 水生态学杂志, 2012, 33(5): 47-53. |
| [35] |
Lacy R C, Pollak J P. Vortex: a stochastic simulation of the extinction process. Version 10.5.5. Brookfield: Chicago Zoological Society, 2021.
|
| [36] |
刘乐和, 吴国犀, 王志玲. 葛洲坝水利枢纽兴建后长江干流铜鱼和圆口铜鱼的繁殖生态. 水生生物学报, 1990, 14(3): 205-215. |
| [37] |
李学梅, 吴兴兵, 杨德国, 朱永久. 基于转录组测序的圆口铜鱼微卫星标记筛选. 水生态学杂志, 2021, 42(4): 97-103. |
| [38] |
危起伟, 王剑伟, 陈大庆, 郑海涛, 龚廷登, 何强, 李翀, 李昀, 廖文根, 刘绍平, 倪朝辉, 王冲, 杨德国, 赵依民. 长江上游珍稀特有鱼类国家级自然保护区科学考察报告. 北京: 科学出版社, 2012.
|
| [39] |
徐薇, 杨志, 乔晔. 长江上游河流开发受威胁鱼类优先保护等级评估. 人民长江, 2013, 44(10): 109-112. |
| [40] |
蒋志刚, 江建平, 王跃招, 张鹗, 张雁云, 李立立, 谢锋, 蔡波, 曹亮, 郑光美, 董路, 张正旺, 丁平, 罗振华, 丁长青, 马志军, 汤宋华, 曹文宣, 李春旺, 胡慧建, 马勇, 吴毅, 王应祥, 周开亚, 刘少英, 陈跃英, 李家堂, 冯祚建, 王燕, 王斌, 李成, 宋雪琳, 蔡蕾, 臧春鑫, 曾岩, 孟智斌, 方红霞, 平晓鸽. 中国脊椎动物红色名录. 生物多样性, 2016, 24(5): 501-551, 615. |
| [41] |
陈桂来, 张薪. 圆口铜鱼的生物学特征及疾病防治技术. 吉林农业, 2011(4): 289. |
| [42] |
杨志, 龚云, 朱迪, 潘磊, 董纯, 刘宏高, 陈小娟, 唐会元. 金沙江中下游圆口铜鱼的繁殖生物学. 水生生物学报, 2018, 42(5): 1010-1018. |
| [43] |
孙志禹, 朱挺兵, 杨德国, 李翀, 吴兴兵. 圆口铜鱼人工驯养繁育进展与展望. 淡水渔业, 2020, 50(1): 107-112. |
| [44] |
周湖海, 田辉伍, 何春, 唐锡良, 樊寒冰, 陈大庆, 段辛斌, 刘绍平. 金沙江下游巧家江段产漂流性卵鱼类早期资源研究. 长江流域资源与环境, 2019, 28(12): 2910-2920. |
| [45] |
程鹏. 长江上游圆口铜鱼的生物学研究[D]. 武汉: 华中农业大学, 2008.
|
| [46] |
高少波, 唐会元, 陈胜, 杨志, 董方勇. 金沙江一期工程对保护区圆口铜鱼早期资源补充的影响. 水生态学杂志, 2015, 36(2): 6-10. |
| [47] |
涂志英, 李丽萍, 袁喜, 王从锋, 黄应平. 圆口铜鱼幼鱼可持续游泳能力及活动代谢研究. 淡水渔业, 2016, 46(1): 33-38. |
| [48] |
杨少荣, 王小明. 金沙江下游梯级水电开发生态保护关键技术与实践. 人民长江, 2017, 48(S2): 54-56, 84. |
| [49] |
段辛斌, 陈大庆, 刘绍平, 池成贵, 杨如恒. 长江三峡库区鱼类资源现状的研究. 水生生物学报, 2002, 26(6): 605-611. |
| [50] |
杨志, 万力, 陶江平, 蔡玉鹏, 张原圆, 乔晔. 长江干流圆口铜鱼的年龄与生长研究. 水生态学杂志, 2011, 32(4): 46-52. |
| [51] |
熊美华, 邵科, 李伟涛, 朱滨. 圆口铜鱼资源变化及物种保护研究进展. 人民长江, 2023, 54(3): 63-71. |
| [52] |
杨志, 张鹏, 唐会元, 龚云, 董纯, 陈小娟, 赵娜. 金沙江下游圆口铜鱼生境适宜度曲线的构建. 生态科学, 2017, 36(5): 129-137. |
| [53] |
段辛斌, 刘绍平, 熊飞, 陈大庆, 杨如恒, 池成贵, 穆天荣. 长江上游干流春季禁渔前后三年渔获物结构和生物多样性分析. 长江流域资源与环境, 2008, 17(6): 878-885. |
| [54] |
胡兴坤, 邵科, 阙延福, 朱滨, 叶卿, 陈志刚, 李成成, 徐火清, 李伟涛. 金沙江下游宜宾段产漂流性卵鱼类早期资源动态变化. 水生态学杂志, 2023, 44(2): 73-80. |
| [55] |
张镀光, 王导群, 利广杰, 唐锡良, 段辛斌, 田辉伍. 乌东德水电站蓄水前库区江段鱼类资源研究. 人民长江, 2022, 53(10): 45-51. |
| [56] |
严莉. 长江铜鱼种群生物学及遗传多样性分析[D]. 武汉: 华中农业大学, 2005.
|
| [57] |
肖琼. 金沙江下游水利工程对鳅科鱼类的变动影响[D]. 长沙: 湖南农业大学, 2015.
|
| [58] |
熊飞, 刘红艳, 段辛斌, 刘绍平, 陈大庆. 长江上游江津和宜宾江段圆口铜鱼资源量估算. 动物学杂志, 2014, 49(6): 852-859. |
| [59] |
Chen S B, Watanabe S. Age dependence of natural mortality coefficient in fish population dynamics. Nippon Suisan Gakkaishi, 1989, 55(2): 205-208. |
| [60] |
Fieberg J, Ellner S P. When is it meaningful to estimate an extinction probability?. Ecology, 2000, 81(7): 2040-2047. |
| [61] |
张先锋, 王克雄. 长江江豚种群生存力分析. 生态学报, 1999, 19(4): 529-533. |
| [62] |
张先锋, 王丁, 王克雄. 漩涡模型及其在白暨豚种群管理中的应用. 生物多样性, 1994, 2(3): 133-139. |
| [63] |
吴斌, 王伟萍, 贺刚, 王海华. 不同模拟情景下长江江豚种群生存力分析. 渔业科学进展, 2021, 42(2): 28-35. |
| [64] |
武祥伟, 李光华, 毕保良, 于虹漫, 孔令富, 冷云, 张宇, 姜志武. 秀丽高原鳅种群生存力分析及最小可存活种群数估算. 水生生物学报, 2017, 41(3): 543-551. |
| [65] |
刘成汉. 四川长江干流主要经济鱼类若干繁殖生长特性. 四川大学学报: 自然科学版, 1980, 17(2): 181-188. |
| [66] |
王康, 肖洋, 毛劲乔, 龚轶青, 张培培, 王刚. 水温变化影响下金沙江下游典型鱼类繁殖期研究. 河海大学学报: 自然科学版, 2023, 51(2): 50-55. |
| [67] |
鲁雪报, 倪勇, 胡美洪, 刘勇, 曲焕韬, 吕小林, 方凯强. 圆口铜鱼人工繁殖的最适水温探讨. 科学养鱼, 2016(10): 6-7. |
| [68] |
张春光, 杨君兴, 赵亚辉. 金沙江流域鱼类. 北京: 科学出版社, 2019.
|
| [69] |
董纯, 罗安红, 陈小娟, 万成炎, 唐会元, 杨志. 不同年龄和性腺发育阶段圆口铜鱼营养成分分析. 水生态学杂志, 2022, 43(2): 108-115. |
| [70] |
Chen Q W, Zhang J Y, Chen Y C, Mo K L, Wang J, Tang L, Lin Y Q, Chen L, Gao Y, Jiang W, Zhang Y X. Inducing flow velocities to manage fish reproduction in regulated rivers. Engineering, 2021, 7(2): 178-186. |
| [71] |
祝茜, 刘焕章. 长江上游圆口铜鱼种群动态. 中国鱼类学会2008学术研讨会论文摘要汇编. 南昌, 2008: 123.
|
| [72] |
田书荣, 王明旭, 姜广顺, 李立, 康祖杰, 黄太福, 桂小杰. 武陵山脉东段华南虎重引入的种群生存力分析. 野生动物学报, 2022, 43(1): 5-16. |
| [73] |
余开, 赵永锋. 践行绿色发展理念促进大水面生态渔业高质量发展. 科学养鱼, 2021(7): 20-23. |
| [74] |
李雷, 危起伟, 吴金明, 杜浩, 谢晓. 长江宜宾江段渔业资源现状调查. 长江流域资源与环境, 2013, 22(11): 1449-1457. |
| [75] |
邬建国. 岛屿生物地理学理论: 模型与应用. 生态学杂志, 1989, 8(6): 34-39. |
| [76] |
Brook B W, O'Grady J J, Chapman A P, Burgman M A, Akçakaya H R, Frankham R. Predictive accuracy of population viability analysis in conservation biology. Nature, 2000, 404(6776): 385-387. |
| [77] |
Shaffer M L. Population viability analysis. Conservation Biology, 1990, 4(1): 39-40. |
| [78] |
杨青瑞, 陈冬红, 骆辉煌, 和晓荣, 张建国. 金沙江中游鲁地拉以上河段水电开发中鱼类保护措施. 水力发电, 2022, 48(2): 8-12. |
| [79] |
中国三峡集团, 三峡集团举办2021赤水河圆口铜鱼大规模增殖放流活动. https://www.thepaper.cn/newsDetail_forward_14476053. 2021-09-13.
|
| [80] |
吴俊东, 王翔, 翁永红, 袁达, 张文传. 乌东德水电站集运鱼系统方案设计. 人民长江, 2022, 53(2): 88-94. |
| [81] |
杨志, 乔晔, 张轶超, 朱迪, 常剑波. 长江中上游圆口铜鱼的种群死亡特征及其物种保护. 水生态学杂志, 2009, 30(2): 50-55. |
| [82] |
Zhang P, Li K F, Wu Y L, Liu Q Y, Zhao P X, Li Y. Analysis and restoration of an ecological flow regime during the Coreius guichenoti spawning period. Ecological Engineering, 2018, 123: 74-85. |