 2024, Vol. 44
2024, Vol. 44文章信息
- 毛军, 其曼古丽·拉提, 乔福生, 胡雷, 刘丹, 王长庭
- MAO Jun, Qimanguli PALATI, QIAO Fusheng, HU Lei, LIU Dan, WANG Changting
- 高寒草甸植物生态化学计量特征的空间格局及其影响因素
- Spatial patterns and driving factors of plants' ecological stoichiometric characteristics in the alpine meadow of western Sichuan Province
- 生态学报. 2024, 44(9): 3660-3675
- Acta Ecologica Sinica. 2024, 44(9): 3660-3675
- http://dx.doi.org/10.20103/j.stxb.202305151023
-
文章历史
- 收稿日期: 2023-05-15
- 网络出版日期: 2024-02-01

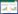




自Redfield发现海洋浮游生物碳(C)、氮(N)、磷(P) 元素的比值(摩尔比约为106∶16∶1) 与海洋表层水体中有机质∶N∶P存在一致性以来, 生态化学计量学蓬勃发展, 其研究对象几乎涵盖了海洋、湖泊、湿地、森林、草地和农田等多种生态系统[1—3]。作为生态化学计量学重要的分支之一, 植物生态化学计量学研究内容囊括了植物器官内各种大量元素(C、氢(H)、氧(O)、N和P等) 和微量元素(铁(Fe)、铜(Cu)、锰(Mn) 和锌(Zn) 等)的含量及比值特征, 以及它们在不同植物器官之间和不同环境梯度下的差异和潜在生态学机制[4]。由于C是构成植物骨架的最基本元素, 约占植物干质量的50%, 而N、P是植物蛋白质、DNA和RNA的重要组成元素, 也是陆地生态系统最主要的限制性营养元素, 目前植物生态化学计量学对这3种元素的研究最多[5—7]。并且, 植物生态化学计量学已经形成了“化学计量内稳性理论”[8]、“生长速率假说”[9]、“N∶P阈值假说”[10—11]、“温度-植物生理假说”和“温度-生物地球化学假说”[12]等理论和多种假说, 这些理论和假说对于理解植物的生理生态过程、生物地球化学循环过程以及判断植物养分限制状况等具有重要的理论和现实意义。
海拔是一个复杂的环境因子, 通过改变气温、降水、光照、土壤理化性质和生长季长度等综合影响植物生长[13]。高海拔地区的植物通过自然选择, 在形态和生理上都表现出很明显的适应性。在形态方面, 植物叶和芽常受到油脂类物质保护, 芽具有鳞片, 植物体表具有蜡粉和密毛, 植物矮小并常呈匍匐状、垫状或莲座状, 这些形态有助于保持较高的温度, 减少严寒的影响[14]。在生理方面, 植物也可以通过调节自身C、N、P含量及其生态化学计量比适应海拔引起的各种环境因子变化。例如, Lino等[15]在安第斯山脉发现蝇子草属植物Silene acaulis的非结构性碳水化合物浓度在高海拔(3150 m) 低于其他海拔(2600 m、2800 m和3000 m), 而Zhao等[16]发现高山嵩草(Kobresia pygmaea) 的可溶性糖、果糖和蔗糖含量均随海拔升高而增加。“温度-植物生理假说”认为, 植物通过增加N、P含量补偿低温引起的代谢速率下降, 这可以保障功能器官生长以提高资源竞争和防御能力[12]。例如, Bu等[17]发现高海拔地区(>3400 m) 203个植物种子N、P含量最高, 且海拔分别解释了N和P含量变化的4.3%和4.0%。但Macek等[18]研究表明海拔4500—6000 m渐尖早熟禾(Poa attenuata) 和扁芒菊(Waldheimia tridactylites) 的N含量随着海拔升高而下降, 同时, 随着海拔增加, 渐尖早熟禾P含量下降而扁芒菊P含量增加。相反, “温度-生物地球化学假说”则认为低温引起土壤微生物活性下降, 土壤N、P有效性低, 植物N、P含量下降[12], 因此在该假说下海拔升高引起植物N、P含量下降。然而, 也有研究表明海拔梯度上植物C、N、P含量及其生态化学计量比没有明显变化, 比如, 吴航等[19]分析了青藏高原54种菊科植物种子C、N、P含量与海拔的关系, 发现不同海拔高度的植物种子中C、N、P含量及其比值差异不显著, 但随着海拔升高, C、N含量具有增加而P含量具有下降的趋势。总之, 目前关于海拔梯度上植物C、N、P含量及其生态化学计量特征如何变化仍具有不确定性, 这种不确定性可能与植物生长环境中主要限制因子(如温度或降水)、土壤基质年龄以及植物自身的生存策略差异具有很大联系, 从而在不同气候环境、土壤基质和不同物种上会得出不同的响应模式[12, 20]。因此, 很有必要从不同植物功能群水平上研究海拔对植物C、N、P含量及其生态化学计量比的影响。此外, 纬度也可以通过调控温度影响植物生长, Reich和Oleksy[12]的研究表明, 全球植物叶片N、P含量在靠近赤道的低纬度地区(气温和生长季长度增加) 下降, N∶P则增加。但Zheng和Shangguan[21]发现黄土高原126种植物叶片N∶P随着纬度和年太阳辐射量的增加和年平均气温、年降水量的下降而增加, 说明高原地区的植物可能具有独特的生存策略。在青藏高原高寒草甸, 草本植物可分为禾本科、莎草科、豆科和杂类草4个植物功能群, 植物功能群是指基于植物分类和植物形态、生理、生活史等特性划分的对环境因子有相似反应、具有某些确定功能特征的物种群[22—23]。这些物种群往往具有不同的资源获取策略[22], 因此它们的C、N、P含量及其生态化学计量比对海拔和纬度的响应方式可能不同。比如: 相对于一些深根系的杂类草植物而言, 浅根系的禾本科和莎草科植物对土壤表层(如0—10 cm) 水分和养分的竞争力更强; 并且, 莎草科植物地上部分低矮、叶片退化, 这些特点降低了低温对莎草科植物生长的影响。因此, 海拔和纬度引起的气温和降水变化对禾本科和莎草科植物C、N、P含量及其生态化学计量比的影响可能较小[24—25]。大多数杂类草植物叶片较宽(如草玉梅(Anemone rivularis) 和黄帚橐吾(Ligularia virgaurea)), 根系分布较深, 这些特征有助于提高对光资源和土壤深层水分和养分的获取能力, 是一类耐干旱、贫瘠的植物[26]。豆科植物通过与根瘤菌形成共生从而具有固氮功能, 其N含量、N∶P可能比非豆科植物高, 但C∶N可能较低[27—28]。因此, 在区域尺度上研究地理格局和不同植物功能群间C、N、P含量及其生态化学计量比的变化, 对于深入认识植物的生态适应性策略至关重要。
青藏高原总面积约为250万km2, 平均海拔4000 m以上, 是世界上面积最大、海拔最高的高原, 素有“世界屋脊”、“世界第三极”和“亚洲水塔”之称[29—30]。高寒草甸是在海拔高、气温低(年平均温度0℃以下) 和水分适中(年降雨量约400—1000 mm左右) 的条件下形成的一种草地类型, 广泛分布于青藏高原东部及周围山地, 是当地牧民生产、生活的物质保障, 也是家畜和野生动物赖以生存的自然栖息地, 同时还具有C封存、水源涵养和生物多样性保护等重要功能[13, 31]。因此, 深入认识高寒草甸植物对高海拔的适应策略以及不同功能群之间的C、N、P生态化学计量比的差异, 从而建立科学合理的草地保护和管理模式, 对于维护青藏高原地区生态安全具有重要意义。根据前面提到的目前关于海拔梯度上植物C、N、P含量及其生态化学计量比变化的不确定性及其各功能群生态适应性策略的差异, 本研究在青藏高原东南缘高寒草甸选取60个样点, 重点研究海拔梯度上(3300—3500 m、3500—3700 m、3700—3900 m、3900—4100 m和4100—4300 m) 4种植物功能群(禾本科、莎草科、豆科和杂类草) C、N、P含量及其生态化学计量比的变化, 结合研究区地形因子(经纬度、坡度)、气候因子(年平均气温和年降水量) 以及土壤理化特征(密度、含水率和pH, C、N、P含量及其比值), 旨在探讨: 1) 植物C、N、P含量及其生态化学计量比在不同海拔和不同植物功能群之间有何差异? 2) 影响不同植物功能群的C、N、P含量及其生态化学计量比的主要因子是什么? 3) 这些因子对不同功能群C、N、P含量及其生态化学计量比的影响途径是什么? 研究结果有助于了解植物功能群C∶N∶P对海拔的响应方式及其潜在生态学机制, 为高寒草甸生态系统植物多样性和生态系统多功能性保护提供理论依据。
1 材料和方法 1.1 研究区域概况本研究区位于青藏高原东南部, 包括四川省阿坝藏族羌族自治州红原、阿坝、若尔盖和壤塘县, 以及甘孜藏族自治州石渠、甘孜、色达、道孚和白玉县, 共9个县(97.88—103.20°E, 30.72—36.10°N; 图 1), 采样点海拔3303—4334 m。研究区气候类型属于寒温带高原季风气候[32], 根据欧盟及欧洲中期天气预报中心等组织发布的ERA5-Land数据集, 研究区9个县2019和2020年的年平均气温(MAT) 在-5.41—1.00℃之间, 年降水量(MAP) 在793.26—1338.91 mm之间(图 1)。

|
| 图 1 采样点分布图和采样区不同海拔年平均气温和年降雨量 Fig. 1 The location of sampling sites and mean annual temperature and precipitation at different altitudes in the sampling area 气象站旁边的数字表示气象站的海拔, 经纬度为采样区经纬度, 不同大写字母表示不同海拔梯度间差异显著(P<0.05) |
植被类型为典型的高寒草甸植被, 物种组成按功能属性可分为禾本科、莎草科、豆科和杂类草4个类群[23]。其中, 禾本科优势植物主要包括垂穗披碱草(Elymus nutans)、草地早熟禾(Poa pratensis) 和川西剪股颖(Agrostis hugoniana) 等; 莎草科优势植物包括矮生嵩草(Kobresia humilis)、线叶嵩草(Kobresia capillifolia) 和双柱头藨草(Scirpus distigmaticus) 等; 豆科植物较少, 主要包括镰荚棘豆(Oxytropis falcata)、披针叶野决明(Thermopsis lanceolata) 和高山豆(Tibetia himalaica) 等; 杂类草较多, 主要包括蔷薇科、菊科、唇形科、毛茛科、龙胆科和伞形科, 其优势植物包括草玉梅、雪白委陵菜(Potentilla nivea)、黄帚橐吾、狼毒(Stellera chamaejasme)、细叶亚菊(Ajania tenuifolia)、乳白香青(Anaphalis acteal)、西伯利亚蓼(Polygonum sibiricum)、珠芽蓼(Polygonum viviparum) 和钝苞雪莲(Saussurea nigrescens) 等。植被生长期集中在5—9月, 生长期为120—140 d, 最高植被高度达45—60 cm[33]。土壤类型主要为亚高山草甸土, 土层深度40 cm以上[34—35]。根据我们以往的研究, 土壤pH介于5.91—6.87之间[36]。
1.2 样品采集与分析依托第二次青藏高原综合科学考察项目, 本研究于2019和2020年的7月下旬至9月上旬(地上生物量高峰期), 参考Cao等[7]和苏昊海等[37]的试验方法, 以海拔200 m为间距, 在海拔3300—3500 m、3500—3700 m、3700—3900 m、3900—4100 m和4100—4300 m上进行不同植物功能样品采集。海拔从低到高, 各海拔段采样点数分别为10、20、10、14和6个, 总计调查了60个样点(图 1)。所有样点之间相隔10 km以上, 均远离公路、住房等基础设施, 以尽可能避免人为干扰的影响。在每个样点选择1个100 m × 100 m的区域作为调查样地。首先, 利用GPS测定并记录样地的海拔、经度和纬度等地理信息。其次, 样地中间布设一条长100 m的样线, 沿着样线随机选取3个0.5 m × 0.5 m样方, 完成植物群落调查(盖度、物种数和株丛数) 之后将样方中的植物按不同功能群(禾本科、莎草科、豆科和杂类草) 齐地面剪下, 植物样品装入写好编号的信封袋中, 带回实验室放置在65 ℃的烘箱中烘干至恒重(72 h以上), 之后称重获得各植物功能群的生物量。然后, 在剪完植物的样方四角和中心位置先取5个环刀(体积100 cm3), 再用土钻(内径5 cm) 随机取5钻土样混合为一个样品, 每次取样深度为10 cm。按照这种方法依次取0—10 cm、10—20 cm和20—30 cm深度的土样。环刀中的土样带回实验室105 ℃烘干48 h至恒重后计算土壤含水率(SWC) 和密度(SBD)。土钻取得的土样过2 mm筛, 剔除根系和凋落物, 自然风干后测定pH、全C、全N和全P含量。
土壤pH采用电极法测定, 水土质量比为2.5∶1[38]。不同植物功能群的样品全部用球磨仪粉碎, 过2 mm筛保证样品混合均匀。使用元素分析仪(Vario Micro cube, Elementar, Berlin, Germany) 测定植物和土壤C、N含量, 植物和土壤样品先分别使用H2SO4-H2O2[38]和HClO4-H2SO4消煮, 再利用钼锑抗比色法测定P含量[39]。本研究中土壤含水率、密度、pH和C、N、P含量为0—30 cm土层的平均值, 植物和土壤C∶N、C∶P和N∶P为质量比。
1.3 数据分析采用单因素方差分析(ANOVA) 检验不同海拔MAP和MAT、不同植物功能群和土壤C含量、N含量、P含量、C∶N、C∶P和N∶P的差异, 显著水平为P < 0.05。分别采用最小显著差异法(LSD) 和Tamhane′s T2法对方差齐和方差不齐时不同海拔高度上述指标进行多重比较。利用降趋势对应分析(DCA) 分别对不同功能群C、N、P含量和C、N、P生态化学计量比进行分析, DCA排序结果中“Lengths of gradient”的第一排序轴梯度长度分别为0.4和0.5, 均小于3.0, 因此选用冗余分析(RDA) 进一步分析不同功能群C、N、P含量及其生态化学计量比与土壤理化性质(C、N、P含量及其化学计量比、土壤pH、密度和含水率)、地形因子(经纬度、坡度) 和气候因子(MAT和MAP) 之间的关系[40]。最后, 根据RDA分析结果得出显著影响不同植物功能群C、N、P含量及其生态化学计量比的环境因子, 利用AMOS 23.0构建结构方程模型分析这些环境因子对4个功能群C、N、P生态化学计量比的影响路径。图形在Origin 2021、Canoco 5.0、ArcMap 10.7.1和AMOS 23.0中绘制。
2 结果 2.1 不同海拔植物功能群C、N、P含量及其生态化学计量比随着海拔升高, 不同植物功能群C含量没有显著变化, 禾本科和杂类草植物N含量整体上增加, 豆科P含量则先增加后下降(表 1)。其中, 禾本科和杂类草N含量在海拔3900—4100 m和4100—4300 m显著高于其他海拔, 豆科P含量在海拔3700—3900 m显著高于4100—4300 m (P < 0.05)。同一海拔下, 仅豆科C、N含量与其他功能群具有显著差异, 即豆科C含量在3300—3500 m、3900—4100 m和4100—4300 m显著低于其他功能群C含量, 3700—3900 m显著低于禾本科C含量, 豆科N含量在3500—3700 m和3700—3900 m显著高于其他功能群。从整个海拔梯度来看, 豆科植物N含量和N∶P最高, 而C含量和C∶N最低, 禾本科C含量和C∶N最高; 研究区植物平均C、N、P含量分别为415.17 g/kg、21.30 g/kg和0.95 g/kg, C∶N、C∶P和N∶P分别为19.39、448.14和23.96 (表 2)。
| 变量Variable | 3300—3500 m | 3500—3700 m | 3700—3900 m | 3900—4100 m | 4100—4300 m |
| 禾本科C含量Grasses C content | 429.66±6.34Aa | 429.86±2.97Aa | 429.62±2.54Aa | 434.76±4.91Aa | 425.42±4.24Aa |
| 莎草科C含量Sedges C content | 433.82±8.18Aa | 431.15±3.29Aa | 413.10±15.62Aab | 422.37±4.60Aa | 426.21±4.85Aa |
| 豆科C含量legume C content | 369.97±20.56Ab | 423.89±7.60Aa | 384.06±22.43Ab | 385.39±24.92Ab | 370.87±24.33Ab |
| 杂类草C含量Forbs C content | 407.97±6.47Aa | 420.22±2.56Aa | 418.34±1.86Aab | 422.76±4.28Aa | 423.91±3.60Aa |
| 禾本科N含量Grasses N content | 17.74±2.20Ba | 18.17±0.80Bb | 18.53±1.63Bb | 24.17±1.48Aa | 21.98±1.43Aba |
| 莎草科N含量Sedges N content | 20.20±1.58Aa | 19.26±0.82Ab | 17.93±1.70Ab | 21.29±1.37Aa | 19.68±2.60Aa |
| 豆科N含量legume N content | 23.97±2.76Aa | 25.14±1.67Aa | 26.64±2.65Aa | 23.62±2.98Aa | 18.15±5.97Aa |
| 杂类草N含量Forbs N content | 20.12±2.44Ba | 19.64±0.87Bb | 19.40±1.73Bb | 25.20±1.14Aa | 25.13±1.60Aa |
| 禾本科P含量Grasses P content | 0.86±0.07Aa | 0.92±0.08Aa | 1.13±0.16Aa | 0.97±0.08Aa | 1.11±0.20Aa |
| 莎草科P含量Sedges P content | 0.79±0.09Aa | 0.96±0.10Aa | 0.92±0.13Aa | 0.82±0.15Aa | 0.90±0.21Aa |
| 豆科P含量legume P content | 0.77±0.10Aba | 0.86±0.08Aba | 1.09±0.16Aa | 0.85±0.12Aba | 0.60±0.19Ba |
| 杂类草P含量Forbs P content | 0.93±0.12Aa | 1.07±0.09Aa | 1.20±0.14Aa | 1.09±0.08Aa | 1.17±0.13 |
| 海拔高度从低到高, 样本数量分别为30、60、30、42和18; 不同大写字母表示不同海拔高度间差异显著, 不同小写字母表示不同植物功能群之间差异显著(P < 0.05) | |||||
| 功能群类别 Function group categories |
碳含量 C content/(g/kg) |
氮含量 N content/(g/kg) |
磷含量 P content/(g/kg) |
碳氮比 C∶N |
碳磷比 C∶P |
氮磷比 N∶P |
| 禾本科Grasses | 429.86 | 20.12 | 1.00 | 22.66 | 459.52 | 21.95 |
| 莎草科Sedges | 425.33 | 19.67 | 0.88 | 20.45 | 491.13 | 23.63 |
| 豆科Legume | 386.84 | 23.50 | 0.83 | 14.68 | 399.76 | 28.78 |
| 杂类草Forbs | 418.64 | 21.90 | 1.09 | 19.76 | 442.15 | 21.47 |
| 均值Mean | 415.17 | 21.30 | 0.95 | 19.39 | 448.14 | 23.96 |
4个功能群的C∶N整体上随着海拔增加而下降(图 2)。禾本科C∶N在海拔3300—3500 m (25.08)、3500—3700 m (24.68) 和3700—3900 m (23.40) 显著高于3900—4100 m (17.54)。莎草科C∶N在3500—3700 m (22.83) 显著高于3700—3900 m (19.38)、3900—4100 m (16.83) 和4100—4300 m (17.80)。豆科C∶N在3500—3700 m (16.39) 显著高于4100—4300 m (12.54)。杂类草C∶N在3500—3700 m (21.69) 显著高于3900—4100 m (16.89, P < 0.05)。除莎草科C∶P在3300—3500 m (595.59) 显著高于3700—3900 m (381.70) 之外, 不同功能群C∶P和N∶P随海拔变化不显著(图 2, P > 0.05)。此外, 不同海拔上禾本科、莎草科、豆科和杂类草的N∶P分别为: 19.52—27.15、19.94—27.53、26.81—31.45和18.69—24.54, 均大于16。

|
| 图 2 不同海拔4种植物功能群碳氮、碳磷和氮磷比 Fig. 2 C∶N, C∶P and N∶P ratios of four plant functional groups at different altitudes 海拔从低到高样本数量分别为30、60、30、42和18;不同大写字母表示不同海拔差异显著, 不同小写字母表示不同功能群差异显著(P < 0.05) |
同一海拔下, 豆科C∶N显著低于其他功能群, 海拔3500—3700 m杂类草C∶N (21.69) 显著低于禾本科和莎草科C∶N (分别为24.68和22.82)。海拔3300—3500 m豆科C∶P (416.80) 显著低于莎草科C∶P (595.59), 而其他海拔下不同功能群C∶P无显著变化。海拔3300—3500 m、3500—3700 m和3700—3900 m豆科N∶P显著高于禾本科和杂类草N∶P, 3900—4100 m和4100—4300 m不同功能群N∶P差异不显著(图 2)。
2.2 不同海拔土壤C、N、P含量及其生态化学计量比土壤N含量和N∶P随着海拔梯度的增加而增加, 但土壤C含量、P含量、C∶N和C∶P没有显著变化(图 3)。具体而言, 土壤N含量在海拔3300—3500 m显著低于3700—3900 m和4100—4300 m, 土壤N∶P在海拔3300—3500 m和3500—3700 m显著低于4100—4300 m。

|
| 图 3 不同海拔土壤碳、氮、磷含量及其生态化学计量比变化 Fig. 3 Changes in soil carbon, nitrogen and phosphorus contents at different elevations 不同大写字母表示不同海拔梯度间差异显著; ns, 差异不显著(P>0.05) |
如图 4所示, 植物群落中禾本科生物量比例和盖度变化与禾本科C、N、P含量及其生态化学计量比没有显著关系; 莎草科N含量与其生物量在群落中的比例显著负相关; 豆科C、N含量与其生物量在群落中的比例显著正相关, 同时, 豆科盖度与豆科C∶N极显著正相关; 群落中杂类草生物量比例与其N含量显著负相关而与C∶N极显著正相关, 杂类草盖度与其C含量、C∶N显著正相关, 但杂类草盖度与杂类草N含量显著负相关。

|
| 图 4 不同植物功能群C、N、P含量及其生态化学计量比与植被特征的Pearson相关性 Fig. 4 Pearson correlations between C, N, P contents and their ecological stoichiometric ratios of different plant functional groups with vegetation characteristics Gra, 禾本科; Sed, 莎草科; Leg, 豆科; Forbs, 杂类草; Com.B, 植物群落生物量; Gra.BR、Sed.BR、Leg.BR和Forbs BR分别是禾本科、莎草科、豆科和杂类草生物量占群落生物量的比例; Com.cov、Gra. cov、Sed. cov、Leg. cov和Forbs cov分别是植物群落、禾本科、莎草科、豆科和杂类草植物的盖度 |
根据不同功能群C、N、P含量与土壤、地形和气候因子的RDA分析(图 5; 表 3), 纬度、MAP、土壤pH、土壤C含量和土壤N∶P是显著影响不同功能群C、N和P含量的因子(P < 0.05), 其解释率分别为12.40%、5.20%、5.10%、4.80%、3.60%和4.00%。结合Pearson相关性分析(图 6), 本研究发现纬度与禾本科C和杂类草N显著正相关(P<0.05), 而与4个植物功能群的P含量极其显著负相关(P < 0.01)。MAP与禾本科和杂类草N含量显著负相关, 但MAP与莎草科、豆科和杂类草P含量显著负相关。土壤pH与杂类草C含量及禾本科、莎草科和杂类草P含量显著负相关, 而与杂类草N显著正相关。土壤N∶P与杂类草C显著正相关, 土壤C含量则只与豆科C含量显著正相关(图 3, 图 6)。此外, 海拔与禾本科N含量和杂类草C、N含量显著正相关(图 6)。

|
| 图 5 不同功能群C、N、P含量与土壤、地形和气候因子冗余分析(RDA) 排序图 Fig. 5 Redundancy analysis (RDA) of C, N, and P contents of different plant functional groups with soil, topographic and climatic factors Gra, 禾本科; Sed, 莎草科; Leg, 豆科; Forbs, 杂类草; SBD, 土壤密度; SWC, 土壤含水率; Altitude, 海拔; Longitude, 经度; Latitude, 纬度; Slope, 坡度; MAP, 年平均降雨量; MAT, 年平均气温 |
| 因子Factor | 解释率 Explains/% |
F | P | 因子Factor | 解释率 Explains/% |
F | P | |
| 纬度Latitude | 12.40 | 6.20 | 0.002** | 土壤C∶N Soil C∶N | 1.40 | 0.90 | 0.542 | |
| 经度Longitude | 1.50 | 1.00 | 0.466 | 土壤C∶P Soil C∶P | 2.40 | 1.50 | 0.172 | |
| 海拔Altitude | 2.30 | 1.40 | 0.196 | 土壤N∶P Soil N∶P | 4.00 | 2.40 | 0.026* | |
| 年平均气温MAT | 2.20 | 1.30 | 0.204 | pH | 4.80 | 2.60 | 0.012* | |
| 年降雨量MAP | 5.20 | 2.70 | 0.006** | SBD | 1.00 | 0.60 | 0.785 | |
| 土壤碳含量Soil C content | 3.60 | 2.10 | 0.005** | SWC | 1.80 | 1.10 | 0.354 | |
| 土壤N含量Soil N content | 3.30 | 2.00 | 0.072 | 坡度Slope | 0.60 | 0.30 | 0.952 | |
| 土壤P含量Soil P content | 3.40 | 1.90 | 0.094 | |||||
| MAT, 年平均气温; MAP, 年降雨量; SBD, 土壤密度; SWC, 土壤含水率; 黑色字体表示差异显著, *表示P ≤ 0.05, ** 表示P ≤ 0.01 | ||||||||

|
| 图 6 不同植物功能群C、N、P含量及其生态化学计量比与土壤理化、地形因子的Pearson相关性 Fig. 6 Pearson correlations between C, N, P contents and their ecological stoichiometric ratios of different plant functional groups with soil physicochemical properties and topographic factors Gra, 禾本科; Sed, 莎草科; Leg, 豆科; Forbs, 杂类草; SBD, 土壤密度; SWC, 土壤含水率; Altitude, 海拔; Longitude, 经度; Latitude, 纬度; Slope, 坡度; MAP, 年降雨量; MAT, 年平均气温 |
纬度、海拔和土壤C、P含量是显著影响不同功能群C∶N、C∶P和N∶P的因子(P < 0.05), 其解释率分别为15.70%、8.90%、5.00%和6.30% (图 7; 表 4)。纬度与禾本科、莎草科和杂类草C∶P和N∶P显著正相关。海拔不仅与4种功能群C∶N极显著负相关, 而且还与豆科C∶P显著负相关(图 6)。土壤C与各功能群C∶N、C∶P和N∶P无显著相关性。土壤P与禾本科和杂类草C∶N显著正相关, 与禾本科的C∶P、N∶P, 莎草科C∶P, 以及杂类草N∶P显著负相关(图 6)。

|
| 图 7 不同植物功能群C∶N、C∶P和N∶P与土壤、地形和气候因子RDA排序图 Fig. 7 Redundancy analysis (RDA) of C∶N, C∶P and N∶P of different plant functional groups with soil, topographic and climatic factors Gra, 禾本科; Sed, 莎草科; Leg, 豆科; Forbs, 杂类草; SBD, 土壤密度; SWC, 土壤含水率; Altitude, 海拔; Longitude, 经度; Latitude, 纬度; Slope, 坡度; MAP, 年降雨量; MAT, 年平均气温 |
| 因子Factors | 解释率 Explains/% |
F | P | 因子Factors | 解释率 Explains/% |
F | P | |
| 纬度Latitude | 15.70 | 8.20 | 0.002** | 土壤C∶N Soil C∶N | 0.80 | 0.50 | 0.762 | |
| 经度Longitude | 2.30 | 1.60 | 0.144 | 土壤C∶P Soil C∶P | 2.20 | 1.50 | 0.14 | |
| 海拔Altitude | 8.90 | 5.10 | 0.002** | 土壤N∶P Soil N∶P | 3.40 | 2.10 | 0.088 | |
| 年平均气温MAT | 1.20 | 0.80 | 0.534 | pH | 1.40 | 1.00 | 0.44 | |
| 年降雨量MAP | 0.80 | 0.50 | 0.764 | SBD | 0.50 | 1.00 | 0.928 | |
| 土壤C Soil C | 5.00 | 3.30 | 0.004** | SWC | 2.40 | 1.60 | 0.146 | |
| 土壤N Soil N | 1.00 | 0.70 | 0.632 | 坡度Slope | 1.20 | 0.80 | 0.528 | |
| 土壤P Soil P | 6.30 | 3.90 | 0.01* | |||||
| MAT, 年平均气温; MAP, 年降雨量; SBD, 土壤密度; SWC, 土壤含水率; 黑色字体表示差异显著, *表示P ≤ 0.05, ** 表示P ≤ 0.01 | ||||||||
构建结构方程模型(SEM) 分析显著影响不同功能群C∶N、C∶P和N∶P的环境因子(表 4) 对它们的影响路径(图 8)。其中, 土壤C含量与各功能群C、N、P生态化学计量比无显著相关性(图 6), 因此模型中没有纳入该指标。总体来看, 地形因子(海拔和纬度) 通过改变气候因子(MAT和MAP) 从而影响植物的C、N、P生态化学计量比。SEM显示海拔和纬度增加共同导致MAT和MAP下降, 且MAT和MAP互相影响(路径系数-0.52, P < 0.001)。MAP对土壤P含量具有极其显著正效应, 土壤P则分别对禾本科N∶P和莎草科C∶P具有显著负效应, 但土壤P对杂类草C∶N具有显著正效应。MAT对4个功能群C∶N、C∶P和N∶P的影响不同, MAT对禾本科C∶N、C∶P和N∶P没有显著影响, 但对莎草科C∶N、C∶P和N∶P具有显著正效应, MAT对豆科C∶P、N∶P以及杂类草C∶P具有显著正效应而对豆科和杂类草的C∶N具有显著负效应。MAP对豆科C∶N具有显著正效应, 而MAP对莎草科和杂类草C∶P和N∶P具有显著负效应。此外, 4个功能群的C∶P和N∶P互相影响(路径系数0.68—0.77, P < 0.001)。

|
| 图 8 环境因子对禾本科、莎草科、豆科和杂类草C∶N、C∶P和N∶P比影响的结构方程模型 Fig. 8 Structural equation models for the effects of environmental factors on the C∶N, C∶P and N∶P ratios of grasses, sedges, legume, and forbs Grasses, 禾本科; Sedges, 莎草科; Legume, 豆科; Forbs, 杂类草; MAP, 年平均降雨量; MAT, 年平均气温; Altitude, 海拔; Latitude, 纬度; *, P < 0.05; **, P < 0.01; ***, P < 0.001; 禾本科模型: χ2=4.040, df=7, GFI=0.984, AGFI=0.916; RMSEA=0.000, P=0.775; 莎草科模型: χ2=9.910, df=13, GFI=0.963, AGFI=0.897; RMSEA=0.000, P=0.701; 豆科模型: χ2=8.197, df=11, GFI=0.968, AGFI=0.896; RMSEA=0.000, P=0.696; 杂类草模型: χ2=9.247, df=12, GFI=0.964, AGFI=0.893; RMSEA=0.000, P=0.682; χ2: 卡方值; df: 自由度; GFI: 适配指数; AGFI: 调整后适配度指数; RMSEA: 近似误差均方根 |
C是构成植物体骨架的基本元素, 约占植物生物量干质量的50%, 并以结构性(如木质素、纤维素) 和非结构性碳水化合物(如葡萄糖、果糖和麦芽糖等) 的形式存在, 结构性碳水化合物作为植物的基本骨架, 在植物体中变化较小[5, 32]。本研究发现海拔梯度上各功能群C含量无显著变化(表 1), 表明研究区植物C含量受海拔升高引起的温湿度、太阳辐射和风速等环境因子变化的影响较小。这与洪江涛等[41]在区域尺度上发现藏北高寒草原紫花针茅(Stipa purpurea) 根系C含量相对稳定, 以及贺合亮等[42]发现青藏高原东部窄叶鲜卑花(Sibiraea angustata) 根、茎和叶等器官C含量相对稳定的研究结果相似。研究区植物平均C含量为415.17 g/kg (表 2), 低于全球492种陆地植物叶片C含量(464 g/kg) 和中国草地生态系统植物叶片C含量(438 g/kg)[2, 43]。不同植物功能群C含量与MAP呈负相关(图 5), 说明本研究区C含量相对较低可能是因为青藏高原东南缘降水量较丰富, 草本植物组织相对幼嫩。同时, 海拔梯度上各植物功能群C含量相对稳定也表明研究区的草本植物并没有分配更多的C到防御性器官(如根和茎的表皮) 以抵御严寒, 其生长策略可能不是保守生长策略, 而是积极资源获取策略[44—45]。
本研究区植物平均N、P含量分别为21.30 g/kg和0.95 g/kg (表 2), N含量接近全球草本植物N含量(21.72 g/kg), 而P含量则低于全球植物(1.58 g/kg)、中国陆生植物(1.21 g/kg) 和全球草本植物P含量(1.64 g/kg)[2, 43], 说明研究区植物P含量较低, P可能是该地区植物生长的限制性养分。海拔梯度上, 3900—4100 m和4100—4300 m禾本科和杂类草N含量显著增加(表 1), 尽管土壤N含量也随着海拔升高而增加(图 3), 但因土壤全N含量不代表植物能够利用的N形态, 禾本科和杂类草N含量与土壤N含量并无显著相关性(图 6), 这些结果表明禾本科和杂类草具有提高自身N含量适应低温环境(图 1) 的生态适应性策略, 符合“温度-植物生理假说”, 即气温下降引起植物代谢速率变慢, 不利于植物生长, 但植物通过增加N或P含量可以补偿代谢速率下降从而适应低温环境[7, 12, 24]。莎草科是青藏高原高寒草甸主要的原生物种, 在长期进化过程中, 这类植物可能通过改变自身形态而不是调节N含量以适应高海拔环境。例如, 莎草科植物通常具有植株低矮和丛生的特点(如矮生嵩草), 这些特征有利于保持较高的温度, 减少低温对其生长的影响, 因而海拔升高没有引起莎草科植物N发生明显改变(表 1)[14]。豆科植物N含量在不同海拔保持稳定则归因于其能够与根瘤菌形成共生关系, 根瘤菌可以把N2合成为含N化合物满足豆科对N的需求[28]。
禾本科、莎草科和杂类草P含量随着海拔升高无显著变化(表 1), 表明这些功能群具有一定的P “生态化学计量学内稳性”。一方面, P是植物细胞质膜、DNA和RNA的重要组成元素[6—7], 植物维持P含量稳定有助于保持正常的生长状态[2]。另一方面, 这可能归因于不同海拔土壤P含量没有显著变化(图 3)。根据“温度-生物地球化学假说”, 温度下降引起土壤微生物活性下降, 土壤N、P有效性降低从而导致植物N、P含量下降[12]。虽然豆科P含量在海拔4100—4300 m显著低于3700—3900 m (表 1), 但豆科P含量与土壤P含量没有显著关系(图 6), 表明海拔4100—4300 m豆科P含量下降并不是土壤P含量变化引起的。豆科P含量与MAP显著正相关(图 6), 这表明4100—4300 m豆科P含量下降是由海拔升高引起MAP下降(图 1) 引起的, 这可能是降水量不足导致土壤有效磷含量或者豆科植物对土壤P的获取、吸收和利用效率较低引起的[12, 37, 46]。总之, 在适应环境变化方面, 不同植物功能群具有调节生理、形态特征或与微生物形成共生关系的策略。
3.2 环境因子对不同植物功能群C∶N、C∶P和N∶P的影响植物C∶N可以表征N素利用效率, C∶N越高, N素利用效率越高[47]。本研究区植物平均C∶N为19.39 (表 2), 高于He等[43]报道的青藏高原高寒草甸植物叶片C∶N (17), 这种差异可能是因为本研究测定的是不同植物功能群整株的N含量, 而由于植物叶片N含量相对其他器官高(如茎), 这使得单独测定植物叶片N含量得出的C∶N会低于测定整个植株N含量得到的C∶N。同时, 本研究得到的C∶N (19.39) 低于全球(43.6) 植物叶片C∶N, 以及黄土高原126种植物叶片C∶N (21.2), 这可能是由于研究区不同植物功能群C含量较低引起的。4个功能群C∶N整体上随着海拔增加而下降(图 2), 表明海拔升高引起各植物功能群N利用效率下降。一方面, 4种植物功能群的C∶N均与MAP显著正相关(图 6, 图 7), 说明海拔升高引起的MAP下降会降低植物N利用效率。这可以解释为水分不足引起土壤N向植物根系表面的扩散率和根系对N的吸收和运输速率下降, 从而降低了植物对土壤N的利用效率[46]。另一方面, 研究区各功能群C含量随着海拔升高相对稳定(表 1), 而禾本科和杂类草植物通过提高自身N含量适应海拔引起的低温环境(图 1), 这必然引起禾本科和杂类草C∶N下降。对于莎草科和豆科而言, 二者的C∶N均与海拔显著负相关, 且豆科C∶N还与MAP显著正相关(图 6; 图 7), 结构方程模型(图 8) 显示MAT对莎草科C∶N具有显著负效应而MAP对豆科C∶N具有显著正效应, 因此, 海拔4100—4300 m低温和少雨(图 1) 导致这2个功能群N利用效率下降, 其内在过程可能是低温引起植物代谢速率变慢所致。此外, He等[48]发现内蒙古温带草原、西藏高寒草原和新疆山地草原植物叶片C∶N在共存物种和不同植被类型间差异显著, 植物生命形式和属性特征解释了70%的C∶N变异, 而生长季平均温度和降水量解释率仅为3%。本研究通过分析植被特征(如各功能群的生物量占群落生物量的比例) 与不同植物功能群C、N、P含量及其生态化学计量比的关系(图 4), 发现不同植物功能群C∶N与各植物功能群生物量占群落生物量的比例没有显著相关性, 但是豆科C∶N和杂类草的C∶N与各自的盖度显著正相关, 说明豆科和杂类草植物的盖度影响其C∶N, 其主要机制可能是盖度通过影响光合作用固定的C从而影响C∶N。因此, 海拔梯度上不同植物功能群内部特征的改变也是造成C∶N差异的原因之一。
生态化学计量学“生长速率假说”认为, 快速生长的有机体需要大量的核糖体核糖核酸(rRNA) 指导蛋白质的合成, 因此C∶P和N∶P较低[9]。本研究区植物平均C∶P、N∶P分别为448.14和23.96 (表 2), C∶P高于黄土高原14种草本植物C∶P (262), N∶P高于全球草本植物N∶P (13.3)、中国草本植物N∶P (13.5) 和He等[43]报道的青藏高原高寒草甸植物叶片N∶P (14.6), 说明研究区植物生长速率相对较慢, 这主要是本研究区植物P含量低引起的。海拔梯度上, 莎草科C∶P在3700—3900 m显著低于3300—3500 m, 表明3700—3900 m莎草科植物生长速率比3300—3500 m快, 其原因可能是更高海拔地区的植物生长季越短, 植物需要快速生长以在较短的时间内完成一系列的生长繁殖过程, 因此, 植物分配更多的P到地上部分从而导致C∶P和N∶P下降[19, 49]。除了莎草科C∶P之外, 海拔梯度上不同植物功能群C∶P、N∶P无显著变化(图 2), 其中N∶P的研究结果与Han等[50]发现我国753种陆生植物叶片N∶P随纬度增加无显著变化的研究结果相似。这反映了不同植物功能群C∶P和N∶P具有一定程度的保持自身C、N、P生态化学计量比相对稳定的能力, 符合生态化学计量学“内稳性”理论。这种“内稳性”的维持有利于植物保持最佳生长状态从而完成一系列的生长和繁殖过程[4]。同时, N∶P也是反映植物生长受N或P限制的重要指标, Koerselman和Arthur[10]的研究表明, N∶P < 14, 植物生长倾向于受N限制; N∶P > 16, 植物生长倾向于受P限制; N∶P介于14和16之间, 植物生长受N、P共同限制或不受N、P限制。Güsewell[11]通过整合大量的施肥实验数据, 认为N∶P小于10和大于20更适合判断植物生长受N限制和P限制。不同海拔梯度上禾本科、莎草科、豆科和杂类草的N∶P分别为: 19.52—27.15、19.94—27.53、26.81—31.45和18.69—24.54, 各植物功能群生长倾向于受P限制, 因此, 在对研究区高寒草甸生态系统进行管理或恢复时可以考虑施P肥。
此外, 本研究还发现与其他功能群相比, 豆科植物N含量和N∶P较高, 而C含量和C∶N较低(表 1; 图 2), 这种研究结果与以往报道的与非豆科植物相比, 豆科植物C∶N高而N∶P低的研究报道一致[43, 49]。一方面, 这种现象归因于豆科植物与根瘤菌形成共生关系从而能够获取更多的N, 且这也间接说明植物的生态适应性策略对其C∶N∶P具有重要影响, 因而在构建植物C、N、P生物地球化学模型中应考虑植物适应性策略的差异[28]。另一方面, 豆科植物C含量低则可能与这类植物常呈匍匐状, 不需要投资更多的C形成结构性碳水化合物(如纤维素和木质素) 以支撑地上部分有关[14]。海拔3500—3700 m杂类草C∶N显著低于禾本科C∶N而高于豆科C∶N (图 2), 表明该海拔下杂类草植物N利用效率低于禾本科植物而高于豆科植物。由于4种功能群的C∶N均与MAP极显著正相关(图 4; 图 6), 因此在同一条件下, 根系较浅的禾本科比杂类草具有更大的优势来利用土壤表层相对丰富的水分和土壤N素, 导致禾本科C∶N高于杂类草C∶N; 但对于豆科植物而言, 其较低的C含量导致它们的C∶N低于其他功能群[26]。
4 结论在区域尺度上, 通过研究青藏高原东南缘60个高寒草甸样地不同植物功能群C、N、P含量及其生态化学计量比空间格局和影响因素, 本研究得出以下结论: 1)海拔梯度上, 高寒草甸不同植物功能群具有一定的生态化学计量学内稳性, 这表现在海拔梯度上各植物功能群C含量, 豆科和莎草科植物N含量, 禾本科、莎草科和杂类草P含量, 以及除莎草科C∶P之外的不同功能群C∶P和N∶P能够保持相对稳定。2) 禾本科和杂类草植物通过提高自身N含量适应高海拔(3900—4100 m和4100—4300 m) 环境, 验证了“温度-植物生理假说”。3)青藏高原东南高寒草甸植物P含量较低, 植物生长受P限制。4) 海拔升高引起的低温和少雨环境导致植物氮素利用效率下降(C∶N低)。5)与其他功能群相比, 豆科植物整体上具有高N含量和N∶P, 低C含量、C∶N和C∶P的特点。总之, 植物功能群自身适应性策略和海拔、纬度共同驱动的气温及降水调控了不同植物功能群C、N、P含量及其生态化学计量比在海拔梯度上的变化模式。
| [1] |
Redfield A C. The biological control of chemical factors in the environment. Science Progress, 1960, 11: 150-170. |
| [2] |
Elser J J, Fagan W F, Denno R F, Dobberfuhl D R, Folarin A, Huberty A, Interlandi S, Kilham S S, McCauley E, Schulz K L, Siemann E H, Sterner R W. Nutritional constraints in terrestrial and freshwater food webs. Nature, 2000, 408(6812): 578-580. DOI:10.1038/35046058 |
| [3] |
曾德慧, 陈广生. 生态化学计量学: 复杂生命系统奥秘的探索. 植物生态学报, 2005, 29(6): 1007-1019. |
| [4] |
田地, 严正兵, 方精云. 植物生态化学计量特征及其主要假说. 植物生态学报, 2021, 45(7): 682-713. |
| [5] |
Ågren G I. Stoichiometry and nutrition of plant growth in natural communities. Annual Review of Ecology, Evolution, and Systematics, 2008, 39: 153-170. DOI:10.1146/annurev.ecolsys.39.110707.173515 |
| [6] |
Tang Z Y, Xu W T, Zhou G Y, Bai Y F, Li J X, Tang X L, Chen D M, Liu Q, Ma W H, Xiong G M, He H L, He N P, Guo Y P, Guo Q A, Zhu J L, Han W X, Hu H F, Fang J Y, Xie Z Q. Patterns of plant carbon, nitrogen, and phosphorus concentration in relation to productivity in China's terrestrial ecosystems. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(16): 4033-4038. |
| [7] |
Cao J J, Wang X Y, Adamowski J F, Biswas A, Liu C F, Chang Z Q, Feng Q. Response of leaf stoichiometry of Oxytropis ochrocephala to elevation and slope aspect. Catena, 2020, 194: 104772. DOI:10.1016/j.catena.2020.104772 |
| [8] |
Elser J J, Acharya K, Kyle M, Cotner J, Makino W, Markow T, Watts T, Hobbie S, Fagan W, Schade J, Hood J, Sterner R W. Growth rate-stoichiometry couplings in diverse biota. Ecology Letters, 2003, 6: 936-943. DOI:10.1046/j.1461-0248.2003.00518.x |
| [9] |
Elser J J, Fagan W F, Kerkhoff A J, Swenson N G, Enquist B J. Biological stoichiometry of plant production: metabolism, scaling and ecological response to global change. New Phytologist, 2010, 186(3): 593-608. DOI:10.1111/j.1469-8137.2010.03214.x |
| [10] |
Koerselman W, Meuleman A F M. The vegetation N ∶ P ratio: a new tool to detect the nature of nutrient limitation. Journal of Applied Ecology, 1996, 33(6): 1441. DOI:10.2307/2404783 |
| [11] |
Güsewell S. N ∶ P ratios in terrestrial plants: variation and functional significance. New Phytologist, 2004, 164(2): 243-266. DOI:10.1111/j.1469-8137.2004.01192.x |
| [12] |
Reich P B, Oleksyn J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(30): 11001-11006. |
| [13] |
Wang A, Wang X, Tognetti R, Lei J P, Pan H L, Liu X L, Jiang Y, Wang X Y, He P, Yu F H, Li M H. Elevation alters carbon and nutrient concentrations and stoichiometry in Quercus aquifolioides in southwestern China. Science of the Total Environment, 2018, 622: 1463-1475. |
| [14] |
尚玉昌. 普通生态学. 3版. 北京: 北京大学出版社, 2010.
|
| [15] |
Lino M C G, Cavieres L A, Zotz G, Bader M Y. Carbohydrate reserves in the facilitator cushion plant Laretia acaulis suggest carbon limitation at high elevation and no negative effects of beneficiary plants. Oecologia, 2017, 183(4): 997-1006. DOI:10.1007/s00442-017-3840-5 |
| [16] |
Zhao N, He N P, Wang Q F, Zhang X Y, Wang R L, Xu Z W, Yu G R. The altitudinal patterns of leaf C ∶ N ∶ P stoichiometry are regulated by plant growth form, climate and soil on Changbai Mountain, China. PLoS one, 2014, 9(4): e95196. DOI:10.1371/journal.pone.0095196 |
| [17] |
Bu H Y, Jia P, Qi W, Liu K, Xu D H, Ge W J, Wang X J. The effects of phylogeny, life-history traits and altitude on the carbon, nitrogen, and phosphorus contents of seeds across 203 species from an alpine meadow. Plant Ecology, 2018, 219: 737-748. DOI:10.1007/s11258-018-0830-6 |
| [18] |
Macek P, Klimeš L, Adamec L, Doležal J, Chlumská Z, de Bello F, Dvorský M, Šeháková K. Plant nutrient content does not simply increase with elevation under the extreme environmental conditions of Ladakh, NW Himalaya. Arctic, Antarctic, and Alpine Research, 2012, 44(1): 62-66. DOI:10.1657/1938-4246-44.1.62 |
| [19] |
吴航, 王顺霞, 卜海燕, 张世挺, 葛文静, 王学经, 齐威, 刘坤. 青藏高原53种菊科植物种子中碳、氮、磷含量与种子大小和海拔的关系. 西北植物学报, 2014, 34(8): 1635-1641. |
| [20] |
Baxter I, Dilkes B P. Elemental profiles reflect plant adaptations to the environment. Science, 2012, 336(6089): 1661-1663. DOI:10.1126/science.1219992 |
| [21] |
Zheng S X, Shangguan Z P. Spatial patterns of leaf nutrient traits of the plants in the Loess Plateau of China. Trees, 2007, 21: 357-370. |
| [22] |
刘育红, 杨元武, 张英. 退化高寒草甸植物功能群与土壤因子关系的冗余分析. 生态与农村环境学报, 2018, 34(12): 1112-1121. |
| [23] |
王长庭, 龙瑞军, 丁路明. 高寒草甸不同草地类型功能群多样性及组成对植物群落生产力的影响. 生物多样性, 2004, 12(4): 403-409. |
| [24] |
Du B M, Ji H W, Peng C, Liu X J, Liu C J. Altitudinal patterns of leaf stoichiometry and nutrient resorption in Quercus variabilis in the Baotianman Mountains, China. Plant and Soil, 2017, 413: 193-202. |
| [25] |
Lambers H. Phosphorus acquisition and utilization in plants. Annual Review of Plant Biology, 2022, 73: 17-42. |
| [26] |
李军豪, 杨国靖, 王少平. 青藏高原区退化高寒草甸植被和土壤特征. 应用生态学报, 2020, 31(6): 2109-2118. |
| [27] |
Yang Y, Liu B R, An S S. Ecological stoichiometry in leaves, roots, litters and soil among different plant communities in a desertified region of Northern China. Catena, 2018, 166: 328-338. |
| [28] |
Wei X R, Reich P B, Hobbie S E. Legumes regulate grassland soil N cycling and its response to variation in species diversity and N supply but not CO2. Global Change Biology, 2019, 25(7): 2396-2409. |
| [29] |
Qiu J. China: The third pole. Nature, 2008, 454(7203): 393-397. |
| [30] |
Dong S K, Sherman R. Enhancing the resilience of coupled human and natural systems of alpine rangelands on the Qinghai-Tibetan Plateau. The Rangeland Journal, 2015, 37(1): i-iii. |
| [31] |
Xiao J N, Dong S K, Zhao Z Z, Han Y H, Li S, Shen H, Ding C X. Stabilization of soil organic carbon in the alpine meadow is dependent on the nitrogen deposition level on the Qinghai-Tibetan Plateau. Ecological Engineering, 2021, 170: 106348. |
| [32] |
杨德春, 胡雷, 宋小艳, 王长庭. 降雨变化对高寒草甸不同植物功能群凋落物质量及其分解的影响. 植物生态学报, 2021, 45(12): 1314-1328. |
| [33] |
王鑫, 王长庭, 胡雷, 宋小艳. 积雪变化对高寒草甸钝苞雪莲(Saussurea nigrescens)繁殖分配及功能属性的影响. 生态学报, 2021, 41(19): 7858-7869. DOI:10.5846/stxb201907051414 |
| [34] |
Yuan Z Q, Jiang X J, Liu G J, Jin H J, Chen J, Wu Q B. Responses of soil organic carbon and nutrient stocks to human-induced grassland degradation in a Tibetan alpine meadow. Catena, 2019, 178: 40-48. |
| [35] |
唐立涛, 毛睿, 王长庭, 李洁, 胡雷, 字洪标. 氮磷添加对高寒草甸植物群落根系特征的影响. 草业学报, 2021, 30(9): 105-116. |
| [36] |
毛军. 川西北退化高寒草甸不同功能群植物-土壤生态化学计量特征[D]. 成都: 西南民族大学, 2021.
|
| [37] |
苏昊海, 张小芳, 牛亚琳, 王海茹, 曹建军. 祁连山狼毒(Stellera chamaejasme) 叶片生态化学计量特征对海拔的响应. 中国沙漠, 2021, 41(6): 205-212. |
| [38] |
Li S J, Wang F W, Chen M F, Liu Z Y, Zhou L Y, Deng J, Dong C J, Bao G C, Bai T S, Li Z, Guo H, Wang Y, Qiu Y P, Hu S J. Mowing alters nitrogen effects on the community-level plant stoichiometry through shifting plant functional groups in a semi-arid grassland. Environmental Research Letters, 2020, 15(7): 074031. |
| [39] |
Zhang W J, Xue X, Peng F, You Q G, Hao A H. Meta-analysis of the effects of grassland degradation on plant and soil properties in the alpine meadows of the Qinghai-Tibetan Plateau. Global Ecology and Conservation, 2019, 20: e00774. |
| [40] |
王洪丹. 黄土区露天煤矿排土场土壤与地形因子对植被恢复的影响研究[D]. 北京: 中国地质大学(北京), 2016.
|
| [41] |
洪江涛, 吴建波, 王小丹. 藏北高寒草原紫花针茅根系碳氮磷生态化学计量学特征. 山地学报, 2014, 32(4): 467-474. |
| [42] |
贺合亮, 阳小成, 李丹丹, 尹春英, 黎云祥, 周国英, 张林, 刘庆. 青藏高原东部窄叶鲜卑花碳、氮、磷化学计量特征. 植物生态学报, 2017, 41(1): 126-135. |
| [43] |
He J S, Wang L, Flynn D F B, Wang X P, Ma W H, Fang J Y. Leaf nitrogen: phosphorus stoichiometry across Chinese grassland biomes. Oecologia, 2008, 155: 301-310. |
| [44] |
所尔阿芝, 字洪标, 罗雪萍, 程瑞希, 王艳丽, 毛睿, 王长庭. 青海省4种常见树木碳氮磷生态化学计量特征. 应用与环境生物学报, 2019, 25(4): 783-790. |
| [45] |
祁鲁玉, 陈浩楠, 库丽洪·赛热别力, 籍天宇, 孟高德, 秦慧颖, 王宁, 宋逸欣, 刘春雨, 杜宁, 郭卫华. 基于植物功能性状的暖温带5种灌木幼苗生长策略. 植物生态学报, 2022, 46(11): 1388-1399. |
| [46] |
Soethe N, Wilcke W, Homeier J, Lehmann J, Engels C. Plant growth along the altitudinal gradient-role of plant nutritional status, fine root activity, and soil properties. Gradients in a Tropical Mountain Ecosystem of Ecuador, 2008, 259-266. |
| [47] |
Zhang J H, He N P, Liu C C, Xu L, Chen Z, Li Y, Wang R M, Yu G R, Sun W, Xiao C W, Chen H Y H, Reich P B. Variation and evolution of C ∶ N ratio among different organs enable plants to adapt to N-limited environments. Global Change Biology, 2020, 26(4): 2534-2543. |
| [48] |
He J S, Fang J Y, Wang Z H, Guo D L, Flynn D F B, Geng Z. Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China. Oecologia, 2006, 149(2): 115-122. |
| [49] |
Klimes L. Life-forms and clonality of vascular plants along an altitudinal gradient in E Ladakh (NW Himalayas). Basic and Applied Ecology, 2003, 4(4): 317-328. |
| [50] |
Han W X, Fang J Y, Guo D L, Zhang Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytologist, 2005, 168(2): 377-385. |