 2024, Vol. 44
2024, Vol. 44文章信息
- 元晓春, 曾泉鑫, 张晓晴, 孙浩, 柏欣宇, 陈琳娜, 李文周, 陈亚婷, 陈岳民, 林开淼
- YUAN Xiaochun, ZENG Quanxin, ZHANG Xiaoqing, SUN Hao, BAI Xinyu, CHEN Linna, LI Wenzhou, CHEN Yating, CHEN Yuemin, LIN Kaimiao
- 3年高氮添加对黄山松土壤真菌网络稳定性的影响
- The impact of reduced by three-year high nitrogen addition on stability of soil fungal network of Pinus taiwanensis
- 生态学报. 2024, 44(8): 3372-3381
- Acta Ecologica Sinica. 2024, 44(8): 3372-3381
- http://dx.doi.org/10.20103/j.stxb.202309272099
-
文章历史
- 收稿日期: 2023-09-27
- 网络出版日期: 2024-02-25






2. 福建师范大学地理科学学院, 福州 350007;
3. 福建戴云山国家级自然保护区管理局, 德化 362500
2. School of Geographical Science, Fujian Normal University, Fuzhou 350007, China;
3. Daiyun Mountain National Nature Reserve Administration Bureau, Dehua 362500, China
土壤真菌作为植物和动物的分解者、共生体或病原体, 在维持土壤健康和生态系统功能(如氮和碳循环)中发挥着关键的作用[1—2]。氮沉降等环境变化会通过改变真菌多样性和群落组成[3—4]、养分获取策略[5]及真菌-宿主相互作用[6—7]等影响生态系统的结构和功能。因此, 了解真菌群落稳定性对缓解其对生态系统服务功能的潜在影响至关重要。先前的研究表明, 诸多环境胁迫(如增温、干旱)会影响真菌网络稳定性[8—9]。相比于细菌, 土壤真菌网络在干旱等环境胁迫下更稳定[9]。然而, 真菌作为一种嗜氮群体, 其群落的稳定性是否受氮沉降的影响尚不清楚。
网络分析是了解氮沉降等环境变化如何影响真菌群落稳定性的一个有效途径。正、负相互作用和/或微生物生态位的相似性的差异将导致群落不稳定[10]。基于此, Hernandez等[11]首次提出内聚力用以预测微生物网络的稳定性。相比于其他稳定性指标(例如, 复杂性等), 该指标能量化物种之间的连通性使预测结果更加准确[10], 该指标已被成功应用于土壤和水体等多个领域[11—13]。通常, 正内聚力表示微生物物种之间的正相互作用和/或高度生态位重叠, 而负内聚力则表示负相互作用和/或不同的生态位。负内聚力和正内聚力的相对比例(负∶正内聚力)与群落的稳定性存在紧密关联[10—11]。当一个群落以负内聚力为主(高的负∶正内聚力)代表群落更稳定, 即群落(1)在环境扰动中变化更有限;或(2)在扰动后更有可能恢复到原有的状态[14], 反之亦然[11]。当存在正相互作用的物种数量减少, 依赖正反馈回路的其他物种会受到负面影响[14]。总之, 尽管真菌群落稳定性可以被很好地表征, 然而, 目前关于氮沉降如何影响土壤微生物内聚力, 从而改变微生物群落稳定性的认识仍然十分有限。
先前研究表明, 氮添加下真菌群落受土壤氮有效性的直接影响[15], 过高的土壤氮饱和程度将对真菌群落稳定性产生负面影响。土壤和叶片δ15N及氮富集指数通常可以用以表征氮饱和程度[16—17]。Egerton-Warburton等[16]通过土壤δ15N的追踪发现氮添加下随着土壤δ15N含量增加, 真菌群落物种相对丰度及丰富性降低, 而较低的真菌多样性通常难以维持其群落的稳定性[11]。此外, 网络模块作为一种分类单元, 不仅可以反映模块中物种之间的空间划分[18]或相似的栖息地偏好[19], 也可以影响群落稳定性[8]。通常同一个模块的微生物物种具有以下特征:1)彼此相互作用更大;2)有独特生态位[11]。高度模块化由于模块与模块之间相对独立而能保持群落的稳定。相反, 低模块化更容易将一个模块内受干扰物种的丰度波动传播到其他模块中[11]。例如, 磷钾肥添加降低真菌网络模块性[20], 这可能对真菌群落稳定性产生潜在威胁。
由于化石燃料燃烧、尾气排放等人类活动, 我国东南部氮沉降量依旧处于高位水平[21]。尽管森林生态系统具有巨大的生态调节功能, 全球11%的天然林所在地的氮沉降水平已超过“临界负荷”的氮阈值[22]。戴云山自然保护区位于我国东南区福建省内。在这里森林树种繁多, 黄山松(Pinus taiwanensis)作为高海拔优势树种, 在气候恶劣和土壤贫瘠的环境下具有其他针叶林无法企及的生态演替功能[23]。前期调查发现黄山松较该保护区内其他树种(如罗浮栲)具有更低的土壤氮含量, 且其土壤微生物对氮添加的响应更为敏感[24—25]。为此, 本研究选取黄山松林为研究对象, 结合高通量测序和网络分析, 对连续三年氮添加下的土壤真菌群落进行分析。本研究的主要目的是探究土壤真菌多样性及其群落稳定性对氮添加如何响应。我们假设, 氮添加下随着土壤真菌多样性和网络模块性的降低, 真菌群落变得更不稳定。
1 材料与方法 1.1 研究区概况研究区位于福建省泉州市德化县戴云山国家自然保护区(25°38′07″—25°43′40″N, 118°05′22″—118°20′15″E), 该地年均温为15.6—19.5℃, 年均降水为1700—2000 mm, 属于典型的亚热带海洋性季风气候[25]。土壤类型为山地红壤, 主要林下植被包括箬竹(Indocalamus tessellatus)、山茶花(Camellia flower)、毛稔(Melastoma sanguineum)等[26]。
1.2 试验设计与取样综合参考福建沿海区域(30—40 kg N hm2 a-1)[20]和附近城市(厦门市郊区, 36.34 kg N hm2 a-1)[27]的氮沉降量, 试验以40 kg N hm2 a-1为氮添加基数。按照随机区组法, 在黄山松林设置三种氮添加处理:对照(CT, 0 kg N hm2 a-1)、低氮(LN, 40 kg N hm2 a-1)、高氮(HN, 80 kg N hm2 a-1), 每个处理设置4个重复。试验区从2018年5月开始通过添加尿素模拟野外氮沉降, 氮添加时间为生长季(3—9月), 每月进行一次。具体样地设计及氮添加方法参照先前的方法[25]。于2019—2021年定期在生长季(5月)采集两个土层的土壤样品。即, 按照五点取样法, 在每个样方的0—10 cm和10—20 cm土层各取5个土芯, 同一样方相同土层的土壤样品在去除石块和根系之后混合成一个样品。将土壤样品装入塑料自封袋用冰袋保鲜带回实验室后, 将土壤样本分为两部分, 一部分过2 mm筛后存于-20℃冰柜中, 用于提取DNA和高通量测序, 另一部分用于测定土壤物理化学性质和同位素δ15N。从每个样地选取三棵长势接近的黄山松, 在树冠中上部采集20片健康叶片混合在一起。叶片采集后用干冰冷藏运往实验室。叶片样本在105℃下杀青5 min分钟, 在65℃下烘干直至达到恒重。将干燥样品用球磨仪进行研磨用于后续试验。
1.3 测定项目与方法土壤pH(水土比为2.5∶1)采用玻璃电极pH计(Starter 300, Ohaus, 美国)测定[25]。土壤含水率(SM)用烘干法测定。土壤总碳(TC)和总氮(TN)采用碳氮元素分析仪(Elementar Vario EL III, Elementar, 德国)测定。矿质氮(MN)用2 mol/L的氯化钾浸提[28], 使用连续流动分析仪(Skalar san++, Skalar, 荷兰)测定提取液中的铵态氮(NH4+-N)和硝态氮(NO3--N)浓度。土壤和叶片样本的δ15N值用同位素质谱仪(Thermo scientific MAT253, 德国)测定。氮富集指数(NEI)通过如下公式计算[17]:
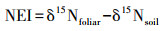
|
其中δ15Nfoliar和δ15Nsoil分别代表黄山松叶片和土壤(0—10 cm)的δ15N值。
取0.5 g鲜土, 参照DNA提取试剂盒(Omega Bio-tek, Norcross, 佐治亚州,美国)试剂盒说明书对所有样品的DNA进行提取, 并用1%琼脂糖凝胶电泳检测DNA质量, 质检合格的于-20℃保存, 交给测序公司(奥维森基因科技有限公司, 北京)进行高通量测序。利用特异性引物ITS1(CTTGGTCATTTAGAGGAAGTAA)和ITS2(TGCGTTCTTCATCGATGC)对真菌进行聚合酶链式反应(PCR)扩增。具体参数参照Cui等[29]的方法。
根据Herren等[10]的方法, 计算了每个样本(j)的正、负内聚力值, 计算公式如下:
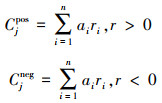
|
其中ai是样本j中物种i的丰度;ri, r > 0和ri, r<0分别为正连通性和负连通性。在每个网络中, 物种i的正连通性(ri,r > 0)和负连通性(ri, r<0)被计算为其与网络中所有其他真菌物种的显著正相关或负相关系数的平均值。负内聚力(Cjneg)和正内聚力(Cjpos)的范围分别为-1到0和0到1, 绝对值越高表示内聚力越大。总内聚力是一种网络复杂性的度量, 计算为正内聚力和负内聚力的绝对值之和。负:正内聚力的绝对值用于表征每个真菌网络的稳定性[11]。
1.4 数据处理采用SPSS 26.0和R 4.1.0软件对数据进行统计分析。通过重复测量方差分析探究氮添加、土层和时间因素及其交互作用对各指标影响的主效应。通过Duncan多重比较(α=0.05)检验指标在同一取样时间、不同处理间的差异显著性。通过vegan包计算土壤真菌多样性, Chao1和Shannon指数分别表征真菌丰富度和多样性。为构建真菌的共现性网络, 选择相对丰度大于0.01%的真菌物种分类的操作分类单元(OTU)[30]。通过vegan包进行主坐标(PCoA)分析, 并通过Anosim检验真菌群落结构的相似性。通过WGCNA包Spearman相关矩阵推断出初始网络关系。网络中的节点代表特定OTU, 连接不同节点的边代表OTU之间的相关关系。为减少假阳性结果, 使用Beniamini-Hochberg方法(FDR)校正所有的P值[31]。选择Spearman相关系数大于0.60且经FDR校正后P<0.01的相关关系来构建共现网络。用Gephi的0.9.2版本对网络进行了可视化。然后, 通过默认参数确定了共现网络中存在显著关联的OTUs, 构建微生物网络模块(模块度高于0.4表明网络具有模块化结构)[32]。柱状图通过Origin 2018软件绘制, 图中数据为平均值(标准误), n=4。
2 结果与分析 2.1 氮添加下土壤的物理化学性质和同位素δ15N氮添加3年内, 土壤pH和SM均无显著的变化(表 1)。0—10 cm土层土壤TC和TN在第3年氮添加下显著增加, 尤其在高氮添加下(P<0.05, 表 1)。与土壤MN相同, δ15Nfoliar在第2—3年氮添加下显著增加(P<0.05), 而δ15Nsoil和NEI仅在高氮添加下显著增加(P<0.05, 表 1)。
| 指标 Indicators |
2019年 | 2020年 | 2021年 | ||||||
| 0—10 cm | 10—20 cm | 0—10 cm | 10—20 cm | 0—10 cm | 10—20 cm | ||||
| pH | LN | -0.02 | 0.06 | 0.09 | 0.1 | 0.01 | 0.05 | ||
| HN | -0.01 | 0.03 | 0.14 | 0.08 | 0.05 | 0.07 | |||
| SM/% | LN | -0.08 | -0.02 | -0.03 | 0.01 | 0.05 | 0.01 | ||
| HN | -0.14 | -0.02 | -0.04 | 0.01 | -0.02 | -0.03 | |||
| TC/(g/kg) | LN | 0.26 | 1.02 | -4.06 | -1.87 | 11.43*** | 0.57 | ||
| HN | -8.71 | 4.73 | -5.55 | -2.55 | 12.25*** | -1.22 | |||
| TN/(g/kg) | LN | -0.31 | -0.19 | -0.24 | -0.01 | 0.2 | 0.28 | ||
| HN | -0.95* | 0.4 | -0.22 | -0.08 | 0.85** | 0.21 | |||
| MN/(mg/kg) | LN | -24.76** | -13.18 | 7.02*** | 8.77*** | 6.45** | 2.68 | ||
| HN | -9.25 | -0.48 | 9.39*** | 10.13*** | 18.43*** | 4.3 | |||
| δ15Nfoliar/‰ | LN | 1.15 | — | 2.40** | — | 2.68* | — | ||
| HN | 1.62 | — | 5.79*** | — | 4.89** | — | |||
| δ15Nsoil/‰ | LN | 0.19 | — | 0.919 | — | 1.33 | — | ||
| HN | -0.02 | — | 1.75** | — | 1.48 | — | |||
| NEI/‰ | LN | 0.97 | — | 1.49 | — | 1.35 | — | ||
| HN | 1.64 | — | 4.04*** | — | 3.41** | — | |||
| pH: 酸碱度pH value; SM: 土壤含水率Soil moisture; TC: 总碳Total carbon; TN: 总氮Total nitrogen; MN: 矿质氮Mineral nitrogen; δ15Nsoil: 土壤δ15N, Soil δ15N; δ15Nfoliar: 叶片δ15N, Foliar δ15N; NEI: 氮富集指数Nitrogen enrichment index; LN: 低氮Low nitrogen; HN: 高氮High nitrogen; 表中数据为氮添加与对照组之间的均值差(均值差=低氮或高氮处理-对照), n=4. 星号代表低氮或高氮处理相比于对照存在显著差异; 0—10 cm和10—20 cm代表土层深度; *: P<0.05; **: P<0.01; ***: P<0.001; —: 缺失值; | |||||||||
重复测量方差分析表明, 氮添加的主效应及其与土层的交互作用对土壤真菌Chao1指数没有显著影响(表 2)。氮添加与氮添加时间以及氮添加、土层与氮添加时间的交互作用对土壤真菌的Chao1和Shannon指数存在显著影响(P<0.05, 表 2)。在0—10 cm土层, 第1—2年氮添加降低或不影响真菌Chao1和Shannon指数, 但第3年氮添加显著提高真菌Chao1和Shannon指数, 尤其在低氮处理下(P<0.05, 图 1)。在10—20 cm土层, 1—3年内氮添加对真菌Chao1指数没有显著影响, 但第3年高氮添加显著提高真菌Shannon指数(P<0.05, 图 1)。土层显著影响真菌的Chao1和Shannon指数(P<0.05, 表 2), 总体上, 0—10 cm土层真菌的Chao1和Shannon指数高于10—20 cm土层(图 1)。此外, PCoA分析表明, 第1—2年内氮添加对黄山松林土壤真菌群落β多样性无显著影响, 但第3年氮添加下真菌群落存在明显的聚类现象(P<0.01), 且不同土层真菌群落β多样性存在显著差异(P<0.001, 图 2)。
| 变异来源 Source of variation |
Chao1指数 Chao1 index |
Shannon指数 Shannon index |
变异来源 Source of variation |
Chao1指数 Chao1 index |
Shannon指数 Shannon index |
|
| N | 0.310 | 2.31 | T × N | 7.49** | 4.81* | |
| D | 84.05*** | 4.20* | T × D | 24.84*** | 0.42 | |
| D × N | 0.59 | 0.49 | T × N ×D | 15.55*** | 7.50** | |
| T | 90.65*** | 0.13 | ||||
| N, D和T分别表示氮添加, 土层和时间; *: P<0.05; **: P<0.01; ***: P<0.001. | ||||||

|
| 图 1 氮添加对土壤真菌α多样性的影响 Fig. 1 Effect of nitrogen addition on soil fungal alpha diversity CT: 对照Control; LN: 低氮Low nitrogen; HN: 高氮High nitrogen; 不同小写字母表示同一取样时间不同处理间差异显著 |

|
| 图 2 氮添加对土壤真菌β多样性的影响 Fig. 2 Effect of nitrogen addition on beta diversity of soil fungi N和D表示氮添加和土层对真菌β多样性的影响; **: P<0.01; ***: P<0.001; n.s.: P>0.05. PCoA: 主坐标分析Principal co-ordinates analysis |
网络分析显示, 氮添加不改变真菌共现网络中的优势物种(门水平)的占比, 但改变了真菌网络中的聚类效果(图 3)。通过网络拓扑属性的分析发现, 与对照相比, 氮添加增加了土壤真菌网络的节点和边的数量, 但平均聚类系数显著降低(图 3)。无论是低氮还是高氮添加, 真菌网络中的正连接的比例随着氮添加水平增加而提高, 而负连接的比例则相反(图 3)。此外, 真菌网络主要分为6大模块, 模块度随着氮添加水平增加而减少(图 3)。

|
| 图 3 不同氮添加水平下真菌群落的共现网络 Fig. 3 Co-occurrence network patterns of fungal communities under different nitrogen addition levels 节点根据门类和模块进行着色; 节点大小代表操作分类单元(OTU)的度, 网络中的连线表示节点(OTU)间存在显著联系 |
氮添加的主效应及其与氮添加时间的交互效应显著影响土壤真菌网络的内聚力(表 3)。与氮添加对土壤真菌多样性和群落结构的影响模式一致, 第3年氮添加对土壤真菌内聚力的影响程度大于第1—2年氮添加。总体上, 在两个土层中都观察到真菌的负内聚力的绝对值和总内聚力均在低氮添加下最大, 在高氮添加下最小(图 4)。在0—10 cm土层, 第3年氮添加显著提高真菌正内聚力和总内聚力。负内聚力的绝对值在不同氮添加水平有差异, 低氮增加真菌负内聚力的绝对值, 而高氮对负内聚力的绝对值无显著影响(图 4)。与对照相比, 第3年高氮显著降低了两个土层的真菌负:正内聚力的绝对值, 而低氮提高或不改变两者比值的绝对值。总体上, 在0—10 cm土层真菌负内聚力及负:正内聚力的绝对值高于10—20 cm土层(图 4)。
| 变异来源 Source of variation |
|负内聚力| | Negative cohesion| |
正内聚力 Positive cohesion |
总内聚力 Total cohesion |
|负内聚力: 正内聚力| | Negative cohesion: Positive cohesion| |
| N | 23.53*** | 4.65* | 10.08** | 27.99*** |
| D | 8.57** | 0.93 | 3.12 | 8.74** |
| D × N | 0.75 | 1.41 | 0.91 | 1.50 |
| T | 33.36*** | 9.55** | 11.73** | 19.61*** |
| T × N | 3.91* | 11.73** | 5.88* | 10.79** |
| T × D | 4.81* | 5.08* | 4.98* | 0.50 |
| T × N ×D | 0.41 | 8.27** | 7.42** | 5.66* |

|
| 图 4 氮添加对真菌内聚力的影响 Fig. 4 Effect of nitrogen addition on soil fungal cohesion |
氮添加与氮添加时间的交互作用对土壤真菌的Chao1和Shannon指数存在显著影响, 这说明真菌多样性对氮添加的响应随年度添加时间存在明显差异。第1—2年氮添加下真菌群落多样性没有变化, 直到第3年氮添加真菌多样性存在显著变化(图 1和图 2)。这说明真菌群落对氮添加初期的响应并不灵敏。先前在沙漠灌木林的研究同样发现真菌群落多样性在两年氮添加下没有显著的变化[33]。第3年氮添加0—10 cm土层的真菌多样性发生了转变, 且这种响应是积极的, 尤其在低氮添加下(图 1), 这与多数研究结果相反[34—36]。由于氮添加会诱导土壤酸性阳离子(H+和Al3+)的增加[37]和pH降低[38], 多项全球尺度的meta分析也曾报道氮添加对微生物α多样性存在很强的负效应, 尤其在长期氮添加下[38—39]。但本研究中, 仅发现土壤有效氮显著增加, 土壤pH并未发生显著变化(表 1)。通常, 氮有效性的增加会改善微生物养分利用策略[40—41], 氮资源的积累有利于真菌的生长, 并促进其多样性增加。而在氮添加初期(1—2年内)真菌多样性未发生显著变化, 可能原因在于真菌群落的底物偏好。相对细菌而言, 生长缓慢的真菌属于贫营养类群, 偏好分解难分解的有机质[42], 因此其对易分解物质的获取不如细菌。基于以上, 推测氮添加后期真菌多样性的增加更多取决于氮有效性的增加而非土壤酸化。此外, 真菌多样性的土层效应十分明显, 10—20 cm土层真菌的α多样性整体低于0—10 cm土层(表 1和图 1), 这与多数研究结果一致[43—44], 由于随土壤深度增加, 土壤资源可用性降低[43]。
3.2 氮添加对真菌网络稳定性的影响平均聚类系数和负:正内聚力的绝对值是网络复杂性和稳定性的重要指标[11]。本研究发现高氮添加下, 真菌网络平均聚类系数远低于对照和低氮(图 3)。且相比于对照和低氮添加, 第3年高氮添加显著降低了两个土层的负:正内聚力的绝对值(图 4)。以上说明高氮添加同时降低了真菌网络的复杂性和稳定性, 这一发现符合复杂性和稳定性存在协同性的生态理论[8]。
关于影响微生物网络不稳定的因素, 先前研究认为微生物多样性降低将破坏群落稳定性[11]。但是, 本研究结果并不支持这个观点, 因为高氮添加提高了土壤真菌多样性(图 1)。因此, 推测高氮添加下土壤真菌网络稳定性降低的原因可能在于:1)氮富集程度增加会导致真菌群落趋于不稳定。先前研究表明真菌群落会直接受到氮有效性的影响[15]。本研究中发现第三年高氮添加下矿质氮含量和氮富集指数显著增加(表 1)。氮富集指数是氮饱和的潜在指标[17, 45]。生态系统氮饱和可能通过影响土壤氮循环[46], 加剧磷限制[47]等过程影响特定微生物的生态位, 进而改变真菌群落的稳定性。而低氮添加对氮富集指数无显著影响(表 1), 这可能是导致真菌群落网络稳定性在低氮处理下不受影响的主要原因。2)高正连接比例和低模块性加剧了高氮添加对真菌网络稳定性的负面影响。通常而言, 微生物网络的不稳定性主要围绕正连接展开[11]。在正连接中, 彼此相互维持的微生物胜过其他微生物, 其中任何一个成员受到扰动都将迅速传播到整个网络[14]。相反, 当负连接中的成员受到的扰动时, 将抑制其他成员受到的扰动[48]。因此, 高氮添加下更高的正连接比例(图 3)可能是真菌群落不稳定的诱发因子。此外, Hernandez等[11]的研究曾发现高环境胁迫下微生物具有更低的模块性。类似地, 本研究中发现更低的模块性存在于高氮添加处理下(图 4)。高度模块化通常由于模块与模块之间相对独立而保持群落的稳定。相反, 低模块化更容易将一个模块内受干扰物种的丰度波动传播到其他模块中[11]。因此, 相比于对照和低氮, 高氮添加下真菌物种跨模块的关联可能更为普遍。这将导致一个模块中影响真菌物种的环境扰动传播到其他模块的可能性更大, 最终影响真菌整个群落的稳定性。综合而言, 尽管本研究提出的假设并非全部成立, 但研究发现随着平均聚类系数和模块性的降低, 高氮添加下形成了一个以正连接为主导的不稳定的真菌网络。
4 结论本研究主要探讨了连续三年氮添加对黄山松土壤真菌多样性和网络稳定性的影响。研究表明, 氮添加下真菌多样性及群落稳定性存在明显的年际变化, 其响应主要集中的氮添加后第3年。第3年氮添加显著增加了真菌多样性, 其主要的影响因素可能是氮的有效性而非土壤pH。相比于低氮, 第3年高氮添加降低了真菌网络稳定性, 这与较高的氮富集程度和网络中较低的正连接比例及模块度有关。综合而言, 本研究表明高浓度氮输入会破坏土壤真菌群落的稳定性, 这可能不利于生态系统功能的维持。
| [1] |
Frąc M, Hannula S E, Bełka M, Jędryczka M. Fungal biodiversity and their role in soil health. Frontiers in Microbiology, 2018, 9: 707. DOI:10.3389/fmicb.2018.00707 |
| [2] |
张晶, 张惠文, 李新宇, 张成刚. 土壤真菌多样性及分子生态学研究进展. 应用生态学报, 2004, 15(10): 1958-1962. DOI:10.3321/j.issn:1001-9332.2004.10.051 |
| [3] |
Zhou J, Jiang X, Zhou B K, Zhao B S, Ma M C, Guan D W, Li J, Chen S F, Cao F M, Shen D L, Qin J. Thirty four years of nitrogen fertilization decreases fungal diversity and alters fungal community composition in black soil in northeast China. Soil Biology and Biochemistry, 2016, 95: 135-143. DOI:10.1016/j.soilbio.2015.12.012 |
| [4] |
赵爱花, 刘蕾, 付伟, 武慧, 陈保冬. 施氮对森林生态系统AM真菌群落组成及多样性的影响. 生态学报, 2020, 40(21): 7576-7587. |
| [5] |
Li L, McCormack M L, Chen F S, Wang H M, Ma Z Q, Guo D L. Different responses of absorptive roots and arbuscular mycorrhizal fungi to fertilization provide diverse nutrient acquisition strategies in Chinese fir. Forest Ecology and Management, 2019, 433: 64-72. DOI:10.1016/j.foreco.2018.10.055 |
| [6] |
Carrara J E, Walter C A, Hawkins J S, Peterjohn W T, Averill C, Brzostek E R. Interactions among plants, bacteria, and fungi reduce extracellular enzyme activities under long-term N fertilization. Global Change Biology, 2018, 24(6): 2721-2734. DOI:10.1111/gcb.14081 |
| [7] |
孙鹏飞, 程瑞梅, 肖文发, 沈雅飞, 曾立雄, 王丽君, 陈天, 张萌, 邢红爽. 模拟氮沉降和接种外生菌根真菌对马尾松(Pinus massoniana)幼苗营养元素的影响. 生态学报, 2023, 43(17): 7263-7273. |
| [8] |
Yuan M M, Guo X, Wu L W, Zhang Y, Xiao N J, Ning D L, Shi Z, Zhou X S, Wu L Y, Yang Y F, Tiedje J M, Zhou J Z. Climate warming enhances microbial network complexity and stability. Nature Climate Change, 2021, 11(4): 343. DOI:10.1038/s41558-021-00989-9 |
| [9] |
de Vries F T, Griffiths R I, Bailey M, Craig H, Girlanda M, Gweon H S, Hallin S, Kaisermann A, Keith A M, Kretzschmar M, Lemanceau P, Lumini E, Mason K E, Oliver A, Ostle N, Prosser J I, Thion C, Thomson B, Bardgett R D. Soil bacterial networks are less stable under drought than fungal networks. Nature Communications, 2018, 9: 3033. DOI:10.1038/s41467-018-05516-7 |
| [10] |
Herren C M, McMahon K D. Cohesion: a method for quantifying the connectivity of microbial communities. The ISME Journal, 2017, 11(11): 2426-2438. DOI:10.1038/ismej.2017.91 |
| [11] |
Hernandez D J, David A S, Menges E S, Searcy C A, Afkhami M E. Environmental stress destabilizes microbial networks. The ISME Journal, 2021, 15(6): 1722-1734. DOI:10.1038/s41396-020-00882-x |
| [12] |
Li F L, Zhang Y, Altermatt F, Yang J H, Zhang X W. Destabilizing effects of environmental stressors on aquatic communities and interaction networks across a major river basin. Environmental Science and Technology, 2023, 57(20): 7828-7839. DOI:10.1021/acs.est.3c00456 |
| [13] |
Danczak R E, Johnston M D, Kenah C, Slattery M, Wilkins M J. Microbial community cohesion mediates community turnover in unperturbed aquifers. mSystems, 2018, 3(4): e00066-18. |
| [14] |
Coyte K Z, Schluter J, Foster K R. The ecology of the microbiome: networks, competition, and stability. Science, 2015, 350(6261): 663-666. DOI:10.1126/science.aad2602 |
| [15] |
Huang S L, Yu C Q, Fu G, Sun W, Li S W, Han F S, Xiao J Y. Effects of short-term nitrogen addition on soil fungal community increase with nitrogen addition rate in an alpine steppe at the source of Brahmaputra. Microorganisms, 2023, 11(8): 1880. DOI:10.3390/microorganisms11081880 |
| [16] |
Egerton-Warburton L M, Graham R C, Allen E B, Allen M F. Reconstruction of the historical changes in mycorrhizal fungal communities under anthropogenic nitrogen deposition. Proceedings of the Royal Society B-Biological Sciences, 2001, 268: 2479-2484. DOI:10.1098/rspb.2001.1844 |
| [17] |
刘瑞强, 何宗明, 黄志群, 范少辉, 万晓华, 杜婷, 苏慧琴. 采伐剩余物管理对杉木人工林土壤和叶片δ15N的长期效应. 林业科学, 2014, 50(12): 7-13. |
| [18] |
Liu S W, Yu H, Yu Y H, Huang J, Zhou Z Y, Zeng J X, Chen P B, Xiao F S, He Z L, Yan Q Y. Ecological stability of microbial communities in Lake Donghu regulated by keystone taxa. Ecological Indicators, 2022, 136: 108695. DOI:10.1016/j.ecolind.2022.108695 |
| [19] |
Shukla R, Bhat A. Patterns and drivers of species co-occurrence networks in a tropical stream fish metacommunity. Hydrobiologia, 2022, 849(12): 2797-2811. DOI:10.1007/s10750-022-04894-w |
| [20] |
Tang S, Zhou J J, Pan W K, Sun T, Liu M J, Tang R, Li Z J, Ma Q X, Wu L H. Effects of combined application of nitrogen, phosphorus, and potassium fertilizers on tea (Camellia sinensis) growth and fungal community. Applied Soil Ecology, 2023, 181: 104661. DOI:10.1016/j.apsoil.2022.104661 |
| [21] |
Wen Z, Xu W, Li Q, Han M J, Tang A H, Zhang Y, Luo X S, Shen J L, Wang W, Li K H, Pan Y P, Zhang L, Li W Q, Collett J L, Zhong B Q, Wang X M, Goulding K, Zhang F S, Liu X J. Changes of nitrogen deposition in China from 1980 to 2018. Environment International, 2020, 144: 106022. DOI:10.1016/j.envint.2020.106022 |
| [22] |
Wang R, Goll D, Balkanski Y, Hauglustaine D, Boucher O, Ciais P, Janssens I, Penuelas J, Guenet B, Sardans J, Bopp L, Vuichard N, Zhou F, Li B G, Piao S L, Peng S S, Huang Y, Tao S. Global forest carbon uptake due to nitrogen and phosphorus deposition from 1850 to 2100. Global Change Biology, 2017, 23(11): 4854-4872. DOI:10.1111/gcb.13766 |
| [23] |
苏松锦, 刘金福, 兰思仁, 洪伟, 李文周. 黄山松研究综述(1960—2014)及其知识图谱分析. 福建农林大学学报(自然科学版), 2015, 44(5): 478-486. |
| [24] |
戴辉, 曾泉鑫, 周嘉聪, 彭园珍, 孙雪琦, 陈静琪, 陈文伟, 陈岳民. 罗浮栲林土壤微生物碳利用效率对短期氮添加的响应. 应用生态学报, 2022, 33(10): 2611-2618. |
| [25] |
元晓春, 崔琚琰, 林开淼, 周嘉聪, 曾泉鑫, 谢欢, 刘苑苑, 徐建国, 陈岳民. 黄山松土壤可溶性有机质对氮添加的响应及其与细菌群落的关联. 应用生态学报, 2021, 32(11): 4085-4094. |
| [26] |
张晓晴, 曾泉鑫, 元晓春, 万晓华, 崔琚琰, 李文周, 林惠瑛, 谢欢, 陈文伟, 吴君梅, 陈岳民. 氮添加诱导的磷限制改变了亚热带黄山松林土壤微生物群落结构. 应用生态学报, 2023, 34(1): 203-212. |
| [27] |
蔡美君, 张剑辉, 朱恒, 吴水平. 厦门郊区大气主要含氮化合物组成特征研究. 环境科学与技术, 2019, 42(S2): 238-247. |
| [28] |
苏涛, 司美茹, 王朝辉, 李生秀. 土壤矿质氮分析方法的影响因素研究. 农业环境科学学报, 2005, 24(6): 1238-1242. DOI:10.3321/j.issn:1672-2043.2005.06.040 |
| [29] |
Cui J Y, Yuan X C, Zhang Q F, Zhou J C, Lin K M, Xu J G, Zeng Y Z, Wu Y, Cheng L, Zeng Q X, Mei K C, Chen Y M. Nutrient availability is a dominant predictor of soil bacterial and fungal community composition after nitrogen addition in subtropical acidic forests. PLoS ONE, 2021, 16(2): e0246263. DOI:10.1371/journal.pone.0246263 |
| [30] |
Feng J, Zeng X M, Zhang Q G, Zhou X Q, Liu Y R, Huang Q Y. Soil microbial trait-based strategies drive metabolic efficiency along an altitude gradient. ISME Communications, 2021, 1(1): 71. DOI:10.1038/s43705-021-00076-2 |
| [31] |
Benjamini Y, Krieger A M, Yekutieli D. Adaptive linear step-up procedures that control the false discovery rate. Biometrika, 2006, 93(3): 491-507. DOI:10.1093/biomet/93.3.491 |
| [32] |
Shi S J, Nuccio E E, Shi Z J, Shi Z J, He Z L, Firestone M K. The interconnected rhizosphere: high network complexity dominates rhizosphere assemblages. Ecology Letters, 2016, 19(8): 926-936. DOI:10.1111/ele.12630 |
| [33] |
She W W, Bai Y X, Zhang Y Q, Qin S G, Feng W, Sun Y F, Zheng J, Wu B. Resource availability drives responses of soil microbial communities to short-term precipitation and nitrogen addition in a desert shrubland. Frontiers in Microbiology, 2018, 9: 186. DOI:10.3389/fmicb.2018.00186 |
| [34] |
Yang Y, Li T, Wang Y Q, Cheng H, Chang S X, Liang C, An S S. Negative effects of multiple global change factors on soil microbial diversity. Soil Biology and Biochemistry, 2021, 156: 108229. DOI:10.1016/j.soilbio.2021.108229 |
| [35] |
Morrison E W, Frey S D, Sadowsky J J, van Diepen L T A, Thomas W K, Pringle A. Chronic nitrogen additions fundamentally restructure the soil fungal community in a temperate forest. Fungal Ecology, 2016, 23: 48-57. DOI:10.1016/j.funeco.2016.05.011 |
| [36] |
陈松鹤, 向晓玲, 雷芳, 邹乔生, 艾代龙, 郑亭, 黄秀兰, 樊高琼. 秸秆覆盖配施氮肥根际土真菌群落及其与小麦产量的关系. 生态学报, 2022, 42(21): 8751-8761. |
| [37] |
Treseder K K. Nitrogen additions and microbial biomass: a meta-analysis of ecosystem studies. Ecology Letters, 2008, 11(10): 1111-1120. DOI:10.1111/j.1461-0248.2008.01230.x |
| [38] |
Zhou Z H, Wang C K, Luo Y Q. Meta-analysis of the impacts of global change factors on soil microbial diversity and functionality. Nature Communications, 2020, 11: 3072. DOI:10.1038/s41467-020-16881-7 |
| [39] |
Dang P F, Li C F, Lu C, Zhang M M, Huang T T, Wan C X, Wang H Y, Chen Y L, Qin X L, Liao Y C, Siddique K H M. Effect of fertilizer management on the soil bacterial community in agroecosystems across the globe. Agriculture, Ecosystems and Environment, 2022, 326: 107795. DOI:10.1016/j.agee.2021.107795 |
| [40] |
Tahovská K, Choma M, Kaštovská E, Oulehle F, Bárta J, Šantrčková H, Moldan F. Positive response of soil microbes to long-term nitrogen input in spruce forest: results from Gårdsjön whole-catchment N-addition experiment. Soil Biology and Biochemistry, 2020, 143: 107732. DOI:10.1016/j.soilbio.2020.107732 |
| [41] |
Sun M Y, Li M C, Zhou Y Q, Liu J, Shi W C, Wu X L, Xie B H, Deng Y, Gao Z. Nitrogen deposition enhances the deterministic process of the prokaryotic community and increases the complexity of the microbial co-network in coastal wetlands. Science of the Total Environment, 2023, 856: 158939. DOI:10.1016/j.scitotenv.2022.158939 |
| [42] |
Blagodatskaya E V, Blagodatsky S A, Anderson T H, Kuzyakov Y. Priming effects in Chernozem induced by glucose and N in relation to microbial growth strategies. Applied Soil Ecology, 2007, 37(1-2): 95-105. DOI:10.1016/j.apsoil.2007.05.002 |
| [43] |
Guasconi D, Juhanson J, Clemmensen K E, Cousins S A O, Hugelius G, Manzoni S, Roth N, Fransson P. Vegetation, topography, and soil depth drive microbial community structure in two Swedish grasslands. FEMS Microbiology Ecology, 2023, 99(8): 1-13. |
| [44] |
Dang Q L, Tan W B, Zhao X Y, Li D, Li Y P, Yang T X, Li R F, Zu G F, Xi B D. Linking the response of soil microbial community structure in soils to long-term wastewater irrigation and soil depth. Science of the Total Environment, 2019, 688: 26-36. DOI:10.1016/j.scitotenv.2019.06.138 |
| [45] |
Fang H J, Yu G R, Cheng S L, Zhu T H, Zheng J J, Mo J M, Yan J H, Luo Y Q. Nitrogen-15 signals of leaf-litter-soil continuum as a possible indicator of ecosystem nitrogen saturation by forest succession and N loads. Biogeochemistry, 2011, 102(1-3): 251-263. DOI:10.1007/s10533-010-9438-1 |
| [46] |
陈洁, 骆土寿, 周璋, 许涵, 陈德祥, 李意德. 氮沉降对热带亚热带森林土壤氮循环微生物过程的影响研究进展. 生态学报, 2020, 40(23): 8528-8538. |
| [47] |
Fleischer K, Dolman A J, van der Molen M K, Rebel K T, Erisman J W, Wassen M J, Pak B, Lu X J, Rammig A, Wang Y P. Nitrogen deposition maintains a positive effect on terrestrial carbon sequestration in the 21st century despite growing phosphorus limitation at regional scales. Global Biogeochemical Cycles, 2019, 33(6): 810-824. DOI:10.1029/2018GB005952 |
| [48] |
Fontaine C, Guimarães P R, Kéfi S, Loeuille N, Memmott J, van der Putten W H, van Veen F J F, Thébault E. The ecological and evolutionary implications of merging different types of networks. Ecology Letters, 2011, 14(11): 1170-1181. DOI:10.1111/j.1461-0248.2011.01688.x |