 2024, Vol. 44
2024, Vol. 44文章信息
- 张志强, 彭秀娟, 裴鑫怡, 王萍, 李文慧, 高歌, 李龙萱, 伍亮
- ZHANG Zhiqiang, PENG Xiujuan, PEI Xinyi, WANG Ping, LI Wenhui, GAO Ge, LI Longxuan, WU Liang
- 增温影响中华蟾蜍蝌蚪表型可塑性的生理代价
- Effects of increased temperatures on phenotypic plasticity and consequential physiological costs in the tadpoles of Bufo gargarizans
- 生态学报. 2024, 44(8): 3329-3336
- Acta Ecologica Sinica. 2024, 44(8): 3329-3336
- http://dx.doi.org/10.20103/j.stxb.202308081698
-
文章历史
- 收稿日期: 2023-08-08
- 网络出版日期: 2024-01-29






生理学权衡是理解生活史进化过程的核心内容之一[1—2]。两栖动物生活史复杂, 增温能加速变态期蝌蚪的发育进程, 促进形态表型发生可塑性改变[3—5], 但其生理基础, 仍不清楚[6—8]。随着免疫学技术介入两栖动物生活史研究, 解决了蝌蚪和变态后个体较小、取材不易等难题, 为阐明特定生活史阶段当前与未来存活之间的关系贡献了新思路, 提供了新方法[9—11]。当前, 常用血液中白细胞的数量动态来指示天然免疫和体液免疫能力的大小, 以嗜中性粒细胞与淋巴细胞的比值(N/L的比值)来表示应激反应能力[12—13], 以对植物血凝素(PHA-P)的反应来代表整合性免疫能力的高低, 在两栖动物成体[14—16]、幼体和蝌蚪[17—19]中, 均有较为广泛的应用, 且测定方法被不断优化。温度[20—22]和气候变化[2]显著影响两栖动物成体的免疫功能, 反应模式随物种而变化。水位加速干涸情况下, 黑斑侧褶蛙(Pelophylax nigromaculatus)蝌蚪完成变态后的幼蛙个体变小, PHA-P最大反应值下降[23], N/L的比值显著上升[24];荧光聚苯乙烯微球暴露和初次清除不影响黑斑侧褶蛙蝌蚪的PHA-P最大反应值, 但N/L的比值增强明显[25]。目前, 关于脊椎动物免疫功能与应激反应能力之间的关系, 仍有争论[26]。
中华蟾蜍(Bufo gargarizans)隶属于无尾目(Anura)蟾蜍科(Bufonidae)蟾蜍属, 在我国分布广泛, 从产卵到发育至幼蟾, 常伴有水温的大幅度波动[27]。中华蟾蜍母蟾产卵时的平均水温约为10℃[27], 在10、15、20、25℃下分别驯化2周, 其蝌蚪的最适温度范围、最高逃避温度和最高致死温度均有上移趋势[28]。分布于合肥地区的中华蟾蜍成体, 对PHA-P的反应无性别差异, 最大反应值出现于注射后12 h[14];水温[3]、水位[29]等均影响中华蟾蜍蝌蚪变态前后的形态表型。本研究参照中华蟾蜍蝌蚪生存的最适温度范围[28]以及合肥地区未来100年的气候变暖趋势[30], 设置了23、25、27、29℃组, 以37期中华蟾蜍蝌蚪为研究对象, 引入内脏器官大小、应激反应和免疫功能等测定指标, 突破了以往研究只关注形态表型的局限性, 将整合揭示变态后幼蟾响应高温的生理策略, 为预估其未来命运提供理论基础。
1 材料与方法 1.1 实验材料与试剂2022年3月初在安徽农业大学智慧广场小池塘(117.15°E, 31.52°N)采集中华蟾蜍卵团, 来源于同一卵团的个体在安徽农业大学动物生理生态学实验室塑料箱(长×宽×高=60 cm ×36 cm ×36 cm)内孵化和饲养, 每箱约1000只, 共3箱。箱体内水温为(22±1)℃, 水深约10 cm。参照Gosner分期鉴定标准, 当蝌蚪发育阶段至26—28期时[31], 以市售青菜叶煮熟后饲喂, 每隔3—5 d换曝气24 h以上的自来水一次。所用的快速瑞氏-吉姆萨染液(D010)购自南京建成生物工程有限公司, PHA-P(Sigma L-8754)购自Sigma-Aldrich公司。BA-210生物显微镜、显游标卡尺和移液器分别为麦克奥迪实业集团有限公司、上海恒量量具有限公司和德国艾本德公司产品。
1.2 实验方法取室内饲养发育至37期的中华蟾蜍蝌蚪, 通过电子天平(赛多利斯科学仪器有限公司, ± 0.1 mg)测其体重, 直尺(± 0.01 mm)测其体全长, 每30只蝌蚪随机置于水温为(23±1)℃、(25±1)℃、(27±1)℃和(29±1)℃的水族箱内(长×宽×高=48 cm×26 cm×30 cm)饲养, 分别简称为23℃组、25℃组、27℃组和29℃组, 加热棒控温, 水深约10 cm, 以泡沫板下系石块来营造登陆环境, 每一水温组做3次平行实验, 共使用蝌蚪360只, 即30只/箱×3箱/温度组×4个温度组=360只。实验开始时, 4个温度组共12个箱体间蝌蚪的体重(F3, 11=0.440, P>0.05)和体全长(F3, 11=2.780, P>0.05)均无显著差异。每天9:00和21:00观察蝌蚪变态情况, 直至所有蝌蚪均发育至46期幼蟾时为止, 统计每一箱内每只蝌蚪从37期发育至46期幼蟾的天数, 以其差值作为变态时长, 以所有蝌蚪个体变态时长的平均值表示该箱个体的变态时长。因各温度组蝌蚪发育至46期幼蟾的个体数不同, 23、25、27、29℃组分别随机共取20、15、22和14只幼蟾, 测其体重和体长后, 留测对PHA-P的反应;23、25、27、29℃组共分别随机取46、38、51和9只幼蟾, 测其体重和体长后, 断腿采血制作血涂片, 并解剖称量心、肝、脾、肺和肾的湿重, 以及去除内容物的胃和小肠的湿重, 测量胃和小肠的长度。23、25、27、29℃组37期蝌蚪的初始数量均为90只, 发育至46期幼蟾的个体数分别为66、53、73和23只, 存活率分别为73.33%、58.89%、81.11%和25.56%。
1.3 指标测定 1.3.1 器官大小指标

|
在46期幼蟾右侧大腿根部注射浓度为2.5 g/L的PHA-P溶液20 μL, 左侧后足不注射任何物质, 注射前后均用70%乙醇消毒被注射部位。在注射前(0 h)和注射后9 h内, 逐小时测量被注射部位皮肤组织的增厚厚度(± 0.01 mm), 确定对PHA-P的反应强度。为避免测量误差, 始终保持以相似角度测量同一部位, 每一测量时间点重复测量3次, 且均由同一人操作。
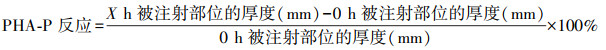
|
每一温度组每一箱内被采集到血液的46期幼蟾, 均取血液约20 μL, 制作血涂片1—2张, 放入37℃烘箱中烘干后, 用快速瑞氏-吉姆萨染色试剂盒染色, 经试剂Ⅰ和试剂Ⅱ分别染1—3和5—8 min后, 用蒸馏水充分冲洗约30 s, 37℃再次烘干后, 用中性树胶封片, 在400倍光学显微镜下镜检观察, 以“几”字形计数每张血涂片中不同视野中嗜中性粒细胞、嗜酸性粒细胞、嗜碱性粒细胞、单核细胞和淋巴细胞的数量, 直至计数至第100个白细胞时为止, 第2张血涂片也做相同处理, 以2张血涂片中每一种类型白细胞数量的平均值表示其百分占比情况。以N/L的比值来表示变态后46期幼蟾的应激反应能力。
1.4 统计分析用统计分析软件SPSS软件包进行数据处理。实验开始时, 统计每一温度组每一箱内30只37期蝌蚪的体重、体全长的平均值, 以3箱的平均值代表该温度组的情况。经one sample Kologorov-Smirnov测试检验数据的方差齐次性, 对不符合正态分布的胃长度系数和变态时长的数据分别进行反正弦和常用对数转换, 因转换后仍不符合正态分布, 故用非参数检验中的多个独立样本的Kruskal-Wallis H法进行统计分析。实验结束时, 用单因素方差分析比较不同水温组每只46期幼蟾体重、体长和变态时长, 以及内脏器官大小、各型白细胞的百分比、N/L的比值和PHA-P最大反应值的组间差异;以水温作为组间因子, 注射前(0 h)和注射后1至9 h为组内因子, 用重复测量方差分析比较水温对PHA-P反应模式的影响, 以及同一水温组内不同时间点之间的差异。文中数值均以平均值±标准误(Mean±SE)表示, P<0.05为差异显著。
2 结果与分析 2.1 增温对中华蟾蜍46期幼蟾体重、体长和变态时长的影响如图 1所示, 体重23℃组显著高于25℃组和27℃组(F3, 211=6.542, P<0.05), 但29℃组与其他各组的差异均不显著;体长23℃组最长, 27℃其次, 均显著长于25℃组(F3, 211=10.075, P<0.05), 但29℃组与其他各组的差异均不显著;变态时长29℃组显著低于23℃组和25℃组(F3, 11=8.713, P<0.05), 说明增温对变态后46期幼蟾的身体大小有先抑制再提升的作用, 但变态时长趋向于显著缩短。

|
| 图 1 增温对中华蟾蜍46期幼蟾体重、体长和变态时长的影响 Fig. 1 Effects of increased temperatures on body mass, body length and metamorphosis duration for Gosner stage 46 toadlets of Bufo gargarizans 柱上不同的小写字母代表温度组间差异显著(P<0.05) |
如图 2所示, 各温度组之间, 心(F3, 140=2.068, P>0.05)、肝(F3, 140=1.636, P>0.05)、脾(F3, 140=2.258, P>0.05)、肺(F3, 140=1.258, P>0.05)和肾(F3, 140=1.826, P>0.05)的湿重系数均无明显的组间差异;胃湿重系数25℃组显著高于27℃组(F3, 140=4.665, P<0.05), 小肠湿重系数23℃和25℃组均显著高于29℃组(F3, 140=3.127, P<0.05);胃长度系数25℃组显著高于23℃组和27℃组(F3, 140=3.115, P<0.05), 小肠长度系数无显著的组间差异(F3, 140=0.972, P>0.05), 说明适度增温有利于登陆后幼蟾胃和小肠的发育, 但偏高的温度不利于胃的机械消化及小肠消化和吸收功能的执行。

|
| 图 2 增温对中华蟾蜍46期幼蟾内脏器官大小的影响 Fig. 2 Effects of increased temperatures on internal organ size for Gosner stage 46 toadlets of Bufo gargarizans |
如图 3所示, 各温度组之间, 嗜中性粒细胞(F3, 163=1.317, P>0.05)和淋巴细胞的百分比(F3, 163=0.614, P>0.05)及N/L的比值(F3, 163=0.042, P>0.05)均无显著的组间差异;嗜酸性粒细胞的百分比25℃组显著高于其他各温度组(F3, 163=10.056, P<0.05);单核细胞的百分比23℃组和27℃组均显著高于25℃组(F3, 163=8.448, P<0.05);嗜碱性粒细胞的百分比23℃组显著高于25℃组(F3, 163=3.342, P<0.05), 说明增温不影响变态后幼蟾的应激反应能力, 但会导致白细胞数量发生动态改变, 表现为促进嗜酸性粒细胞增殖, 抑制嗜碱性粒细胞和单核细胞增殖。

|
| 图 3 增温对中华蟾蜍46期幼蟾白细胞数量及N/L的比值的影响 Fig. 3 Effects of increased temperatures on the number of different subtypes of leukocyte, and the ratio of neutrophils to lymphocytes (ratio of N/L) for Gosner stage 46 toadlets of Bufo gargarizans N/L的比值:嗜中性粒细胞与淋巴细胞的比值 |
如图 4所示, 23℃组(F9, 171=96.837, P<0.05)、25℃组(F9, 126=73.649, P<0.05)和27℃组(F9, 189=72.040, P<0.05)幼蟾PHA-P反应均在注射后5 h达到最大反应值, 4 h其次, 5 h时和4 h时均显著高于其他测量时间点;29℃组幼蟾PHA-P反应在注射后5 h达到最大反应值, 4 h时和6 h时其次, 5 h时显著高于其他测量时间点(F9, 117=33.518, P<0.05), 说明增温不影响PHA-P最大反应值的出现时间早晚。

|
| 图 4 增温对中华蟾蜍46期幼蟾对植物血凝素的反应的影响 Fig. 4 Effects of increased temperatures on the responsiveness to phytohemagglutinin-P (PHA-P) for Gosner stage 46 toadlets of Bufo gargarizans |
如图 5所示, 29℃组幼蟾PHA-P最大反应值显著高于其他温度组(F3, 67=3.072, P<0.05), 说明高温处理显著增强了PHA-P的反应强度。

|
| 图 5 增温对中华蟾蜍46期幼蟾植物血凝素最大反应值的影响 Fig. 5 Effects of increased temperatures on maximal phytohemagglutinin-P (PHA-P) response for Gosner stage 46 toadlets of Bufo gargarizans |
温度强烈影响两栖动物变态前后的表型可塑性, 通常随着温度升高, 变态时长缩短, 存活率下降, 身体趋向于变小。例如, 在平均水温为(15.7±7)℃、(21±1)℃和(26±1)℃的条件下, 中华蟾蜍蝌蚪的变态时长随水温增加而缩短, 变态后幼蟾的体长和体宽均趋向于下降[3]。本研究中, 从23℃升温至29℃组, 中华蟾蜍变态后幼蟾的体重和体长下降之后至29℃又有所回升, 但变态时长也明显缩短, 存活率趋向于大幅度下降。研究表明, 分布于浙江丽水的中华蟾蜍蝌蚪对水位变化表现出较差的可塑性, 在其早期发育阶段, 水位深浅只影响体长, 在逐减水位组最长、恒低水位组最短, 体重未见明显变化;水位变化也不影响其变态时长, 以及变态后幼蟾的体长和体重等指标, 这可能与中华蟾蜍在冬季繁殖的生活习性, 以及栖息地易于发生干旱和处于波动状态的水位有关[29]。与水位相比, 中华蟾蜍蝌蚪变态前后的身体大小对高温刺激更为敏感, 表现出了更强的可塑性。
内脏器官系数可反映两栖动物的营养和生理状态, 变态前后器官大小的适应性改变对其未来存活尤为重要[32—33]。本研究中, 增温并未显著影响中华蟾蜍变态后幼蟾的心、肝、脾、肺和肾等内脏器官的大小, 但胃和小肠的大小对高温的响应策略不同。心等重要内脏器官维持相对大小不变, 有助于充分发挥各自的功能;胃湿重系数和长度系数, 以及小肠湿重系数均在25℃组最高, 分别在27℃组和29℃组最低, 但小肠长度系数未见组间差异。(20±2)℃的室温条件下, 从27期至46期的变态发育过程中, 中华蟾蜍消化道长度在38期最长, 之后逐渐缩短[34];从42—46期, 中华蟾蜍处于从蝌蚪变为幼蟾的关键阶段, 其口无法执行摄食功能[34]。登陆后, 变态后幼蟾的胃容积扩大, 预示贮存和机械消化食物的能力有所增强, 小肠湿重增加, 与其较强的消化吸收能力有关, 这些变化均有助于幼蟾存活。然而, 25℃有利于中华蟾蜍蝌蚪胃和小肠发育, 但继续升高的温度却导致胃大小和小肠湿重缩减, 可能不利于幼蟾捕食和执行消化吸收功能。
3.2 中华蟾蜍蝌蚪变态后幼蟾免疫功能和应激水平对高温的响应免疫指标的变化可指示动物体的健康状态[1], 各型白细胞的百分比、N/L的比值和PHA-P反应是两栖动物生态免疫学研究中应用较多的指标[10], 但关于免疫与生长、免疫与应激能力之间的关系, 仍有争论[23—26]。
成体两栖动物中, 常用血液中皮质醇含量来代表其应激水平[12—13], 在受到外界环境条件胁迫后, 皮质醇含量在短时间内即可产生动态变化, 且其含量常与免疫功能表现为负相关关系[35—36], 但也有例外[37]。与成体相比, 蝌蚪和两栖动物幼体血量较少, 不适合以皮质醇含量作为衡量其应激反应能力的指标。N/L的比值是一个较好的替代指标, 该指标具有需血量少(低至3 μL)、计数准确等优点[10]。循环血中嗜中性粒细胞常是机体抵御感染的第一道防线, 淋巴细胞则具有参与免疫应答和免疫识别的功能, 前者数量增加, 后者数量减少, 两者的比值增大, 意味着发生了较强的炎症反应[12—13]。然而, N/L的比值的峰值出现时间通常滞后于皮质醇[12—13]。随着温度升高, 中华蟾蜍变态后幼蟾的N/L的比值均处于稳定平衡的状态, 说明增温未触发机体的炎症反应, 也未破坏机体的免疫平衡。在水温为21、23、25、27℃条件下, 增温可加速黑斑侧褶蛙37期蝌蚪的变态过程, 但对46期幼蛙N/L的比值也无明显影响[19]。然而, 当面临水位下降风险时, 变态后黑斑侧褶蛙46期幼蛙的N/L的比值快速干涸组显著高于其他水位处理组[24]。此外, 对黑斑侧褶蛙37期蝌蚪进行50 nm低、中和高浓度荧光标记聚苯乙烯微球溶液处理后, 连续测定了暴露7 d(7 d时)、清水清除7 d(14 d时)和14 d(21 d时)时的N/L的比值, 发现N/L的比值只在21 d时清水组低于其他浓度组, 低、中和高浓度组都从7 d时至21 d时显著增加[25]。
中华蟾蜍变态后幼蟾25℃组嗜酸性粒细胞的百分比占比最高, 而嗜碱性粒细胞和单核细胞的百分比都占比最低。嗜酸性粒细胞、嗜碱性粒细胞及单核细胞均能执行天然免疫功能[12], 它们的相对值却随增温变化趋势相反, 说明免疫系统内部的不同组分之间对高温的反应并不一致。适度升高的温度会增加细菌、寄生虫侵害有机体的概率, 亦会激活幼蟾登陆后杀伤细菌、寄生虫的功能, 而更高的29℃的水温则使细菌、寄生虫滋生概率增加, 但却抑制了其天然免疫功能。然而, 29℃条件下, 幼蟾对PHA-P的反应的最大值却显著高于23、25、27℃组, 提示由PHA-P所诱导的细胞介导的免疫能力及体液免疫能力均在高温刺激下得到了显著增强。PHA-P是一种有丝分裂原, 反应机制复杂, 被认为是一种集合了细胞介导的免疫能力和/或体液免疫能力的整合性免疫应答反应[10, 15], 对PHA-P的最大反应值的出现时间早晚受发育阶段[17, 19]、温度[7]、水位[23]和微塑料[25]等因素影响, 但这些因素并不影响对PHA-P的反应的强度。值得注意的是:本研究中, 当水温为29℃时, 中华蟾蜍变态为幼蟾的存活率只有25.56%。能存活个体对PHA-P的反应保持高强度, 可能与高温导致的细菌和寄生虫感染强度增加有关, 提示登陆后被激活的免疫功能可能有助于其当前存活, 但不利于未来存活。
4 结论增温导致中华蟾蜍登陆后幼蟾的变态加速、白细胞数量动态改变和PHA-P反应强度增加, 但对应激反应能力无影响, 存活率下降, 被激活的免疫功能可能有利于幼蟾快速适应陆地环境, 但不利于其未来存活。
| [1] |
张志强, 王德华. 免疫能力与动物种群调节和生活史权衡的关系. 应用生态学报, 2005, 16(7): 1375-1379. |
| [2] |
Rollins-Smith L A, Le Sage E H. Heat stress and amphibian immunity in a time of climate change. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 2023, 378(1882): 20220132. DOI:10.1098/rstb.2022.0132 |
| [3] |
刘莉, 李成, 李乃兵, 徐宏发, 王跃招. 不同水温条件下中华蟾蜍蝌蚪的表型可塑性研究. 四川动物, 2006, 25(2): 214-217. |
| [4] |
王守红, 李豪, 刘露莎, 张兴其, 江建平. 温度对饰纹姬蛙蝌蚪生长的影响. 动物学杂志, 2018, 53(2): 191-197. |
| [5] |
Ruthsatz K, Peck M A, Dausmann K H, Sabatino N M, Glos J. Patterns of temperature induced developmental plasticity in anuran larvae. Journal of Thermal Biology, 2018, 74: 123-132. DOI:10.1016/j.jtherbio.2018.03.005 |
| [6] |
Gervasi S S, Foufopoulos J. Costs of plasticity: responses to desiccation decrease post-metamorphic immune function in a pond-breeding amphibian. Functional Ecology, 2008, 22(1): 100-108. DOI:10.1111/j.1365-2435.2007.01340.x |
| [7] |
张志强, 王佳慧, 金冰艳, 陈利, 杨雨晴, 刁迎珠, 伍亮. 温度对黑斑侧褶蛙变态时长及PHA-P反应的影响. 生态学杂志, 2021, 40(12): 4029-4033. |
| [8] |
张志强, 杨雨晴, 陈利, 金冰艳, 王佳慧, 刁迎珠, 伍亮. 水温升高对黑斑侧褶蛙蝌蚪变态时长和血液参数的影响. 安徽农业大学学报, 2022, 49(2): 254-258. |
| [9] |
Brock P M, Murdock C C, Martin L B. The history of ecoimmunology and its integration with disease ecology. Integrative and Comparative Biology, 2014, 54(3): 353-362. DOI:10.1093/icb/icu046 |
| [10] |
张志强. 动物生态学研究中免疫学参数的选择及其优缺点分析. 四川动物, 2015, 34(1): 145-148. |
| [11] |
Rollins-Smith L A. Amphibian immunity-stress, disease, and climate change. Developmental and Comparative Immunology, 2017, 66: 111-119. DOI:10.1016/j.dci.2016.07.002 |
| [12] |
Davis A K, Maney D, Maerz J. The use of leukocyte profiles to measure stress in vertebrates: a review for ecologists. Functional Ecology, 2008, 22: 760-772. DOI:10.1111/j.1365-2435.2008.01467.x |
| [13] |
Davis A K, Maney D L. The use of glucocorticoid hormones or leucocyte profiles to measure stress in vertebrates: What's the difference?. Methods in Ecology and Evolution, 2018, 9(6): 1556-1568. DOI:10.1111/2041-210X.13020 |
| [14] |
Zhang Z Q, Jin C C, Qu K S, Caviedes-Vidal E. Immune responsiveness to phytohemagglutinin displays species but not sex differences in three anuran species. PeerJ, 2017, 5: e3181. DOI:10.7717/peerj.3181 |
| [15] |
Zamara-Camacho F J. Integrating time progression in ecoimmunology studies: beyond immune response intensity. Current Zoology, 2019, 65(2): 205-212. DOI:10.1093/cz/zoy045 |
| [16] |
彭兵龙, 张昊滨, 陈可为, 杨会会, 连丽燕, 汪寅, 张志强. 中华蟾蜍足垫部位对植物血凝素的反应及其组织学和血液学变化. 动物学杂志, 2019, 54(1): 87-98. |
| [17] |
高慧清, 张琼文, 孙思琪, 连丽燕, 王红, 张志强. 变态期花背蟾蜍蝌蚪对植物血凝素的反应模式及其生态学意义. 生态学杂志, 2017, 36(9): 2542-2548. |
| [18] |
Zamara-Camacho F J, Comas M. Early swelling response to phytohemagglutinin is lower in older toads. PeerJ, 2018, 6: e6104. DOI:10.7717/peerj.6104 |
| [19] |
张志强, 王泽洋, 许洋溢, 陈利, 王佳慧, 王永泰. 黑斑侧褶蛙变态前后身体大小的变化及对植物血凝素的反应模式. 安徽农业大学学报, 2020, 47(4): 519-523. |
| [20] |
肖锋, 颉志刚, 庄建洲. 高温刺激对蟾蜍免疫活性和能量消耗的影响. 生态学报, 2015, 35(9): 3087-3092. |
| [21] |
Moretti E H, Titon S C M, Junior B T, Marques F S, Gomes F R. Thermal sensitivity of innate immune response in three species of Rhinella toads. Comparative Biochemistry and Physiology Part A, Molecular and Integrative Physiology, 2019, 237: 110542. DOI:10.1016/j.cbpa.2019.110542 |
| [22] |
Bakewell L, Kelehear C, Graham S P. Impacts of temperature on immune performance in a desert anuran (Anaxyrus punctatus). Journal of Zoology, 2021, 315(1): 49-57. DOI:10.1111/jzo.12891 |
| [23] |
伍亮, 高歌, 史爽, 李龙萱, 彭秀娟, 李文慧, 王萍, 张志强. 水位变化影响蝌蚪存活、形态表型和植物血凝素反应. 应用生态学报, 2022, 33(12): 3427-3432. |
| [24] |
伍亮, 李龙萱, 高歌, 史爽, 王月妍, 冯文红, 鲍传和, 张志强. 水位对黑斑侧褶蛙蝌蚪形态表型、白细胞数量和应激反应的影响. 动物学杂志, 2023, 58(1): 117-125. |
| [25] |
刁迎珠, 裴鑫怡, 王萍, 史爽, 李文慧, 彭秀娟, 李冉, 伍亮, 鲍传和, 张志强. 荧光聚苯乙烯微球对蝌蚪应激水平和免疫功能的影响. 生态学杂志, 2023, 42(5): 1142-1149. |
| [26] |
Costantini D. A meta-analysis of impacts of immune response and infection on oxidative status in vertebrates. Conservation Physiology, 2022, 10(1): coac018. DOI:10.1093/conphys/coac018 |
| [27] |
费梁, 胡淑琴, 叶昌媛, 田婉淑, 江建平, 吴贯夫, 李健, 王宜生. 中国动物志两栖纲中卷无尾目. 北京: 科学出版社, 2009.
|
| [28] |
王立志. 恒温和变温驯化对大蟾蜍蝌蚪热耐受性的影响. 生态学报, 2014, 34(4): 1030-1034. |
| [29] |
唐小芬, 樊晓丽, 林植华, 姚婷婷, 李香, 金晶, 周存通. 浙江丽水中华大蟾蜍和黑眶蟾蜍蝌蚪对水位变化的表型响应. 生态学报, 2015, 35(3): 911-918. |
| [30] |
王文本, 王玉红, 刘文海. 合肥市快速发展模式下城郊局地气候变化特征分析. 智能城市, 2019, 5(14): 29-31. |
| [31] |
Gosner K L. A simplified table for staging anuran embryos and larvae. Herpetologica, 1960, 16(3): 183-190. |
| [32] |
Patmann M D, Shewade L H, Schneider K A, Buchholz D R. Xenopus tadpole tissue harvest. Cold Spring Harbor Protocols, 2017. DOI:10.1101/pdb.prot097675 |
| [33] |
连丽燕, 高慧清, 孙嘉珞, 孙思琪, 张琼文, 张志强. 变态前后花背蟾蜍蝌蚪消化器官大小及各型白细胞百分比的适应性变化. 生态学杂志, 2018, 37(4): 1204-1210. |
| [34] |
王念, 吴民耀, 王宏元, 李忻怡. 中华蟾蜍蝌蚪变态过程中消化系统的重构. 西北农林科技大学学报, 2010, 38(12): 7-12. |
| [35] |
Lima A S, Ferreira L D F, Silva D P, Gomes F R, Titon S C M. Thermal sensitivity of Bullfrog's immune response kept at different temperatures. Journal of Experimental Zoology Part A, Ecological and Integrative Physiology, 2020, 333(10): 767-778. DOI:10.1002/jez.2436 |
| [36] |
Lima A S, de Figueredo A C, Floreste F R, Neto P G G, Gomes F R, Titon S C M. Temperature extreme events decrease endocrine and immune reactive scope in bullfrogs (Lithobates catesbeianus). Integrative and Comparative Biology, 2022, 62(6): 1671-1682. DOI:10.1093/icb/icac105 |
| [37] |
Gomes F R, Madelaire C B, Moretti E H, Titon S C M, Assis V R. Immunoendocrinology and ecoimmunology in Brazilian anurans. Integrative and Comparative Biology, 2022, 62(6): 1654-1670. DOI:10.1093/icb/icac014 |