 2024, Vol. 44
2024, Vol. 44文章信息
- 巢林, 欧梦菲, 陈健, 卢卫福, 马琳, 李忠国, 黄柏华, 明安刚, 张建兵, 胡宝清, 刘艳艳
- CHAO Lin, OU Mengfei, CHEN Jian, LU Weifu, MA Lin, LI Zhongguo, HUANG Bohua, MING Angang, ZHANG Jianbing, HU Baoqing, LIU Yanyan
- 喀斯特11种典型生态恢复树种凋落叶分解及其对土壤碳排放的激发效应
- Leaf litter decomposition and its priming effect of eleven typical ecological restoration tree species in the karst area of China
- 生态学报. 2024, 44(8): 3397-3407
- Acta Ecologica Sinica. 2024, 44(8): 3397-3407
- http://dx.doi.org/10.20103/j.stxb.202307291630
-
文章历史
- 收稿日期: 2023-07-29
- 网络出版日期: 2024-01-29






2. 中国科学院华南植物园, 中国科学院退化生态系统植被恢复与管理重点实验室, 广州 510650;
3. 湖南会同森林生态系统国家野外科学观测研究站, 会同 418307;
4. 中国林业科学研究院热带林业实验中心, 广西友谊关森林生态系统国家定位观测研究站, 凭祥 532600
2. Key Laboratory of Vegetation Restoration and Management of Degraded Ecosystems, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China;
3. Huitong National Research Station of Forest Ecosystem, Huitong 418307, China;
4. Experimental Center of Tropical Forestry, Chinese Academy of Forestry, Guangxi Youyiguan Forest Ecosystem Research Station, Pingxiang 532600, China
喀斯特生态系统(约占世界陆地面积的15%)是陆地生态系统的重要组成部分, 在全球碳(C)循环和气候调节方面发挥着至关重要的作用[1—2]。中国西南喀斯特地区是全球三大岩溶集中分布区中面积最大、岩溶作用最为强烈的地区, 同时也是石漠化最严重和生态环境最脆弱的地区之一[3—4]。尽管喀斯特土壤富含C, 是一个巨大的碳库[5], 但是由于土层浅薄且不连续, 平均土壤厚度仅为2—9 cm, 土壤中的C集中于微生物活跃的表层, 使其对气候变暖和CO2浓度升高等全球变化更为敏感[6]。因此, 即使喀斯特土壤碳库发生小幅变化, 也可能对区域甚至全球气候产生重大影响。
喀斯特生态系统石漠化治理和植被恢复关键在于增加植被覆盖度、保土蓄水、提高土壤肥力, 而植物凋落物在保持水土、提高土壤肥力和促进生态系统养分循环等方面扮演着重要角色[7—9], 其分解过程对喀斯特生态系统物质循环、能量转换以及生态系统服务功能的发挥与维持具有重要意义[9]。凋落物分解主要受气候条件、凋落物质量和分解者的调控[10], 其中凋落物质量是决定其分解快慢的第一要素[11]。植物凋落物不仅是土壤有机碳(SOC)的主要来源, 还可以通过激发效应改变SOC输出。土壤激发效应是指凋落物等外源碳输入导致原有SOC分解加速或减慢的现象[12]。研究表明正激发效应可促进SOC分解增加12倍, 而负激发效应能降低90%的SOC分解[13]。此外, 引入土壤激发效应可以提高地球系统模型对土壤碳储量的预测能力[14]。目前, 对于西南喀斯特区凋落物分解的研究主要集中在凋落物量、组成特征和动态变化[7, 15—16]、养分归还特征[17]、凋落物化学计量特征[18]、落叶树种与常绿树种凋落叶分解差异[19]等方面, 而关于喀斯特地区凋落物分解诱导的土壤激发效应研究报道较少。
植树造林是治理喀斯特退化生态系统的重要手段之一, 而选择适宜的造林树种对植被恢复至关重要[20]。尽管前人研究已筛选出一大批适于喀斯特石漠化治理的优良树种[3, 21], 但是对于生态恢复树种在土壤肥力提升、固碳效应等方面的研究较为缺乏, 不足以科学地支撑喀斯特退化生态系统造林树种的选择。本研究选择了11种适应性强、耐干旱贫瘠的生态恢复优良树种, 在喀斯特石山树木园进行了研究。分析了这些物种的凋落叶性状特征, 探究了不同树种凋落叶分解及其土壤激发效应的差异, 以期明确典型生态恢复树种凋落叶性状与凋落叶分解及其激发效应强度之间的关系, 为喀斯特退化生态系统恢复树种的科学选择提供理论依据。
1 材料与方法 1.1 研究区概况研究地位于广西凭祥市中国林业科学研究院热带林业实验中心大青山石山树木园(106°39′50″—106°59′30″E, 21°57′47″—22°19′27″N), 该园由我国著名林学家吴中伦院士主持设计, 建于1980年, 总面积36.8 hm2, 是我国目前唯一以收集保存热带亚热带岩溶石山树种为重点的树木园。该树木园所处地区属于南热带季风气候, 雨热同期, 年平均气温21℃, 年平均降水量1500 mm, 干湿季分明, 雨季集中于4—10月。地貌类型为典型的喀斯特峰丛洼地, 海拔150—800 m, 土壤类型以碳酸盐岩发育成的石灰土为主。经过40多年的生态恢复, 该树木园已经从退化严重的残次生林群落转变为物种丰富的近自然森林群落, 引进和保存树种118科393属680种, 其中石山树种334种, 经中试及推广实验, 已经筛选出50多种适应石山生境的优良造林树种。
1.2 研究方法 1.2.1 土壤和凋落物采集本研究采集的土壤来自广西弄岗国家级自然保护区弄岗保护站附近的农田耕作层(0—20 cm)。该农田种植C4甘蔗已有连续50年以上的历史。土壤C和N含量分别为18.9 g/kg和2.2 g/kg, C∶N比为8.6, pH为4.9。C4甘蔗土的δ13C值为-14.3‰。土壤风干、充分混匀并过2 mm筛, 剔除可见的植物残体和细石。
2020年11月, 在大青山石山树木园选择东京桐(Deutzianthus tonkinensis)、光皮梾木(Cornus wilsoniana wanger)、蚬木(Excentrodendron tonkinense)、青冈栎(Cyclobalanopsis glauca)、海南椴(Hainania trichosperma)、任豆(Zenia insignis)、顶果树(Acrocarpus fraxinifolius)、降香黄檀(Dalbergia odorifera)、割舌树(Walsura robusta)、南酸枣(Choerospondias axillaris)和望天树(Parashorea chinensis)等11种典型喀斯特恢复树种, 每个树种选择3—4棵成熟植株, 收集对应树种200 g新鲜凋落叶。收集完成后, 将凋落叶放入60℃烘箱烘干至恒重。烘干后的凋落物样品分为两部分:一部分用球磨仪进行研磨, 用于测定相关化学指标;另一部分凋落叶剪成1 cm2正方形, 用于开展凋落叶分解实验。
1.2.2 实验设计本研究利用C3植物凋落叶和C4甘蔗土壤δ13C自然丰度的差异来区分凋落叶来源CO2和土壤来源CO2。称150 g干重C4土壤放入1 L培养瓶, 于25℃条件下进行为期10 d的预培养, 以最大限度减少土壤扰动对实验结果的影响。预培养结束后, 将0.5 g凋落叶添加至土壤表层, 每种凋落叶处理设置4个重复, 同时设置4个对照处理组不添加凋落物。接下来, 开展为期180 d的培养实验, 并通过添加去离子水, 使土壤含水率在整个实验期间维持在田间饱和持水量的60%。在培养第3、7、13、20、30、45、60、90、140 d和180 d进行土壤气体采集。
1.2.3 指标测定与计算凋落叶性状∶凋落叶干物质含量(LDMC, mg/g)为凋落叶干重与其鲜重的比值;采用标准方法测定凋落叶C、N、P、水溶性C、木质素、半纤维素、纤维素和单宁含量等。C和N含量运用CN元素分析仪测定(ElementarVario, 湖南, 德国)。为测定凋落物可溶性C, 往2 g凋落物粉末样品中加入60 ml去离子水, 并放入振荡器上振荡30 min, 将过滤后的浸提液放入总有机碳(TOC)分析仪中进行测定(Vario TOC cube, Elementar Analysis system GmbH, 朗根塞尔博德,德国)。木质素、半纤维素和纤维素(300 mg凋落物样品)依据美国国家可再生能源实验室测定标准[22], 采用两歩酸水解法进行分馏, 水解的液体用于量化纤维素和半纤维素, 运用高效液相色谱仪(HPLC) (Agilent-1260, Agilent Technologies, 圣克拉拉,加利福利亚州,美国) 测定。单宁含量根据Hagerman(2011)所描述的酸-丁醇方法进行测定[23]。将凋落物样品放入硫酸和高氯酸中进行酸化, 酸化后的溶液用于测定凋落物初始的P、Ca、K、Mg和Mn含量, 运用电感耦合等离子体质谱测定(ICP-MS) (Elan DRC-e; PerkinElmer, 诺沃克,康涅狄格州,美国)。培养实验结束后, 挑出培养瓶土壤中未分解凋落叶残体, 小心去除黏着在凋落叶残体表面的土壤颗粒后, 将凋落叶残体置于60℃条件下烘干至恒重, 并称重计算凋落叶质量损失百分比(ML, %)。计算公式为:
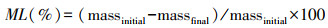
|
(1) |
式中, massinitial和massfinal分别为凋落叶初始干重和实验结束后剩余干重。
土壤呼吸和激发效应:土壤呼吸测定流程如图 1所示。在既定采样时间节点, 用无CO2的空气冲洗培养瓶后, 关闭培养瓶气阀开关。在25℃恒温培养箱内培养12 h后, 应用便携式气泵和真空气袋采集培养瓶中的气体, 运用Picarro G2131-i高精度CO2同位素分析仪测定CO2浓度和δ13C同位素值。

|
| 图 1 土壤呼吸测定流程 Fig. 1 Procedures for soil respiration measurements |
土壤呼吸计算公式如下:
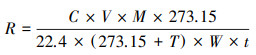
|
(2) |
式中, R为CO2-C释放量(μg C g-1土壤d-1);C为测定CO2浓度(μmol/mol);V为培养瓶体积(L);M为C的摩尔质量(12 g/mol);22.4(L)是理想气体在1个标准大气压和温度为273.15 K条件下的摩尔体积;W为土壤的干重(g);t为CO2累积时间(d);T为培养温度(25℃)。
采用质量守恒方程来区分凋落叶来源和土壤来源CO2:
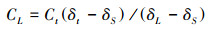
|
(3) |
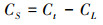
|
(4) |
式中, Ct为来源于土壤呼吸总的CO2-C(Ct=CL+CS);CL为来源于C3凋落叶的C;CS为来源于C4土壤的C;δt为来源于凋落叶处理土壤所释放CO2的同位素值δ13C;δL和δs分别为C3凋落叶和C4土壤同位素值δ13C。
土壤激发效应(PE, %)计算公式为:
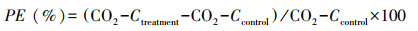
|
(5) |
式中, CO2-Ctreatment表示添加凋落物处理土壤总的CO2释放量;CO2-Ccontrol为来源于对照土壤CO2释放量。SOC分解的绝对变化量为CO2-Ctreatment减去CO2-Ccontrol。
1.3 数据处理与分析统计分析前, 对所有数据进行正态分布和同质性检验, 根据需要将凋落叶木质素和磷含量进行Log10转化。采用单因素方差分析(One-way ANOVA)和多重比较(Tukey HSD)检验比较不同树种凋落叶质量损失和激发效应强度的差异性;运用皮尔逊(Pearson)相关分析和线性回归分析对凋落叶性质与凋落物质量损失和激发效应强度之间的相关性进行分析, 并用逐步回归分析建立多元回归模型, 基于赤池信息准则(AICc)筛选最优模型。利用主成分分析(PCA)进一步探讨凋落叶性质与质量损失和激发效应之间的关系。Pearson相关分析在R4.2.1软件中完成, PCA分析在Canoco 5.0软件中进行, 使用OriginPro 2018绘图。
2 结果与分析 2.1 凋落叶性状特征及其变异系数11个生态恢复树种凋落叶性状表现出较大的变异性(表 1)。11个树种凋落叶质量损失相差3倍(CV=30%), 而凋落物激发效应强度变异幅度相对较大(CV=90%)。凋落叶C相关化学性质中, 单宁含量变异最大(CV=62%), 而总碳含量变异最小(CV=5%)。P和Mg含量呈现出高度变异性(CV>100%), 而K、Ca和Mn等其他养分相关化学性质变异程度相近(CV=31%—39%), 属于中等程度变异。化学计量比特征中, N/P的变异最大(CV=77%, 相差32倍), 而C/N的变异幅度最小(CV=24%, 相差2倍)。
| 凋落叶特征 Leaf litter traits |
均值 Mean |
中位数 Median |
最小值 Minimum |
最大值 Maximum |
变异系数 Coefficient of variation/% |
| 质量损失ML/% | 35.31 | 33.86 | 16.50 | 49.93 | 30.26 |
| 激发效应PE/% | 37.55 | 53.54 | -22.54 | 81.12 | 89.91 |
| 叶干物质含量LDMC/(mg/g) | 399.13 | 381.64 | 219.46 | 664.08 | 28.11 |
| 碳C/(mg/g) | 431.20 | 428.12 | 390.66 | 473.65 | 5.20 |
| 非结构性碳NSC/(mg/g) | 42.23 | 43.91 | 24.08 | 53.34 | 20.33 |
| 水溶性碳WSC/(mg/g) | 231.72 | 229.33 | 134.09 | 375.59 | 35.36 |
| 木质素Lignin/(mg/g) | 124.46 | 114.35 | 93.58 | 182.76 | 23.89 |
| 纤维素Cellulose/(mg/g) | 126.44 | 115.61 | 70.92 | 216.42 | 30.57 |
| 半纤维素Hemicellulose/(mg/g) | 83.55 | 86.75 | 46.96 | 162.39 | 38.72 |
| 单宁Tannin/(mg/g) | 36.18 | 27.44 | 9.45 | 68.96 | 62.28 |
| 氮N/(mg/g) | 12.72 | 11.59 | 9.05 | 22.83 | 31.20 |
| 磷P/(mg/g) | 0.28 | 0.16 | 0.06 | 1.38 | 133.41 |
| 钾K/(mg/g) | 5.86 | 5.14 | 2.85 | 11.14 | 39.72 |
| 钙Ca/(mg/g) | 31.50 | 31.81 | 17.25 | 50.67 | 31.44 |
| 镁Mg/(mg/g) | 0.19 | 0.04 | 0.01 | 1.06 | 157.22 |
| 锰Mn/(mg/g) | 2.63 | 2.93 | 0.96 | 3.69 | 32.41 |
| 木质素/氮Lignin/N | 10.50 | 9.44 | 6.97 | 19.90 | 35.99 |
| 碳/氮C/N | 36.45 | 39.33 | 19.87 | 47.32 | 23.93 |
| 碳/磷C/P | 3525.00 | 2775.03 | 343.64 | 7665.19 | 63.13 |
| 氮/磷N/P | 113.18 | 76.53 | 8.74 | 281.11 | 77.00 |
| ML: 质量损失Mass loss; PE: 激发效应Priming effect; 叶干物质含量: Leaf dry matter content; C: 碳Carbon; NSC: 非结构性碳Non-structure carbohydrates; WSC: 水溶性碳Water soluble carbon; N: 氮Nitrogen; P: 磷Phosphorus; K: 钾potassium; Ca: 钙Calcium; Mg: 镁Magnesium; Mn: 锰Manganese | |||||
不同生态恢复树种凋落叶质量损失百分比和激发效应强度均存在极显著差异(图 2, P<0.001)。11个树种凋落叶质量损失率最大与最小值相差3倍, 其中海南椴的凋落叶分解速度最快, 质量损失达到50%, 而青冈栎凋落叶质量在整个培养期间仅损失16.5%, 其分解速度最慢(图 2)。与对照土壤CO2释放量2.3 mg C kg-1土壤d-1相比, 凋落叶处理土壤CO2释放量为5.1 mg C kg-1土壤d-1, 是对照土壤平均每天CO2释放量的2.2倍(图 2)。除了海南椴、任豆和割舌树为负的激发效应之外, 其余8个生态恢复树种凋落叶都促进了土壤有机碳分解, 产生了正的激发效应(图 2)。总体而言, 11种生态恢复树种凋落叶输入平均促进土壤有机碳分解达37.6%, 其中望天树凋落叶分解诱导的土壤激发效应强度最大(81%), 而割舌树的激发效应强度最小(-6.9%)。

|
| 图 2 凋落叶质量损失百分比、土壤CO2释放量及土壤激发效应 Fig. 2 The percentage mass loss of leaf litter and its priming effect 不同字母表示各处理之间存在显著差异(P<0.05) |
相关性与线性回归分析结果显示, 凋落叶分解与非结构性碳(NSC)和水溶性碳(WSC)含量(图 3和图 4)呈显著正相关, 与LDMC、纤维素和Mn含量(图 3和图 4)呈显著负相关关系;然而, 凋落叶性质与土壤激发效应之间无显著相关关系(图 3)。通过逐步回归分析不同生态恢复树种凋落叶分解与凋落物性质之间的关系, 发现WSC、K和Ca相结合可以解释凋落叶分解变异的98%(表 2)。

|
| 图 3 凋落叶性质与凋落物分解和激发效应之间的相关性 Fig. 3 Matrix of Pearson correlation coefficients between the mass loss of leaf litter and litter properties ML: 质量损失Mass loss; PE: 激发效应Priming effect; LDMD: 叶干物质含量Leaf dry matter content; C: 碳Carbon; NSC: 非结构性碳Non-structure carbohydrates; WSC: 水溶性碳Water soluble carbon; Lignin: 木质素; Cellulose:纤维素;Hemicellulose:半纤维素;Tannin:单宁; N: 氮Nitrogen; P: 磷Phosphorus; K: 钾potassium; Ca: 钙Calcium; Mg: 镁Magnesium; Mn: 锰Manganese;*代表差异性(P<0.05) |

|
| 图 4 11个生态恢复树种凋落叶分解与凋落物性质之间的关系 Fig. 4 Relationships between the mass loss of leaf litter and key litter properties |
| 模型 Model |
赤池信息准则指数 Akaike information criterion |
凋落叶性质 Leaf litter traits |
决定系数R2 | 显著性 Significance level |
| 1 | 14.99332 | WSC; K; Ca | 0.981 | P<0.0001 |
| 2 | 22.59612 | WSC; K | 0.954 | P<0.0001 |
| 3 | 31.62306 | WSC | 0.876 | P<0.0001 |
| WSC: 水溶性碳Water soluble carbon; K: 钾potassium; Ca: 钙Calcium | ||||
对11个树种凋落叶分解、激发效应和化学性质进行主成分分析, 发现前两个排序轴可以解释48.5%的凋落叶性状变异(图 5)。第一轴主要由ML、C和养分相关化学性质(WSC、NSC、纤维素和Mn)以及LDMC等指标组成, 解释了25.8%的变异。第二轴主要由化学计量比特征和N含量等指标组成, 解释了22.7%的变异。从PCA分析图可以更直观的看出, 凋落叶质量的损失与WSC和NSC呈正相关, 但与半纤维素、Mn和LDMC呈负相关, 这与相关性和回归分析结果一致(图 5)。

|
| 图 5 喀斯特11个典型生态恢复树种凋落叶性质主成分分析 Fig. 5 Principal component analysis (PCA) of twenty leaf litter properties from 11 ecological restoration tree species |
本研究发现凋落叶C含量变异最小(CV=5%), 表明11个生态恢复树种具有类似的C含量。尽管不同生态恢复树种凋落叶C含量变化不明显, 但是不同形态碳化合物, 如水溶性碳、纤维素和单宁等C相关化学性状变异较大(CV=30%—62%), 这主要是由于不同化学形态碳化合物在分子量大小、结构复杂性和化学键类型等方面存在较大差异引起的。11个生态恢复树种凋落叶平均C含量为431.2 mg/g, 远低于弄岗喀斯特森林6个常见树种的599.8 mg/g[7], 但高于桂西北喀斯特原生林群落的402.5 mg/g和次生林群落的389.6 mg/g[18], 与贵州普定喀斯特生态系统观测研究站6个典型喀斯特树种的434.4 mg/g[19]以及西双版纳植物园喀斯特森林群落的442.7 mg/g相近[16]。
与C相关化学性状相比, 凋落叶养分相关化学性状, 尤其是P和Mg含量呈现出高度变异(CV>130%), 表明不同生态恢复树种养分利用与重吸收方面可能存在较大差异。研究表明喀斯特地区土壤普遍受P元素限制[1, 24], 因此不同树种可能采取不同的养分利用与获取策略以满足植物生长对P的需求, 比如一些树种利用根系直接吸收土壤中的P元素, 另一些树种则通过菌根真菌促进植物对P吸收[25]。此外, 为了最大程度上降低植物对外部环境养分的依赖, 植物叶片在衰老凋亡前会将养分转移到其他鲜活组织, 叶片养分重吸收这一过程是植物利用养分的重要策略之一[26]。在P匮乏的喀斯特地区, 植物叶片P元素的重吸收过程显得尤为重要, 而不同恢复树种叶片P元素重吸收效率的差异性则可能是导致凋落叶P含量差异巨大的另一个重要原因[27—28]。
3.2 凋落物分解及其诱导的土壤激发效应植物凋落叶C相关性状、养分含量和物理性质等是调控凋落物分解的重要因素[29]。不同生态恢复树种凋落叶性状呈现出较大变异, 从而导致不同树种凋落叶分解存在极显著差异。叶干物质含量是叶片结构成分(非活性组分, 木质素和纤维素等)相对于叶片总质量的度量[30], 反映了叶片密度、结构支撑和对食草动物防御能力, 可以表征凋落叶质量高低, 因此能够较好的预测凋落叶分解速率[31]。本研究发现, 低LDMC树种的凋落叶比高LDMC树种分解更快, 与之前的研究结果相一致[32—33]。这主要因为高叶干物质含量意味着叶片具有更高比例的维管束组织和厚壁组织, 反映在化学成分方面则表现出凋落叶具有更高的纤维素、不可溶性糖和木质素等[34—35]。本研究还发现叶干物质含量与纤维素含量呈正相关趋势, 而纤维素含量与凋落叶质量损失之间呈显著负相关关系进一步解释了为何高干物质含量树种凋落叶分解更慢。
非结构性碳水化合物和水溶性碳是植物凋落物中易分解的组分。在凋落物分解初期, 易分解组分可以快速释放进入土壤并被微生物利用[8, 36]。研究发现, 与非喀斯特生态系统相比, 喀斯特生态系统土壤微生物更容易受到碳限制的影响[37]。因此, 在凋落物分解早期阶段, 非结构性碳水化合物和水溶性碳的快速释放能够为土壤微生物提供碳源, 促进微生物的生长和活性, 从而加速凋落物的分解[38—39]。前人研究表明, 凋落物中的Mn元素在木质素分解酶的作用过程中起到辅助因子和调节剂的作用, Mn含量越高, 凋落物的分解速度越快[40—42]。然而, 本研究发现凋落叶Mn含量与凋落物分解呈显著负相关, 这可能是由于凋落物中Mn含量过高则可能对土壤微生物产生抑制或毒害作用, 从而减缓凋落物分解。凋落物分解通常受到N和木质素的控制[11, 43], 但多元回归分析表明, 凋落叶分解则主要受WSC、K和Ca含量控制。在为期200天的室内培养实验过程中, 11种生态恢复树种凋落叶平均分解程度为35%(范围在16.5%—49.9%)。这个分解程度高于亚热带森林30个树种为期12周的凋落叶分解培养实验(平均分解程度为17.8%, 范围在9.0%—35.1%)[44], 但低于温带森林35个树种为期6年凋落叶野外分解实验结果(平均分解程度为77%)[45], 与不同海拔梯度为期1年的凋落叶分解实验结果类似(平均分解程度为40%, 范围在11%—76%)[46]。这表明本研究所观察到的凋落叶质量损失在之前相关室内培养或野外原位凋落分解实验的范围之内。因此, 探讨凋落叶质量损失与其初始化学性质之间的关系是可靠的。前人研究还指出, 凋落物中Mg和Mn等元素含量是影响凋落物分解速率的重要因素[42]。因此, 未来研究应关注K、Ca、Mg和Mn等重要化学组分对凋落物分解的影响, 而不仅仅局限于C、N和P这3种元素[47]。
土壤激发效应强度和方向受到多种生物和非生物因素的调控, 包括凋落物属性(碳输入数量和质量)、土壤理化性质(SOC含量、养分有效性、pH等)和微生物群落(生物量、群落组成和活性)等[1, 12, 38]。本研究重点关注生态恢复树种凋落叶性状与土壤激发效应之间的关联, 发现凋落叶性状与土壤激发效应之间没有显著相关关系。然而, 之前的研究发现, 在亚热带森林中, 凋落叶的碳相关化学性质是控制激发效应强度的主要因素[48], 这在一定程度上反映喀斯特生态系统植物凋落物诱导的激发效应及其控制因素可能与非喀斯特生态系统存在一定差异。下一步研究工作将进一步考虑喀斯特生态恢复树种凋落物分解诱导的激发效应及其控制因素。
4 结论本研究发现, 凋落叶性状, 诸如叶干物质含量、非结构性碳、水溶性碳、纤维素和锰含量能够较好地预测喀斯特生态恢复树种凋落叶的分解速度。然而, 这些特征与凋落叶分解诱导的土壤激发效应之间没有相关性。从生态系统物质循环与能量流动角度出发, 在植被恢复初期, 可以选择光皮梾木、海南椴、顶果木和降香黄檀等凋落叶分解速度较快的树种, 以促进土壤养分的归还与利用, 推动植被的恢复与发展。相反, 海南椴、任豆和割舌树等生态恢复树种凋落叶输入会抑制土壤有机碳分解, 即产生负激发效应, 可以作为植被恢复中后期优先选择的树种, 有利于退化生态系统土壤碳的固持。
| [1] |
Chen L Y, Liu L, Qin S Q, Yang G B, Fang K, Zhu B, Kuzyakov Y, Chen P D, Xu Y P, Yang Y H. Regulation of priming effect by soil organic matter stability over a broad geographic scale. Nature Communications, 2019, 10: 5112. DOI:10.1038/s41467-019-13119-z |
| [2] |
Jiang Z C, Lian Y Q, Qin X Q. Rocky desertification in Southwest China: impacts, causes, and restoration. Earth-Science Reviews, 2014, 132: 1-12. DOI:10.1016/j.earscirev.2014.01.005 |
| [3] |
王克林, 岳跃民, 陈洪松, 吴协保, 肖峻, 祁向坤, 张伟, 杜虎. 喀斯特石漠化综合治理及其区域恢复效应. 生态学报, 2019, 39(20): 7432-7440. DOI:10.5846/stxb201909051849 |
| [4] |
袁道先, 章程. 岩溶动力学的理论探索与实践. 地球学报, 2008, 29(3): 355-365. DOI:10.3321/j.issn:1006-3021.2008.03.009 |
| [5] |
Li D J, Wen L, Yang L Q, Luo P, Xiao K C, Chen H, Zhang W, He X Y, Chen H S, Wang K L. Dynamics of soil organic carbon and nitrogen following agricultural abandonment in a karst region. Journal of Geophysical Research: Biogeosciences, 2017, 122(1): 230-242. DOI:10.1002/2016JG003683 |
| [6] |
Ahmed A R, Pichler V, Homolák M, Gömöryová E, Nagy D, Pichlerová M, Gregor J. High organic carbon stock in a karstic soil of the Middle-European Forest Province persists after centuries-long agroforestry management. European Journal of Forest Research, 2012, 131(6): 1669-1680. DOI:10.1007/s10342-012-0608-7 |
| [7] |
郭屹立, 李冬兴, 王斌, 白坤栋, 向悟生, 李先琨. 北热带喀斯特季节性雨林土壤和6个常见树种凋落物的C、N、P化学计量学特征. 生物多样性, 2017, 25(10): 1085-1094. |
| [8] |
胡仪, 吴福忠, 吴秋霞, 康自佳, 岳楷, 杨玉盛, 倪祥银. 三个亚热带森林优势种凋落物非结构性碳水化合物含量的季节动态. 生态学报, 2022, 42(5): 1901-1910. DOI:10.5846/stxb202103050606 |
| [9] |
窦百强, 容丽, 俞国松, 李恒. 茂兰喀斯特森林自然保护区凋落叶分解动态. 水土保持通报, 2019, 39(2): 94-100. |
| [10] |
Bradford M A, Veen G F, Bonis A, Bradford E M, Classen A T, Cornelissen J H C, Crowther T W, De Long J R, Freschet G T, Kardol P, Manrubia-Freixa M, Maynard D S, Newman G S, Logtestijn R S P, Viketoft M, Wardle D A, Wieder W R, Wood S A, van der Putten W H. A test of the hierarchical model of litter decomposition. Nature Ecology & Evolution, 2017, 1(12): 1836-1845. |
| [11] |
Yang K, Zhu J, Zhang W, Lu D, Zhang Y, Zheng X, Xu S, Wang G G. Litter decomposition and nutrient release from monospecific and mixed litters: comparisons of litter quality, fauna and decomposition site effects. Journal of Ecology, 2022, 110(7): 1673-1686. DOI:10.1111/1365-2745.13902 |
| [12] |
Kuzyakov Y. Priming effects: interactions between living and dead organic matter. Soil Biology & Biochemistry, 2010, 42(9): 1363-1371. |
| [13] |
Perveen N, Barot S, Maire V, Cotrufo M F, Shahzad T, Blagodatskaya E, Stewart C E, Ding W X, Siddiq M R, Dimassi B, Mary B, Fontaine S. Universality of priming effect: an analysis using thirty five soils with contrasted properties sampled from five continents. Soil Biology & Biochemistry, 2019, 134: 162-171. |
| [14] |
Guenet B, Camino-Serrano M, Ciais P, Tifafi M, Maignan F, Soong J L, Janssens I A. Impact of priming on global soil carbon stocks. Global Change Biology, 2018, 24(5): 1873-1883. DOI:10.1111/gcb.14069 |
| [15] |
俞国松, 王世杰, 容丽, 冉景丞. 茂兰喀斯特森林主要演替群落的凋落物动态. 植物生态学报, 2011, 35(10): 1019-1028. |
| [16] |
Zhu X A, Zou X, Lu E F, Deng Y, Luo Y, Chen H, Liu W J. Litterfall biomass and nutrient cycling in karst and nearby non-karst forests in tropical China: A10-year comparison. Science of the Total Environment, 2021, 758: 143619. DOI:10.1016/j.scitotenv.2020.143619 |
| [17] |
卢晓强, 杨万霞, 丁访军, 丁晖, 吴军, 曹铭昌, 崔鹏, 徐海根. 茂兰喀斯特地区原始林凋落物量动态与养分归还. 生态与农村环境学报, 2014, 30(5): 614-619. |
| [18] |
曾昭霞, 王克林, 曾馥平, 宋同清, 刘孝利, 宋希娟. 桂西北喀斯特区原生林与次生林凋落叶降解和养分释放. 生态学报, 2012, 32(9): 2720-2728. DOI:10.5846/stxb201103240372 |
| [19] |
Liu C C, Liu Y G, Guo K, Zhao H W, Qiao X G, Wang S J, Zhang L, Cai X L. Mixing litter from deciduous and evergreen trees enhances decomposition in a subtropical karst forest in southwestern China. Soil Biology & Biochemistry, 2016, 101: 44-54. |
| [20] |
蔡道雄, 卢立华. 浅谈石漠化治理的对策及造林技术措施. 世界林业研究, 2002, 15(2): 76-80. DOI:10.3969/j.issn.1001-4241.2002.02.012 |
| [21] |
黄甫昭, 李冬兴, 王斌, 丁涛, 向悟生, 文淑均, 郭屹立, 李先琨. 喀斯特季节性雨林优势种群生态位特征及其对石漠化地区植被修复的启示. 广西科学, 2018, 25(5): 599-610. |
| [22] |
Sluiter A, Hames B, Ruiz R, Scarlata C, Sluiter J, Templeton D, Crocker D. Determination of structural carbohydrates and lignin in biomass. Laboratory Analytical Procedure, 2008, 1617: 1-16. |
| [23] |
Hagerman A E. The Tannin Handbook. Miami University, Oxford, OH, 2011.
|
| [24] |
巢林, 曾鑫, 欧梦菲, 黄忻婷, 张建兵, 刘艳艳. 喀斯特山区不同土地利用方式对土壤微生物生物量、酶活性及其化学计量的影响. 山地学报, 2023, 41(1): 28-40. |
| [25] |
Shi J C, Zhao B Y, Zheng S, Zhang X W, Wang X L, Dong W T, Xie Q J, Wang G, Xiao Y P, Chen F, Yu N, Wang E T. A phosphate starvation response-centered network regulates mycorrhizal symbiosis. Cell, 2021, 184(22): 5527-5540. DOI:10.1016/j.cell.2021.09.030 |
| [26] |
Sun X B, Li D J, Lü X T, Fang Y T, Ma Z L, Wang Z C, Chu C J, Li M M, Chen H. Widespread controls of leaf nutrient resorption by nutrient limitation and stoichiometry. Functional Ecology, 2023, 37(6): 1653-1662. DOI:10.1111/1365-2435.14318 |
| [27] |
陆姣云, 段兵红, 杨梅, 杨晗, 杨惠敏. 植物叶片氮磷养分重吸收规律及其调控机制研究进展. 草业学报, 2018, 27(4): 178-188. |
| [28] |
Chen H, Reed S C, Lü X T, Xiao K C, Wang K L, Li D J. Global resorption efficiencies of trace elements in leaves of terrestrial plants. Functional Ecology, 2021, 35(7): 1596-1602. DOI:10.1111/1365-2435.13809 |
| [29] |
Sun Z L, Tian P, Zhao X C, Wang Y P, Wang S Z, Fang X M, Wang Q K, Liu S E. Temporal shifts in the explanatory power and relative importance of litter traits in regulating litter decomposition. Forest Ecosystems, 2022, 9: 100072. DOI:10.1016/j.fecs.2022.100072 |
| [30] |
Chomel M, Guittonny‐Larchevêque M, Fernandez C, Gallet C, DesRochers A, Pare D, Jackson B G, Baldy V. Plant secondary metabolites: a key driver of litter decomposition and soil nutrient cycling. Journal of Ecology, 2016, 104(5): 1527-1541. |
| [31] |
Kazakou E, Vasseur F, Sartori K, Baron E, Rowe N, Vile D. Secondary metabolites have more influence than morphophysiological traits on litter decomposability across genotypes of Arabidopsis thaliana. New Phytologist, 2019, 224(4): 1532-1543. DOI:10.1111/nph.15983 |
| [32] |
Kazakou E, Violle C, Roumet C, Pintor C, Gimenez O, Garnier E. Litter quality and decomposability of species from a Mediterranean succession depend on leaf traits but not on nitrogen supply. Annals of Botany, 2009, 104(6): 1151-1161. DOI:10.1093/aob/mcp202 |
| [33] |
Tao J, Zuo J, He Z, Wang Y P, Liu J C, Liu W D, Cornelissen J H C. Traits including leaf dry matter content and leaf pH dominate over forest soil pH as drivers of litter decomposition among 60 species. Functional Ecology, 2019, 33(5): 1798-1810. |
| [34] |
Kazakou E, Vile D, Shipley B, Gallet C, Garnier E. Co-variations in litter decomposition, leaf traits and plant growth in species from a Mediterranean old-field succession. Functional Ecology, 2006, 20(1): 21-30. DOI:10.1111/j.1365-2435.2006.01080.x |
| [35] |
Poorter H, Niklas KJ, Reich PB, Oleksyn J, Poot P, Mommer L. Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control. New Phytologist, 2012, 193(1): 30-50. DOI:10.1111/j.1469-8137.2011.03952.x |
| [36] |
吴秋霞, 吴福忠, 胡仪, 康自佳, 张耀艺, 杨静, 岳楷, 倪祥银, 杨玉盛. 亚热带同质园11个树种新老叶非结构性碳水化合物含量比较. 植物生态学报, 2021, 45(7): 771-779. |
| [37] |
Chen H, Li D J, Mao Q G, Xiao K C, Wang K L. Resource limitation of soil microbes in karst ecosystems. Science of the Total Environment, 2019, 650: 241-248. DOI:10.1016/j.scitotenv.2018.09.036 |
| [38] |
Soong J L, Fuchslueger L, Marañon-Jimenez S, Torn M S, Janssens I A, Penuelas J, Richter A. Microbial carbon limitation: the need for integrating microorganisms into our understanding of ecosystem carbon cycling. Global Change Biology, 2020, 26(4): 1953-1961. DOI:10.1111/gcb.14962 |
| [39] |
吴小健, 石焱, 朱嘉宁, 孙麟钧, 马祥庆. 全球变暖对森林凋落物的影响. 世界林业研究, 2023, 36(1): 26-32. |
| [40] |
李强, 周道玮, 陈笑莹. 地上枯落物的累积、分解及其在陆地生态系统中的作用. 生态学报, 2014, 34(14): 3807-3819. DOI:10.5846/stxb201211271684 |
| [41] |
Peng Y, Li Y J, Song S Y, Chen Y Q, Chen G T, Tu L H. Nitrogen addition slows litter decomposition accompanied by accelerated manganese release: a five-year experiment in a subtropical evergreen broadleaf forest. Soil Biology and Biochemistry, 2022, 165: 108511. DOI:10.1016/j.soilbio.2021.108511 |
| [42] |
Chen G T, Sun Y, Chen Y Q, Chen Y Q, Ma W, Zhong Q, Li Y J, Qiu L J, Zhou S X, Chen G, Xie J L, Tu L H. Manganese indicates root decomposition rates across soil layer, root order, and tree species: Evidence from a subtropical forest. Soil Biology and Biochemistry, 2023, 181: 109023. DOI:10.1016/j.soilbio.2023.109023 |
| [43] |
Cornwell W K, Cornelissen J H C, Amatangelo K, Dorrepaal E, Eviner V T, Godoy O, Hobbie S E, Hoorens B, Kurokawa H, Pérez-Harguindeguy N, Quested H M, Santiago L S, Wardle D A, Wright I J, Aerts R, Allison S D, Van Bodegom P, Brovkin V, Chatain A, Callaghan T V, Díaz S, Garnier E, Gurvich D E, Kazakou E, Klein J A, Read J, Reich P B, Soudzilovskaia N A, Vaieretti M V, Westoby M. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecology Letters, 2008, 11(10): 1065-1071. DOI:10.1111/j.1461-0248.2008.01219.x |
| [44] |
Jiang L, Wang H, Li S, Fu X, Dai X, Yan H, Kou L. Mycorrhizal and environmental controls over root trait-decomposition linkage of woody trees. New Phytologist, 2021, 229(1): 284-295. DOI:10.1111/nph.16844 |
| [45] |
Sun T, Hobbie S E, Berg B, Zhang H G, Wang Q K, Wang Z W, Hättenschwiler S. Contrasting dynamics and trait controls in first-order root compared with leaf litter decomposition. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(41): 10392-10397. |
| [46] |
Fujii S, Cornelissen J H C, Berg M P, Mori A S. Tree leaf and root traits mediate soil faunal contribution to litter decomposition across an elevational gradient. Functional Ecology, 2018, 32(3): 840-852. DOI:10.1111/1365-2435.13027 |
| [47] |
贾丙瑞. 凋落物分解及其影响机制. 植物生态学报, 2019, 43(8): 648-657. |
| [48] |
Chao L, Liu Y Y, Freschet G T, Zhang W D, Yu X, Zheng W H, Guan X, Yang Q P, Cheng L C, Dijkstra F A, Wang S L. Litter carbon and nutrient chemistry control the magnitude of soil priming effect. Functional Ecology, 2019, 33: 876-888. DOI:10.1111/1365-2435.13278 |