 2024, Vol. 44
2024, Vol. 44文章信息
- 钱恬, 魏宇宸, 王明明, 郭晓伟, 罗忠奎
- QIAN Tian, WEI Yuchen, WANG Mingming, GUO Xiaowei, LUO Zhongkui
- 全球和生物群系尺度土壤有机碳分布控制因子及其分异规律
- Distinct controlling factors on the distribution of soil organic carbon stocks at global and biomes scales
- 生态学报. 2024, 44(8): 3382-3396
- Acta Ecologica Sinica. 2024, 44(8): 3382-3396
- http://dx.doi.org/10.20103/j.stxb.202212113558
-
文章历史
- 收稿日期: 2022-12-11
- 采用日期: 2023-06-24






土壤固碳是有效缓解全球气候变化的重要途径[1—2], 阐明其在不同空间尺度和不同土层深度上的环境影响因子, 是增加土壤碳汇和减少土壤碳源的重要理论基础。土壤的碳源、汇变化取决于植物碳输入和土壤有机碳(SOC)分解之间的净平衡, 这种输入与分解在自然因素和人为管理的影响下呈现出高度的空间异质性[3]。因此, 碳循环研究的关键问题之一是揭示不同空间(水平和垂直)尺度上SOC的环境影响因素。气候、土壤、植被、地形以及人类活动是影响SOC的五个重要影响因素, 大量研究表明, 这些SOC控制因子在不同的空间尺度上发挥不同的作用[4], 即存在SOC控制因子的尺度效应[5]。在较小尺度上(如小于几百平方米), SOC主要受土壤性质、地形和人类活动的控制[3];在流域尺度, 降水、地貌、下垫面和土地利用类型起主导作用[6—7];在更大的区域到全球尺度上, 气候则产生显著影响[8]。此外, 人类活动, 如土地利用和管理, 也调节着不同空间尺度上SOC的存储和再分配[9]。土壤剖面纵向尺度上, SOC主控因子也随深度发生显著变化[10]。有研究表明, 相较于气候, 表层土壤SOC与植被有更高的相关性, 但全剖面SOC的绝对量则主要受气候的调控[11—12];也有研究认为, 气候对不同深度土壤的SOC的调控没有显著差异[13]。
针对不同尺度上SOC控制因子, 研究往往需要建立因变量(即SOC)与多个环境协变量之间的关系, 但很少考虑空间自相关对结果的影响, 因此造成环境控制因子解释的不确定性。空间自相关是地理空间数据的基本特性, 即数据的空间距离越近、则数据越相似[14], 被认为是构建统计模型的不利条件。由于数据分布的不均匀性, 在数据稀疏区域, 这种空间自相关的空间结构无法被计算, 而如果假定空间数据独立[15], 却没有对自相关数据进行排除, 则同一特征间的空间自相关关系将被错误地理解为环境变量对因变量的解释能力[16], 从而引起模型的过拟合与外推能力的降低。因此, 想要提高全球SOC模型的拟合精度, 就需要利用一定的空间统计方法表征并筛除SOC的空间自相关性, 获得独立数据集后, 建立SOC与环境协变量间的模型。
为探寻排除自相关数据后不同尺度上各土壤深度SOC与环境影响因子的关系, 本文选用了全球113571个来自包括世界土壤信息服务(WoSIS)数据库[17]、北半球永久冻土区土壤数据集[4]、中国第二次土壤普查结果[18]等的土壤剖面SOC和理化性质测量数据, 评估各环境因子在全球尺度和生物群系尺度上对土壤有机碳密度(SOCd, kg/m3土壤)的影响差异。这些土壤数据经缺失值补充和土层标准化后, 设定不同的空间自相关数据筛选阈值, 结合对应气候、土壤、植被、地形与人类活动等38种环境协变量信息。然后根据数据所处的地理空间位置, 筛选出全球以及生物群系范围内的对应子集, 对每个子集使用基于机器学习的极致梯度提升决策树(XGBoost), 建立SOCd与环境协变量间的机器学习模型, 探讨空间自相关在不同尺度上对模型解释性和SOCd主控因子的影响, 分析SOCd与环境因子间的相关关系和SOCd在不同空间尺度上的主控因子差异。
1 材料与方法 1.1 土壤SOC数据在本研究中, 使用了WoSIS数据库[17]、冻土数据集[4]、以及中国土壤二普数据库[18]。WoSIS整理并管理了全球最大的土壤剖面观测数据库, 本研究选用2019年经过质量控制和标准化的数据集(ISRIC数据中心), 共包括110856个剖面和471301个土层。冻土SOC数据集由Mishra等人汇编整理, 包括0—0.3 m, 0—1 m的2530个剖面和1—2 m的875个剖面, 这些冻土数据弥补了高纬度地区土壤数据的不足[4]。中国土壤二普数据共2473个剖面、8749个土层。本研究对土壤数据进行进一步筛选, 排除重复和有明显错误的SOC数据(SOC, g C/kg soil)(如矿质土层深度值为负、深层深度值小于上层深度值、SOC大于1000等)的土层, 随后基于公式计算矿质土壤SOCd:
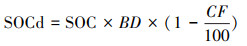
|
(1) |
式中, BD是细土(土粒直径 < 2mm)的容重(kg/dm3), CF是土壤中砾石(土粒直径>2 mm)含量的体积百分比(%)。
为尽可能利用所得SOC测量值, 需要对BD和CF的缺失值进行补充。以BD为例:把除SOC、土壤质地、BD的其它土壤理化性质(WoSIS提供的32个数值型土壤理化信息, 如pH、土壤阳离子交换量(CEC)等)作为协变量, 从存在BD值的数据集中随机重采样(Boostrap), 获得80%样本量的新子集, 这个子集被用于XGBoost统计预测模型的训练, 以上采样与建模过程独立重复50次, 且每次都使用对应模型与训练集数据进行模型精度(用R2来表示)验证(图 1展示了其中一次重复的BD和CF的模型精度验证图), 得到BD的模型预测精度R2=0.85, CF的模型预测精度R2=0.70。在对BD缺失值进行预测时, 每个土层样点都会基于土壤理化性质实测值与上述方法中建立的50个模型得到50个预测结果, 本研究仅保留预测结果变异系数小于15%的结果的平均值。CF缺失值的补充方法与之一致。随后基于公式(1)计算各土层SOCd。

|
| 图 1 BD和CF的模型精度验证图 Fig. 1 Model accuracy verification of BD and CF BD:细土(土粒直径 < 2mm)的容重(kg/dm3);CF:土壤中砾石(土粒直径>2 mm)含量的体积百分比(%) |
土壤剖面数据集存在的另一问题是采集土层深度不一, 这里使用等面积二次平滑样条函数[19—21]对剖面SOCd进行回归, 使用均方根误差获取每个土壤剖面样条回归函数的最佳参数, 并将结果统一到0—5、5—15、15—30、30—60、60—100、100—200 cm这6个标准深度土层。当剖面深度高于某标准土层的上限时, 该层以下的不做计算(如某一剖面的采样深度为0—70 cm, 则不计算100—200 cm土层的SOCd), 最后合计得到113571个土壤剖面数据(图 2)。

|
| 图 2 土壤剖面样点在全球各生物群系的分布 Fig. 2 Location of soil profiles across globe biomes |
这些土壤剖面涵盖了9类主要的生物群系类型, 包括热带/亚热带森林(TSF)、热带/亚热带草原(TSG)、温带森林(TF)、温带草原(TG)、地中海/山地灌木林(MS)、北方森林(BF)、冻原(Tundra)、沙漠(Deserts)和农田(Croplands)[22], 其土壤剖面分布与数量见表 1。
| 生物群系类型 Biome types |
各层土壤样点计数Counts of soil samples in each layer | 总计 Total |
|||||
| 0—5 cm | 5—15 cm | 15—30 cm | 30—60 cm | 60—100 cm | 100—200 cm | ||
| 热带/亚热带森林 Tropical/Subtropical Forests |
7345 | 7023 | 6544 | 6137 | 5053 | 2660 | 34762 |
| 热带/亚热带草原 Tropical/Subtropical Grasslands |
10192 | 8908 | 7607 | 7102 | 6155 | 3505 | 43469 |
| 温带森林Temperate Forests | 9575 | 9660 | 8960 | 8699 | 7805 | 5586 | 50285 |
| 温带草原Temperate Grasslands | 4024 | 3987 | 3892 | 3463 | 3152 | 2532 | 21050 |
| 地中海/山地灌木 Mediterranean/montane Shrublands |
5917 | 5844 | 5678 | 5518 | 5098 | 3678 | 31733 |
| 北方森林Boreal Forest | 1094 | 1520 | 1411 | 1455 | 1141 | 586 | 7207 |
| 苔原Tundra | 322 | 385 | 271 | 243 | 216 | 109 | 1546 |
| 沙漠Deserts | 3240 | 3741 | 4154 | 3708 | 3325 | 2144 | 20312 |
| 作物Croplands | 19083 | 18230 | 17401 | 16336 | 15471 | 11401 | 97922 |
| 总计Total | 60792 | 59298 | 55918 | 52661 | 47416 | 32201 | 308286 |
本研究使用经纬度定位剖面点对应的环境协变量, 这些变量包括以温度与降水为主的19种生物气候因子[23], 基于自动化地球科学分析系统(SAGA GIS)和全球高程数据[23]计算得到的13种地形, 以及2种植物光谱数据[24]、人口[25]与生产总值[26]等2种人类活动信息, 土壤饱和渗透系数[27]和土壤母质信息(表 2)。其中, 土壤饱和渗透系数(Ks)是基于土壤水力参数的物理背景、利用土壤质地推演出的一项土壤水力参数, 它仅以砂、粉砂、粘土的百分比含量作为预测输入[28], 所以在研究中被认为是一项与土壤质地相关联的土壤性质。
| 变量类型 Variable types |
代码 Codes |
解释 Explanation |
时空分辨率 Resolution |
来源 Sources |
| 气候Climate | aT | 年均温Mean Annual Temperature | 1970—2000, 30 arcsecond | WorldClim, https://www.worldclim.org/data/bioclim.html |
| ragTm | 平均日较差Mean Diurnal Range (Mean of monthly (max temp-min temp)) |
|||
| isoT | 等温性 Isothermality (BIO2/BIO7) (×100) |
|||
| sdT | 温度季节性 Temperature Seasonality (sd ×100) |
|||
| maxThm | 最暖月最高温 Max temperature of warmest month |
|||
| minTcm | 最冷月最低温 Min temperature of coldest month |
|||
| ragTa | 温度年度范围 Temperature annual range (BIO5-BIO6) |
|||
| mTwq | 最湿季均温 Mean temperature of wettest quarter |
|||
| mTdq | 最旱季均温 Mean temperature of driest quarter |
|||
| mThq | 最暖季均温 Mean temperature of warmest quarter |
|||
| mTcq | 最冷季均温 Mean temperature of coldest quarter |
|||
| aP | 年均降水Annual precipitation | |||
| Pwm | 最湿月降水 Precipitation of wettest month |
|||
| Pdm | 最旱月降水 Precipitation of driest month |
|||
| cvP | 降水季节性 Precipitation Seasonality (Coefficient of Variation) |
|||
| Pwq | 最湿季降水 Precipitation of wettest quarter |
|||
| Pdq | 最旱季降水 Precipitation of driest quarter |
|||
| Phq | 最暖季降水 Precipitation of warmest quarter |
|||
| Pcq | 最冷季降水 Precipitation of coldest quarter |
|||
| 地形Terrian | TWI | 地形湿度指数Terrian wetness index | 2017, 1 km | 基于高程数据(https://www.worldclim.org/data/bioclim.html, wc2.1_30s_elev.tif), 利用自动化地球科学分析系统(SAGA GIS)计算。 |
| CArea | 集水区Catchment Area | |||
| ModCArea | 优化集水区Modified-catchment area | |||
| Flowpath | 水流路径Flowpath | |||
| TRI | 地形粗糙度指数Terrain ruggedness index | |||
| TPI | 地形位置指数Terrian position index | |||
| Roughness | 粗糙度Roughness | |||
| Slope | 坡度Slope | |||
| Aspect | 坡向Aspect | |||
| FlowDir | 流向Flow Direction | |||
| CPlanform | 平面形式曲率Planform curve | |||
| CProfile | 轮廓曲率Profile curve | |||
| CMcnab | 麦克纳布曲率Mcnab curve | |||
| 植被 Vegetation |
NPP | 净初级生产力Net primary productivity | 1982—2011年, 1 km | 中分辨率成像光谱仪(MODIS), https://lpdaac.usgs.gov/products/mod17a2hv061/ |
| SIF | 日光诱导叶绿素荧光 Solar-induced chlorophyll fluorescence |
2017—2019年, 2 km | 全球二氧化碳监测科学实验卫星 (TanSAT)[23] |
|
| 人类 Human activites |
Pop | 人口Population | 1990—2015, 30 arcsecond | 全球环境历史数据库(HYDE), https://www.pbl.nl/en/image/links/hyde https://www.pbl.nl/en/image/links/hyde |
| GDP | 地区生产总值Gross domestic product | 0—2000, 30 arcsecond | 数据知识库(Dryad), https://datadryad.org/stash/dataset/doi:10.5061/dryad.dk1j0 | |
| 土壤 Soil |
Ks | 土壤饱和渗透系数 Saturated Hydraulic Conductivity |
1 km | Zhang et al.[27] |
| SPM | 土壤母质Soil parent material | FAO, https://www.fao.org/land-water/land/land-governance/land-resources-planning-toolbox/category/details/en/c/1026564/ |
除此之外, 生物群系划分图被用于各尺度上SOCd分布的控制因子及其分异规律的研究, 它是由MODIS土地覆盖图[29]和世界陆地生态区[30]聚合而成的生物群系空间层, 以代表九种生物群系类型(图 2)。
1.3 自相关数据筛选为分析自相关数据对模型解释性和外推性的影响, 研究首先基于剖面的空间信息与SOCd值计算每个土层样点的区域莫兰指数(I), 公式为:
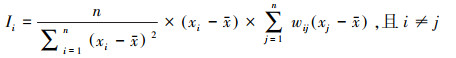
|
(2) |
式中, xi和xj是相同土层内样点i和j的SOCd值, x和n分别表示研究区域内样点的SOCd平均值和个数, w是标准化的空间权重值。I范围在-1到1之间, 当I显著> 0时, 该样点存在空间正自相关, 当I显著 < 0时, 该样点存在空间负自相关。设立H0假设:空间样点是随机分布的, 然后计算每个剖面土层的临界值(Z得分), 获得与之对应的自相关显著性水平(P值):
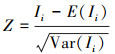
|
(3) |
式中, P值表示H0正确时的概率, P值越接近0, 则说明该样点空间自相关性越强。本研究设定, 当P值≤0.1时, 该数据存在空间自相关。
1.4 分析方法本文采用多边形连接模型与土壤—景观模型相结合的思路[18], 即根据生物群系类型对全球所有数据进行划分, 再对每一个子集以及全球总数据集, 都分别根据土壤剖面实测数据和对应的气候、土壤、植被、地形、人类活动等环境控制因子, 获取全球尺度与生物群系尺度上的SOC统计模型[31]。为研究全球尺度上空间自相关对统计模型准确性的影响, 利用现有的6个标准土层上的SOCd-环境协变量数据集, 分别进行子数据集提取以及训练、验证集划分, 包括如下步骤:
(a) 所有数据集记为P0, 表示不剔除自相关。P0同时作为训练集与验证集(这个验证集里的环境协变量需要基于训练模型再次预测, 并与实测数据进行对比。下文所述验证集的用途与之一致);
(b) 所有P值>0.1的数据集记为P1, 即剔除自相关后的P0的子集。P1同时作为训练集与验证集;
(c) 取P0, 遵循8∶2原则随机划分出训练集(P0—8)与验证集(P0—2);
(d) 取P1, 遵循8∶2原则随机划分出训练集(P1—8)与验证集(P1—2);
(e) 选用步骤d中的验证集(P1—2), P0减去该验证集的剩余所有数据作为训练集(P0—P1—2), 这一步是为了检测自相关数据对统计模型外推性能的影响。
研究进一步探寻排除空间自相关后不同尺度上SOC控制因子的变化:
(f) 全球尺度上的研究数据集与步骤d一致;
(g) 生物群系尺度上, 首先取P1, 根据9个生物群系类型提取子集, 每个子集遵循8∶2原则随机划分出训练集与验证集。
在此筛选基础上, 使用每一子集的训练数据建立XGBoost模型, 对应验证集被用于模型R2的计算。每个协变量的相对重要性由模型给出, R2与相对重要性的乘积即为该协变量对SOCd变异性的解释率。
在这里, XGBoost是一种使用梯度手段进行多棵分类决策树的迭代增强算法, 它对每一次迭代的残差进行泰勒展开, 并加上正则化项作为目标函数, 可以在集成中避免过拟合现象。XGBoost能够对高维变量进行高效处理, 同时也能对缺失特征分裂的方向进行学习。本研究使用XGBoost模型, 获取每个子集SOCd与该空间位置的气候、土壤、植被、地形与人类活动等5项环境协变量的关系:
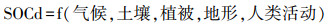
|
(4) |
10折交叉验证被用以优化与约束XGBoost模型参数。为了评估样点分布不均、缺失BD和CF插补以及土层深度标准化所导致的不确定性, 以上所有训练与验证集划分、模型建立与评价都基于不同的随机数种子独立重复25次。在相同的空间尺度与土层深度上, 步骤a与b计算得到的R2差异, 可以表征空间自相关数据对模型解释性的影响, 这里的解释性指得是模型拟合数据集内部关系的能力, 不包括模型外推性。在相同土层上, 步骤d与e计算得到的R2差异, 可以表征空间自相关数据对模型外推性能的影响。在相同土层上, 步骤c与e计算得到的R2差异, 可以表征如果不经自相关筛选, 模型的外推能力会被多大程度的高估。研究汇总了不同生物群系与标准土层上各环境类型对SOCd的解释程度, 以及模型中的主控环境协变量及其在解释模型中的重要性占比, 这里及下文提及的重要性占比都是指该变量对SOCd的解释率相对占比。
2 结果与分析 2.1 全球尺度土壤有机碳分布的控制因子在未剔除空间自相关时, 基于气候、土壤、植被、地形和人类活动等五项环境因素, XGBoost模型在全球尺度上的6个标准土层中由上至下分别解释64%、61%、61%、59%、53%、55%的SOCd空间变异。当对训练数据进行空间自相关筛除后, 模型对训练集的解释性显著降低, 在由上至下的标准土层中分别降低了16%、19%、16%、20%、17%、13%。当这些模型被用于外推预测时, 排除空间自相关数据不能显著提高模型对外部独立数据集的解释率, 平均只能解释各土层27%、26%、27%、24%、22%和19%的变异。值得注意的是, 当选用独立验证集进行外推验证、但没有在验证集中筛除自相关变量时, 会导致模型过拟合, 而且随土壤深度的增加, 空间自相关对模型解释率造成的误差变大(图 3)。

|
| 图 3 空间自相关对全球尺度土壤有机碳模型解释性与外推性的影响(R2) Fig. 3 The influence of spatial autocorrelation on model performance (R2) in terms of interpretability and extrapolation at the global scale a—e:对应1.4中的分类步骤; a:同时使用P0作为训练和验证数据集;b:同时使用P1作为训练和验证数据集;c:使用P0—8为训练集、P0—2为验证集;d:使用P1—8为训练集、P1—2为验证集;e:使用P0—P1—2为训练集、P1—2为验证集 |
图 4比较了全球尺度上、不同自相关筛选条件下5种环境类型对SOCd的绝对影响。在排除自相关后, 气候类变量为SOCd贡献了17%—30%的空间变异。随土壤深度的增加, 气候在模型所有环境变量中的重要性占比不断上升, 但模型对SOCd的解释率随之下降, 导致气候对SOCd的绝对解释程度没有发生显著波动。图 5展示了具体环境协变量对SOCd的相对重要性。在各项气候型环境协变量中, 剖面所处位置的温度和降水随时间序列的变异程度与变化范围是较为重要的SOCd变异贡献变量, 如温度季节性、温度年际范围、降水季节性等。除此之外, 最暖季均温和最冷季降水也是重要的环境影响因子。在表层0—30 cm的土壤中, 土壤性质的重要性仅次于气候, 约解释了15%—21%的SOCd空间变异, 在所有考虑到的土壤性质中, 最重要的是土壤饱和渗透系数(Ks), 其相对重要性约为25%—30%。但随土壤深度的下降, 土壤性质的相对重要性降低。综合各个变量的相对重要性, 地形变量的总体影响在各土层中相对一致, 在未筛除自相关变量时约为9%, 筛除后约为10%。植被与人类活动对SOCd的影响较小, 合计仅占5%—10%, 但是人口和叶绿素荧光指数的贡献却在所有环境协变量中相对重要性占比很高。

|
| 图 4 全球尺度上自相关筛选及5种类型的环境变量对SOCd的总体相对影响 Fig. 4 The overall relative influence of five environmental variables on SOCd with and without controlling spatial autocorrelation at the global scale SOCd:土壤有机碳密度Soil organic carbon density |

|
| 图 5 全球尺度上环境协变量解释SOCd空间变异的相对重要性 Fig. 5 The relative importance of individual environmental variables for influencing SOCd with and without controlling spatial autocorrelation at the global scale RD:原始数据, 即保留空间自相关;SAC:筛除空间自相关;aT:年均温;ragTm:平均日较差;isoT:等温性;sdT:温度季节性;maxThm:最暖月最高温;minTcm:最冷月最低温;ragTa:温度年度范围;mTwq:最湿季均温;mTdq:最旱季均温;mThq:最暖季均温;mTcq:最冷季均温;aP:年均降水;Pwm:最湿月降水;Pdm:最旱月降水;cvP:降水季节性;Pwq:最湿季降水;Pdq:最旱季降水;Phq:最暖季降水;TWI:地形湿度指数;CArea:集水区;ModCArea:优化集水区;Flowpath:水流路径;TRI:地形粗糙度指数;TPI:地形位置指数;Roughness:粗糙度;Slope:坡度;Aspect:坡向;FlowDir:流向;CPlanform:平面形式曲率;CProfile:轮廓曲率;CMcnab:麦克纳布曲率;NPP:净初级生产力;SIF:日光诱导叶绿素荧光;Pop:人口;GDP:地区生产总值;Ks:土壤饱和渗透系数;SPM:土壤母质;Pcq:最冷季降水 |
总体上, 气候、土壤与地形是影响全球尺度上SOCd空间分布最为关键的控制因子。无论是否排除空间自相关数据, 气候、土壤与地形在所有环境协变量内部的总体相对重要性占比都在80%以上。排除空间自相关后, 这三者对SOCd的总体绝对解释率在30%以上。随土壤深度的增加, 土壤特性的影响减小, 而气候和地形的影响没有显著变化。
2.2 生物群系尺度土壤有机碳分布的控制因子基于五类环境影响因子, 在考虑空间自相关的情况下, XGBoost模型在9种不同生物群系类型和6个标准土层深度上解释了45%—77%的SOCd空间分布变异(图 6)。其中, 苔原土壤中XGBoost模型的平均解释比例高达77%。温带森林和温带草原的被解释程度相对较低, 为46%和47%。

|
| 图 6 模型对不同生物群系和土层深度的SOCd的解释性 Fig. 6 Model performance among different biomes and soil layer depths |
当对自相关数据进行排除后, 在热带/亚热带的森林与草原中, 气候是最主要的环境影响类型, 约能解释24%—28%的SOCd变异;紧随其后的是土壤, 约为14%—18%;地形贡献了11%—12%的解释性;植被与人类活动的重要性仅有3%—5%。虽然气候是最重要的环境影响因素, 但在具体环境协变量中, Ks的相对重要性最高, 在0—30 cm土层中, 森林和草原中Ks重要性占比都约为24%, 但在更深层土层中(30—100 cm), 土壤性质对草原SOC的影响更大, 达到30%。在温带森林和草原、地中海/山地灌木林、沙漠和农田中, 气候依然是最主要的环境变量, 其重要性占比分别为25%、25%、26%、33%和31%, 地形紧接其后, 约为11%、14%、12%、13%和10%。在冻土苔原中, 气候、地形和植被分别对模型提供了37%、24%和10%的解释占比(图 7)。在0—5 cm的土层中, 地形轮廓曲率的重要性高达16%, 它可以用以衡量植被生长的难易程度和土壤水分流失情况。苔原的温度极值情况也反应了SOCd的水平, 在0—30 cm的土层中, 最暖季平均温度对SOCd的贡献率约为12%, 且与之呈负相关关系(图 8)。北方森林尤为特殊, 因为这里影响SOCd的主要环境变量并非气候而是地形, 在0—15 cm土层中, 地形与气候的贡献分别约为33%和18%, 在15—30 cm土层中, 地形的相对重要性高达43%, 而气候只占14%;在深层土层中(100—200 cm), 气候的重要性反而增加(图 7)。人类活动也是一项非常重要的SOC影响变量。在全球尺度下, 人口的相对重要性占比约为11%—13%, GDP为3%—7%, 而在生物群系尺度, GDP的影响占比则高于人口密度, 且在地中海/山地灌木生物群系类型中, GDP对表层土壤SOCd的空间变异解释了6.5%。

|
| 图 7 各生物群系与标准土层上环境协变量对SOCd的绝对解释率 Fig. 7 Influence of environmental covariates to SOCd across biomes and soil layer depths 图中数字:绝对解释率(%);TSF:热带/亚热带森林;TSG:热带/亚热带草原;TF:温带森林;TG:温带草原;MS:地中海/山地灌木;BF:北方森林;Tundra:苔原;Deserts:沙漠;Croplands:作物 |

|
| 图 8 各生物群系与标准土层上SOCd主控因子的相对重要性 Fig. 8 Relative influence of the most important six individual environmental covariates across biomes and soil layer depths 图中数字:主控因子在模型中的相对重要性(%) |
空间自相关是空间生态环境大数据的典型特征, 对土壤有机碳而言, 本文的研究发现, 空间自相关数据显著提高不同尺度上SOCd模型的总体解释率, 但也给模型带来多方面的误差:1)相邻SOCd数据间的自相关特征使损失函数向聚类样本偏移, 从而增加模型在信息稀疏区域的误差;2)部分自相关数据提供的有效信息被错误地分配给环境协变量, 且高自相关区域内的主控因子会获得更高的解释率提升;3)模型不能对SOCd-环境协变量间的统计特征与空间自相关特征进行有效融合, 这也导致了数据的空间自相关特征信息的丢失, 随尺度变小, 这种误差会越来越严重。
消除空间自相关影响的方法之一是直接删去那些空间自相关的数据, 除此之外还可以通过迭代残差分析等方式[32—33]。本研究在全球尺度上对空间自相关数据进行了筛选, 并在此基础上, 进一步探讨了在所有数据都是空间独立的前提下, 尺度差异对模型解释率的影响。研究结果发现, XGBoost模型对生物群系尺度的SOCd空间变异的解释率显著高于全球尺度。模型解释性随尺度降低而升高的可能原因之一是:尽管本研究基于P值对数据进行了自相关性检验和数据筛除, 但在分区域进行莫兰指数计算时, 与全球范围相比, 每个生物群系区域的边缘与面积比更大, 因此其边缘效应被更加地放大, 一些在全球尺度空间自相关筛除时会被删去的边缘数据得以保留, 证据是各生物群系尺度下的R2与全球尺度RD的R2更加接近(图 3)。另一原因是, 环境协变量以及土壤理化性质的变异程度在不同尺度上存在差异, 随尺度降低, 土壤变异程度降低, 使得模型更容易捕捉数据之间的非线性关系[34—35]。
3.2 不同尺度土壤有机碳分布的控制因子差异SOCd的控制因子及其解释率在不同尺度、不同土层显著不同[36]。本研究发现, 在深层土层中(60—200 cm), SOCd的解释率得到了提高, 尤其是在苔原、热带/亚热带森林和温带森林中, 解释率都增加了约12%。北方森林全剖面土壤SOC的解释率与土壤深度成正相关, 细究其环境类型变量的相对重要性可以发现, 随土壤深度的增加, 气候型变量的解释率占比并没有显著变化, 而地形、植被、人类活动与土壤性质的占比有着不同程度的提高, 这说明在未来深层SOC的研究中, 应该更多考虑非气候型环境协变量的影响。
不同生物群系类型中, SOCd分布的控制因子变化及其随土壤深度的变化也存在较大差异。在热带/亚热带森林, 反应植物生长状态的NPP与SIF, 以及决定植物生长发育的气候和土壤因素对SOCd变异的解释率最高。在热带/亚热带草原, 随深度增加, SOCd主控因子由气候转向土壤性质, 这是可能出于草地植被根系的分布强烈影响土壤有机碳在土壤剖面的垂直分布格局, 因此草地深层土壤中空气与水分的可获取性强烈支配了SOC的含量;Schuur等的研究同样证实了SOC储量的变异性有时更受氧化还原和氧气可用性的影响[37]。在北方森林和苔原, 温度对SOCd的解释率低, 这可能是因为低于阈值的低温导致由温度变化推动的SOC周转速率变异不大, 但苔原因为缺乏植被的保护, 土壤侵蚀程度与SOC暴露流失易受降水的影响[38], 从而提高气候变量的总体重要性。在苔原地区, 降水型变量对SOC影响的重要性随土层加深逐步上升, 且降水量的变异程度越高, SOC损失越大, 这可能是因为降水导致有机质通过淋溶、侵蚀等方式流失[3], 但也有利于地表植物源碳向土壤的输入, 表层SOC的流失更大程度上被植物碳源的输入抵消。
人类活动也是一项非常重要的SOCd影响因素, 但因其调控机制复杂多样、活动内容与强度难以评价、动态性最高[39], 目前相关研究中尚没有统一指标, 这将导致对深层土壤有机碳储量的低估[40]。在全球尺度上, 人口的影响更加强烈, 而在具体生物群系内部, 地区生产总值(GDP)的重要性更高。这其实是一个非常明显的尺度效应现象。在世界范围内, 人口密度能够反应当地人类活动的强弱, 是一个低频发生的变量, 而生产方式受限于气候与历史发展的影响, 因此即使是相同的生产强度, 生产方式差异所带来的高频GDP变化也是显著的, 从而GDP在大尺度统计中的贡献能力被掩盖。除此之外, 土地利用变化、土壤管理措施等变量已被证明对增加SOC储量起着重要作用[41], 但在本研究中没有被使用。
3.3 不同尺度土壤有机碳与环境协变量间的相关关系研究通过计算SOCd与环境协变量之间的相关系数发现(如图 9所示):整体上, 各生物群系尺度上的SOCd与气候、土壤、植被、地形与人类活动等环境因素的关系与全球尺度上的趋于一致, 都具有显著相关性。温度的变异程度以及绝大多数降水相关的环境协变量为正相关, 温度均值、极值与最旱季降水量为负相关。在全球尺度上, 高纬度土壤中SOC的储量更高[42];但在生物群系尺度上, 尤其是在温带森林中, 纬度与SOCd呈负相关。地形协变量与SOC的相关性较弱, 只有水流路径、地形粗糙度指数、粗糙度、坡度等相关性较为显著。关于地形因素对SOC影响程度并没有普遍的一致意见, 一些研究表明[43], 从全球、亚大陆到区域尺度, 地形位置、坡度和坡向对SOC变异的影响较小, 这与本研究的结果存在一定不同;但也有研究发现在大于100 m的尺度上, 地形因素较为重要[44]。植被因素中的净初级生产力、人类活动中的GDP与SOC的相关性也较好, 在全球尺度表层土壤中(0—15 cm)的相关系数分别为0.20和0.18, 其中植被通过控制碳的输入影响SOC储存, 被认为是预测表层土壤中SOC的最好指标[45]。

|
| 图 9 各生物群系与标准土层上SOCd与环境协变量的相关关系系数 Fig. 9 Correlation coefficients between SOCd and environmental covariates across different biomes and soil layer depths Globe:全球 |
研究通过分析自相关训练数据对模型的影响发现:无论是否考虑空间自相关, 模型难以基于训练数据挖掘外部独立区域SOCd的空间变异性, 因此在做SOC预测时, 必须严格规定适用范围。本研究模型拟合结果表明, 随自相关数据的排除, 环境协变量的解释贡献降低了13%—20%不等, 这揭示了高空间自相关性数据对全球SOCd分布做出的解释。在不同尺度利用统计模型进行SOC的计算时, 都需要讨论SOC与控制因子的关系。因此, 出于数据来源和尺度降级策略的不同, 研究者的结果也各不相同[35, 46]。辅助变量和数据源的分辨率、预测模型的误差、置信空间的选择等因素也都需被考虑在内, 但本文无涉及。
统计模型本身也存在着一些缺陷。首先, 土壤属性尤其是粘土比例、微生物活动以及植物输入分配情况与SOC分布密切相关[39], 而它们恰好是统计模型难以直接获取的数据。另一方面, SOCd预测需要运用到深度与水平两个方向上的方法学, 而深度上SOCd的预测与补齐仍处于黑箱模型与平滑回归(如Bishop等提出的等面积平滑样条法[20])相结合的阶段, 与水平预测的输入数据及方法学相对独立, 这就导致了纵向尺度的误差被带入到了全球尺度的预测中。除此之外, 随SOC协变量的不断更新, 出于黑箱模型的不可解释性, 研究区SOC统计模型需要被重新建立[19—21], 新增协变量也无法对已有结果做出直接修正。因此, 对不同空间尺度的各种土壤和环境因素的机制机理层面的分析, 对于探究它们对空间尺度SOC变化具有重要意义, 而本研究仅进行了统计分析, 从而限制了模型的随时间变化与协变量修改的动态更新能力。
4 结论通过对空间自相关的控制, 本研究发现:全球约有13%—20%的SOCd变异可以被实测数据的自相关性所解释。排除自相关数据后, 气候、土壤、植被、地形和人类活动等五项环境影响因子驱动的XGBoost模型, 在全球尺度0—5、5—15、15—30、30—60、60—100、100—200 cm的标准土层中分别只能解释27%、26%、27%、24%、22%和19%的SOCd变异。如果不考虑自相关, 这些因子的解释性将被高估。在9种不同生物群系类型下解释了45%—77%的SOCd变异。基于控制自相关后的研究结果, 发现:SOCd的环境主控因子在不同尺度和深度上都存在不同。在全球尺度上, 气候对SOCd变异的解释率更高, 而在生物群系尺度上, 气候、土壤与地形都具有较高的解释性。在不同空间尺度上, SOC呈现出明显的主控因子差异:在全球尺度上, 气候起决定性作用, 而在生物群系尺度上, 大多数生物群系类型中气候的解释率最高, 但地形与土壤属性在苔原和北方森林中起决定性作用。在土壤全剖面(0—2 m)中, 表层和底层SOC的控制因子也存在显著差异。为了更加有效的管理土壤有机碳和预测土壤有机碳变化, 必须考虑土壤有机碳控制因子在不同空间尺度和深度上的差异, 这能为合理利用土壤资源、促进土壤碳汇和减少土壤碳排放提供科学依据, 对更好地制定相关管理和保护SOC政策并确保土壤资源的可持续利用也至关重要。
| [1] |
Köchy M, Hiederer R, Freibauer A. Global distribution of soil organic carbon-Part 1: Masses and frequency distributions of SOC stocks for the tropics, permafrost regions, wetlands, and the world. SOIL, 2015, 1: 351-365. DOI:10.5194/soil-1-351-2015 |
| [2] |
Lehmann J, Solomon D, Kinyangi J, Dathe L, Wirick S, Jacobsen C. Spatial complexity of soil organic matter forms at nanometre scales. Nature Geoscience, 2008, 1(4): 238-242. DOI:10.1038/ngeo155 |
| [3] |
Wiesmeier M, Urbanski L, Hobley E, Lang B, von Lützow M, Marin-Spiotta E, van Wesemael B, Rabot E, Ließ M, Garcia-Franco N, Wollschläger U, Vogel H J, Kögel-Knabner I. Soil organic carbon storage as a key function of soils-A review of drivers and indicators at various scales. Geoderma, 2019, 333: 149-162. DOI:10.1016/j.geoderma.2018.07.026 |
| [4] |
Mishra U, Hugelius G, Shelef E, Yang Y H, Strauss J, Lupachev A, Harden J W, Jastrow J D, Ping C L, Riley W J, Schuur E A G, Matamala R, Siewert M, Nave L E, Koven C D, Fuchs M, Palmtag J, Kuhry P, Treat C C, Zubrzycki S, Hoffman F M, Elberling B, Camill P, Veremeeva A, Orr A. Spatial heterogeneity and environmental predictors of permafrost region soil organic carbon stocks. Science Advances, 2021, 7(9): eaaz5236. DOI:10.1126/sciadv.aaz5236 |
| [5] |
Li J Q, Pei J M, Pendall E, Fang C M, Nie M. Spatial heterogeneity of temperature sensitivity of soil respiration: a global analysis of field observations. Soil Biology and Biochemistry, 2020, 141: 107675. DOI:10.1016/j.soilbio.2019.107675 |
| [6] |
Fu B J, Chen L D, Ma K M. The effect of land use chang on the regional environment in the yangjuangou catchment in the loess plateau of China. Acta Geographica Sinica, 1999, 54(003): 241-246. |
| [7] |
姚丽贤, 周修冲, 蔡永发, 陈婉珍. 不同采样密度下土壤特性的空间变异特征及其推估精度研究. 土壤, 2004, 36(5): 538-542. DOI:10.3321/j.issn:0253-9829.2004.05.013 |
| [8] |
Ruiz-Colmenero M, Bienes R, Eldridge D J, Marques M J. Vegetation cover reduces erosion and enhances soil organic carbon in a vineyard in the central Spain. CATENA, 2013, 104: 153-160. DOI:10.1016/j.catena.2012.11.007 |
| [9] |
Amundson R, Berhe A A, Hopmans J W, Olson C, Sztein A E, Sparks D L. Soil science. Soil and human security in the 21st century. Science, 2015, 348(6235): 1261071. DOI:10.1126/science.1261071 |
| [10] |
Luo Z K, Viscarra-Rossel R A, Qian T. Similar importance of edaphic and climatic factors for controlling soil organic carbon stocks of the world. Biogeosciences, 2021, 18: 2063-2073. DOI:10.5194/bg-18-2063-2021 |
| [11] |
Jenkinson D S, Coleman K. The turnover of organic carbon in subsoils. Part 2. Modelling carbon turnover. European Journal of Soil Science, 2008, 59(2): 400-413. DOI:10.1111/j.1365-2389.2008.01026.x |
| [12] |
Jobbágy E G, Jackson R B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecological Applications, 2000, 10(2): 423-436. DOI:10.1890/1051-0761(2000)010[0423:TVDOSO]2.0.CO;2 |
| [13] |
Hicks Pries C E, Castanha C, Porras R C, Torn M S. The whole-soil carbon flux in response to warming. Science, 2017, 355(6332): 1420-1423. DOI:10.1126/science.aal1319 |
| [14] |
Tobler W R. A computer movie simulating urban growth in the Detroit region. Economic Geography, 1970, 46(sup1): 234-240. |
| [15] |
Pontius R G, Cornell J D, Hall C A S. Modeling the spatial pattern of land-use change with GEOMOD2: application and validation for Costa Rica. Agriculture, Ecosystems & Environment, 2001, 85(1/2/3): 191-203. |
| [16] |
谢花林, 刘黎明, 李波, 张新时. 土地利用变化的多尺度空间自相关分析——以内蒙古翁牛特旗为例. 地理学报, 2006, 61(4): 389-400. DOI:10.3321/j.issn:0375-5444.2006.04.006 |
| [17] |
Batjes N, Ribeiro E, Oostrum A V. Standardised soil profile data to support global mapping and modelling (WoSIS snapshot 2019). Earth System Science Data, 2020, 12: 299-320. DOI:10.5194/essd-12-299-2020 |
| [18] |
施建平, 宋歌. 中国土种数据库——基于第二次土壤普查的全国性土壤数据集. 中国科学数据, 2016, 1(2): 1-12. |
| [19] |
Ponce-Hernandez R, Marriott F H C, Beckett P H T. An improved method for reconstructing a soil profile from analyses of a small number of samples. Journal of Soil Science, 1986, 37(3): 455-467. DOI:10.1111/j.1365-2389.1986.tb00377.x |
| [20] |
Bishop T F A, McBratney A B, Laslett G M. Modelling soil attribute depth functions with equal-area quadratic smoothing splines. Geoderma, 1999, 91(1/2): 27-45. |
| [21] |
Malone B P, McBratney A B, Minasny B, Laslett G M. Mapping continuous depth functions of soil carbon storage and available water capacity. Geoderma, 2009, 154(1/2): 138-152. |
| [22] |
Xiao L J, Wang G C, Wang M M, Zhang S, Sierra C A, Guo X W, Chang J F, Shi Z, Luo Z K. Younger carbon dominates global soil carbon efflux. Global Change Biology, 2022, 28(18): 5587-5599. DOI:10.1111/gcb.16311 |
| [23] |
Pearson R G, Stanton J C, Shoemaker K T, Aiello-Lammens M E, Ersts P J, Horning N, Fordham D A, Raxworthy C J, Ryu H Y, McNees J, Akçakaya H R. Life history and spatial traits predict extinction risk due to climate change. Nature Climate Change, 2014, 4(3): 217-221. DOI:10.1038/nclimate2113 |
| [24] |
Du S S, Liu Y Y. TanSat Solar-Induced Chlorophyll Fluorescence Product. (2020-6-7)[2021-9-1]. https://zenodo.org/record/3883434#.Y05ACi-KGpY.
|
| [25] |
Klein Goldewijk K, Beusen A, Janssen P. Long-term dynamic modeling of global population and built-up area in a spatially explicit way: HYDE 3.1. The Holocene, 2010, 20(4): 565-573. DOI:10.1177/0959683609356587 |
| [26] |
Kummu M, Taka M, Guillaume J H A. Gridded global datasets for gross domestic product and human development index over 1990-2015. Scientific Data, 2018, 5(1): 1-15. DOI:10.1038/s41597-018-0002-5 |
| [27] |
Zhang Y G, Schaap M G, Zha Y Y. A high-resolution global map of soil hydraulic properties produced by a hierarchical parameterization of a physically based water retention model. Water Resources Research, 2018, 54(12): 9774-9790. DOI:10.1029/2018WR023539 |
| [28] |
Kosugi K. Three-parameter lognormal distribution model for soil water retention. Water Resources Research, 1994, 30(4): 891-901. DOI:10.1029/93WR02931 |
| [29] |
Channan S, Collins K, Emanuel W. Global mosaics of the standard MODIS land cover type data. (2017-6-7)[2021-9-1]. University of Maryland and the Pacific Northwest National Laboratory.
|
| [30] |
Olson D M, Dinerstein E, Wikramanayake E D, Burgess N D, Powell G V N, Underwood E C, D'Amico J A, Itoua I, Strand H E, Morrison J C, Loucks C J, Allnutt T F, Ricketts T H, Kura Y, Lamoreux J F, Wettengel W W, Hedao P, Kassem K R. Terrestrial ecoregions of the world: a new map of life on earth. BioScience, 2001, 51(11): 933. DOI:10.1641/0006-3568(2001)051[0933:TEOTWA]2.0.CO;2 |
| [31] |
McBratney A B, Mendonça Santos M L, Minasny B. On digital soil mapping. Geoderma, 2003, 117(1/2): 3-52. |
| [32] |
周国法. 具有空间自相关残差的回归模型及其应用. 生物数学学报, 1997, 12(2): 158-163. |
| [33] |
于尚洋. GLMM中基于空间自相关和迭代残差的空间聚类检测[D]. 秦皇岛: 燕山大学,.
|
| [34] |
管孝艳, 杨培岭, 吕烨. 基于多重分形理论的农田土壤特性空间变异性分析. 应用基础与工程科学学报, 2011, 19(5): 712-720. DOI:10.3969/j.issn.1005-0930.2011.05.003 |
| [35] |
王小艳, 冯跃华, 李云, 武彪, 陈山, 李香玲, 王旭, 莫银化, 宋碧. 黔中喀斯特山区村域稻田土壤理化特性的空间变异特征及空间自相关性. 生态学报, 2015, 35(9): 2926-2936. DOI:10.5846/stxb201404300863 |
| [36] |
Tian H W, Zhang J H, Zhu L Q, Qin J T, Liu M, Shi J Q, Li G D. Revealing the scale- and location-specific relationship between soil organic carbon and environmental factors in China's north-south transition zone. Geoderma, 2022, 409: 115600. DOI:10.1016/j.geoderma.2021.115600 |
| [37] |
Schuur E A G, Chadwick O A, Matson P A. Carbon cycling and soil carbon storage in mesic to wet Hawaiian montane forests. Ecology, 2001, 82(11): 3182-3196. DOI:10.1890/0012-9658(2001)082[3182:CCASCS]2.0.CO;2 |
| [38] |
Yang Y B, Fang X M, Galy A, Yang R S. Carbonate composition and its impact on fluvial geochemistry in the NE Tibetan Plateau region. Chemical Geology, 2015, 410: 138-148. |
| [39] |
Viscarra Rossel R A, Webster R, Bui E N, Baldock J A. Baseline map of organic carbon in Australian soil to support national carbon accounting and monitoring under climate change. Global Change Biology, 2014, 20(9): 2953-2970. |
| [40] |
Luo Z K, Wang E L, Sun O J. Can no-tillage stimulate carbon sequestration in agricultural soils? A meta-analysis of paired experiments. Agriculture, Ecosystems & Environment, 2010, 139(1/2): 224-231. |
| [41] |
Tiemann L K, Grandy A S, Atkinson E E, Marin-Spiotta E, McDaniel M D. Crop rotational diversity enhances belowground communities and functions in an agroecosystem. Ecology Letters, 2015, 18(8): 761-771. |
| [42] |
Poggio L, Sousa L D, Batjes N H, Heuvelink G B M, Kempen B, Ribeiro E, Rossiter D. SoilGrids 2.0: producing soil information for the globe with quantified spatial uncertainty. SOIL, 2021, 7(1): 217-240. |
| [43] |
Gray J M, Humphreys G S, Deckers J A. Relationships in soil distribution as revealed by a global soil database. Geoderma, 2009, 150(3-4): 309-323. |
| [44] |
Hobley E, Wilson B, Wilkie A, Gray J, Koen T. Drivers of soil organic carbon storage and vertical distribution in Eastern Australia. Plant Soil, 2015, 390: 111-127. |
| [45] |
Frank D A, Pontes A W, McFarlane K J. Controls on soil organic carbon stocks and turnover among north American ecosystems. Ecosystems, 2012, 15(4): 604-615. |
| [46] |
左昕弘, 赖佳鑫, 刘峰, 石孝均. 基于地统计学和空间自相关的土壤有机质空间异质性分析及分区——土地整治视角. 中国农业资源与区划, 2022, 43(03): 240-252. |