 2024, Vol. 44
2024, Vol. 44文章信息
- 赵鑫田, 陈开超, 李浡睿, 孙启兴, 周洁, 孙楠, 廖珂, 李赛强, 梅慧芳, 罗旭, 杨晓东
- ZHAO Xintian, CHEN Kaichao, LI Borui, SUN Qixing, ZHOU Jie, SUN Nan, LIAO Ke, LI Saiqiang, MEI Huifang, LUO Xu, YANG Xiaodong
- 荒漠灌木与城市绿化灌木功能性状及其对夏季热浪响应机理的差异
- Differences in functional traits and the response mechanism to summer heat wave between desert native shrubs and urban greening shrubs
- 生态学报. 2024, 44(7): 2837-2848
- Acta Ecologica Sinica. 2024, 44(7): 2837-2848
- http://dx.doi.org/10.20103/j.stxb.202306301398
-
文章历史
- 收稿日期: 2023-06-30
- 网络出版日期: 2024-01-12





2. 宁波市林场, 宁波 315440;
3. 新疆大学绿洲生态教育部重点实验室, 乌鲁木齐 830046;
4. 新疆大学生态与环境科学学院, 乌鲁木齐 830046
2. Ningbo Forest Farm of Zhejiang Province, Ningbo 315440, China;
3. Key Laboratory of Oasis Ecology, Ministry of Education, Urumqi 830046, China;
4. Institute of Resources and Environment Science, Xinjiang University, Urumqi 830046, China
受全球气候变化的影响, 夏季热浪现象在世界各地频繁发生, 并呈增加趋势[1]。例如, 2022年, 欧洲和亚洲的6月中旬气温突破40℃, 且最高气温远高于其他年份同期值(27℃)[1]。7月中旬, 葡萄牙、北非和伊朗的最高气温分别达到45℃、48℃和52℃, 均创下当地历史最高纪录。中国于7月12日发布高温红色预警后, 截至8月28日, 在我国其中23个省份共记录到7555次40℃以上的高温。比同期前五年(2017—2021)的平均水平高11.6倍[2]。作为一种自然灾害, 夏季热浪影响公众健康和生命安全[3], 还会通过高温和热气流冲击使植物干旱胁迫加剧, 因水力失败或水力失衡引起大量出现植株结构性受损, 甚至死亡, 进而改变群落结构, 引起生物多样性丧失、减弱生态系统的功能和稳定性[4]。
热浪对植物的影响可能因物种类型而异[5]。根据植物生长的区位和人类管理的程度, 植物可分为乡土原生植物和城市绿化物种[6]。原生植物在没有人为影响情况下自然生长和繁殖[7]。相反, 城市绿化物种则需要被培育、维护和管理, 以形成城市景观, 提高城市生态质量[8]。由于长期适应性环境选择和人工栽培, 城市绿化植物处在有利环境中, 其应对自然灾害的能力弱于本地物种[9]。同时, 城管部门在绿化植物在筛选时主要考虑的是外观、气味、花色等景观属性特征, 忽略抗逆能力。因此, 在城市热浪期间, 绿化植物受到高温和热气流冲击的影响高于本地乡土物种, 其诱发干旱胁迫导致的受损程度、落叶比例和死亡株数均高于后者。但是, 由于夏季热浪在最近几年影响时长和范围各不相同, 其研究窗口期变动大和事件突发的特点使得国内外有关植物响应热浪的研究相对不足[10—11], 少有成果分析过乡土种和城市绿化种的受损程度, 揭示它们应对高温和热气流冲击的机理。
水力相关性状的差异及其整合可能是揭示城市绿化植物和荒漠原生物种应对热浪机理不同的一个重要视角[12]。水力性状是指植物在长期进化过程中形成的形态、解剖、结构和生理特性, 如蒸腾速率、气孔导度、Huber值、P50和枝条木质部水力导度等[13]。这些性状与叶片和大气之间的水分吸收、输送和水蒸气交换有关[14]。已有研究证实, 当植物遭受热浪的高温和热气流冲击时, 这些性状的差异将导致植物应对干旱胁迫的能力发生变化, 进而影响受损的不同[15]。例如, 具有较高P50或较低导水率的物种优先经历水力失衡(/水力失败), 或由于气孔关闭(碳饥饿)而缺乏营养, 导致植株受损, 甚至死亡[16]。此外, 植物对热浪引起干旱胁迫的适应并不是通过单一性状, 而是通过多种性状的组合实现的[2, 17]。换句话说, 植物利用不同性状之间的互补关系和权衡来提高对热浪的适应[12]。这种特征组合的适应被生态学家称为性状整合[18]。研究发现, 性状整合在缓解高温引起的干旱胁迫方面比单个性状更有效, 并且可以更准确地预测植物受损[19]。植物性状的整合度越高, 意味着植物对干旱胁迫的适应性更强, 在热浪期间不易受损[20—21]。然而, 在夏季热浪期, 从水力学和性状整合的角度分析城市绿化植物和乡土原生植物响应热浪机理方面的研究亟待深入。
中国新疆维吾尔自治区位于亚洲中部, 是距离海洋最远的地方。降雨量不足和蒸发量高导致当地植物长期遭受干旱胁迫[22]。据新疆气象中心报道, 受全球气候变化影响, 近年来当地夏季平均气温已从23.2℃上升至26.4℃, 极端最高气温从32℃上升到39℃。2019年、2020年和2022年, 连续发生夏季热浪事件。2022年热浪被认为是自1961年以来最长、最强的一次。本文在2022年夏季热浪期间, 选择古尔班通古特沙漠西部边缘的新疆维吾尔自治区精河县和艾比湖湿地自然保护区为研究地点, 测量了沙漠中9种乡土灌木物种和县城内8种常见绿化灌木在水力相关性状和性状整合方面的差异。本文科学问题是水力相关性状和其整合在荒漠乡土植物和城市绿化植物间的差异, 导致了它们应对夏季热浪引起干旱胁迫的机理不同。对这个问题的解读, 可为干旱地区的绿化植物的筛选提供理论支持。
1 研究区概况研究区位于中国西北部新疆维吾尔自治区古尔班通古特沙漠西部边缘的精河县, 以及其邻接的艾比湖湿地自然保护区。受典型大陆性气候影响, 精河县和艾比湖湿地自然保护区年平均降水量不足100 mm, 年平均蒸发量超过1600 mm。日照时数约2800 h, 极端最高气温44℃, 极端最低气温-33℃, 年平均气温6—8℃, 平均海拔189 m。当地土壤类型以灰褐色荒漠土为主, 盐渍化程度高, pH值为7.52—9.29。
2 物种选择和实验测定 2.1 物种选择考虑精河县和艾比湖湿地自然保护区的基底景观为荒漠稀疏灌丛, 当地的荒漠乡土乔木仅有胡杨一个物种, 为减小统计上误差, 本项目在研究绿化植物和乡土植物对热浪引起干旱胁迫响应时, 仅选择了灌木物种, 没有考虑乔木种。在精河县面积较大的公园内, 基于以下两点选择城市绿化植物: (1) 人工种植绿化灌木; (2) 在城市中广泛分布, 是人工种植的优势灌木种。研究选取了8种城市绿化灌木, 分别为红瑞木(Cornus alba)、紫叶李(Prunus cerasifera)、小叶女贞(Ligustrum quihoui)、紫穗槐(Amorpha fruticose)、丁香花(Syringa oblata)、虎榛子(Ostryopsis davidiana)、蔷薇(Rosa multiflora)和风箱果(Physocarpus amurensis)。为排除微地形和小气候等非生物因素的影响, 这些绿化灌木都集中分布在精河县的人民公园和91团团部公园周边。
在艾比湖湿地自然保护区的东大桥站至托托镇的绿洲和荒漠的交错区内, 选择当地的优势灌木为研究对象。将当地9种优势灌木为作为研究对象。它们分别是白刺(Nitraria tangutorum)、骆驼刺(Alhagi sparsifolia)、琵琶柴(Reaumuria songonica)、沙拐枣(Calligonum mongolicum)、盐节木(Halocnemum strobilaceum)、铃铛刺(Halimodendron halodendron)、盐穗木(Halostachys caspica)、盐爪爪(Kalidium foliatum)和黑果枸杞(Lycium ruthenicum)。为减小微环境对实验的影响, 交错区取样区选择在地势平缓处, 其范围不超过3 km。
2.2 夏季热浪植物受损程度测定在原生荒漠灌木和城市绿化灌木调查的区域, 针对每个物种分别调查了100株个体因热浪引起干旱胁迫的受损程度。热浪对植物的影响被定义为以下5类:正常、轻微、中度、重度和植株死亡(表 1)。调查时间段在2022年8月中下旬, 处于新疆新疆气象中心的记录北疆夏季热浪的末期(精河县2022年的夏季热浪的持续时间为6月1日—8月24日), 植物受损情况可反馈夏季热浪对植物在整个持续期的影响。
| 编号 Number |
受损级别 Damage degree |
特征 Characteristics |
编号 Number |
受损级别 Damage degree |
特征 Characteristics |
|
| 1 | 正常 | 植株未有任何变化 | 4 | 重度 | 叶片大部分凋落, 部分枝条干枯死亡 | |
| 2 | 轻微 | 少数叶片变黄, 有落叶现象 | 5 | 植株死亡 | 叶片完全凋落, 植物枯死 | |
| 3 | 中度 | 较多的叶片变黄并落叶 |
对8种城市绿化灌木和9种荒漠乡土灌木, 各随机选择3—5棵植株, 测定功能性状。本文实验测定时间处在夏季热浪后期, 由于干旱胁迫对植物的不利影响会随着时间的推移而累积。本文获得数值恰好捕捉了夏季热浪期温度和热气流冲击引起干旱胁迫对植物的最大伤害。此外, 为真实反映夏季热浪对两种灌木的影响, 城市绿化植物在实验期不做任何处理, 园林管理部门按照正常绿化管理要求实施浇水管护。
2.3.2 功能性状的测定植物抗旱性状可分为水力性状、光合性状、碳经济性状和叶性状。其均可以通过影响水分运输、水力安全、水分吸收、叶片和大气间的水分交换等过程影响植物对干旱胁迫的响应。本文中这几种性状测定方法如下:
(1) 水力性状
本文选择的水力性状有:黎明前枝叶水势、正午枝叶水势、准稳态导水率、P50、胡伯尔值、干材密度和解剖特征(导管密度和直径)。测定方法如下:
① 黎明前枝叶水势和正午枝叶水势
对于每个灌木种选择的个体, 随机选取3—5个叶片完整且无虫害的树枝, 然后将其剪断。按照前人的方法对样品进行预处理和保存[23]。之后使用便携式露点水势计(WP4C, Decagon Devices, Pullman, Washington, USA)和压力室仪器(1505D-EXP; PMS Instrument Company, Albany, OR, USA)分别测量叶片水势和枝水势。所有叶片测量重复三次。凌晨和中午水势采样时间分别为当地时间05:30—07:30和12:30—14:30。
② 准稳态导水率、P50、胡伯尔值和干材密度
使用植物水电导率测量仪(HPFM-Gen3, Dynamax Inc., Houston, USA)测量木质部栓塞脆弱性曲线, 获得木质部导水率(枝条比导率和准稳态导水率)。枝条的采样和后续的实验预处理参照Venturas等人的方法[24]。根据Sperry等人提出采样压力室仪器(1505D-EXP; PMS Instrument Company, Albany, OR, USA)和高压流量计来确定导水率损失百分比(PLC; %)[25]。然后通过拟合PLC曲线得到P50(导水率损失50%时的水势)[26]。实验过程中压力范围根据Sperry等人的研究方法确定[27]。具体地, 从0 MPa开始, 之后将0.5 MPa设为增量, 依此在0.5, 1.0, 1.5, 2.0, 2.5, 3.0, 3.5 MPa, 以及随后的几个压力值分别测定导水率, 直至栓塞达到95%以上停止。实际工作中, 参考Yang等人对干旱区植物的研究成果, 压力值一般不超过3.5Mpa[23]。之后, 木材横截面积(SA; m2)和树枝上所有叶子的总面积(TLA; m2)分别使用游标卡尺和叶面积仪(Li-3100C, Lincoln, NE, USA)进行测量。胡伯尔值由公式(1)计算[28]。为了减小蒸腾的影响, 枝条采样在当地时间06:00-07:00进行。枝条的干材密度采用排水法测定[23]。
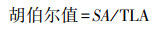
|
(1) |
③ 水力解剖特征
从测定导水率的茎段上截取一段, 用于测定木质部解剖结构特征。样品固定软化之后, 采用石蜡切片法将其制成永久切片[29]。具体步骤如下:
ⅰ) 固定样品:从测定脆弱曲线的枝条上截取1根3—5 cm的枝段, 将其切成5 cm长的条状, 放入FAA固定液(使用甲醛原液、冰醋酸原液、70%乙醇配置)中固定, 时间大约24 h; 经乙醇脱水、透明、石蜡浸蜡、模具包埋、切片机切片、番红染色与树胶封片等过程。
ⅱ) 导管直径(D, μm)测定:所有切片样品通过显微镜(Leica Microsystems CMS GmbH; Wetzlar Germany; Model:DM3000 LED)在50倍镜和400倍镜下摄像。获取横切面照片, 随后使用Image J图像处理软件分析照片。选取每个物种的每个重复在50倍镜下的照片各一张, 每张照片选择3个不同位置的30°扇形切面, 然后测量所选区域中所有导管直径。得到每张照片中所有导管直径的值, 并计算每张照片中所有导管直径的平均值来表示物种的一个重复, 每个物种20个重复[30]。
ⅲ) 导管密度(N, 个/mm2)测定:在测量导管直径同一张照片中, 选择3个不同角度的扇形切面, 对3个扇形切面中的导管数量进行计数, 随后计算扇形切面的面积(不包括髓、韧皮部与树皮), 导管密度通过导管个数与面积的比值获得。
(2) 叶性状
本文选择的叶性状有干物质含量和比叶面积。对每个选定的个体, 选择3—5个叶片完好、无虫害的枝条, 然后剪掉3—5个叶片。所有样品带回实验室后, 分别用叶面积仪和电子分析天平测定叶片面积(LA, cm2)和鲜重(LFW, g), 准确度分别为<0.0001 g。采用烘箱干燥法测定叶片干质量(LDW, g), 由式2—3分别计算叶片干物质含量(LDMC, g/g)和比叶面积(SLA, m2/kg)。
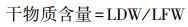
|
(2) |
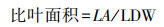
|
(3) |
(3) 光合性状
本文选择的叶性状有叶片气孔导度(Gs, H2O μmol m-2 s-1)、蒸腾速率(Tr, H2O μmol m-2 s-1)、净光合速率(Pn, μmolCO2 m-2 s-1)和内在水分利用效率(WUEi, mol mol-1)。采用Li-6400XT便携式光合作用测量系统(Lincoln, NE, USA)测定。为控制和减小微气候环境变化对光合性状的影响, 叶室的温度、相对湿度、CO2浓度和光照均人工控制, 数值分别设置为35℃、70%、400 μmol/mol和1600 μmol m-2 s-1。这几个值均是当地夏季影响植物光合作用的最优气象条件。此条件下测定光合性状为最大值。这种控制可以避免因为光照、温度和CO2浓度等因子的日内变化引起测量偏差。
(4) 碳经济性状
本文选择的碳经济性状有植物可溶性糖、淀粉和非结构性碳含量。可溶性糖的测定采用80%的乙醇离心预处理样本在采用蒽酮法测定[31]; 淀粉测定采用蒽酮显色法。非结构性碳含量根据可溶性糖和淀粉含量进行计算。具体见公式4。
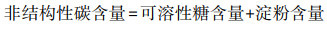
|
(4) |
采用独立样本T检验比较了荒漠原生灌木与城市绿化灌木在功能性状和热浪引起损伤程度上的差异。利用R程序包“igraph”测量每个物种的性状整合[32]。所有分析均在R统计软件(4.0版本)中实现。
3 结果分析 3.1 荒漠乡土灌木和城市绿化灌木的受损程度荒漠乡土灌木接近74%的个体在夏季热浪中的未受损伤, 仅有26%的植株轻微或中度受损, 没有个体达到重度(4级)和植株死亡(5级)的受损程度(图 1)。但对于城市绿化灌木, 大约56%的个体受到夏季热浪的损伤。其中, 大约有4%和3%的个体达到了重度和植株死亡的受损程度。城市绿化灌木的轻微、重度和植株死亡的个体数均高于荒漠乡土灌木。

|
| 图 1 夏季热浪期间荒漠乡土灌木(Dns)和城市绿化灌木(Ugs)的受损程度的差异 Fig. 1 Difference in damage degree to desert native shrubs (Dns) and urban greening shrubs (Ugs) during summer heatwaves |
荒漠乡土灌木植物的枝水势(黎明前水势和正午水势)的平均值低于城市绿化植物, 且前者的黎明前和正午叶水势均显著低于后者(P<0.05)。枝比导率、准稳态导水率、胡伯尔值和P50在两种类型植物之间没有统计上的显著差别(P>0.05), 但荒漠乡土灌木的枝比导率、准稳态导水率、胡伯尔值的平均值高于城市绿化灌木, 相反P50的平均值在前者略低于后者。所有荒漠乡土灌木的性状的变异性或数值范围远大于城市绿化植物(图 2)。

|
| 图 2 荒漠乡土灌木与城市绿化灌木水力性状的差异 Fig. 2 Differences in hydraulic traits between desert native shrubs and urban greening shrubs 性状的差异采用独立样本t检验检验; 相同字母表示处理间差异不显著, 不同字母表示处理间差异显著; P<0.05 |
在叶片性状中, 荒漠乡土灌木的比叶面积和导管密度显著高于城市绿化灌木(P<0.05), 但干物质含量和导管直径成相反模式(P<0.05), 干材密度没有显著性差异(P>0.05)(图 3)。碳经济性状中, 荒漠乡土灌木的淀粉和非结构性碳含量和城市绿化灌木无显著差异(P > 0.05), 但前者的可溶性碳含量显著高于后者(P<0.05) (图 3)。在光合性状方面, 城市绿化灌木的气孔导度显著高于荒漠乡土灌木(P<0.05), 而水分利用效率成相反模式(P<0.05), 但净光合速率和蒸腾速率无显著差异(P > 0.05) (图 3)。

|
| 图 3 荒漠乡土灌木和城市绿化灌木间在叶性状、碳经济和光合性状上的差异 Fig. 3 Differences in leaf, photosynthetic, and carbon economic traits between desert native shrubs and urban greening shrubs 性状的差异采用独立样本t检验检验; 相同字母表示处理间差异不显著, 不同字母表示处理间差异显著; P<0.05 |
G表示性状整合的网络密度。它是根据节点数N和边界数L计算获得的[G=2L/N (N-1)]。G的数值越高, 表示性状间整合度就越高, 性状间总体联系就越密切。本文研究结果表明, 荒漠乡土灌木的性状整合度(G=0.39)高于城市绿化植物(G=0.2)(图 4)。荒漠乡土灌木性状整合的关键节点是与水力性状和气孔调节相关的性状, 比如准稳态导水率、P50、气孔导度、蒸腾速率、净光合速率、水分利用效率和导管密度。这些关键节点的性状至少与其他11个性状有联系。此外, 枝比导率、胡伯尔值、可溶性糖含量、非结构碳含量和导管直径也与至少7个其他性状紧密结合(图 4)。在城市绿化灌木的性状整合上, 比叶面积、干物质含量、枝叶水势和水分利用效率是关键节点, 它们分别与7个其他性状相关联(图 4)。

|
| 图 4 城市绿化灌木的性状整合及其与荒漠乡土灌木的差别 Fig. 4 Difference in trait integration between urban greening shrubs and desert native shrubs Ψlpd、Ψlmd、Ψspd分别为黎明前叶水势、正午叶水势、黎明前枝水势和正午枝水势, 单位均是MPa; Ks、K、Hv和P50分别表示枝条的枝比导率、准稳态导水率、胡伯尔值和导水率损失50%时的水势, 单位分别为kg s-1 m-1 MPa-1、10-3kg s-1 MPa-1、m2/m2和Mpa。Dns和Ugs分别为荒漠原生植物与城市绿化植物; SLA、LDMC、SWD、D和N分别是比叶面积、叶片干物质含量、枝条干材密度、导管直径和导管密度的缩写; 它们的单位分别为m2/kg、g/g、g/cm3、μm和mm-2; SC、SS和NSC是淀粉、可溶性糖和非结构性碳的缩写, 它们的单位均为%; Gs、Tr、Pn和WUEi分别是气孔导度、蒸腾速率、净光合速率和内在水分利用效率的缩写。这几个指标的单位分别为H2O2μmol m-2 s-1、μmol m-2 s-1、μmolCO2 m-2 s-1和mol/mol; Dns和Ugp分别代表荒漠乡土灌木和城市绿化灌木; 各种间性状间的正相关和负相关分别用实线和虚线表示; 线越粗表示相关性越强; P<0.05 |
荒漠原生灌木在遭受夏季热浪后, 未出现4级以上受损的个体, 3级受损个体也不到总数的3%, 但城市绿化灌木受损个体占总数的56%以上, 甚至还有整株死亡。这表明在夏季热浪影响下城市绿化灌木的损伤远大于荒漠乡土灌木。这主要取决于两方面:(1) 长期环境适应性和遗传特性使两者受损程度存在差异。相对荒漠乡土灌木, 城市绿化灌木耗水量大而抗热浪引起干旱胁迫的能力小。城市管理部门筛选绿化物种时, 优先考虑的是植物的外貌和其他景观特征(比如植物花色、叶片形状、气味等), 以及苗木的易获得性, 很少顾及植物抗环境胁迫的遗传特性[10]。这使得城市绿化植物主要来源于环境条件优越的生境。比如, 本文调查的绿化植物紫叶李、小叶女贞和紫穗槐, 原生地主要在温带季风气候区或者海拔较高降水量较多的地区。未有人为干扰情景下, 它们在荒漠上生存受限。相反, 原生荒漠灌木在长期进化中形成了适应干旱的众多特征。比如琵琶柴、白刺、盐节木、盐穗木和盐爪爪的叶片退化; 铃铛刺和沙拐枣叶片上的白粉。在遭遇夏季热浪时, 绿化灌木在抗热浪能力的不足导致其死亡率增加; (2) 正常绿化灌溉导致绿化植物受损增加[33]。为了保证城市绿化植物正常的生长, 干旱地区的城市管理部门定期对绿化植物进行灌溉。尽管这种管理使得绿化植物在正常情况下能生长, 但在夏季热浪发生时, 较高的气温使得土壤蒸发和植物蒸腾水分量远高于正常情景。这种情景下, 按照以往正常的管理方式, 将不能满足植物的耗水要求, 容易因干旱胁迫导致受损[34]。此外, 在干旱区荒漠, 夏季热浪使得城市缺水现象严重, 尽管按期浇灌, 但蒸腾增加, 可能进一步加重了绿化灌木承受的干旱胁迫。
4.2 荒漠乡土植物相较城市绿化植物具有应对夏季热浪的水力性状优势本研究发现, 荒漠灌木物种的水力性状要优于城市绿化灌木。这表明荒漠植物在水力学方面优势, 如增加水分输送和吸水能力, 减少了水分蒸腾损失等, 以降低夏季热浪引起干旱胁迫的影响, 使它们相对城市绿化灌木获得更高生存概率。水势是衡量植物抗旱性的重要指标[35]。在本研究中, 荒漠灌木物种枝叶水势相对城市绿化灌木较低, 表明在高温和热浪冲击的影响下, 荒漠灌木物种更低的叶片与土壤间水势差使其更容易从土壤中吸收水分[36]。此外, 荒漠灌木物种正午和黎明前的水势差小于城市绿化灌木, 表明它的再水合能力强于城市绿化灌木[12]。这种优势让其在干旱胁迫引起木质部的栓塞可以通过再水合过程进行修复, 减少高温和热浪冲击的影响。水力结构由木质部导管的形态结构决定, 它影响植物水分输送策略[37]。在本研究中, 荒漠乡土灌木物种的水力结构(枝比导率、准稳态导水率和胡伯尔值以及导管密度)均高于城市灌木。这表明, 荒漠乡土灌木比城市绿化灌木从根和茎向叶的供水能力更好, 水分输送效率也更高[38]。这使得荒漠乡土灌木物种在夏季热浪期更能承受高温和热气流冲击的影响, 减弱它们引起干旱胁迫增加导致的水分过度损失。这一点也可以用P50数据进行佐证, 尽管荒漠乡土灌木的数值和城市绿化灌木间未有显著差异, 但前者的平均值和最低值均小于后者。
荒漠乡土灌木更优的抗旱能力也可以通过光合性状反映。乡土物种的气孔导度低于城市绿化灌木, 但水分利用效率表现出相反的模式。这表明荒漠灌木物种只需要更少的水就可以合成等量光合有机物。较低耗水量更有利于减弱夏季热浪引起干旱胁迫的影响。此外, 植物的抗旱性在一定程度上取决于它们抵抗栓塞发生的能力或栓塞后修复的能力[39]。木材密度和被认为是植物抗栓塞的重要指标。这是因为木材密度与木质部导管的硬度呈正相关, 较高的木材密度使植物不太容易发生栓塞[47]。本研究发现荒漠灌木物种的木材密度高于城市绿化灌木。导管特性(直径和密度)也影响植物栓塞形成。荒漠乡土灌木较小的导管直径和较大导管密度能够提升木质部的脆弱性, 减小干旱诱发栓塞的风险[41]。
4.3 荒漠原生植物相较城市绿化植物应对夏季热浪的机理更多荒漠乡土灌木物种的性状整合度高于城市绿化物种。这表明荒漠乡土灌木在经受夏季热浪影响时能更好地协调功能性状间的关系, 采取多种机理将高温和热浪冲击诱发干旱胁迫的负面影响降至最低, 由此导致它比城市绿化的受损更小[42—43]。Yang等人[15]和刘丛聪等[44]学者也得到了类似的结果。他们发现, 性状整合水平较高的植物比整合水平较低的植物更能抵御逆境。在荒漠灌木的性状整合中, 水力输送有关性状和气孔调节性状(准稳态导水率、P50、干物质含量、气孔导度、蒸腾速率、净光合速率、水分利用效率、导管密度)是性状整合网络的关键节点, 荒漠灌木物种主要通过增加水分输送和采取气孔调节来减少夏季热浪的影响[26, 45]。例如, 准稳态导水率较高的植物通常具有较高的叶面积和光合特征[17], 因为导水率的增加可以将更多的水从根系和树干输送到叶片, 保证光合和蒸腾所需的水分供给。同时, 这一过程的正反馈提升的蒸腾拉力, 也可以克服重力和木质部水分运输摩擦力, 将更多的水分从根系提升至叶片。
荒漠乡土灌木和城市绿化灌木在性状整合中关键节点的不同反映了其应对热浪机理的差异。光合性状和气孔属性作为节点, 表明荒漠乡土灌木可以通过气孔调节来减少高温和热浪冲击引起干旱胁迫的影响[46]。在干旱胁迫下, 由于水势差过大, 乡土灌木关闭气孔, 以降低木质部栓塞导致水力衰竭的风险。同时, 围绕水分运输性状为节点, 可能表明荒漠灌木物种中也出现了另一种适应方式[47]。例如, 遭受高温和热浪冲击时, 气孔没有立刻关闭, 而是通过增加蒸腾拉力来吸收和输送更多的水分, 以减少干旱胁迫[48—49]。相反, 在绿化植物的性状整合中, 与叶片有相关的性状(比叶面积、干物质含量和水分利用效率)为关键节点, 且关键点的数量较少。这表明, 当城市绿化灌木遭受夏季热浪时, 它们的应对机制较为简单。在短期内, 它们只能通过调节自身光合作用和增加溶质势来减少自身水分的消耗和蒸腾丧失。同时, 荒漠灌木与城市灌木中的P50很少与其他性状相关, 可能是木质部栓塞脆弱性与植物短期抗旱性之间的关系不明显。在荒漠地区, 植物长期受到干旱胁迫, 导致木质部导管永久堵塞[50], 因而P50在夏季热浪期的变化不大。这种现象在植物水分生理学上被称为“空化疲劳”[39]。
5 结论和展望遭受2022年夏季热浪的危害后, 荒漠乡土灌木的受损程度低于城市绿化灌木。荒漠植物在水力性状的优势, 导致它的水分运输、吸收和减少蒸腾失水的能力要强于城市绿化灌木。荒漠乡土灌木的性状整合度也高于城市绿化灌木。荒漠植物通过协调功能性状间的关系, 组合优势, 采取多种机理(比如气孔调节、水力运输调节和增加蒸腾拉力)将高温和热浪冲击诱发干旱胁迫的负面影响降低。对城市绿化灌木而言, 其性状整合的关键节点数较少且以叶性状为主, 其遭遇夏季热浪时, 防御策略较简单, 主要利用减小蒸腾的气孔调节机理。
夏季热浪对植物的影响除干旱胁迫外, 还存在高温灼伤。此外, 夏季热浪对植株的损害是一个持续积累的过程。在未来实验中, 利用盆栽控制实验, 结合户外真实环境条件对绿化植物和乡土植物的水力相关性状进行测定, 同步在不同时段连续观测叶片温度变化以及高温灼伤过程, 可更好揭示两种植物在应对热浪机理上的差异, 以及由此导致受损程度的不同。另外, 在干旱荒漠区, 水分和盐分同时对植物产生负向影响, 且两者很难分离。在夏季热浪期, 强烈蒸发会引起地表积累的盐分快速积累, 由此通过盐胁迫进一步加强根系水分吸收困难, 加重干旱胁迫的影响。未来也应该针对夏季热浪影响下水盐的协同效应进行研究。
2022年人类有记录以来北半球最严重的夏季热浪已离去, 但全球气候变化背景下热浪的未来情景不知。2022年夏季热浪或许是未来温度最低的一个起始, 因此, 它对城市绿化生态系统的损害可能是巨大的。我们建议, 城市管理部门在筛选绿化植物时, 除景观特点外, 优先应该考虑植物的水力特性, 筛选在水分吸收、运输、低蒸腾损失上更有优势的物种。此外, 为提高城市绿化植物应对夏季热浪的能力, 在干旱半干旱地区, 可选择当地的原生乡土植物作为绿化物种。
| [1] |
Lu R Y, Xu K, Chen R D, Chen W, Li F, Lv C Y. Heat waves in summer 2022 and increasing concern regarding heat waves in general. Atmospheric and Oceanic Science Letters, 2023, 16(1): 100290. DOI:10.1016/j.aosl.2022.100290 |
| [2] |
Liu X J, Tian G J, Feng J M, Hou H L, Ma B R. Adaptation strategies for urban warming: assessing the impacts of heat waves on cooling capabilities in Chongqing, China. Urban Climate, 2022, 45: 101269. DOI:10.1016/j.uclim.2022.101269 |
| [3] |
Amengual A, Homar V, Romero R, Brooks H E, Ramis C, Gordaliza M, Alonso S. Projections of heat waves with high impact on human health in Europe. Global and Planetary Change, 2014, 119: 71-84. DOI:10.1016/j.gloplacha.2014.05.006 |
| [4] |
冯雷, 李旭东. 高温热浪对人类健康影响的研究进展. 环境与健康杂志, 2016, 33(2): 182-188. |
| [5] |
Qu L P, De Boeck H J, Fan H H, Dong G, Chen J Q, Xu W B, Ge Z Q, Huang Z J, Shao C L, Hu Y L. Diverging responses of two subtropical tree species (Schima superba and Cunninghamia lanceolata) to heat waves. Forests, 2020, 11(5): 513. DOI:10.3390/f11050513 |
| [6] |
Asgarzadeh M, Vahdati K, Lotfi M, Arab M, Babaei A, Naderi F, Pir Soufi M, Rouhani G. Plant selection method for urban landscapes of semi-arid cities (a case study of Tehran). Urban Forestry & Urban Greening, 2014, 13(3): 450-458. |
| [7] |
Berthon K, Thomas F, Bekessy S. The role of 'nativeness' in urban greening to support animal biodiversity. Landscape and Urban Planning, 2021, 205: 103959. DOI:10.1016/j.landurbplan.2020.103959 |
| [8] |
Teixeira C P, Fernandes C O, Ryan R, Ahern J. Attitudes and preferences towards plants in urban green spaces: implications for the design and management of Novel Urban Ecosystems. Journal of Environmental Management, 2022, 314: 115103. DOI:10.1016/j.jenvman.2022.115103 |
| [9] |
Moser A, Rötzer T, Pauleit S, Pretzsch H. Structure and ecosystem services of small-leaved lime (Tilia cordata Mill.) and black locust (Robinia pseudoacacia L.) in urban environments. Urban Forestry & Urban Greening, 2015, 14(4): 1110-1121. |
| [10] |
Esperon-Rodriguez M, Rymer P D, Power S A, Challis A, Marchin R M, Tjoelker M G. Functional adaptations and trait plasticity of urban trees along a climatic gradient. Urban Forestry & Urban Greening, 2020, 54: 126771. |
| [11] |
张小燕, Alison Kim Shan Wee, Tadashi Kajita, 朱俊杰, 曹坤芳. 两种红树植物不同种源幼苗对模拟高温热浪的光合生理响应差异. 植物科学学报, 2021, 39(4): 424-432. |
| [12] |
Lawren S, Christine S. Leaf venation: structure, function, development, evolution, ecology and applications in the past, present and future. The New Phytologist, 2013, 198(4): 983-1000. DOI:10.1111/nph.12253 |
| [13] |
张富崇, 于明含, 何莹莹, 王春媛, 丁国栋, 唐颖. 毛乌素沙地降水格局变化对油蒿木质部解剖特征的影响. 生态学报, 2023, 43(16): 6661-6669. |
| [14] |
贺鹏程, 叶清. 基于植物功能性状的生态学研究进展: 从个体水平到全球尺度. 热带亚热带植物学报, 2019, 27(5): 523-533. |
| [15] |
Choat B, Ball M C, Luly J G, Holtum J A M. Hydraulic architecture of deciduous and evergreen dry rainforest tree species from north-eastern Australia. Trees, 2005, 19(3): 305-311. DOI:10.1007/s00468-004-0392-1 |
| [16] |
McDowell N, Pockman W T, Allen C D, Breshears D D, Cobb N, Kolb T, Plaut J, Sperry J, West A, Williams D G, Yepez E A. Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought?. The New Phytologist, 2008, 178(4): 719-739. DOI:10.1111/j.1469-8137.2008.02436.x |
| [17] |
Yang X D, Anwar E, Zhou J, He D, Gao Y C, Lv G H, Cao Y E. Higher association and integration among functional traits in small tree than shrub in resisting drought stress in an arid desert. Environmental and Experimental Botany, 2022, 201: 104993. DOI:10.1016/j.envexpbot.2022.104993 |
| [18] |
Pavlicev M, Cheverud J M, Wagner G P. Measuring morphological integration using eigenvalue variance. Evolutionary Biology, 2009, 36(1): 157-170. DOI:10.1007/s11692-008-9042-7 |
| [19] |
Joshi J, Stocker B D, Hofhansl F, Zhou S X, Dieckmann U, Prentice I C. Towards a unified theory of plant photosynthesis and hydraulics. Nature Plants, 2022, 8(11): 1304-1316. DOI:10.1038/s41477-022-01244-5 |
| [20] |
Reich P B. The world-wide 'fast-slow' plant economics spectrum: a traits manifesto. Journal of Ecology, 2014, 102(2): 275-301. DOI:10.1111/1365-2745.12211 |
| [21] |
Powell T L, Wheeler J K, Oliveira A A R, Costa A C L, Saleska S R, Meir P, Moorcroft P R. Differences in xylem and leaf hydraulic traits explain differences in drought tolerance among mature Amazon rainforest trees. Global Change Biology, 2017, 23(10): 4280-4293. DOI:10.1111/gcb.13731 |
| [22] |
Cao S P, He Y, Zhang L F, Chen Y, Yang W, Yao S, Sun Q. Spatiotemporal characteristics of drought and its impact on vegetation in the vegetation region of Northwest China. Ecological Indicators, 2021, 133: 108420. DOI:10.1016/j.ecolind.2021.108420 |
| [23] |
Yang X D, Anwar E, Xu Y L, Zhou J E, Sha L B, Gong X W, Ali A, Gao Y C, Liu Y J, Ge P. Hydraulic constraints determine the distribution of heteromorphic leaves along plant vertical height. Frontiers in Plant Science, 2022, 13: 941764. DOI:10.3389/fpls.2022.941764 |
| [24] |
Venturas M D, MacKinnon E D, Jacobsen A L, Pratt R B. Excising stem samples underwater at native tension does not induce xylem cavitation. Plant, Cell & Environment, 2015, 38(6): 1060-1068. |
| [25] |
Sperry J S, Donnelly J R, Tyree M T. A method for measuring hydraulic conductivity and embolism in xylem. Plant, Cell and Environment, 1988, 11(1): 35-40. DOI:10.1111/j.1365-3040.1988.tb01774.x |
| [26] |
Meinzer F C, Johnson D M, Lachenbruch B, McCulloh K A, Woodruff D R. Xylem hydraulic safety margins in woody plants: coordination of stomatal control of xylem tension with hydraulic capacitance. Functional Ecology, 2009, 23(5): 922-930. DOI:10.1111/j.1365-2435.2009.01577.x |
| [27] |
Yang X D, Yan E R, Chang S X, Da L J, Wang X H. Tree architecture varies with forest succession in evergreen broad-leaved forests in Eastern China. Trees, 2015, 29(1): 43-57. DOI:10.1007/s00468-014-1054-6 |
| [28] |
Hultine K R, Koepke D F, Pockman W T, Fravolini A, Sperry J S, Williams D G. Influence of soil texture on hydraulic properties and water relations of a dominant warm-desert phreatophyte. Tree Physiology, 2006, 26(3): 313-323. DOI:10.1093/treephys/26.3.313 |
| [29] |
林金星. 木材制片技术的改进. 植物学通报, 1993, 10(3): 61-64. |
| [30] |
Trifilò P, Lo Gullo M A, Nardini A, Pernice F, Salleo S. Rootstock effects on xylem conduit dimensions and vulnerability to cavitation of Olea europaea L. Trees, 2007, 21(5): 549-556. DOI:10.1007/s00468-007-0148-9 |
| [31] |
Hewitt B R. Spectrophotometric determination of total carbohydrate. Nature, 1958, 182(4630): 246-247. |
| [32] |
Csárdi G, Nepusz T. The igraph software package for complex network research. International Journal of Complex Systems, 2006. |
| [33] |
段洪浪, 吴建平, 刘文飞, 廖迎春, 张海娜, 樊后保. 干旱胁迫下树木的碳水过程以及干旱死亡机理. 林业科学, 2015, 51(11): 113-120. |
| [34] |
Esperon-Rodriguez M, Power S A, Tjoelker M G, Marchin R M, Rymer P D. Contrasting heat tolerance of urban trees to extreme temperatures during heatwaves. Urban Forestry & Urban Greening, 2021, 66: 127387. |
| [35] |
Delzon S, Cochard H. Recent advances in tree hydraulics highlight the ecological significance of the hydraulic safety margin. New Phytologist, 2014, 203(2): 355-358. DOI:10.1111/nph.12798 |
| [36] |
Choat B, Jansen S, Brodribb T J, Cochard H, Delzon S, Bhaskar R, Bucci S J, Feild T S, Gleason S M, Hacke U G, Jacobsen A L, Lens F, Maherali H, Martínez-Vilalta J, Mayr S, Mencuccini M, Mitchell P J, Nardini A, Pittermann J, Pratt R B, Sperry J S, Westoby M, Wright I J, Zanne A E. Global convergence in the vulnerability of forests to drought. Nature, 2012, 491(7426): 752-755. DOI:10.1038/nature11688 |
| [37] |
Blackman A. Alternative pollution control policies in developing countries. Review of Environmental Economics and Policy, 2010, 4(2): 234-253. DOI:10.1093/reep/req005 |
| [38] |
罗丹丹, 王传宽, 金鹰. 木本植物水力系统对干旱胁迫的响应机制. 植物生态学报, 2021, 45(9): 925-941. |
| [39] |
Hacke U G, Sperry J S. Functional and ecological xylem anatomy. Perspectives in Plant Ecology, Evolution and Systematics, 2001, 4(2): 97-115. DOI:10.1078/1433-8319-00017 |
| [40] |
Hacke U G, Sperry J S, Pittermann J. Drought experience and cavitation resistance in six shrubs from the Great Basin, Utah. Basic and Applied Ecology, 2000, 1(1): 31-41. DOI:10.1078/1439-1791-00006 |
| [41] |
Anderegg W R L, Anderegg L D L. Hydraulic and carbohydrate changes in experimental drought-induced mortality of saplings in two conifer species. Tree Physiology, 2013, 33(3): 252-260. DOI:10.1093/treephys/tpt016 |
| [42] |
Ma C C, Gao Y B, Guo H Y, Wang J L, Wu J B, Xu J S. Physiological adaptations of four dominant Caragana species in the desert region of the Inner Mongolia Plateau. Journal of Arid Environments, 2008, 72(3): 247-254. DOI:10.1016/j.jaridenv.2007.05.009 |
| [43] |
Funk J L, Larson J E, Vose G. Leaf traits and performance vary with plant age and water availability in Artemisia californica. Annals of Botany, 2021, 127(4): 495-503. DOI:10.1093/aob/mcaa106 |
| [44] |
Liu C C, Li Y, He N P. Differential adaptation of lianas and trees in wet and dry forests revealed by trait correlation networks. Ecological Indicators, 2022, 135: 108564. DOI:10.1016/j.ecolind.2022.108564 |
| [45] |
Ziegler C, Coste S, Stahl C, Delzon S, Levionnois S, Cazal J, Cochard H, Esquivel-Muelbert A, Goret J Y, Heuret P, Jaouen G, Santiago L S, Bonal D. Large hydraulic safety margins protect Neotropical canopy rainforest tree species against hydraulic failure during drought. Annals of Forest Science, 2019, 76(4): 115. DOI:10.1007/s13595-019-0905-0 |
| [46] |
Villagra M, Campanello P I, Bucci S J, Goldstein G. Functional relationships between leaf hydraulics and leaf economic traits in response to nutrient addition in subtropical tree species. Tree Physiology, 2013, 33(12): 1308-1318. DOI:10.1093/treephys/tpt098 |
| [47] |
Gleason S M, Wiggans D R, Bliss C A, Comas L H, Cooper M, DeJonge K C, Young J S, Zhang H H. Coordinated decline in photosynthesis and hydraulic conductance during drought stress in Zea mays. Flora, 2017, 227: 1-9. DOI:10.1016/j.flora.2016.11.017 |
| [48] |
Tardieu F, Simonneau T. Variability among species of stomatal control under fluctuating soil water status and evaporative demand: modelling isohydric and anisohydric behaviours. Journal of Experimental Botany, 1998, 49: 419-432. DOI:10.1093/jxb/49.Special_Issue.419 |
| [49] |
Domec J C, Johnson D M. Does homeostasis or disturbance of homeostasis in minimum leaf water potential explain the isohydric versus anisohydric behavior of Vitis vinifera L. cultivars? Tree Physiology, 2012, 32(3): 245-248. DOI:10.1093/treephys/tps013 |
| [50] |
Jupa R, Plavcová L, Flamiková B, Gloser V. Effects of limited water availability on xylem transport in Liana Humulus lupulus L. Environmental and Experimental Botany, 2016, 130: 22-32. DOI:10.1016/j.envexpbot.2016.05.008 |