 2024, Vol. 44
2024, Vol. 44文章信息
- 苏立城, 陈晓珊, 罗志忠, 胡英, 陈煜佳, 吴道铭, 曾曙才
- SU Licheng, CHEN Xiaoshan, LUO Zhizhong, HU Ying, CHEN Yujia, WU Daoming, ZENG Shucai
- 氮添加对森林土壤有机碳库固存及CO2排放的影响研究进展
- Effects of nitrogen addition on the organic carbon sequestration and CO2 emissions in forest soils: a review
- 生态学报. 2024, 44(7): 2717-2733
- Acta Ecologica Sinica. 2024, 44(7): 2717-2733
- http://dx.doi.org/10.20103/j.stxb.202305250867
-
文章历史
- 收稿日期: 2023-05-24
- 网络出版日期: 2024-01-12

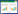




森林生态系统是陆地生态系统中最大的碳库, 森林土壤则是陆地生态系统中最大的有机碳库, 据权威研究估算, 全球有约50% 的地上碳和40% 的地下碳储存在森林生态系统中, 目前全球森林生态系统的碳储量达到了(861±66) Gt, 其中森林土壤的碳储量约为(383±30) Gt[1]。森林生态系统不仅是温室气体的“源”, 也是温室气体的“汇”, 森林土壤呼吸作用所释放的二氧化碳(CO2)是温室气体的重要来源之一, 且极易受人类活动的干扰[2], 近年来, 由于温室效应加剧, 围绕如何提高森林土壤有机碳库固存能力并减缓CO2排放的研究受到了广泛关注。
氮是限制植物生长和土壤微生物生命活动的主要营养元素, 外源氮的添加对提高森林净初级生产力、促进森林可持续发展而言具有重要意义。但在森林经营过程中, 氮沉降或施肥措施引起的大量氮素携入可能会改变土壤养分供应和理化性质稳态, 导致土壤酸化, 从而增加土壤微生物生命活动的环境压力并改变其群落结构特征和生态功能[3], 具体表现为影响输入有机质(如凋落物、细根周转和根系分泌物等)的降解、固定和土壤有机碳(SOC)的矿化过程, 最终影响森林土壤的碳循环过程并改变土壤有机碳库的固存和CO2排放通量[4—5]。由于森林土壤中的碳、氮循环过程存在紧密的耦合作用, 因此氮也是影响森林土壤碳循环过程的重要因素之一, 即使森林土壤中的氮含量发生微小变化, 也可能会破坏森林土壤的碳氮平衡, 继而影响有机碳库的稳定性和改变CO2的排放通量, 对全球气候变化产生重大影响[6]。因此, 明确森林土壤碳循环过程对氮添加的响应, 对预测全球气候变化条件下森林土壤有机碳库稳定性及寻求降低CO2排放通量的方法而言具有重要的指导意义。
1 森林生态系统的土壤碳循环过程按照赋存形态, 土壤中的碳可分为无机碳和有机碳两类, 无机碳包括CO2(吸附的CO2和溶解的CO2)、碳酸(H2CO3)、重碳酸盐(HCO3-)、碳酸盐(CO32-)及固态的碳酸钙(CaCO3), 性质较稳定[7]; 有机碳主要来源于死亡的动植物残体、牲畜粪便和绿肥等有机物料, 经土壤微生物的分解腐化后形成一类特殊、复杂的高分子有机化合物, 即土壤有机质(SOM), 其中所含的碳元素为SOC[8]。森林生态系统的土壤碳循环主要可分为3个过程, 包括:①动物粪便及残体、植物地上凋落物和根系生命活动的碳输入过程; ②土壤碳库不同形态碳组分之间的迁移、转化过程; ③土壤呼吸作用和可溶性有机碳(DOC)、无机碳的解吸附、溶解的碳输出过程。这3个过程并不存在先后顺序, 是共同进行且相互交织的。
经研究发现, 森林土壤的碳输入途径主要有两条(图 1):(1)植物地上凋落物和细根周转产物在土壤微生物的作用下分解腐化为SOM[9]; (2)植物根系生命活动过程中的渗出物(小分子有机酸、氨基酸和糖等)和分泌物(有机酸、酶类、植物激素、氨基酸和糖类等)所形成的根际沉积碳经土壤微生物吸收后转化为SOM[10]。森林土壤中碳组分的迁移和转化是指: 输入的有机物质在土壤微生物的作用下, 或经由微生物呼吸作用大部分吸收转化为微生物生物量碳组分(MBC)、胞外多糖或腐殖质等SOM, 未被吸收的碳素则会矿化为CO2释放; 或经由植物根系呼吸作用一部分碳素参与根系生命活动, 另一部分矿化为CO2释放。但土壤中被矿化的CO2并非全部都会释放到大气中, 其中的一部分CO2会经由土壤淋溶和沉淀作用通过SOC→CO2→HCO3-(aq)→CaCO3(s)途径转化为碳酸盐补充土壤无机碳库[11—12]。而转化为SOM固存在土壤有机碳库中的SOC也不稳定, 在外界因素的作用下会发生迁移与转化。研究表明, 土壤中不同性质和功能的碳组分之间会相互影响和转化, 而土壤有机碳库和无机碳库之间同样也可以相互作用和转化[13]。森林土壤的碳输出是指土壤呼吸作用和无机碳解吸附、溶解导致的CO2释放过程, 其中绝大部分的CO2由土壤呼吸所释放, 包括生物学过程(植物根系呼吸、土壤微生物呼吸和土壤动物呼吸)和非生物学过程(土壤有机质的化学氧化分解过程), 而植物根系和土壤微生物呼吸作用所释放的CO2是森林土壤碳循环中最重要的碳输出过程[14]。

|
| 图 1 森林生态系统中的土壤碳循环过程 Fig. 1 Schematic overview of soil carbon cycling in forest ecosystem |
相关Meta分析表明[15—18], 森林土壤的碳循环过程极其复杂, 不仅与其他元素如氮、磷循环过程相互耦合, 同时还受到气候、森林经营、人为活动、土壤理化环境以及土壤微生物、相关酶活性等众多因素的共同影响。
2 氮添加对森林土壤有机碳库固存的影响 2.1 氮添加对森林土壤有机碳输入的影响氮是植物生长的主要限制性营养元素, 不仅是植物体内蛋白质和叶绿素的主要成分, 也是核酸、各类酶、磷脂和某些生长激素的组成成分, 有“生命元素”之称, 可促进或抑制植物的生长固碳[19]。促进作用表现为当氮适量时, 可促进植物体合成蛋白质、提高相关酶活性来增加叶面积和叶绿素含量, 进而提高光合作用速率和碳同化能力[20]; 抑制作用表现为当氮添加不足或过量时, 造成植物体内营养失衡, 抑制植物叶片形态结构和生理功能, 导致植物光合作用过程受限, 降低碳同化速率[21]。因此, 氮添加可通过促进或抑制植物光合作用来影响植物的生长发育和有机物积累, 继而改变植物的地上凋落物产量、化学组成和地下碳分配格局来影响有机物质的回归, 并调节土壤微生物的相关特性来影响SOC输入[22]。即氮添加对SOC输入的影响主要可表现为两个方面:一是通过影响植物生长固碳后地上凋落物的相关特性和微生物的分解速率来影响SOC输入[23—24]; 二是通过影响植物的地下碳分配格局来调节细根周转速率和改变根系分泌物、渗出物的产生来影响SOC输入[25—27]。
第一方面, 氮添加可能会改变凋落物的产量和化学组成, 同时通过影响土壤微生物群落特征来调节凋落物的分解速率, 由此影响凋落物释放SOC[28—29]。研究表明, 氮添加会改变森林土壤的养分供应差异来影响植物的生长和碳同化, 直接促进或抑制凋落物的产生并调节凋落物的数量和化学组成(C/N、木质素/N等), 这会导致凋落物的可分解性(难易程度)和分解速率改变[30—31]; 其次, 氮添加引起的森林土壤理化性质改变和养分供应差异也会影响土壤微生物的群落组成、活性和功能多样性, 进而促进或抑制凋落物分解腐化为SOC组分[32—33]。
氮的添加会促进森林生态系统的净初级生产力[34]。其中, 当氮添加量较低时, 凋落物产量有所提高且基质的C/N和木质素/N呈下降趋势, 致使其所含的耐分解化合物成分减少, 同时提高了相关微生物和酶的活性, 促进凋落物分解腐化为SOM, 由此增加SOC输入[35—36]。而当氮添加量较高时, 尽管凋落物的产量和基质成分与较低量氮添加时相比无明显差异, 但由于土壤微生物和酶活性的下降导致其分解腐化凋落物的速率明显受限, 对SOC输入的贡献则表现为无显著影响或略有上升[37]。Hobbie等人[38]发现, 适量氮添加提高了森林的凋落物数量, 并使凋落物中木质素含量显著下降, 其分解腐化速率显著提高。Carly等[39]的研究则证实凋落物数量的增加可以提高森林土壤中己糖和酚酸含量, 刺激了土壤微生物和酶的活性而提高凋落物的分解速率。Thomas等[40]和Xia等[41]也发现, 在低氮添加水平下(30 kg N hm-2 a-1), 森林土壤中的木质素分解酶类活性显著降低, 有效减缓了腐殖质的分解速率, 增加了土壤中难分解有机碳的积累。而付琦等[42]通过梯度氮添加试验发现, 施氮增加了森林的凋落物数量和N含量, 即降低了基质的C/N, 促进SOC积累, 但随着施氮量增加, 年SOC输入量呈下降趋势。Saiya-Cork等[43]的研究也表明, 低氮添加量(30 kg N hm-2 a-1)使森林土壤中的纤维素酶、脲酶、酸性磷酸酶和糖苷酶等活性显著升高, 显著促进凋落物分解为SOM, 但是随着氮添加量的增加, 相关酶活性显著降低, SOM量下降。郭绮雯等[44]的梯度氮添加研究表明, 中低浓度的氮添加显著促进了凋落物中纤维素和半纤维素等易分解组分的分解, 并释放SOC, 而高氮添加(300 kg N hm-2 a-1)下则表现为抑制作用。周世兴等[45]的研究则发现, 氮添加会抑制凋落物中木质素和纤维素等难分解碳组分的分解, 降低SOC的输入, 且抑制作用随氮添加量的增加而显著加强。
第二方面, 氮添加通过改变根际土壤的理化性质或养分有效性来调节根系生命活动, 此过程中, 由于根系的生理更新作用, 不断产生的细根一部分会替代老根, 其余的则会和被替代的老根一起凋亡分解, 该过程称为细根周转, 细根周转产物即根系凋落物[46]; 同时, 根系在生命活动过程中也会向土壤释放出有机物质, 包括碳水化合物、氨基酸、有机酸、脂肪酸、肽、核苷和酶类等, 称为根系分泌物和根系渗出物[47]。氮添加通过调节根际养分有效性可影响细根周转速率以及根系的生物量和化学组成, 或改变根际土壤的理化性质, 使根系响应继而主动或被动向土壤释放有机物质(分泌物或渗出物)[48]。根系凋落物和根系分泌物、渗出物都属于有机物质, 是森林土壤有机碳库的重要来源之一。
细根周转途径输入SOC的过程与细根产量、周转速率以及分解速率密切相关。研究表明, 适量氮添加会提高森林植物的细根产量和周转速率而增加根系凋落物的产量, 同时会降低根系凋落物的C/N和木质素/N; 且随着氮添加量增多, 倾向于提高根际胞外酶中纤维素酶活性并抑制木质素酶活性[49]。
一项针对全球不同森林生态系统细根周转对氮添加响应的荟萃分析就表明, 氮添加使森林植物细根产量平均增加了27%, 细根周转速率及土壤碳储量均有效提高[50]。Li等[51]研究发现, 当氮添加量不超过50 kg N hm-2 a-1时, 森林的细根产量和周转速率随氮添加量的增加而提高, 且土壤碳储量与细根产量之间存在显著正相关。而Ferreira等[52]发现, 在氮添加量高于60 kg N hm-2 a-1时会抑制植物细根的生物量和周转速率, 且抑制程度随添加量上升而加重。Li等[53]则发现当氮添加量高于100 kg N hm-2 a-1时, 细根生物量和周转速率会受到显著抑制。Sun等[54]发现, 氮添加使5种不同森林类型的细根C/N和木质素/N都呈下降趋势, 且提高了细根周转速率, 但根系凋落物的分解速率均随氮添加量的升高而显著下降, 分析根际胞外酶的活性后发现, 氮添加促进了纤维素酶活性而抑制了木质素活性。关于土壤微生物如何响应氮添加来调节胞外酶分泌和活性, 有研究就指出:氮限制条件下, 氮添加会提高土壤养分有效性, 导致更多的碳向微生物分配, 促进微生物分泌胞外酶, 从而提高微生物体外修饰作用, 此举有利于促进分解和降解外部有机物并转化为SOC; 而当氮添加过量时, 土壤的酸化作用抑制会微生物分泌可降解复杂底物的胞外酶活性(如分解木质素的过氧化物酶、酚氧化酶等), 从而降低微生物对根系凋落物的分解速率和释放SOC[55]。Carrara等[56]发现在高氮添加量下植物会降低地下碳分配和土壤微生物活性, 导致根系生物量下降了24.7%、根际AM真菌生物量减少了55.9%、胞外酶数量下降18.7%, SOC转化率大大降低。上述的相关研究表明, 氮添加会降低植物细根的C/N和木质素/N, 同时调节土壤微生物胞外酶的分泌及其活性来影响可分解性和分解速率来输入SOC。
此外, 根系分泌物和渗出物所形成的碳沉积也是SOC的重要来源, 其中以根系分泌物为主。有研究表明, 植物通过根系分泌物形式向根际土壤输入的有机化合物约占光合产物的5%—21%, 分泌物的数量和成分受到植物种类、林龄以及根际土壤微环境等的影响[57]。根系分泌物的释放主要受根际土壤养分有效性的调节, 低养分有效性条件下会限制根系分泌物的分泌, 而高养分有效性可促进根系分泌物的释放, 氮添加则倾向于增加根际土壤的养分有效性[58—60]。Qiao等[61]对亚高山针叶树种进行了氮添加试验, 发现氮添加下植物根系分泌物显著增多, 以酚酸类化合物为主。庞丽等[62]研究了氮添加对马尾松根系分泌物酸性磷酸酶和有机酸的影响, 结果显示, 马尾松不同家系根系分泌物对氮添加的响应均表现为促进作用。Ataka等[63]测定氮添加处理下寒温带森林根系分泌物的分泌速率, 结果显示氮添加显著促进了根系分泌物的分泌速率和产量, 使根际土壤中的DOC含量显著提高。Iiu等[64]也发现, 氮添加使温带森林植物白杨和白桦根系分泌物的分泌速率和根际土壤总有机碳(TOC)含量显著提高。因此, 氮添加倾向于促进根系分泌物和渗出物的释放, 同时调节基质的化学计量比及其可分解性, 并最终被土壤微生物吸收转化。Berhongaray等[65]的研究结论显示, 森林生态系统的SOC输入主要来源于植被地下碳分配途径的贡献, 地下碳分配(根系凋落物和根系分泌物、渗出物等)对SOC的转化效率高达76%;而地上凋落物对SOC转化效率仅约9%, 绝大部分经由微生物矿化作用转化为CO2释放。这与Clemmensen等[66]、Cotrufo等[67]的研究结论一致。
由于氮添加使森林生态系统地上地下凋落物、根系分泌物和渗出物等有机质增多, 土壤养分有效性提高, 刺激了土壤微生物的体内修饰来促进外源有机质的分解, 这提高了土壤的呼吸作用, 使CO2排放通量升高。同时, 受到外源有机质的刺激, 土壤中原有的SOM会产生激发效应, 加速SOC的矿化, 该过程也会使土壤的CO2排放通量升高。因此, 氮添加对凋落物分解腐化向土壤输入SOC的实际影响还存在不确定性。但从全球尺度来看, 绝大部分的氮添加研究, 无论是基于野外的氮沉降观测还是人为的施氮试验, 均提高了森林土壤的碳储量, 即促进森林生态系统的SOC输入[68—70]。
2.2 氮添加对森林土壤有机碳库及其组分的影响森林土壤有机碳库是由不同固存时间和分解阶段的有机质所构成的混合体, 包括植物源SOC和微生物源SOC, 依据功能和性质的差异可分为活性有机碳、缓性(慢性)有机碳和惰性有机碳[71]。其中, 活性有机碳易被吸收、分解和矿化, 是植物营养、土壤微生物能量和CO2释放的主要来源[72]。植物源碳(凋落物、根系凋落物、根系分泌物等)是森林土壤最主要的碳输入, 并在土壤微生物的介导下参与土壤碳循环过程。由于土壤中的碳、氮循环过程是紧密耦合的, 因此氮添加所引起的土壤氮循环响应也会改变其中的许多碳循环过程, 进而影响土壤有机碳库及其组分的稳定性[73—74]。氮添加影响森林土壤有机碳库组成和稳定性的途径主要为以下3条:①影响植物碳同化和生理代谢过程, 来改变凋落物、根系分泌物等植物源有机碳在数量及化学成分上的差异; ②改变土壤养分有效性来影响土壤微生物的生命活动, 进而调控SOM的降解、稳定和转化, 来改变不同SOC组分间的含量; ③改变土壤的理化性质如pH、土壤团聚体结构和密度等, 来破坏土壤环境对SOC的物理保护机制, 进而影响土壤有机碳库各组分的稳定性[75—76]。
作为陆地生态系统有机质的主要分解者, 土壤微生物不仅控制了SOC的矿化过程, 同时还是驱动各类有机碳组分降解转化的“动力引擎”, 主导着土壤的碳循环过程[77]。土壤微生物通过“体外修饰”及“体内周转”途径将植物源碳降解转化, 或以残体的形式(腐殖质)储存于土壤中; 或被同化为微生物生物量和自身代谢产物, 以微生物源碳形式储存于土壤中。土壤微生物体外修饰即通过分泌胞外酶来分解或转化大分子植物源碳底物, 使其转化为腐殖质; 土壤微生物体内周转即直接吸收小分子植物源碳底物后经同化作用合成为自身生物量。其中, 经土壤微生物“体内周转”途径所同化的碳素, 会随着微生物的生命活动转化为微生物生物量和自身代谢产物, 并储存在土壤中成为有机碳库的一部分, 这类有机碳组分被称之为微生物源碳。关于以微生物“体内周转”过程为驱动力的微生物源碳形成过程, Liang等[78]在2017年形象的提出了“土壤微生物碳泵(MCP)”的概念, 在该研究中, Liang等人系统地梳理了土壤碳循环过程中土壤微生物调控SOC形成和稳定的相关途径, 并提出了“土壤MCP概念体系”, 这是目前有关土壤微生物介导并参与SOC形成的最新理论[78], Liang等[79]还系统地阐述了土壤MCP概念体系的相关内涵和影响因素。
SOC的激发效应策略可以解释土壤微生物在底物利用中“体内周转”和“体外修饰”作用对氮添加的响应[80]。土壤微生物的群落组成主要受养分有效性驱动, 依据养分吸收偏好的差异可将不同微生物分为r生活史策略(rate-strategistis)和K生活史策略(Kapazitätsgrenze-strategistis)[81]。其中, 细菌多为r策略型微生物, 它们繁殖力强、生殖期短、迭代快, 能快速分解吸收易降解的有机物质并大量投入到繁殖迭代中, 而不是用于维持个体生长, 主要利用外源底物[82]; 而真菌多为K策略型微生物, 通常生长缓慢、发育周期和繁殖周期较长, 所吸收的能量主要用于维持个体生存以提高存活率和竞争能力, 倾向于分解吸收难降解的复杂有机物质, 主要利用原有SOM来获取能量[83]。在SOC矿化过程中, 土壤中有机质的成分及结构差异会影响微生物的底物利用, 进而影响土壤微生物的群落结构和功能, 反之亦然[84]。在氮限制森林生态系统中, 土壤养分有效性偏低, 底物不足, 此时K策略微生物的碳代谢过程占主导地位, 可迅速分泌胞外酶来分解固存的SOM以获取养分, 该过程为SOC正激发效应, 即微生物的氮挖掘机制, 该过程会倾向于降低土壤有机碳库固存并导致活性有机碳组分含量提高[85]。而随着氮的添加, 土壤养分有效性上升, 底物逐渐增多, 此时r策略微生物迅速生长并逐渐占据优势, 优先利用外源底物获取能量, 从而降低对SOM的分解, 该过程为SOC负激发效应, 即微生物底物偏好机制, 该过程有利于SOM的稳定, 并提高了MBC含量, 有利于土壤有机碳库固存[86—87]。因此在富氮森林生态系统中, 不管氮添加与否, 均倾向于r策略微生物主导SOC激发, 促进土壤有机碳库固存, 并提高稳定性有机碳组分含量。
氮添加通过调控底物的化学组成差异来影响土壤微生物碳利用效率(CUE)的高低。CUE是指微生物通过“体内周转”合成每单位生物量碳与所吸收的底物有机碳之比, 该比值的热力学限制为0.88[88]。即微生物在吸收底物转化为自身生物量过程中, 至少需要消耗12%的碳提供能量, 但在实际中, 由于环境条件的限制, 维持土壤微生物生命活动所消耗的碳从未低于12%[89]。研究表明, 底物的可分解性越高则CUE越高, 也就是说, 底物越容易被微生物吸收转化的话, 微生物耗能就越低, 更能促进MBC组分积累而增加土壤有机碳库[90]。根据推导, 若凋落物和根系分泌物等底物所含的复杂化合物成分越低, 则越有利于SOC转化和固存。底物的可分解性取决于其养分有效性。例如, 当底物的C:N:P化学计量比升高时, 基质中的难分解化合物含量也会升高(即高分子量有机物质, 如纤维素、半纤维和木质素等), 土壤微生物在分解底物转化为自身生物量过程中, 会被迫分泌更多的胞外酶去捕获短缺的营养元素来维持所合成的微生物生物量C ∶ N ∶ P化学计量比平衡, 此过程会消耗更多的碳来提供能量[91—92]。由于氮添加提高了土壤养分有效性, 促进森林植物生长和碳同化, 提高了植物的凋落物产量和地下碳分配并降低了基质的C/N和木质素/N, 从而使其分解、转化过程中的CUE有效提高, 因此相关的研究普遍得出氮添加促进了森林土壤有机碳库固存和组分稳定性提高的结论[93—94]。
根据Meta分析, 在全球尺度上, 氮添加(沉降)使森林生态系统碳汇每年增加了276—448Tg, 其中约40% 为森林土壤有机碳库固存[95]。例如, 以氮限制森林生态系统为例, Vandeneden等[96]在为期9年的研究中发现, 氮添加使温带森林土壤中的纤维素酶和多酚氧化酶活性显著下降, 抑制了SOM的分解速率, 引起土壤有机碳库增加的同时也提高了难分解有机碳的含量。当土壤形成富氮环境时亦是如此, Hyvonen等[68]、Du等[97]的研究均表明, 随着氮添加量增加会导致土壤中有机碳矿化速率降低而促进SOC积累, 并促进DOC组分、LOC组分和ROC组分转化为稳定的有机碳组分固定。Liu等[98]发现, 长期氮添加会降低森林土壤中微生物对惰性有机碳的分解速率, 进而使森林土壤有机碳库中惰性有机碳比例不断上升, 有利于土壤碳库的长期固存。郭洁云等[99]采用整合分析法, 对172篇已发表的森林土壤氮添加论文进行了综合分析, 结果表明氮添加显著增加了森林凋落物产量, 显著降低了凋落物的C/N; 绝大多数的研究都显示氮添加提高了土壤有机碳库固存并改变了各组分含量, 使有机质层土壤中活性有机碳的DOC和MBC组分含量提高, 而矿质层土壤中活性有机碳的DOC和MBC组分含量下降, 惰性有机碳含量提高。
除了上述土壤微生物介导所引起的土壤有机碳库及其组分含量对氮添加的响应变化过程, 土壤的物理保护机制也对氮添加下的土壤有机碳库固存和稳定过程起到了一定作用[100—101]。土壤微生物源源不断的迭代过程会导致微生物死亡残体和部分代谢产物持续增加, 这些微生物残留物类有机质随着时间的推移趋向于在土壤中积累并稳定固存, 该过程称之为“续埋效应”[78]。研究认为, 在“续埋效应”过程中, 土壤团聚体可吸收SOC形成团聚体结合态有机碳, 使其避免被分解, 从而促进有机碳库稳定[102—103]; 此外, 土壤的矿物质也可以吸附SOC或直接与其结合形成矿物结合态有机碳[104], 该现象被Xiao等[105]称为矿物碳泵效应。目前认为, 氮添加会有利于土壤团聚体形成并吸收SOC[106]。此外, 相关研究表明, 氮添加会改变SOC的垂直分布格局, 使有机质层SOC向矿质层转移, 并在矿物质的物理保护作用下成为矿物结合态有机碳组分(惰性SOC), 增加有机碳库的稳定性[107—109]。
如表 1列举的研究结果所示, 受氮添加量、施氮方式、施氮持续时间以及气候、土壤基质和森林植被类型等因素的共同影响, 森林土壤有机碳库中不同组分对氮添加的响应并非呈现线性关系。尽管未能揭示不同森林生态系统土壤有机碳库对氮添加的具体响应规律, 但梳理氮添加对不同森林生态系统SOC输入、转化和分解过程的相关研究结论, 有助于预测未来氮沉降(施氮)对不同森林生态系统碳汇功能的影响, 以便及时调整施肥策略以提高森林碳汇能力缓解全球温室效应。
| 气候带及森林类型Climate zones and forest type | 氮添加量Nitrogen addition | 结论Conclusions | 参考文献References |
| 热带山地雨林Tropical montane rainforests | 50 kg N hm-2 a-1 | 土壤TOC含量随着氮添加持续时间的延长而增加; 氮添加使土壤有机碳库各组分呈现垂直分布格局差异, 具体表现为表层土壤中DOC组分含量显著提高, 但深层土壤中含量反而下降, 腐殖质(稳定性SOC)含量提高。 | [110] |
| 热带雨林Tropical rain forest | 0 kg N hm-2 a-1; 50 kg N hm-2 a-1 | 在氮添加7 a后, 土壤中AM真菌生物量显著减少, 胞外酶活性降低, MBC含量显著降低, 稳定性有机碳组分含量下降, 土壤有机碳库发生减持。 | [111] |
| 亚热带常绿阔叶林Subtropical evergreen broad-leaved forest | 100 kg N hm-2 a-1 | 为期5 a的氮添加对森林土壤TOC、C/N影响均不显著, 但活性SOC含量表现为随土层深度增加而减小的趋势。 | [112] |
| 亚热带常绿阔叶林Subtropical evergreen broad-leaved forest | 0 kg N hm-2 a-1; 50 kg N hm-2 a-1; 100 kg N hm-2 a-1; 150 kg N hm-2 a-1 | 所有氮添加处理均显著促进了土壤中纤维素酶、葡萄糖苷酶和酚酸过氧化物酶的活性, 土壤有机碳库固存随氮添加量的增加而增多, 但MBC的含量却随氮添加的增多而显著降低。 | [113] |
| 亚热带高山常绿阔叶林Subtropical alpine evergreen broad-leaved forest | 0 kg N hm-2 a-1; 50 kg N hm-2 a-1; 150 kg N hm-2 a-1; 300 kg N hm-2 a-1 | 氮添加使凋落物中的C/N下降, 促进有机碳库固存, 且SOC中的MBC组分含量随氮添加量增加呈下降趋势, 高氮(300 kg N hm-2 a-1)下显著抑制。 | [114] |
| 温带针阔混交林Temperate mixed coniferous and broad-leaved forest | 0 kg N hm-2 a-1; 40 kg N hm-2 a-1; 80 kg N hm-2 a-1; 120 kg N hm-2 a-1 | 土壤中TOC含量、活性有机碳含量、颗粒态有机碳含量均随氮添加量增加呈先上升后下降的趋势; 与对照相比, 氮添加促进了土壤团聚体形成, 并与SOC结合成团聚体结合态有机碳, 促进SOC积累和稳定。 | [115] |
| 温带落叶针叶Temperate deciduous-coniferous forest | 50 kg N hm-2 a-1 | 长期的氮添加(20 a)促进了土壤有机碳库固持, 且导致SOC垂直分布格局发生改变, 上层土壤(0—30 cm)活性SOC含量显著提高, 而下层土壤中活性SOC含量下降。 | [116] |
| 温带亚高山针叶林Temperate subalpine coniferous forest | 25 kg N hm-2 a-1 | 长期的(17 a)氮添加使土壤pH下降, 导致SOC含量下降了3.9%, 其中的MBC组分含量下降了22%。分析发现SOC含量减少是由于SOM的分解速率提高以及MBC输入量减少所致。 | [85] |
| 温带针叶林、阔叶林Temperate coniferous forests and broad-leaf forests | 0 kg N hm-2 a-1; 50 kg N hm-2 a-1; 150 kg N hm-2 a-1 | 氮添加使土壤有机碳库显著增加, 但SOC各组分对不同氮添加水平的响应机制存在差异:低氮处理下土壤微生物活性提高、活性SOC含量显著增加, 而高氮处理下土壤微生物活性显著下降, 难分解SOC含量显著提高。 | [117] |
| 青藏高原高寒针叶林Alpine coniferous forests of the Tibetan plateau | 0 kg N hm-2 a-1; 8 kg N hm-2 a-1; 40 kg N hm-2 a-1 | 氮添加显著增加了土壤表层SOC含量, 并促进了土壤团聚体的形成, 与活性SOC结合为团聚体结合态有机碳, 并向深层土壤转移。 | [118] |
| TOC:总有机碳Total organic carbon; DOC:可溶性有机碳Dissolved organic carbon; SOC:土壤有机碳Soil organic carbon; a:年Annus; MBC:微生物量碳Microbial biomass carbon; C:碳Carbon; N:Nitrogen; SOM:Soil organic matter | |||
释放CO2是土壤碳循环中的碳输出过程, 由土壤呼吸作用和无机碳解吸附、溶解过程产生, 其中主要由土壤呼吸作用所释放。土壤呼吸作用包括植物根系自养呼吸、土壤微生物和动物的异氧呼吸以及含碳化合物的化学氧化过程[119]。研究表明, 森林土壤释放CO2的途径主要来源于植物根系自养呼吸和土壤微生物异氧呼吸, 其中又以土壤微生物的异氧呼吸为主, 占森林土壤CO2总排放量的60% 以上[120—121]。土壤微生物通过呼吸作用氧化有机物质来获取能量并释放CO2, 用以维持其生命活动和生态功能, 即SOC矿化, 该过程的速率和程度会直接影响CO2排放, 而通过SOC矿化途径所释放的CO2可占到森林土壤CO2总排放量的70% 以上[122—123]。因此, 微生物呼吸作用的大小可表征土壤的CO2排放强度。
在氮添加条件下, 土壤有机碳库与SOC矿化、土壤微生物之间会发生复杂的交互联系。如图 2所示, 氮添加会提高土壤的养分有效性, 促进植物根系养分吸收, 提高了植物的碳同化能力, 继而增加了植物源碳输入, 在土壤微生物介导下, 植物源碳输入会产生激发效应引起SOC矿化, 而土壤微生物的活性和生物量又受氮素有效性的调控。同时, 氮添加会促进植物根系生命活动, 由此提高根系自养呼吸释放CO2; 氮添加引起的土壤酸化也会诱导土壤无机碳溶解释放CO2[97]。因此, 目前研究结论可以明确的是:①氮添加提高了植物的根系呼吸作用, 因此促进了根系途径释放CO2[124]; ②氮添加降低了土壤pH, 促进了土壤无机碳的溶解并释放CO2[125]。除此以外, 由于土壤有机碳的激发效应存在正激发和负激发, 正激发会促进SOC积累而减少CO2排放, 负激发会促进SOC分解而增加了CO2排放, 而激发效应的方向受到外源有机质底物数量和化学组成、土壤微生物群落组成和功能、土壤环境条件等众多因素的影响, 导致激发效应途径的CO2排放与否难以确定, 因此难以准确判断不同森林土壤CO2排放通量对氮添加的响应规律。总的来说, 不同森林土壤CO2排放对氮添加的响应分为促进、抑制和无显著影响[126—128]。

|
| 图 2 氮添加对森林土壤CO2排放的影响机制 Fig. 2 Mechanism of the effect of nitrogen addition on CO2 emissions from forest soil |
在氮限制森林中, 氮添加提高了土壤养分有效性, 倾向于促进土壤微生物活性和植物生长, 由此提高森林土壤的呼吸作用而促进CO2排放。促进机制包括:①氮添加促进植物生长发育使根系生物量增多, 提高了根系自养呼吸速率; ②氮添加促进了森林植物的光合碳同化, 其凋落物回归和地下碳分配为土壤提供了更多底物, 促进了SOC矿化的正激发效应; ③氮添加为土壤微生物提供了氮源而促进其生长繁殖, 提高了异氧呼吸速率, 同时土壤微生物活性的提高进一步促进了SOC矿化的正激发效应。
因此, 在氮限制的森林生态系统中, 氮添加往往可促进森林土壤释放CO2。王泽西等[129]研究发现, 氮添加显著促进了氮限制高山森林土壤呼吸速率, 使森林土壤呼吸通量提高了11%—15%, CO2排放通量随施氮量增加先升高后降低, 在中氮添加水平(50 kg N hm-2 a-1)达到最高。Strömgren等[130]在温带氮限制云杉林中连续施氮22年, 发现森林土壤有机质层和矿质层SOC矿化速率分别提高了118%和19%, 土壤CO2排放通量提高了31%。Sun等[131]发现氮添加不仅显著促进了青藏高原地区高山针叶林土壤活性有机碳的矿化, 还促进了惰性有机碳的分解, 使土壤CO2排放通量明显提高。一些研究发现, 氮添加量和氮素形态会影响其促进森林土壤CO2排放的效果。Sun等[131]发现, 在养分贫瘠地区, 森林土壤的CO2排放量随氮添加量升高而显著增多。李睿达等[132]在热带桉树人工林施肥研究中发现氮添加显著促进了土壤CO2排放, 且随着施氮量的增加而显著增多。Geng等[133]在温带养分贫瘠的针阔混交林中开展氮添加试验, 发现当添加量低于100 kg N hm-2 a-1时土壤CO2排放通量显著提高, 而当氮添加量超过100 kg N hm-2 a-1后, 会显著抑制CO2排放。Li等[134]对青藏高原针阔混交林开展的梯度氮添加结论也与该研究一致。
3.3 氮添加对森林土壤CO2排放的抑制作用持续的氮添加会造成土壤酸化, 而土壤pH过低被认为是毒害效应, 会使土壤微生物活性降低, 呼吸作用受限, 继而抑制CO2的排放量[135]。由此可得, 氮添加抑制森林土壤CO2排放的机制可能是:①氮添加降低了土壤的C/N和pH, 造成土壤微生物活性和生物量下降, 因此降低了微生物的异氧呼吸; ②氮添加使根际土壤养分失衡, 使植物根系生物量和细根周转速率受到影响, 从而降低了根系的自养呼吸; ③氮添加造成的氮富集效应抑制了土壤微生物和胞外酶活性, 降低微生物群落的碳源获取能力, 提高了CUE, 降低了SOC矿化的正激发效应, 从而减少了CO2释放[136]。
大量研究表明, 在非氮限制的森林生态系统中, 氮添加通常会抑制土壤微生物的活性而降低SOC的矿化速率, 从而抑制了森林土壤的CO2排放[95]。无论是室内培养试验还是野外试验, 若土壤属于富氮状态, 氮添加均有可能会抑制土壤CO2排放[137]。Wang等[138]采集了富氮的亚热带针叶林的土壤进行室内氮添加培养试验, 发现氮添加使土壤呼吸速率下降了6.6%, SOC矿化速率明显抑制。Lu等[139]在热带富氮森林中开展野外氮添加研究, 发现不同梯度的氮添加均显著降低了SOC矿化速率, CO2排放通量与对照相比显著减少。Zhang等[140]在亚热带富氮森林中开展的梯度氮添加试验也与Lu等人的研究结论一致。Micks等[141]则发现, 在温带富氮森林中, 氮添加使土壤呼吸速率下降, 同时CO2排放通量受到抑制。
氮添加量和施氮时间是影响森林土壤CO2排放通量的关键因素[142]。短期的低氮添加量通常促进土壤CO2排放或无影响、而长期的低氮添加量往往可以促进土壤CO2排放; 短期的高氮添加量通常抑制土壤CO2排放或无影响、而长期的高氮添加量往往可以抑制土壤CO2排放[143—144]。Forsmark等[145]在氮贫瘠的温带针叶中开展了为期12年的梯度氮添加试验, 发现土壤呼吸作用对低氮添加量的响应并不敏感, 土壤CO2排放通量与对照相比无显著差异, 但高氮添加量下微生物呼吸作用受到抑制, 土壤CO2排放通量显著降低。张雪等[146]通过盆栽试验, 证实短期的低氮添加对马尾松盆栽土壤CO2的释放量有促进作用, CO2释放速率在氮添加后的短期内显著提高, 但在施氮后期迅速下降。而Chen等[147]则发现, 高氮添加(100 kg N hm-2 a-1)初期对土壤呼吸速率并无显著影响, 但随着时间的推移, 土壤呼吸速率显著降低, CO2排放通量明显下降。
3.4 氮添加对森林土壤CO2排放量无显著影响由于森林土壤所释放的CO2主要来源于土壤的呼吸作用, 若氮添加并未对森林土壤的呼吸作用产生显著影响, 那么森林土壤CO2排放量对氮添加的响应特征可能也表现为无显著影响。关于氮添加对森林土壤CO2排放并未产生显著影响的原因, 可能受氮添加量不足和施氮时间过短的影响。有研究表明, 短期的氮添加可能不会改变森林土壤的CO2排放通量。Sun等[148]以亚热带湿地松林土壤开展为期1年的氮添加试验, 发现该森林土壤的CO2排放通量并未受到影响。Maestrini等[149]在温带阔叶林中以60 kg N hm-2 a-1的氮添加量每月等量添加1年后, 发现土壤CO2排放通量与对照相比无显著差异。
此外, 在土壤理化性质和微生物的共同作用下, 森林土壤的CO2排放量可能也不会产生显著波动。Allison等[150]在寒带针叶林中开展氮添加试验, 发现氮添加降低了土壤微生物生物量的C/N, 但并未改变微生物的生物量, 对土壤呼吸速率及CO2排放通量无影响。Fernández等[151]在地中海地区的针阔混交林中进行氮添加试验, 发现氮添加主要与土壤温度协同影响森林土壤的呼吸作用, 但与不施肥的森林土壤相比, 总CO2排放量并无显著差异。Mori等[152]发现, 在马来西亚的原始热带雨林地区, 由于受磷限制影响, 氮添加未显著改变土壤自养呼吸和异氧呼吸速率, 从而并未对CO2排放通量造成显著影响。Koehler等[153]在热带森林中进行的氮添加试验(125 kg N hm-2 a-1)同样也显示对森林土壤CO2排放通量无显著影响。在富磷的非热带地区, 也得出了类似Mori等和Koehler等的研究结论, Gao等[154]研究了氮添加对富磷环境的亚热带混交林土壤温室气体排放通量的影响, 发现氮添加对该森林土壤的CO2排放通量无显著影响。
4 结论与展望总体而言, 氮添加通过提高森林土壤的养分有效性而提高了森林的净初级生产力, 促进了凋落物和根际沉积碳(细根周转产物、根系分泌物及渗出物等)产量, 同时降低了上述基质中的大分子化合物含量, 使其可分解性提高; 尽管不同森林生态系统的土壤微生物群落特征及生态功能对氮添加的响应规律存在错综复杂的耦合和交互影响, 但均倾向于促进植物源SOC的积累并趋向于稳定, 有效提高了土壤有机碳库固存和稳定性。氮添加对森林土壤的CO2释放规律并不呈现简单的线性关系, 由于受到底物数量和质量、微生物群落组成和活性、土壤理化性质、气候、植被类型等众多因素的共同影响, 不同森林生态系统的CO2排放量可能表现为促进、抑制或无影响。
森林土壤有机碳库在全球碳循环过程中扮演着重要的角色, 对维系生态系统碳循环稳定和缓解温室效应而言举足轻重。近年来, 林业化肥滥用现象日渐严重, 加剧了温室效应, 在全球气候变暖的背景下, 氮素输入对森林土壤碳循环过程的影响越来越受到关注。由于受到植被类型、土壤理化性质、微生物种类和气候等因素的多重影响, 氮添加对森林土壤有机碳库固存机制及CO2排放的研究尚不全面, 人们对氮添加条件下森林植被的地下碳分配格局规律也缺乏系统认识, 此外森林土壤有机碳库中活性SOC向惰性SOC转化的稳定机制及其与土壤微生物的耦联关系并不清晰。因此, 在今后的研究中可加强以下方面的突破:
(1) 加强长期氮添加对森林土壤有机碳库固存及稳定性影响的研究。研究发现, 土壤有机碳库及其组分通常对短期的氮添加无响应, 而对长期的氮添加有显著响应。建立长期的氮添加对森林土壤有机碳库影响的研究可更好地掌握土壤有机碳库的稳定固存机制, 以便有效预测氮沉降对森林土壤有机碳库的长期动态影响, 这有利于寻找提高森林土壤有机碳库固存能力和降低温室气体的森林碳汇途径。
(2) 加强全球气候变暖背景下土壤微生物碳代谢过程对氮添加(氮沉降)的响应研究。氮添加主要引起土壤氮素有效性增加, 这一变化会直接引起土壤微生物群落响应, 进而影响CUE、改变土壤呼吸作用强度, 而气候变暖也会影响微生物的活性和生态功能, 这进一步造成了森林生态系统CO2排放规律的不确定性。加强上述研究有望缓解目前的温室效应困局。
(3) 进一步探索氮沉降背景下森林植被差异对SOC分解、转化和固定过程的影响机制。明确不同森林生态系统植物多样性对土壤微生物种群结构和生态功能的影响, 如优势植物是否参与(或主导)了构建土壤微生物群落生态功能, 继而影响森林土壤的碳循环过程。
(4) 借助微生物组学技术, 从土壤—植物—微生物互作角度全面研究氮添加下土壤微生物群落及功能多样性改变对森林土壤有机碳输入及输出的影响。借助微生物组学技术可以更全面、系统地解析氮添加对森林土壤微生物群落和功能多样性的影响, 有助于进一步阐明土壤微生物对SOC输入和输出的影响。
(5) 加强氮添加对森林根际碳沉积途径输入SOC的相关机制研究。目前的研究主要集中在氮添加影响下凋落物回归后的土壤碳循环过程响应及相关介导机制, 但森林土壤碳库的碳输入过程还包括植物地下碳分配调节下的根际沉积碳途径, 而该过程对氮添加的响应十分复杂, 关于氮添加下的植物—菌根互作机制、根际分泌物和渗出物响应规律、根际沉积物化学计量学特征等规律尚无明确的科学结论。
(6) 结合其他关键影响因子研究氮添加对森林土壤碳循环过程的耦合效应。森林土壤的碳循环是一个复杂而综合的过程, 受到众多因素的共同影响。因此有必要结合其他关键因子如气候变暖、CO2浓度升高、降雨等来研究氮添加对森林土壤碳循环的耦合作用, 此举有助于分析在全球气候变化下氮添加对森林土壤有机碳库固存的影响。
| [1] |
Pan Y D, Birdsey R A, Fang J Y, Houghton R, Kauppi P E, Kurz W A, Phillips O L, Shvidenko A, Lewis S L, Canadell J G, Ciais P, Jackson R B, Pacala S W, McGuire A D, Piao S L, Rautiainen A, Sitch S, Hayes D. A large and persistent carbon sink in the world′s forests. Science, 2011, 333(6045): 988-993. DOI:10.1126/science.1201609 |
| [2] |
Han M G, Zhu B. Changes in soil greenhouse gas fluxes by land use change from primary forest. Global Change Biology, 2020, 26(4): 2656-2667. DOI:10.1111/gcb.14993 |
| [3] |
Mondal P, McDermid S S, Qadir A. A reporting framework for Sustainable Development Goal 15: multi-scale monitoring of forest degradation using MODIS, Landsat and Sentinel data. Remote Sensing of Environment, 2020, 237: 111592. DOI:10.1016/j.rse.2019.111592 |
| [4] |
Li W B, Zhang H X, Huang G Z, Liu R X, Wu H J, Zhao C Y, McDowell N G. Effects of nitrogen enrichment on tree carbon allocation: A global synthesis. Global Ecology and Biogeography, 2020, 29(3): 573-589. DOI:10.1111/geb.13042 |
| [5] |
林婉奇, 薛立. 基于BIOLOG技术分析氮沉降和降水对土壤微生物功能多样性的影响. 生态学报, 2020, 40(12): 4188-4197. |
| [6] |
Nandal A, Yadav S S, Rao A S, Meena R S, Lal R. Advance methodological approaches for carbon stock estimation in forest ecosystems. Environmental Monitoring and Assessment, 2023, 195(2): 315. DOI:10.1007/s10661-022-10898-9 |
| [7] |
Batjes N H. Total carbon and nitrogen in the soils of the world. European Journal of Soil Science, 2014, 65(1): 10-21. DOI:10.1111/ejss.12114_2 |
| [8] |
Lal R. Soil carbon dynamics in cropland and rangeland. Environmental Pollution, 2002, 116(3): 353-362. DOI:10.1016/S0269-7491(01)00211-1 |
| [9] |
Porcal P, Dillon P J, Molot L A. Temperature dependence of photodegradation of dissolved organic matter to dissolved inorganic carbon and particulate organic carbon. PLoS One, 2015, 10(6): e0128884. DOI:10.1371/journal.pone.0128884 |
| [10] |
Kuzyakov Y, Domanski G. Carbon input by plants into the soil. Review. Journal of Plant Nutrition and Soil Science, 2000, 163(4): 421-431. DOI:10.1002/1522-2624(200008)163:4<421::AID-JPLN421>3.0.CO;2-R |
| [11] |
Yang Y S, Guo J F, Chen G S, Yin Y F, Gao R, Lin C F. Effects of forest conversion on soil labile organic carbon fractions and aggregate stability in subtropical China. Plant and Soil, 2009, 323(1-2): 153-162. DOI:10.1007/s11104-009-9921-4 |
| [12] |
Neu V, Ward N D, Krusche A V, Neill C. Dissolved organic and inorganic carbon flow paths in an Amazonian transitional forest. Frontiers in Marine Science, 2016, 3: 1-15. |
| [13] |
陈宗定, 许春雪, 安子怡, 王亚平, 孙德忠, 王苏明. 土壤碳赋存形态及分析方法研究进展. 岩矿测试, 2019, 38(2): 233-244. |
| [14] |
Xu X F, Schimel J P, Thornton P E, Song X, Yuan F M, Goswami S. Substrate and environmental controls on microbial assimilation of soil organic carbon: a framework for Earth system models. Ecology Letters, 2014, 17(5): 547-555. DOI:10.1111/ele.12254 |
| [15] |
Zhang X Z, Guan D X, Li W B, Sun D, Jin C J, Yuan F H, Wang A Z, Wu J B. The effects of forest thinning on soil carbon stocks and dynamics: A meta-analysis. Forest Ecology and Management, 2018, 429: 36-43. DOI:10.1016/j.foreco.2018.06.027 |
| [16] |
Sun Z L, Liu S G, Zhang T A, Zhao X C, Chen S, Wang Q K. Priming of soil organic carbon decomposition induced by exogenous organic carbon input: A meta-analysis. Plant and Soil, 2019, 443(1-2): 463-471. DOI:10.1007/s11104-019-04240-5 |
| [17] |
De Stefano A, Jacobson M G. Soil carbon sequestration in agroforestry systems: a meta-analysis. Agroforestry Systems, 2018, 92(2): 285-299. |
| [18] |
Aguilera E, Lassaletta L, Gattinger A, Gimeno B S. Managing soil carbon for climate change mitigation and adaptation in Mediterranean cropping systems: A meta-analysis. Agriculture, Ecosystems & Environment, 2013, 168: 25-36. |
| [19] |
Leghari S J, Wahocho N A, Laghari G M, HafeezLaghari A, MustafaBhabhan G, HussainTalpur K. Role of nitrogen for plant growth and development: a review. Advances in Environmental Biology, 2016, 10(9): 209-219. |
| [20] |
Kraiser T, Gras D E, Gutiérrez A G, González B, Gutiérrez R A. A holistic view of nitrogen acquisition in plants. Journal of Experimental Botany, 2011, 62(4): 1455-1466. DOI:10.1093/jxb/erq425 |
| [21] |
Chen B Q, Cao J H, Wang J K, Wu Z X, Xie G S. Development and implementation of site-specific fertilizer recommendation model based on nutrient balance for rubber plantation. Agronomy Journal, 2011, 103(2): 464-471. DOI:10.2134/agronj2010.0244 |
| [22] |
Van Diepen L T A, Frey S D, Sthultz C M, Morrison E W, Minocha R, Pringle A. Changes in litter quality caused by simulated nitrogen deposition reinforce the N-induced suppression of litter decay. Ecosphere, 2015, 6(10): art205. |
| [23] |
Paul E A. The nature and dynamics of soil organic matter: plant inputs, microbial transformations, and organic matter stabilization. Soil Biology and Biochemistry, 2016, 98: 109-126. DOI:10.1016/j.soilbio.2016.04.001 |
| [24] |
Luo Z K, Feng W T, Luo Y Q, Baldock J, Wang E L. Soil organic carbon dynamics jointly controlled by climate, carbon inputs, soil properties and soil carbon fractions. Global Change Biology, 2017, 23(10): 4430-4439. DOI:10.1111/gcb.13767 |
| [25] |
Lu W J, Liu N, Zhang Y J, Zhou J Q, Guo Y P, Yang X. Impact of vegetation community on litter decomposition: Evidence from a reciprocal transplant study with 13C labeled plant litter. Soil Biology and Biochemistry, 2017, 112: 248-257. DOI:10.1016/j.soilbio.2017.05.014 |
| [26] |
Sokol N W, Sanderman J, Bradford M A. Pathways of mineral-associated soil organic matter formation: integrating the role of plant carbon source, chemistry, and point of entry. Global Change Biology, 2019, 25(1): 12-24. DOI:10.1111/gcb.14482 |
| [27] |
Chen Z J, Geng S C, Zhou X Y, Gui H R, Zhang L L, Huang Z Q, Wang M H, Zhang J H, Han S J. Nitrogen addition decreases soil aggregation but enhances soil organic carbon stability in a temperate forest. Geoderma, 2022, 426: 116112. DOI:10.1016/j.geoderma.2022.116112 |
| [28] |
Man M L, Pierson D, Chiu R, Anaraki M T, VandenEnden L, Ye R X, Lajtha K, Simpson M J. Twenty years of litter manipulation reveals that above-ground litter quantity and quality controls soil organic matter molecular composition. Biogeochemistry, 2022, 159(3): 393-411. DOI:10.1007/s10533-022-00934-8 |
| [29] |
Xu X, Shi Z, Li D J, Rey A, Ruan H H, Craine J M, Liang J Y, Zhou J Z, Luo Y Q. Soil properties control decomposition of soil organic carbon: Results from data-assimilation analysis. Geoderma, 2016, 262: 235-242. DOI:10.1016/j.geoderma.2015.08.038 |
| [30] |
Jewell M D, Shipley B, Paquette A, Messier C, Reich P B. A traits-based test of the home-field advantage in mixed-species tree litter decomposition. Annals of Botany, 2015, 116(5): 781-788. DOI:10.1093/aob/mcv105 |
| [31] |
Vigulu V, Blumfield T J, Reverchon F, Bai S H, Xu Z H. Nitrogen and carbon cycling associated with litterfall production in monoculture teak and mixed species teak and flueggea stands. Journal of Soils and Sediments, 2019, 19(4): 1672-1684. DOI:10.1007/s11368-019-02275-w |
| [32] |
Entwistle E M, Romanowicz K J, Argiroff W A, Freedman Z, Morris J J, Zak D R. Anthropogenic N deposition alters the composition of expressed class Ⅱ fungal peroxidases. Applied and environmental microbiology, 2018, 84(9): e02816-17. |
| [33] |
Moore J A M, Anthony M A, Pec G J, Trocha L K, Trzebny A, Geyer K M, van Diepen L T A, Frey S D. Fungal community structure and function shifts with atmospheric nitrogen deposition. Global Change Biology, 2021, 27(7): 1349-1364. DOI:10.1111/gcb.15444 |
| [34] |
Fatemi F R, Fernandez I J, Simon K S, Dail D B. Nitrogen and phosphorus regulation of soil enzyme activities in acid forest soils. Soil Biology and Biochemistry, 2016, 98: 171-179. DOI:10.1016/j.soilbio.2016.02.017 |
| [35] |
Barantal S, Schimann H, Fromin N, Hättenschwiler S. C, N and P fertilization in an Amazonian rainforest supports stoichiometric dissimilarity as a driver of litter diversity effects on decomposition. Proceedings of the Royal Society B-Biological Sciences, 2014, 281(1796): 20141682. DOI:10.1098/rspb.2014.1682 |
| [36] |
Zhang T A, Chen H Y H, Ruan H H. Global negative effects of nitrogen deposition on soil microbes. The ISME Journal, 2018, 12(7): 1817-1825. DOI:10.1038/s41396-018-0096-y |
| [37] |
张晓曦, 周雯星, 王丽洁, 刘楚波, 米皓皓. 模拟氮沉降对凋落物早期混合分解中相互作用的影响. 生态学杂志, 2022, 41(9): 1708-1716. |
| [38] |
Hobbie S E. Interactions between litter lignin and nitrogenitter lignin and soil nitrogen availability during leaf litter decomposition in a Hawaiian montane forest. Ecosystems, 2000, 3(5): 484-494. DOI:10.1007/s100210000042 |
| [39] |
Phillips C A, Elberling B, Michelsen A. Soil carbon and nitrogen stocks and turnover following 16 years of warming and litter addition. Ecosystems, 2019, 22(1): 110-124. DOI:10.1007/s10021-018-0256-y |
| [40] |
Thomas D C, Zak D R, Filley T R. Chronic N deposition does not apparently alter the biochemical composition of forest floor and soil organic matter. Soil Biology and Biochemistry, 2012, 54: 7-13. DOI:10.1016/j.soilbio.2012.05.010 |
| [41] |
Xia M X, Talhelm A F, Pregitzer K S. Long-term simulated atmospheric nitrogen deposition alters leaf and fine root decomposition. Ecosystems, 2018, 21(1): 1-14. DOI:10.1007/s10021-017-0130-3 |
| [42] |
付琦, 邢亚娟, 闫国永, 董雄德, 张军辉, 王庆贵. 北方森林凋落物动态对长期氮沉降的响应. 生态环境学报, 2019, 28(7): 1341-1350. |
| [43] |
Saiya-Cork K R, Sinsabaugh R L, Zak D R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biology and Biochemistry, 2002, 34(9): 1309-1315. DOI:10.1016/S0038-0717(02)00074-3 |
| [44] |
郭绮雯, 段文标, 刘玉萍, 陈立新, 李常准, 于颖颖, 李少然, 李亦菲. 凋落物添加和模拟氮磷沉降对红松凋落物木质素降解和碳释放的影响. 生态学报, 2021, 41(16): 6621-6632. |
| [45] |
周世兴, 黄从德, 向元彬, 韩博涵, 肖永翔, 唐剑东. 模拟氮沉降对华西雨屏区天然常绿阔叶林凋落物木质素和纤维素降解的影响. 应用生态学报, 2016, 27(5): 1368-1374. |
| [46] |
Wang R L, Yu G R, He N A P. Root community traits: scaling-up and incorporating roots into ecosystem functional analyses. Frontiers in Plant Science, 2021, 12: 690235. DOI:10.3389/fpls.2021.690235 |
| [47] |
Bardgett R D, Mommer L, De Vries F T. Going underground: root traits as drivers of ecosystem processes. Trends in Ecology & Evolution, 2014, 29(12): 692-699. |
| [48] |
Badri D V, Vivanco J M. Regulation and function of root exudates. Plant, Cell & Environment, 2009, 32(6): 666-681. |
| [49] |
Li W B, Jin C J, Guan D X, Wang Q K, Wang A Z, Yuan F H, Wu J B. The effects of simulated nitrogen deposition on plant root traits: a meta-analysis. Soil Biology and Biochemistry, 2015, 82: 112-118. DOI:10.1016/j.soilbio.2015.01.001 |
| [50] |
Yuan Z Y, Chen H Y H. A global analysis of fine root production as affected by soil nitrogen and phosphorus. Proceedings Biological Sciences, 2012, 279(1743): 3796-3802. |
| [51] |
Li X W, Zhang C L, Zhang B B, Wu D, Shi Y F, Zhang W, Ye Q, Yan J H, Fu J M, Fang C L, Ha D, Fu S L. Canopy and understory nitrogen addition have different effects on fine root dynamics in a temperate forest: implications for soil carbon storage. New Phytologist, 2021, 231(4): 1377-1386. DOI:10.1111/nph.17460 |
| [52] |
Ferreira G W D, Oliveira F C C, Silva L O G, Souza J J L L, Soares E M B, Araújo E F, Silva I R. Nitrogen alters initial growth, fine-root biomass and soil organic matter properties of a Eucalyptus dunnii maiden plantation in a recently afforested grassland in Southern Brazil. Forests, 2018, 9(2): 62. DOI:10.3390/f9020062 |
| [53] |
Mei L, Gu J C, Zhang Z W, Wang Z Q. Responses of fine root mass, length, production and turnover to soil nitrogen fertilization in Larix gmelinii and Fraxinus mandshurica forests in Northeastern China. Journal of Forest Research, 2010, 15(3): 194-201. DOI:10.1007/s10310-009-0176-y |
| [54] |
Sun T, Dong L L, Wang Z W, Lü X T, Mao Z J. Effects of long-term nitrogen deposition on fine root decomposition and its extracellular enzyme activities in temperate forests. Soil Biology and Biochemistry, 2016, 93: 50-59. DOI:10.1016/j.soilbio.2015.10.023 |
| [55] |
Weand M P, Arthur M A, Lovett G M, McCulley R L, Weathers K C. Effects of tree species and N additions on forest floor microbial communities and extracellular enzyme activities. Soil Biology and Biochemistry, 2010, 42(12): 2161-2173. DOI:10.1016/j.soilbio.2010.08.012 |
| [56] |
Carrara J E, Walter C A, Hawkins J S, Peterjohn W T, Averill C, Brzostek E R. Interactions among plants, bacteria, and fungi reduce extracellular enzyme activities under long-term N fertilization. Global Change Biology, 2018, 24(6): 2721-2734. DOI:10.1111/gcb.14081 |
| [57] |
Jones D L, Hodge A, Kuzyakov Y. Plant and mycorrhizal regulation of rhizodeposition. The New Phytologist, 2004, 163(3): 459-480. DOI:10.1111/j.1469-8137.2004.01130.x |
| [58] |
Zhu S S, Vivanco J M, Manter D K. Nitrogen fertilizer rate affects root exudation, the rhizosphere microbiome and nitrogen-use-efficiency of maize. Applied Soil Ecology, 2016, 107: 324-333. DOI:10.1016/j.apsoil.2016.07.009 |
| [59] |
Huang X Q, Liu S Z, Liu X, Zhang S R, Li L, Zhao H T, Zhao J, Zhang J B, Cai Z C. Plant pathological condition is associated with fungal community succession triggered by root exudates in the plant-soil system. Soil Biology and Biochemistry, 2020, 151: 108046. DOI:10.1016/j.soilbio.2020.108046 |
| [60] |
Vives-Peris V, de Ollas C, Gómez-Cadenas A, Pérez-Clemente R M. Root exudates: from plant to rhizosphere and beyond. Plant Cell Reports, 2020, 39(1): 3-17. DOI:10.1007/s00299-019-02447-5 |
| [61] |
Qiao M F, Xiao J, Yin H J, Pu X Z, Yue B S, Liu Q. Analysis of the phenolic compounds in root exudates produced by a subalpine coniferous species as responses to experimental warming and nitrogen fertilisation. Chemistry and Ecology, 2014, 30(6): 555-565. DOI:10.1080/02757540.2013.868891 |
| [62] |
庞丽, 张一, 周志春, 丰忠平, 储德裕. 模拟氮沉降对低磷胁迫下马尾松不同家系根系分泌和磷效率的影响. 植物生态学报, 2014, 38(1): 27-35. |
| [63] |
Ataka M, Sun L J, Nakaji T, Katayama A, Hiura T. Five-year nitrogen addition affects fine root exudation and its correlation with root respiration in a dominant species, Quercus crispula, of a cool temperate forest, Japan. Tree Physiology, 2020, 40(3): 367-376. DOI:10.1093/treephys/tpz143 |
| [64] |
Liu G C, Yan G Y, Huang B B, Sun X Y, Xing Y J, Wang Q G. Long-term nitrogen addition alters nutrient foraging strategies of Populus davidiana and Betula platyphylla in a temperate natural secondary forest. European Journal of Forest Research, 2022, 141(2): 307-320. DOI:10.1007/s10342-022-01441-z |
| [65] |
Berhongaray G, Cotrufo F M, Janssens I A, Ceulemans R. Below-ground carbon inputs contribute more than above-ground inputs to soil carbon accrual in a bioenergy poplar plantation. Plant and Soil, 2019, 434(1/2): 363-378. |
| [66] |
Clemmensen K E, Bahr A, Ovaskainen O, Dahlberg A, Ekblad A, Wallander H, Stenlid J, Finlay R D, Wardle D A, Lindahl B D. Roots and associated fungi drive long-term carbon sequestration in boreal forest. Science, 2013, 339(6127): 1615-1618. DOI:10.1126/science.1231923 |
| [67] |
Cotrufo M F, Soong J L, Horton A J, Campbell E E, Haddix M L, Wall D H, Parton W J. Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nature Geoscience, 2015, 8(10): 776-779. DOI:10.1038/ngeo2520 |
| [68] |
Schulte-Uebbing L, de Vries W. Global-scale impacts of nitrogen deposition on tree carbon sequestration in tropical, temperate, and boreal forests: A meta-analysis. Global Change Biology, 2018, 24(2): e416-e431. |
| [69] |
Gundale M J. The impact of anthropogenic nitrogen deposition on global forests: Negative impacts far exceed the carbon benefits. Global Change Biology, 2022, 28(3): 690-692. DOI:10.1111/gcb.15959 |
| [70] |
Jia X Y, Zhong Y Q W, Liu J, Zhu G Y, Shangguan Z P, Yan W M. Effects of nitrogen enrichment on soil microbial characteristics: From biomass to enzyme activities. Geoderma, 2020, 366: 114256. DOI:10.1016/j.geoderma.2020.114256 |
| [71] |
Parton W J, Schimel D S, Cole C V, Ojima D S. Analysis of factors controlling soil organic matter levels in great Plains grasslands. Soil Science Society of America Journal, 1987, 51(5): 1173-1179. DOI:10.2136/sssaj1987.03615995005100050015x |
| [72] |
Qi R M, Li J, Lin Z A, Li Z J, Li Y T, Yang X D, Zhang J J, Zhao B Q. Temperature effects on soil organic carbon, soil labile organic carbon fractions, and soil enzyme activities under long-term fertilization regimes. Applied Soil Ecology, 2016, 102: 36-45. DOI:10.1016/j.apsoil.2016.02.004 |
| [73] |
Schmidt M W I, Torn M S, Abiven S, Dittmar T, Guggenberger G, Janssens I A, Kleber M, Kögel-Knabner I, Lehmann J, Manning D A C, Nannipieri P, Rasse D P, Weiner S, Trumbore S E. Persistence of soil organic matter as an ecosystem property. Nature, 2011, 478(7367): 49-56. DOI:10.1038/nature10386 |
| [74] |
Ni H W, Jing X Y, Xiao X, Zhang N, Wang X Y, Sui Y Y, Sun B, Liang Y T. Microbial metabolism and necromass mediated fertilization effect on soil organic carbon after long-term community incubation in different climates. The ISME Journal, 2021, 15(9): 2561-2573. DOI:10.1038/s41396-021-00950-w |
| [75] |
孟祥萍. 麦玉两熟制下氮肥与秸秆还田的农田碳氮固持和温室气体减排效应研究[D]. 杨凌: 西北农林科技大学, 2021.
|
| [76] |
Rosinger C, Rousk J, Sandén H. Can enzymatic stoichiometry be used to determine growth-limiting nutrients for microorganisms? - A critical assessment in two subtropical soils. Soil Biology and Biochemistry, 2019, 128: 115-126. DOI:10.1016/j.soilbio.2018.10.011 |
| [77] |
Smith P, Cotrufo M F, Rumpel C, Paustian K, Kuikman P J, Elliott J A, McDowell R, Griffiths R I, Asakawa S, Bustamante M, House J I, Sobocká J, Harper R, Pan G, West P C, Gerber J S, Clark J M, Adhya T, Scholes R J, Scholes M C. Biogeochemical cycles and biodiversity as key drivers of ecosystem services provided by soils. SOIL, 2015, 1(2): 665-685. DOI:10.5194/soil-1-665-2015 |
| [78] |
Liang C, Schimel J P, Jastrow J D. The importance of anabolism in microbial control over soil carbon storage. Nature Microbiology, 2017, 2(8): 17105. DOI:10.1038/nmicrobiol.2017.105 |
| [79] |
梁超, 朱雪峰. 土壤微生物碳泵储碳机制概论. 中国科学: 地球科学, 2021, 51(5): 680-695. |
| [80] |
Leff J W, Bardgett R D, Wilkinson A, Jackson B G, Pritchard W J, De Long J R, Oakley S, Mason K E, Ostle N J, Johnson D, Baggs E M, Fierer N. Predicting the structure of soil communities from plant community taxonomy, phylogeny, and traits. The ISME Journal, 2018, 12(7): 1794-1805. DOI:10.1038/s41396-018-0089-x |
| [81] |
Dini-Andreote F, Stegen J C, van Elsas J D, Salles J F. Disentangling mechanisms that mediate the balance between stochastic and deterministic processes in microbial succession. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(11): E1326-E1332. |
| [82] |
Lange M, Eisenhauer N, Sierra C A, Bessler H, Engels C, Griffiths R I, Mellado-Vázquez P G, Malik A A, Roy J, Scheu S, Steinbeiss S, Thomson B C, Trumbore S E, Gleixner G. Plant diversity increases soil microbial activity and soil carbon storage. Nature Communications, 2015, 6: 6707. DOI:10.1038/ncomms7707 |
| [83] |
Averill C, Turner B L, Finzi A C. Mycorrhiza-mediated competition between plants and decomposers drives soil carbon storage. Nature, 2014, 505(7484): 543-545. DOI:10.1038/nature12901 |
| [84] |
Wurzburger N, Wright S J. Fine-root responses to fertilization reveal multiple nutrient limitation in a lowland tropical forest. Ecology, 2015, 96(8): 2137-2146. DOI:10.1890/14-1362.1 |
| [85] |
Boot C M, Hall E K, Denef K, Baron J S. Long-term reactive nitrogen loading alters soil carbon and microbial community properties in a subalpine forest ecosystem. Soil Biology and Biochemistry, 2016, 92: 211-220. DOI:10.1016/j.soilbio.2015.10.002 |
| [86] |
Luo Y, Zang H D, Yu Z Y, Chen Z Y, Gunina A, Kuzyakov Y, Xu J M, Zhang K L, Brookes P C. Priming effects in biochar enriched soils using a three-source-partitioning approach: 14C labelling and 13C natural abundance. Soil Biology and Biochemistry, 2017, 106: 28-35. DOI:10.1016/j.soilbio.2016.12.006 |
| [87] |
Zhang Q F, Feng J G, Li J, Huang C Y, Shen Y W, Cheng W X, Zhu B A. A distinct sensitivity to the priming effect between labile and stable soil organic carbon. The New Phytologist, 2023, 237(1): 88-99. DOI:10.1111/nph.18458 |
| [88] |
Manzoni S, Taylor P, Richter A, Porporato A, Ågren G I. Environmental and stoichiometric controls on microbial carbon-use efficiency in soils. New Phytologist, 2012, 196(1): 79-91. DOI:10.1111/j.1469-8137.2012.04225.x |
| [89] |
Spohn M, Pötsch E M, Eichorst S A, Woebken D, Wanek W, Richter A. Soil microbial carbon use efficiency and biomass turnover in a long-term fertilization experiment in a temperate grassland. Soil Biology and Biochemistry, 2016, 97: 168-175. DOI:10.1016/j.soilbio.2016.03.008 |
| [90] |
Spohn M, Klaus K, Wanek W, Richter A. Microbial carbon use efficiency and biomass turnover times depending on soil depth - Implications for carbon cycling. Soil Biology and Biochemistry, 2016, 96: 74-81. DOI:10.1016/j.soilbio.2016.01.016 |
| [91] |
Sinsabaugh R L, Manzoni S, Moorhead D L, Richter A. Carbon use efficiency of microbial communities: stoichiometry, methodology and modelling. Ecology Letters, 2013, 16(7): 930-939. DOI:10.1111/ele.12113 |
| [92] |
程淑兰, 方华军, 徐梦, 耿静, 何舜, 于光夏, 曹子铖. 氮沉降增加情景下植物-土壤-微生物交互对自然生态系统土壤有机碳的调控研究进展. 生态学报, 2018, 38(23): 8285-8295. |
| [93] |
Lehmann J, Hansel C M, Kaiser C, Kleber M, Maher K, Manzoni S, Nunan N, Reichstein M, Schimel J P, Torn M S, Wieder W R, Kögel-Knabner I. Persistence of soil organic carbon caused by functional complexity. Nature Geoscience, 2020, 13(8): 529-534. DOI:10.1038/s41561-020-0612-3 |
| [94] |
Nave L E, DeLyser K, Domke G M, Holub S M, Janowiak M K, Kittler B, Ontl T A, Sprague E, Sucre E B, Walters B F, Swanston C W. Disturbance and management effects on forest soil organic carbon stocks in the Pacific Northwest. Ecological Applications, 2022, 32(6): e2611. DOI:10.1002/eap.2611 |
| [95] |
de Vries W, Du E Z, Butterbach-Bahl K. Short and long-term impacts of nitrogen deposition on carbon sequestration by forest ecosystems. Current Opinion in Environmental Sustainability, 2014, 9/10: 90-104. DOI:10.1016/j.cosust.2014.09.001 |
| [96] |
VandenEnden L, Anthony M A, Frey S D, Simpson M J. Biogeochemical evolution of soil organic matter composition after a decade of warming and nitrogen addition. Biogeochemistry, 2021, 156(2): 161-175. DOI:10.1007/s10533-021-00837-07||| |
| [97] |
Du Y H, Guo P, Liu J Q, Wang C Y, Yang N, Jiao Z X. Different types of nitrogen deposition show variable effects on the soil carbon cycle process of temperate forests. Global Change Biology, 2014, 20(10): 3222-3228. DOI:10.1111/gcb.12555 |
| [98] |
Liu J, Wu N N, Wang H, Sun J F, Peng B, Jiang P, Bai E. Nitrogen addition affects chemical compositions of plant tissues, litter and soil organic matter. Ecology, 2016, 97(7): 1796-1806. DOI:10.1890/15-1683.1 |
| [99] |
郭洁芸, 王雅歆, 李建龙. 氮添加对中国陆地生态系统植物-土壤碳动态的影响. 生态学报, 2022, 42(12): 4823-4833. |
| [100] |
Bradford M A, Wieder W R, Bonan G B, Fierer N, Raymond P A, Crowther T W. Managing uncertainty in soil carbon feedbacks to climate change. Nature Climate Change, 2016, 6(8): 751-758. DOI:10.1038/nclimate3071 |
| [101] |
Li J Q, Zhu T, Singh B K, Pendall E, Li B, Fang C M, Nie M. Key microorganisms mediate soil carbon-climate feedbacks in forest ecosystems. Science Bulletin, 2021, 66(19): 2036-2044. DOI:10.1016/j.scib.2021.03.008 |
| [102] |
Schimel J P, Schaeffer S M. Microbial control over carbon cycling in soil. Frontiers in Microbiology, 2012, 3: 348. |
| [103] |
Buckeridge K M, Mason K E, McNamara N P, Ostle N, Puissant J, Goodall T, Griffiths R I, Stott A W, Whitaker J. Environmental and microbial controls on microbial necromass recycling, an important precursor for soil carbon stabilization. Communications Earth & Environment, 2020, 1(1): 36. |
| [104] |
Monreal C M, Sultan Y, Schnitzer M. Soil organic matter in nano-scale structures of a cultivated Black Chernozem. Geoderma, 2010, 159(1/2): 237-242. |
| [105] |
Xiao K Q, Zhao Y, Liang C, Zhao M Y, Moore O W, Otero-Fariña A, Zhu Y G, Johnson K, Peacock C L. Introducing the soil mineral carbon pump. Nature Reviews Earth & Environment, 2023, 4(3): 135-136. |
| [106] |
Huang B B, Yan G Y, Liu G C, Sun X Y, Wang X C, Xing Y J, Wang Q G. Effects of long-term nitrogen addition and precipitation reduction on glomalin-related soil protein and soil aggregate stability in a temperate forest. CATENA, 2022, 214: 106284. DOI:10.1016/j.catena.2022.106284 |
| [107] |
Lu M, Zhou X H, Luo Y Q, Yang Y H, Fang C M, Chen J K, Li B. Minor stimulation of soil carbon storage by nitrogen addition: A meta-analysis. Agriculture, Ecosystems & Environment, 2011, 140(1/2): 234-244. |
| [108] |
Johnson D W, Curtis P S. Effects of forest management on soil C and N storage: meta analysis. Forest Ecology and Management, 2001, 140(2/3): 227-238. |
| [109] |
Nave L E, Vance E D, Swanston C W, Curtis P S. Impacts of elevated N inputs on north temperate forest soil C storage, C/N, and net N-mineralization. Geoderma, 2009, 153(1/2): 231-240. |
| [110] |
Cusack D F, Silver W L, Torn M S, McDowell W H. Effects of nitrogen additions on above- and belowground carbon dynamics in two tropical forests. Biogeochemistry, 2011, 104(1/3): 203-225. |
| [111] |
Ma S H, Chen G P, Tian D, Du E Z, Xiao W, Jiang L, Zhou Z, Zhu J L, He H B, Zhu B, Fang J Y. Effects of seven-year nitrogen and phosphorus additions on soil microbial community structures and residues in a tropical forest in Hainan Island, China. Geoderma, 2020, 361: 114034. DOI:10.1016/j.geoderma.2019.114034 |
| [112] |
储炳银, 李培玺, 孙孟瑶, 王晶晶, 滕臻, 徐小牛. 氮磷添加对亚热带常绿阔叶林土壤碳氮及其稳定性同位素的影响. 中南林业科技大学学报, 2021, 41(8): 100-107. |
| [113] |
Wang C, Lu X K, Mori T, Mao Q G, Zhou K J, Zhou G Y, Nie Y X, Mo J M. Responses of soil microbial community to continuous experimental nitrogen additions for 13 years in a nitrogen-rich tropical forest. Soil Biology and Biochemistry, 2018, 121: 103-112. DOI:10.1016/j.soilbio.2018.03.009 |
| [114] |
潘禹, 宋娅丽, 王克勤, 张雨鉴, 郑兴蕊. 模拟N沉降对滇中亚高山典型森林凋落物分解及土壤微生物的影响. 林业科学研究, 2021, 34(3): 88-97. |
| [115] |
曹子铖, 程淑兰, 方华军, 徐梦, 耿静, 卢明珠, 杨艳, 李玉娜. 温带针阔叶林土壤有机碳动态和微生物群落结构对有机氮添加的响应特征. 土壤学报, 2020, 57(4): 963-974. |
| [116] |
Forstner S J, Wechselberger V, Müller S, Keibinger K M, Díaz-Pinés E, Wanek W, Scheppi P, Hagedorn F, Gundersen P, Tatzber M, Gerzabek M H, Zechmeister-Boltenstern S. Vertical redistribution of soil organic carbon pools after twenty years of nitrogen addition in two temperate coniferous forests. Ecosystems, 2019, 22(2): 379-400. DOI:10.1007/s10021-018-0275-8 |
| [117] |
Frey S D, Ollinger S, Nadelhoffer K, Bowden R, Brzostek E, Burton A, Caldwell B A, Crow S, Goodale C L, Grandy A S, Finzi A, Kramer M G, Lajtha K, LeMoine J, Martin M, McDowell W H, Minocha R, Sadowsky J J, Templer P H, Wickings K. Chronic nitrogen additions suppress decomposition and sequester soil carbon in temperate forests. Biogeochemistry, 2014, 121(2): 305-316. DOI:10.1007/s10533-014-0004-0 |
| [118] |
Chang R Y, Zhou W J, Fang Y T, Bing H J, Sun X Y, Wang G X. Anthropogenic nitrogen deposition increases soil carbon by enhancing new carbon of the soil aggregate formation. Journal of Geophysical Research: Biogeosciences, 2019, 124(3): 572-584. DOI:10.1029/2018JG004877 |
| [119] |
Schlesinger W H, Andrews J A. Soil respiration and the global carbon cycle. Biogeochemistry, 2000, 48(1): 7-20. DOI:10.1023/A:1006247623877 |
| [120] |
Metcalfe D B, Fisher R A, Wardle D A. Plant communities as drivers of soil respiration: pathways, mechanisms, and significance for global change. Biogeosciences, 2011, 8(8): 2047-2061. DOI:10.5194/bg-8-2047-2011 |
| [121] |
Hursh A, Ballantyne A, Cooper L, Maneta M, Kimball J, Watts J. The sensitivity of soil respiration to soil temperature, moisture, and carbon supply at the global scale. Global Change Biology, 2017, 23(5): 2090-2103. DOI:10.1111/gcb.13489 |
| [122] |
Fenn K M, Malhi Y, Morecroft M D. Soil CO2 efflux in a temperate deciduous forest: environmental drivers and component contributions. Soil Biology and Biochemistry, 2010, 42(10): 1685-1693. DOI:10.1016/j.soilbio.2010.05.028 |
| [123] |
Wang C, Liu D W, Bai E. Decreasing soil microbial diversity is associated with decreasing microbial biomass under nitrogen addition. Soil Biology and Biochemistry, 2018, 120: 126-133. DOI:10.1016/j.soilbio.2018.02.003 |
| [124] |
Peng Y, Chen G T, Li S, Hu H L, Hu T X, Liu L, Tang Y, Tu L H. Nitrogen additions reduce rhizospheric and heterotrophic respiration in a subtropical evergreen broad-leaved forest. Plant and Soil, 2018, 431(1/2): 449-463. |
| [125] |
An H, Wu X Z, Zhang Y R, Tang Z S. Effects of land-use change on soil inorganic carbon: A meta-analysis. Geoderma, 2019, 353: 273-282. DOI:10.1016/j.geoderma.2019.07.008 |
| [126] |
Liao C, Li D, Huang L, Yue P Y, Liu F, Tian Q X. Higher carbon sequestration potential and stability for deep soil compared to surface soil regardless of nitrogen addition in a subtropical forest. PeerJ, 2020, 8: e9128. DOI:10.7717/peerj.9128 |
| [127] |
Gilliam F S, Burns D A, Driscoll C T, Frey S D, Lovett G M, Watmough S A. Decreased atmospheric nitrogen deposition in eastern North America: Predicted responses of forest ecosystems. Environmental Pollution, 2019, 244: 560-574. DOI:10.1016/j.envpol.2018.09.135 |
| [128] |
Wang C M, Yang X T, Xu K. Effect of chronic nitrogen fertilization on soil CO2 flux in a temperate forest in North China: a 5-year nitrogen addition experiment. Journal of Soils and Sediments, 2018, 18(2): 506-516. DOI:10.1007/s11368-017-1760-y |
| [129] |
王泽西, 陈倩妹, 黄尤优, 邓慧妮, 谌贤, 唐实玉, 张健, 刘洋. 川西亚高山森林土壤呼吸和微生物生物量碳氮对施氮的响应. 生态学报, 2019, 39(19): 7197-7207. |
| [130] |
Coucheney E, Strömgren M, Lerch T Z, Herrmann A M. Long-term fertilization of a boreal N orway spruce forest increases the temperature sensitivity of soil organic carbon mineralization. Ecology and Evolution, 2013, 3(16): 5177-5188. DOI:10.1002/ece3.895 |
| [131] |
Sun S Q, Wu Y H, Zhang J, Wang G X, DeLuca T H, Zhu W Z, Li A D, Duan M, He L. Soil warming and nitrogen deposition alter soil respiration, microbial community structure and organic carbon composition in a coniferous forest on eastern Tibetan Plateau. Geoderma, 2019, 353: 283-292. DOI:10.1016/j.geoderma.2019.07.023 |
| [132] |
李睿达, 张凯, 苏丹, 逯非, 万五星, 王效科, 郑华. 施氮对桉树人工林生长季土壤温室气体通量的影响. 生态学报, 2015, 35(18): 5931-5939. |
| [133] |
Geng J, Cheng S L, Fang H J, Yu G R, Li X Y, Si G Y, He S, Yu G X. Soil nitrate accumulation explains the nonlinear responses of soil CO2 and CH4 fluxes to nitrogen addition in a temperate needle-broadleaved mixed forest. Ecological Indicators, 2017, 79: 28-36. DOI:10.1016/j.ecolind.2017.03.054 |
| [134] |
Li D D, Liu Q, Yin H J, Luo Y Q, Hui D F. Differential responses and controls of soil CO2 and N2O fluxes to experimental warming and nitrogen fertilization in a subalpine coniferous spruce (Picea asperata Mast.) plantation forest. Forests, 2019, 10(9): 808. DOI:10.3390/f10090808 |
| [135] |
Liang L L, Eberwein J R, Allsman L A, Grantz D A, Jenerette G D. Regulation of CO2 and N2O fluxes by coupled carbon and nitrogen availability. Environmental Research Letters, 2015, 10(3): 034008. DOI:10.1088/1748-9326/10/3/034008 |
| [136] |
Xu C H, Xu X, Ju C H, Chen H Y H, Wilsey B J, Luo Y Q, Fan W. Long-term, amplified responses of soil organic carbon to nitrogen addition worldwide. Global Change Biology, 2021, 27(6): 1170-1180. DOI:10.1111/gcb.15489 |
| [137] |
Liao C, Tian Q X, Liu F. Nitrogen availability regulates deep soil priming effect by changing microbial metabolic efficiency in a subtropical forest. Journal of Forestry Research, 2021, 32(2): 713-723. DOI:10.1007/s11676-020-01148-0 |
| [138] |
Wang Q K, Wang S L, He T X, Liu L, Wu J B. Response of organic carbon mineralization and microbial community to leaf litter and nutrient additions in subtropical forest soils. Soil Biology and Biochemistry, 2014, 71: 13-20. DOI:10.1016/j.soilbio.2014.01.004 |
| [139] |
Lu X K, Mao Q G, Wang Z H, Mori T, Mo J M, Su F L, Pang Z Q. Long-term nitrogen addition decreases soil carbon mineralization in an N-rich primary tropical forest. Forests, 2021, 12(6): 734. DOI:10.3390/f12060734 |
| [140] |
Zhang H, Liu Y H, Zhou Z Y, Zhang Y Y. Inorganic nitrogen addition affects soil respiration and belowground organic carbon fraction for a Pinus tabuliformis forest. Forests, 2019, 10(5): 369. DOI:10.3390/f10050369 |
| [141] |
Micks P, Aber J D, Boone R D, Davidson E A. Short-term soil respiration and nitrogen immobilization response to nitrogen applications in control and nitrogen-enriched temperate forests. Forest Ecology and Management, 2004, 196(1): 57-70. DOI:10.1016/j.foreco.2004.03.012 |
| [142] |
Chen F, Yan G Y, Xing Y J, Zhang J H, Wang Q G, Wang H L, Huang B B, Hong Z M, Dai G H, Zheng X B, Liu T. Effects of N addition and precipitation reduction on soil respiration and its components in a temperate forest. Agricultural and Forest Meteorology, 2019, 271: 336-345. DOI:10.1016/j.agrformet.2019.03.021 |
| [143] |
Nottingham A T, Turner B L, Stott A W, Tanner E V J. Nitrogen and phosphorus constrain labile and stable carbon turnover in lowland tropical forest soils. Soil Biology and Biochemistry, 2015, 80: 26-33. DOI:10.1016/j.soilbio.2014.09.012 |
| [144] |
Hudson R J M, Gherini S A, Goldstein R A. Modeling the Global Carbon Cycle: nitrogen fertilization of the terrestrial biosphere and the "missing" CO2 sink. Global Biogeochemical Cycles, 1994, 8(3): 307-333. DOI:10.1029/94GB01044 |
| [145] |
Forsmark B, Nordin A, Maaroufi N I, Lundmark T, Gundale M J. Low and high nitrogen deposition rates in northern coniferous forests have different impacts on aboveground litter production, soil respiration, and soil carbon stocks. Ecosystems, 2020, 23(7): 1423-1436. DOI:10.1007/s10021-020-00478-8 |
| [146] |
张雪, 梅莉, 宋利豪, 刘力诚, 赵泽尧. 模拟氮沉降对马尾松土壤微生物群落结构及温室气体释放的影响. 生态学报, 2019, 39(6): 1917-1925. |
| [147] |
Chen X M, Liu J X, Deng Q, Yan J H, Zhang D Q. Effects of elevated CO2 and nitrogen addition on soil organic carbon fractions in a subtropical forest. Plant and Soil, 2012, 357(1/2): 25-34. |
| [148] |
Sun L Y, Li L, Chen Z Z, Wang J Y, Xiong Z Q. Combined effects of nitrogen deposition and biochar application on emissions of N2O, CO2 and NH3 from agricultural and forest soils. Soil Science and Plant Nutrition, 2014, 60(2): 254-265. DOI:10.1080/00380768.2014.885386 |
| [149] |
Maestrini B, Abiven S, Singh N, Bird J, Torn M S, Schmidt M W I. Carbon losses from pyrolysed and original wood in a forest soil under natural and increased N deposition. Biogeosciences, 2014, 11(18): 5199-5213. DOI:10.5194/bg-11-5199-2014 |
| [150] |
Allison S D, Czimczik C I, Treseder K K. Microbial activity and soil respiration under nitrogen addition in Alaskan boreal forest. Global Change Biology, 2008, 14(5): 1156-1168. DOI:10.1111/j.1365-2486.2008.01549.x |
| [151] |
Fernández-Alonso M J, Díaz-Pinés E, Rubio A. Drivers of soil respiration in response to nitrogen addition in a Mediterranean mountain forest. Biogeochemistry, 2021, 155(3): 305-321. DOI:10.1007/s10533-021-00827-2 |
| [152] |
Mori T, Imai N, Yokoyama D, Mukai M, Kitayama K. Effects of selective logging and application of phosphorus and nitrogen on fluxes of CO2, CH4 and N2O in lowland tropical rainforests of Borneo. Journal of Tropical Forest Science, 2017, 29(2): 248-256. |
| [153] |
Koehler B, Corre M D, Steger K, Well R, Zehe E, Sueta J P, Veldkamp E. An in-depth look into a tropical lowland forest soil: nitrogen-addition effects on the contents of N2O, CO2 and CH4 and N2O isotopic signatures down to 2-m depth. Biogeochemistry, 2012, 111(1): 695-713. |
| [154] |
Gao W L, Yang H, Li S G, Kou L A. Responses of soil CO2, CH4 and N2O fluxes to N, P, and acid additions in mixed forest in subtropical China. Journal of Resources and Ecology, 2017, 8(2): 154-164. DOI:10.5814/j.issn.1674-764X.2017.02.006 |