 2024, Vol. 44
2024, Vol. 44文章信息
- 李愈哲, 黄麟, 樊江文
- LI Yuzhe, HUANG Lin, FAN Jiangwen
- 灌丛资源及其管理利用研究进展
- Research progress of shrub resources and their management and utilization
- 生态学报. 2024, 44(6): 2646-2659
- Acta Ecologica Sinica. 2024, 44(6): 2646-2659
- http://dx.doi.org/10.20103/j.stxb.202304240858
-
文章历史
- 收稿日期: 2023-04-24
- 网络出版日期: 2023-10-31

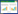




灌丛是重要的植被类型之一, 主要由丛生、没有明显主干且高度不超过5 m的木本高位芽植物所构成[1], 主要分布在气候条件干燥或寒冷、森林不易生长的环境中。在森林或草地自然分布的区域, 受人为或自然因素干扰, 也可能形成次生灌丛。灌丛与森林、草地的关联紧密, 但其特征与森林、草地各有异同。灌丛、灌木、灌木地以及在其中生存的野生动物、植物、微生物的有机整体构成了灌丛资源。灌木提供多种多样的效用, 是人类可利用的重要自然资源, 能作为饲料、薪材、木材、药材等, 发展畜牧业, 提供林副产品, 从而产生可观经济价值。灌丛发达的根系、坚强的生命力, 可以发挥防沙固土、涵养水源、水土保持等生态防护价值[2—3]。此外, 部分灌木物种作为园林美学观赏植物, 还可提供旅游观光、风景游憩等社会文化价值。
由于大部分灌丛植被长期以灌木林的定位纳入森林管理, 目前国内针对灌丛的研究文献较森林、草地少, 多数研究将其作为森林的一类进行简述。并且针对灌木、灌丛的概念释义不清晰, 区分标准存在较多争议[4]。近年来, 在气候和人类活动的驱动下, 灌丛植被的分布范围、覆盖面积快速变化[5], 同时灌丛物种和植被在生态建设恢复中获得更广泛应用[6], 部分灌丛植物经济价值逐渐得到深度开发[7], 对灌丛及其资源价值的认识得以不断演进发展。以灌丛、灌木、灌木林、灌木资源以及若干重要灌木物种的种、属名作为检索关键词, 通过CiteSpace软件统计分析2007—2023年发表在CNKI北大核心的1997篇文献。领域关键词知识图谱共有111个节点, 469条连线, 节点密度0.0768。其中“灌丛”出现的频次最高为100次, 其次是“植被类型”和“土壤养分”(图 1), 分别为79和77次。研究机构共现可视化的117个节点机构, 按出现次数由高到低排序, 中国科学院大学出现次数最高为180次, 是该领域发文量最多的机构, 中国科学院西北高原生物研究所、北京师范大学资源学院、内蒙古农业大学沙漠治理学院等院校机构均有较高出现次数。在Web of science核心合集数据库的5459篇有效文献中, “vegetation”是最高的共现关键词, 出现频次485次。其次是“conservation”和“diversity”, 分别为466和442次。研究机构出现次数由高到低排序, Chinese Academy of Sciences出现次数394次, 是该领域发文量最多的机构, 其次是United States Department of Agriculture (USDA)和University of California System, 分别为266次和179次。

|
| 图 1 2007—2023年关键词共现和研究机构合作可视化知识图谱 Fig. 1 Visualization knowledge map of key words co-occurrence and cooperation between research institutes from 2007—2023 |
研究领域内积极的发展变化促使对灌丛及灌丛资源的认知、管理产生新的更高要求。在这一背景下, 如何准确识别评估我国灌丛资源总量、状态及其变化趋势, 通过发展应用高效精准的灌丛资源管理利用体系, 发挥、维护其生态、生产价值是领域内亟待解决的科学问题。本研究剖析了灌丛相关术语, 介绍了我国灌丛资源分布概况、价值构成, 旨在通过剖析灌丛和灌丛资源研究历史, 在分析凝练当前灌丛管理利用领域面临问题矛盾的基础上, 对未来灌丛资源研究应着力方向开展探讨展望。
1 灌丛相关定义 1.1 灌木灌木类的木本植物分布广泛, 受不同的自然环境条件、经营管理方式以及不同文化认知影响, 在不同的分类体系和定义中, 对灌木的高度存在较多定义差异和分歧(表 1)。通常以有无明显主干及高度来区分乔木和灌木[5]。但在不同学科及同类学科定义中, 有将灌木高度限定为3 m的, 也有限定为5 m的[4, 8, 12], 这是因为我国灌木物种的一般高度存在明显的地域差异, 如青藏高原高寒高海拔区域分布的金露梅高度多在1 m以下[13—14], 而华南的一些次生性灌木树种如构树、桑树、山矾则可以长到5 m或更高[15], 甚至在一定条件下, 也可形态上生长为小乔木或乔木状植株。
| 术语 Term |
定义 Definition |
资料来源 Source |
学科范围 Discipline |
| 灌木 Shrub | 多年生木本植物, 通常无明显主干, 分枝从近地面处开始, 无树冠和枝下高区分, 高一般在3 m以下。 | 国有林区营造林检查验收规则LY/T 1571-2000 | 森林培育学 |
| 无明显主干, 分枝靠近地面, 无树冠和枝下高之分, 成熟植株高在3 m以下的多年生木本植物。 | 林学名词(2016年第二版) | 森林植物学 | |
| 没有明显主干或者无主干、呈丛生状态的矮木本植物。在园林植物中可分为观花、观果、观枝干等几类。常见灌木有玫瑰、杜鹃、牡丹、女贞、小檗、黄杨、沙地柏、铺地柏、连翘、迎春、月季等。 | 草业大辞典 | 草原学 | |
| 灌木群落 Shrub community |
包括一切由无主干、从地面开始分枝的簇生木本植物为主的植物群落, 高度一般在5 m以下, 在世界上分布极其广泛。它有常绿的, 也有落叶的;有针叶的, 也有阔叶和硬叶的, 类型极其多样。 | 草业大辞典 | 草原学 |
| 灌丛 Shrubland | 灌丛包括一切以灌木占优势所组成的植被类型。群落高度一般在5米以下, 盖度大于30%—40%, 更主要是灌丛建群种多为簇生的灌木生活型, 包括原生性灌丛和人为或其他因素影响较长期存在的相对稳定的次生植被。 | 中国植被[8] | 植物生态学 |
| 以灌木占优势的一种植被类型。其植株无明显主干, 建群种多干簇生, 高度在5 m以下, 盖度大于30%—40%, 具有比较郁闭的植被层, 多为中旱生植物。对高大灌木组成的灌丛, 也有称之为“灌木林”的。灌丛的生态适应性很广, 在干燥和寒冷气候条件下, 一般乔木树种难以生存的地方仍能生长。 | 中国农业百科全书:林业卷(上) | 农学 | |
| 小的或矮化的灌木或树木占优势的植被。一般多发育在比乔木群落环境条件差的地方。鲁贝尔[9]把世界灌木群落与乔木群落对应, 分为以下6大类:(1)热带常雨灌木群落。(2)阔叶常绿灌木群落。(3)硬叶灌木群落。(4)夏绿灌木群落。(5)雨绿灌木群落。(6)针叶灌木群落。 | 英汉生态学词典 | 森林生态学 | |
| 以中生性灌木占优势所组成的植被类型。群落高度一般均在5 m以下, 盖度大于30%—40%, 常分布于气候过于干燥或寒冷, 森林难以形成的地方。 | 常用林业名词解释[10] | 森林生态学 | |
| 以中生性灌木为主, 生存比较持久稳定, 高度在5 m以下, 覆盖度大于30%的矮小丛生的木本植物群落。 | 中国资源科学百科全书[11] | 植物资源学 |
限定高度的优势是易于做出判断, 但劣势是不同自然条件下和处于不同生长周期的植被生长状态存在差别, 易出现误判。此外, 还有的定义选择罗列出常见的灌木物种来定义灌木, 但是在物种识别方面过于繁杂, 罗列难以穷举且仍然存在区域差异。通过区域性的研究调查, 形成基于量化的高度指标并考虑当地灌木植物物种点的区域性灌木物种名录可能将更好的解决相关争议和问题。
1.2 灌丛与灌丛资源灌丛建群种多为中生、簇生的灌丛生活型的灌木物种, 灌丛是陆地生态系统一个重要的组成部分。在实际应用中, 为与森林相区分, 同时方便大规模作业, 往往以植被冠层高度来区分灌丛和森林。相关研究中, 灌丛也多以高度限定5 m, 郁闭度0.3—0.4为认定标准[16—17]。不同定义来源均强调灌丛的较强适应性, 能分布在乔木难以分布的地区。
当前从资源角度考虑, 尚未有清晰的存在高度共识的“灌丛资源”概念[18]。参考森林资源的概念[19]“森林资源是森林、林木、林地以及依托森林、林木、林地生存的野生动物、植物和微生物等的总称”。认为灌丛资源的概念可相应定义为灌丛、散生灌木、灌丛地以及依托灌丛、散生灌木、灌丛地生存的野生动物、植物和微生物等的总称。本文讨论的灌丛资源依据这一定义, 将涵盖符合条件的自然植被中的灌丛资源和人工建设形成的灌丛资源。
2 灌丛分布与面积根据中华人民共和国植被图(1 : 100万)数据[20—21], 我国灌丛面积约为9060万hm2(图 2), 主要分布在我国西南、西北和华北部分地区。其中, 西藏、四川、云南3省灌丛面积最大, 达800万hm2以上, 而天津、江苏和山东灌丛面积最小, 均在8万hm2以下(图 2)。

|
| 图 2 中国灌丛植被分布和国各省灌丛面积 Fig. 2 Distribution of shrubland and provincial shrubland area of China 图中所展示数据来源于中华人民共和国植被图(1 : 100万), 数据来源网址:https://www.plantplus.cn/doi/10.12282/plantdata.0155 |
灌丛在我国的生态适应范围极广, 是我国山地和干旱地区极为常见的植被类型, 在青藏高原和云贵高原的山地垂直带上分布有原生灌丛, 而在我国各地分布的次生性灌丛类型具有明显的地区性和生物生态学差异[2, 11, 22]。东北地区山地次生灌丛, 常由胡枝子属、榛属、柳属等灌木植物组成;华北、西北半干旱地区中山带森林破坏后, 常形成由锦鸡儿、荆条、山杏、山桃等灌木形成次生灌丛;西南地区高山、亚高山及中山带的阳坡常由硬叶栎类形成次生灌丛;西北地区常分布沙棘、柽柳、杠柳为优势种的灌丛;亚热带山地丘陵区灌丛物种更为多样, 多种常绿栎类、柃属、冬青、檵木、桃金娘等作为优势种能够形成常绿灌丛;热带地区有以露兜树、酒饼簕、箣竹、仙人掌等树种组成的不同灌丛[23]。
虽然灌丛总面积在全国生态系统类型中占比不到10%, 但在我国是仅次于森林的陆地生态系统土壤保持和碳固定主体[24], 在提供调节服务功能中占有重要地位。随着原生植被的退化以及退化植被的恢复及重建, 中国灌丛植被的分布面积增长迅速[22], 深入研究掌握灌丛植被的特征特性, 有效发掘利用其资源属性, 优化其管理利用方式应引起重视。
3 我国灌丛资源的生态、经济和社会价值在特定的区域内, 灌丛有相当多的实用价值[11, 16], 灵活运用灌丛资源的保护或采伐机制, 以达到生态和经济效益双赢效果非常重要。本文将灌丛资源的常见利用价值和利用方式分别归纳为生态价值、经济植物价值、饲用价值和风景游憩价值。
3.1 生态价值在生态环境保护方面, 灌丛植被具备根系发达、生命力顽强, 其在固碳释氧、防风固沙、涵养水源[16]、水土保持、稳定生态系统方面发挥着重大作用(表 2)。2015年我国灌丛生态系统价值量总量达到22388.15亿元[24]。但不同地区的灌丛发挥的主要生态效益各有特点。
| 价值类型Value type 生态价值Ecological value |
空间范围 Spatial scope |
物质量 Material quantity |
价值量/亿元 Value quantity |
| 防风固沙Windbreak and sand fixation | 全国 | 2.57亿t | 385.53 |
| 物种保育Species conservation | 内蒙古 | — | 261.93 |
| 土壤保育Soil conservation | 全国 | 70.38亿t | 886.75 |
| 水源涵养Water conservation | 全国 | 118.93亿m3 | 839.74 |
| 气候变化减缓Climate change mitigation | 内蒙古 | 23145135.73 t (CO2当量) | 755.79 |
| 微气候调节Microclimate regulation | 全国 | 降温量3389.70 kJ;增湿量55.4万t | 296.56 |
生长在干旱、半干旱地区, 用于保护和改善荒漠化的树种被称作“沙生灌木”, 是沙地造林的重要原生物种, 也是发挥生态效益的重要灌丛资源。在西北地区石羊河下游的民勤绿洲形成的白刺灌丛沙堆, 在减少风沙危害、遏制流沙前移、稳定绿洲环境等方面起重要生态作用[25]。河西走廊的多枝柽柳灌丛、白刺灌丛、油蒿半灌丛[3]和华北地区浑善达克沙地的黄柳灌丛、西伯利亚杏灌丛、小叶锦鸡儿灌丛等也同样能发挥非常重要的防风固沙作用[26]。
为防治沙漠化和改良土壤, 以往常会利用灌丛防风固沙的生态特性, 通过建植人工灌丛来改善目标区域生态环境[27]。据统计, 经过建植灌丛, 我国2009年至2014年期间, 荒漠化土地净减少了12120 km2, 沙化土地面积减少了9902 km2, 沙化逆转速度变快, 荒漠和沙地持续缩减、生态功能增强, 治理成效显著[28]。其中, 柠条和油蒿具有耐旱、生命力强等特性, 是荒漠地区最常应用的人工灌丛, 在我国北方发挥重要生态功能[29—30]。
分布在青藏高原高寒区域的金露梅灌丛草甸, 发挥水源涵养、水热平衡及生态屏障的功能[13]。西南地区的滇石栎灌丛[31]、华北地区山西省的皂柳群落、野皂角群落等[32]、西北地区的温性及高寒灌丛都产生了涵养水源、水土保持等诸多重要生态价值。
3.2 经济植物价值部分灌丛及其衍生产品, 可药用、食用、提取芳香油、观赏和环境绿化、美化(表 3)。常见药用灌木如山杏, 其种仁可供药用并可榨油, 在我国东北和华北地区大量生产种仁, 供内销和出口[33], 常用灌木来源药材还有有小檗、狼牙刺、小柱花等。杜鹃类则常应用于提取芳香油[11]。常见观赏型灌木有连翘、忍冬、黄杨、月季、杜鹃、牡丹、茉莉等。随着生态园林模式得到社会广泛认可, 灌木在园林景观营造中的作用不可忽视。园林植物群落中的灌木发挥着乔木与地面、建筑物与地面之间的连贯和过渡作用。灌木的高度是人在园林中最方便接触、观赏到的植物类群, 并且灌木的物候和季相变化明显, 能使人们在时间上形成韵律和节奏感[34]。
| 价值类型Value type | 举例Example | |
| 经济价值Economic value | 薪炭 | 重要的生产热能燃料灌丛植物, 如沙棘、小叶锦鸡儿、紫穗槐、柽柳、梭梭等。 |
| 水果、干果 | 以果品为主要产出的灌丛植物, 如沙棘、悬钩子属、茶藨子属、越橘属、刺梨、欧李、扁桃、榛子等 | |
| 工业原料、油料和药材 | 以工业原料、油料和药材为主要产出的灌丛植物, 栀子、茶、油茶、文冠果、枸杞、麻黄连翘等 | |
| 其它灌丛经济产品 | 沤肥、沼气生产等其它经济价值 | |
| 社会价值Social value | 风景游憩价值 | 香山公园黄栌红叶、贵州百里杜鹃风景名胜区百里杜鹃、西南高山杜鹃景区、云南古茶园等 |
在木材方面因为灌丛植被植株较矮小, 通常作为薪材被采伐, 尤其是在交通不便、煤火资源不足或缺乏的山区, 人们生活用柴主要来源于灌丛。木材方面的作用没有森林强大。但在特定区域, 灌丛资源能同时发挥畜牧饲用和木材的作用。在浑善达克沙地, 西伯利亚杏、黄柳与地下芦苇、赖草等禾草植物结合, 可同时获得木材和牧草[26]。
3.3 饲用价值在放牧方面, 灌丛的效益比草地小, 对于可放牧利用的灌丛资源, 多数研究常常称其为“灌丛草地”。可供牲畜食用的灌丛种类能发展畜牧业和饲料产业, 合理地开发利用灌丛可以分担草地的放牧压力, 获取畜牧收入和经济收益。
放牧强度、放牧方式能影响灌丛、草地的生态环境状态, 但灌丛和草地受到的影响是不一致的[35]。宋美娟[36]在研究中指出:长期放牧甘肃马鹿可以有效抑制杯腺柳+小叶金露梅+高山绣线菊灌丛扩张, 利于草本植物生长, 但不利于灌丛生长;长期放牧牦牛+藏羊有利于提高高寒灌丛草地水土保持功能, 保护土壤微生物多样性, 有利于杯腺柳+小叶金露梅+高山绣线菊灌丛扩张, 但影响草本植物的生长。周华坤等[14]在青藏高原金露梅高寒灌丛草场中的研究指出, 随着放牧强度增加, 占优势地位的灌木和禾草被典型杂类草替代。过度放牧下, 极易引起原生灌丛逐渐退化为杂类草草地甚至黑土滩-次生裸地[13, 37]。
部分灌丛植物承担了与草地相似的饲草供给功能, 但灌丛与草地的特性不同。对于能有良好牧草地的地区, 要控制好放牧压力, 避免草地最终演变为不适合放牧的稀疏灌丛地[38]。对于环境较恶劣而灌丛占优势的地区, 也需要控制好对灌丛资源的消耗[39—40], 避免饲用价值的退化、衰竭。总之, 合适的放牧强度有利于保持灌丛植被的生产力和饲用价值。
3.4 风景游憩价值灌丛基于其植被特征和构成物种的独特审美属性[41], 形成了诸多富有特色的风景游憩资源。如贵州省西北部的百里杜鹃风景名胜区, 以大面积原生杜鹃林为主要特色, 杜鹃林带宽1—3 km, 绵延50余千米已形成国家5A级旅游景区。此外, 部分色叶灌木物种如黄栌、观花灌木物种如高山杜鹃、部分具有文化传承价值的灌木如古茶等在不同地区形成诸多灌丛风景区, 大量的灌木、灌丛风景游憩资源仍有待开发利用。
4 灌丛资源与森林、草地资源关系 4.1 灌丛资源与森林资源当前森林资源管理实践中, 将灌木视为林木的一类, 因此大量灌丛资源依照目前分类体系, 属于森林资源的范畴[19]。若将森林资源中的林木限制为森林植被中最为主要的乔木, 把灌木剥离出来归入灌丛资源, 则灌丛资源与森林资源将能够更清晰区隔开来。
已有研究表明, 森林的主要功能、价值与灌丛具有高度的重合(表 3), 但一般情况下单位面积灌丛不如森林的产出丰盛[2, 42—43]。同时, 灌木因其分枝多, 以及本身生物学特性决定了灌丛难以发挥用材林的作用, 部分森林功能如材用, 灌丛难以替代。尽管如此, 灌丛亦有森林不具备的一些特殊功用价值, 灌丛在山区土地贫瘠, 不具备成林条件的区域, 能发挥高产薪炭和经济来源的重要作用[44]。此外, 在园林景观营造中, 灌木具有独特价值, 它作为中间层在乔木与地面、建筑物与地面之间发挥连贯和过渡的作用。在生态方面, 灌木容易种植成活, 且具有水土保持、改良土壤等多种生态功能, 因此灌丛在沙化和荒漠化防治方面是重要的植被类型。在生境薄弱的退耕地区, 针对研究区灌木物种特点, 已有研究强调了引种培植灌木物种, 发挥灌丛资源优势的必要性[16]。我国森林管理常涵盖灌丛, 但实际管理实践中, 森林与灌丛、草地管理措施差异很大。生境条件恶劣区域由于自然禀赋的限制, 即使种植乔木树种, 也往往不能发展出可自我维持的稳定森林植被, 种植植物依赖灌溉且易出现成片死亡或呈现为灌木形态的“老头树”。因此要高效可持续地在底子薄禀赋差的生态脆弱区开展生态恢复, 应当基于区域自然禀赋和天然分布的植物群落物种构成, 适当重视灌丛资源的开发应用和灌丛植被的培植恢复。
4.2 灌丛资源与草地资源草地资源指生长多年生草本植物为主的、可供放养或割草饲养牲畜的土地。以往草地资源的阐释中, 以草本植物为主, 灌木覆盖度小于40%的土地在相关标准中被认定为草地[45—46]。由此, 草地资源不可避免的涵盖纳入了部分零散分布的灌丛资源。
草地最主要的利用方式是放牧, 灌丛的利用方式也包含放牧。但因为草地的饲用价值要高于灌丛, 所以一般具备草地资源的地区, 灌丛资源常不作为主要的饲料资源来源。但是在干旱时期, 适口性较好的灌木能分担草本植物的采食压力[47]。所以对于土壤较贫瘠, 容易发生干旱的地区, 灌丛的饲用价值仍十分重要。
草地与灌丛有着密切的联系, 在自然演替过程中它们是相邻的两个阶段。并且在生态恢复的研究和实践中, 草灌结合的治理方式也十分常见。在河西地区干旱、土壤贫瘠多盐的特征下, 生态的恢复与建设常将灌丛与草地植被恢复结合实施[3];在退化沙地, 可以通过结合草地放牧管理和人工灌丛建植恢复健康的草地生态系统[48]。但是两者对气候、地形、海拔、人为因素的响应存在差异。气候因子变化对灌丛结构的影响强于草本层[27], 此外在海拔因子方面, 随着海拔的升高灌木层的盖度随海拔升高而先上升后下降, 而草本层则与灌木层变化趋势相反[49]。所以, 在灌草结合的生态恢复实践中, 还应考虑灌木层、草本层各自的环境因子响应特性。
此外近年来, 草地还存在愈发明显的“灌丛化”现象趋势。草地灌丛化指草地普遍发生灌木或木本植物密度、盖度及生物量增加的现象[50—51]。在畜牧业视角下, 灌丛化往往意味着群落中草本植物总量和比例减少, 适口性高的牧草发生了退化, 不适口且生命力强的灌木开始成为优势植被, 由此饲用价值降低, 由此草地灌丛化被当作过度放牧的标志之一[47]。
5 我国灌丛资源研究中存在的问题 5.1 灌丛及灌丛资源概念、标准缺乏统一共识以往研究中我国灌丛面积的统计结果相差较大(表 4)。胡会峰[52]等由1∶400万的中国植被图估算的我国6种主要灌丛植被总面积约为15462万hm2, 在我国其分布面积仅次于草地, 而高于森林。王兵等[43]由2008年第七次全国森林资源清查结果中估算, 我国灌丛面积达5365万hm2, 占全国森林总面积的21.9%, 占国土面积的5.57%。王庆慧等[2]在其研究中, 根据各省市政府公布统计公报数据统计得出, 2009年我国灌丛面积约为4341万hm2, 2015年我国灌丛面积约为4329万hm2。张蔷等[53]根据2015年中华人民共和国环保部和中国科学院的数据指出, 我国灌丛分布面积6920万hm2, 占国土面积的7.3%。胡雄蛟等[24]采用30 m空间分辨率的光学卫星遥感数据和生态系统特征相结合的分类方法, 将灌丛生态系统分为常绿阔叶灌木林、落叶阔叶灌木林、常绿针叶灌木林和稀疏灌木林4类, 统计得到2015年我国灌丛面积6750万hm2。
| 年份 Year |
面积Area/万hm2 | 估算数据来源 Data source |
年份 Year |
面积Area/万hm2 | 估算数据来源 Data source |
|
| 2006 | 15462 | 1∶400万的中国植被图 | 2015 | 4329 | 各省市政府公布统计公报数据 | |
| 2008 | 5365 | 第七次全国森林资源清查结果 | 2015 | 6920 | 中华人民共和国环保部和中国科学院数据 | |
| 2009 | 4341 | 各省市政府公布统计公报数据 | 2015 | 6750 | 光学卫星遥感数据和生态系统特征相结合 |
通过对比可知, 虽然各研究间灌丛面积差异巨大, 但同一标准下统计的不同年份同类型灌丛面积仍彼此接近, 处在可比区间。在不同统计和研究中, 对于“灌草丛”、“灌木林”、“矮林”等是否归类为灌丛没有一致的观点和处理。例如, 胡会峰[52]等把矮林归类为灌丛资源时, 推算出的全国灌丛面积就比其他研究高出约三倍。此外, 灌丛在不同学科里的定义也存在一些矛盾, 如以往资源学角度强调灌丛不包括森林遭破坏后形成的次生灌丛群落, 需包括受长期人为影响而持久处于灌丛状态的植被。但实际上, 灌丛的扩张一方面是森林或草地遭到不合理利用后, 发生了逆行演替, 退化为次生林、次生灌丛和灌草丛[54]。另一方面也存在人为对退化的草地植被进行干预, 通过封育、人工建植等处理, 促进荒漠生态系统正向演替, 恢复形成的灌丛[48, 51]。部分阶段性的灌丛亦生产大量灌丛的林副产品、提供灌丛生态服务, 基于此灌丛资源应将阶段性灌丛包含其中。此外, 由于大范围的粗略统计和相关标准中, 常以冠层顶部高度来区分林、灌, 常会将成长中的幼龄林误识别为灌丛, 对于有明显主干的矮林, 尽管高度不够但也不应该归类于灌丛, 否则随群落发育生长, 其属性将快速转变。
综上所述, 在灌丛进行定义时, 应该以植被高度和建群物种作为主要的标准。若地区差异过大, 应采取分区域定义的方法, 对不同地区采取针对性的标准, 并在不同学科中统一。
5.2 缺乏细致系统的灌丛资源分类管理制度在生态恢复和保护层面, 我国对天然灌丛的管理延用了部分森林管理方式, 其主要管理方式有:(1)封禁保护、(2)封育保护、(3)抚育改造。封禁保护是使保护区域内的植被在自然状态下自我恢复的有效手段, 适用于地块偏远难以到达的区域。而封育保护则是加入人为干预, 促进植被保护恢复。其中, 围栏封育是退化、沙化植被生态系统恢复最常用的一项措施[51]。抚育改造则用于改善天然和人工灌丛植被, 采用平茬、直播、除草等技术, 优化目标灌丛生产力等状态指标, 满足人类利用需求。
由于气候等自然禀赋和历史经验习惯的差异, 灌丛管理利用在不同地区存在明显差别。在我国西南地区, 喀斯特地貌广泛分布, 地表崎岖、土壤贫瘠, 石漠化严重。西南地区有较多的亚热带/热带灌丛植被[55], 区域内最常见的灌丛管理方式为抚育改造, 以促进灌丛转变为森林。如在滇中地区, 会通过滇石栎灌丛抚育改造的方式加速群落演替, 发挥该灌丛植被的涵养水源和水土保持功能[31]。在西南亚高山地利用直播、栽植技术补充种源, 在已分布茂密灌丛区域, 则采用调控灌丛冠幅和适当去除地被, 促进灌丛演替为森林[56]。此外, 我国西北和华北干旱、半干旱人工灌丛, 通常也采用抚育改造方式进行管理, 常见方式包括补植、平茬、去除林下植被等。该区域灌丛抚育改造的目标是扩大目标灌丛的分布和覆盖面积[57], 具体措施上小叶锦鸡儿、山竹子等豆科植物以直播, 柽柳类灌木以扦插为主, 差不嘎蒿等蒿类则两种方式均有实施。在西北灌丛管理实践中, 人工柠条、四合木、沙柳等老龄灌丛还会通过平茬抚育改造来改善林间生境[58]、提升灌丛资源利用效率[59]和生态防护功能[60]。去除林下植被在灌丛管理中指清理和防除灌丛冠层下的杂草。对于干旱半干旱黄土高原丘陵区域, 平茬和除草被认为是最适合的灌丛管理方式组合, 可有效提高该地区人工柠条灌丛的生态功能[61]。多山偏僻的山地区域, 丰富灌丛资源常被人类进行开垦利用。但无节制的砍伐开垦农田, 不仅造成灌丛资源快速损失, 还可能引发区域生态环境恶化。这些地区除在关键区域(如陡坡灌丛)实施封禁保护措施外, 还需宣传科普灌丛保护重要性, 同时建立完善引导性政策, 调整当地农耕畜牧产业结构, 在生态恢复的同时改善农牧民生活水平[62—63]。
灌丛是一个内部组成复杂且差异巨大的植被类型, 不同自然禀赋下, 干旱荒漠区域灌丛、半干旱半湿润林草过渡带灌丛、南方湿热区灌丛和青藏高原高寒、高山灌丛应制定符合区域实际的差异性管理模式。在不同功能上, 经济生产灌丛管理又区别于生态灌丛, 更强调生产效率和经济效益的最大化, 当前经济灌丛植被常依据其园林观赏、薪材、医用、药用类型等具体资源功用建立各自不同的管理利用体系, 但其经验、习惯做法尚未有成体系的分类归纳。对于灌丛管理, 我国已有大量经验积累和技术总结, 但因灌丛长期附属于森林管理, 尚未有系统性地归纳。灌丛植被的最适管理方式应区别于森林、草地, 反映自身特点。在生态功能导向的灌丛建设中做到“宜林则林、宜草则草、宜灌则灌”。在生产功能导向的灌丛建设管理中, 针对不同区域不同功能的灌丛植被管理亟需发展出一套成体系的贴合区域自然禀赋, 适应生产功能提升为侧重目标的分区分类管理制度。
5.3 草地灌丛化影响灌丛资源的总量、分布和功能状态草原灌丛化是干旱半干旱区草原生态系统中原生灌木/木本植物趋于增加的现象[64—65]。这一现象在全球范围内广泛发生, 快速的灌丛化过程及影响是干旱半干旱区研究领域广受关注的重要生态问题[66]。综合多数研究发现和结论, 灌丛化是多因素共同作用的结果[67—69], 主要驱动因素包括过度放牧、草原火、气候暖湿化、CO2浓度升高等自然和人为因子变化。草原灌丛化过程使地表景观由原本草本植物占优势快速转为灌木植物占优势, 其间原有的草地产出资源和功能类型快速向灌丛转变[70]。发生灌丛化后, 生态系统所产生、提供的生态服务和经济价值的总量和组成都发生极大变化。据研究, 灌丛斑块在草原中有明显的“肥岛”效应[71], 能够富集累积周围环境中的水分、养分, 提升土壤肥力。在更高水分和肥力的支持下, 植被的生产力和碳固定均趋于加强[72]。灌丛化发生后, 随着盖度和反照率等指标的变化, 原有的光照、热量资源分配也会改变[73]。在多样性影响方面, 斑块尺度上的研究与群落尺度上获得的结果常存在差异[74], 但总体上灌木盖度增加通过促进土壤养分聚集, 能增加灌木冠层下的植被、动物、土壤微生物的生物多样性[75]。由于灌丛冠层的庇护, 牲畜难以啃食致密灌丛冠层下的植物[76], 因此灌丛对于生长在其下层的草本植物起到一定庇护作用, 在其冠层下常形成大量小生境[77—78], 从而改善了微气候条件和增强物种多样性和生物量[79]。此外, 灌丛化伴随某些真菌、细菌生物量增加[80], 从而对土壤水分、营养物质循环、土壤表层稳定性等表现出积极作用。
许多研究认为灌丛化是草地退化或者沙漠化的一种表现形式[81], 在干旱区域的研究中, 这一判断源于灌丛化区域土壤沙化的常见情形, 但这很难确定是干旱沙化导致了植被灌丛化, 亦或是灌丛化导致了沙化发生。干旱区的灌丛常缺乏冠层下草本植物, 地表径流和土壤侵蚀增加[82], 由此灌木入侵在部分干旱区研究中被认为是沙漠化的驱动因素[83]。但灌木斑块对生态系统结构和功能的作用会随着灌木密度、盖度的增加而改变。总体上, 更多研究认为相比原有草地, 灌丛化使生态系统结构和功能趋于复杂[84], 有益于提高生态系统稳定性[75]。但灌丛化对植被生态功能的影响是否是积极的, 则可能受到所处气候条件和自然禀赋的极大约束, 特别是取决于区域水热条件能否支持高盖度灌丛长期稳定存在。
6 未来研究展望尽管灌丛资源相比森林和草原更加分散, 欠缺空间上的连续性, 其生态功能和生产价值在不同灌丛类型间变异极大, 获得的科研关注较少。然而, 随着近年来的气候变化和人类活动加剧, 灌丛的分布快速扩展, 灌丛资源量及可利用的灌丛区域快速增加;灌丛在森林不能耐受环境(如高寒、干旱、贫瘠等)中的独特适应性, 使其在生态保护、恢复应用中的特殊价值被逐步认识接纳;部分经济灌丛植物近年来得到广泛应用, 在自然禀赋较差、环境严苛的贫困区域形成了多样的灌丛脱贫产业。这些变化趋势对灌丛资源研究提出了系列迫切需求, 包括如何动态精准评估我国灌丛资源总量、组成和分布如何?如何准确评价不同区域灌丛化的生态、经济综合得失?如何便捷经济的实现灌丛植被变化快速识别?对灌丛资源如何开展科学管理, 高效维持提升其生态、生产价值?综上, 通过对当前灌丛资源研究进展的梳理及存在争议的分析, 我们认为灌丛资源未来研究应聚焦如下几个方面:
6.1 深入厘清草地快速灌丛化的过程、原因和生态影响目前世界范围内关于草地灌丛化的驱动因素、机制仍存较多争议。自然因素中气候暖湿化和暖干化、CO2施肥效应均被认为是不同区域灌丛化的促进因素[85—87]。澳大利亚和北美[88]的灌丛化现象被认为与草原火灾后的灌丛群落替代和耐火的灌木树种有关。在人类活动因素中, 灌丛化主要归咎于过度放牧导致草本物种抑制和耐饲灌木幼苗的更新[89—90]。不同区域的灌丛化的主要驱动因素可能存在很大不同。部分研究指出了灌丛化驱动因素和转变速度可能受当地气候条件调控[73, 91], 但内在机制、规律亟需深入探索厘清。
各灌丛化区域的植被转变过程和灌丛蔓延速度也需要更详细的高时空分辨率监测, 这不仅有助于了解灌丛资源总量和分布的变化, 也将帮助更准确了解灌丛化的原因和调控机制[3]。此外, 尽管较多研究关注灌丛化对畜牧生产的不利影响, 但综合全面评估草地灌丛化的生态影响[42], 特别是评估灌丛化对区域碳收支[92]、养分周转和区域气候反馈的研究还相对匮乏, 灌丛化与区域气候间交互作用关系研究仍未闭环。因此在现有研究基础上, 归纳总结不同区域差异化的自然禀赋条件下, 灌丛化发生的原因、过程差别, 总结其生态影响的变异规律极为必要, 是后续干预调控和科学管理灌丛化过程的基础。
6.2 发展灌丛植被的多基遥感和人工智能识别技术灌丛资源总量及动态的精准掌握依赖于高时空分辨率的灌丛植被监测。相比人工监测, 遥感监测实施效率高, 人力成本低, 在森林、草地等植被监测中已被广泛应用[93—94]。此外, 遥感数据源多样, 除卫星遥感数据如陆地卫星Landsat系列、哨兵Sentinel系列、和国内的高分系列、环境系列、资源系列等数据外, 驻空飞艇、固定翼/旋翼无人机等空基遥感也快速发展, 逐渐应用于植被或其它地物的提取中[95]。近年来, 日光诱导叶绿素荧光(SIF)数据在植被的识别研究中尤受关注[96—97]。这些新技术新方法在灌丛资源的精准识别监测中具有巨大的应用潜力。
此外, 随着人工智能(AI)快速发展, 与遥感大数据和云计算平台结合, 有效地提高了数据挖掘效率[98—100]。在植被识别领域, 已有研究基于无人机遥感图像, 结合野外实地调查样本, 应用贝叶斯、K最邻近、决策树、随机森林等方法对荒漠区灌丛植物特定物种进行了识别[101]。相信随着包括卷积神经网络等深度学习技术的发展和广泛应用, 基于遥感的灌丛植被分布、覆盖度、物种、生物量等特征的识别技术将不断完善, 从而推动促进灌丛资源总量、组成的精准动态监测。
6.3 健全灌丛资源科学系统分类实现分区分类高效管理利用相比其他类型植被资源, 空间上邻近的不同类型间灌丛资源功能定位差异更大, 毗连区域内的灌丛可能分别承担着防风固沙、药材香料原料来源、水果干果生产、游憩景观等完全不同的功能[102]。灌丛资源类型和价值大小的空间变异也非常明显, 如北方灌丛需侧重的防沙固沙功能[103—104]、西南喀斯特区域灌丛承担的固土保肥功能[105]、南方茶和油茶灌丛的经济生产功能[106]等都具有明晰的区域差异。再者同类灌丛在不同气候禀赋下的不同地区, 其本身的发展演替过程、适合的管理利用模式、高效可行的管理技术措施, 存在独特区域特点和丰富的地方经验积累。依据以上特点, 在对灌丛资源的总量、组成及其分布能够实现高效精准监测的基础上, 科学依据灌丛资源的功能定位、所处区域、建群物种生物学特点对灌丛资源进行科学系统分类, 进而系统总结凝练现有管理利用经验, 实现分区分类精准管理。这将极大提升灌丛资源经营管理效率, 从而帮助实现稳固提升灌丛生态服务和生产价值, 推动灌丛资源研究和综合利用。
| [1] |
Sankaran M, Hanan N P, Scholes R J, Ratnam J, Augustine D J, Cade B S, Gignoux J, Higgins S I, Le Roux X, Ludwig F, Ardo J, Banyikwa F, Bronn A, Bucini G, Caylor K K, Coughenour M B, Diouf A, Ekaya W, Feral C J, February E C, Frost P G H, Hiernaux P, Hrabar H, Metzger K L, Prins H H T, Ringrose S, Sea W, Tews J, Worden J, Zambatis N. Determinants of woody cover in African savannas. Nature, 2005, 438(7069): 846-849. DOI:10.1038/nature04070 |
| [2] |
王庆慧, 李婧贤, 彭羽, 蓝文娟. 我国灌丛生态系统服务功能价值评估. 江苏农业科学, 2019, 47(4): 233-237. |
| [3] |
金自学. 河西走廊灌丛植被的生态学研究. 农村生态环境, 2001, 17(2): 17-21. |
| [4] |
Anderson J R. A land use and land cover classification system for use with remote sensor data. US Government Printing Office, 1976.
|
| [5] |
Di Gregorio A. Land cover classification system: classification concepts and user manual: LCCS. Food & Agriculture Org., 2005.
|
| [6] |
Zhao H L, Zhou R L, Su Y Z, Zhang H, Zhao L Y, Drake S. Shrub facilitation of desert land restoration in the Horqin Sand Land of Inner Mongolia. Ecological Engineering, 2007, 31(1): 1-8. DOI:10.1016/j.ecoleng.2007.04.010 |
| [7] |
闫志坚, 杨持, 高天明. 我国西部灌木植被的生态经济价值. 生态经济, 2007, 23(3): 129-130, 138. DOI:10.3969/j.issn.1671-4407.2007.03.030 |
| [8] |
吴征镒主编中国植被编辑委员会编著. 中国植被. 北京: 科学出版社, 1980.
|
| [9] |
Rübel E, Brockmann-Jerosch H. Pflanzengesellschaften der erde. Verlag Uber, Berlin, 1930. |
| [10] |
中国林业科学研究院林业科技信息研究所. 林业专业知识服务系统--林业术语. 2022.
|
| [11] |
《中国资源科学百科全书》编辑委员会. 中国资源科学百科全书. 北京: 中国大百科全书出版社, 2000.
|
| [12] |
王国宏, 郭柯, 谢宗强, 唐志尧, 蒋延玲, 方精云. 《中国植被志》研编规范的若干说明、补充与修订. 植物生态学报, 2022, 46(3): 368-372. |
| [13] |
戴黎聪, 郭小伟, 柯浔, 杜岩功, 张法伟, 李以康, 李茜, 林丽, 曹广民, 朋措吉, 舒锴. 青藏高原金露梅灌丛草甸植物群落对退化演替的响应. 生态环境学报, 2019, 28(4): 732-740. |
| [14] |
周华坤, 赵新全, 汪诗平, 赵亮, 徐世晓. 青藏高原高寒灌丛植被对长期放牧强度试验的响应特征. 西北植物学报, 2008, 28(10): 2080-2093. DOI:10.3321/j.issn:1000-4025.2008.10.025 |
| [15] |
Zhang R F. Cooling effect and control factors of common shrubs on the urban heat island effect in a southern city in China. Scientific Reports, 2020, 10: 17317. DOI:10.1038/s41598-020-74559-y |
| [16] |
李宜翀. 沁河源灌丛植被植物多样性及其分布特征[D]. 临汾: 山西师范大学, 2020.
|
| [17] |
黑龙江农业百科全书编辑委员会. 黑龙江农业百科全书. 北京: 中国大百科全书出版社, 1993.
|
| [18] |
郝庆, 封志明, 赵丹丹, 魏晓. 自然资源治理的若干新问题与研究新趋势. 经济地理, 2019, 39(6): 1-6. |
| [19] |
国家林业局调查规划设计院. 森林资源术语. 中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2011: 36.
|
| [20] |
张新时. 中华人民共和国植被图(1: 1000000). 北京: 地质出版社, 2007.
|
| [21] |
中国科学院中国植被图编辑委员会. 中华人民共和国植被图(1: 1000000). 2021.
|
| [22] |
谢宗强, 唐志尧. 中国灌丛生态系统碳储量的研究. 植物生态学报, 2017, 41(1): 1-4. |
| [23] |
中国农业出版社. 智汇三农--农业专业知识服务平台. 2022.
|
| [24] |
胡雄蛟, 宋昌素, 范馨悦, 肖燚, 徐卫华, 欧阳志云. 中国灌丛生态系统主要调节服务及价值评估. 北京林业大学学报: 社会科学版, 2021, 20(3): 58-64. |
| [25] |
孙涛, 贾志清, 刘虎俊, 钱雅玲. 绿洲荒漠过渡带不同演替阶段白刺灌丛植被和沙堆形态特征相关性分析. 兰州大学学报: 自然科学版, 2020, 56(3): 396-403, 411. |
| [26] |
郭建英, 余伟莅, 胡小龙, 王利兵. 浑善达克沙地灌丛植被的生态学研究. 内蒙古林业科技, 2005, 31(4): 5-8. DOI:10.3969/j.issn.1007-4066.2005.04.002 |
| [27] |
刘任涛. 沙地灌丛的"肥岛"和"虫岛"形成过程、特征及其与生态系统演替的关系. 生态学杂志, 2014, 33(12): 3463-3469. |
| [28] |
王喜明, 薛振华, 王雅梅. 沙生灌木资源利用. 北京: 中国林业出版社, 2020.
|
| [29] |
刘学东, 陈林, 杨新国, 赵伟, 李学斌. 荒漠草原2种柠条(Caragana korshinskii)和油蒿(Artemisia ordosica)灌丛土壤养分"肥岛"效应. 西北林学院学报, 2016, 31(4): 26-32, 92. DOI:10.3969/j.issn.1001-7461.2016.04.05 |
| [30] |
郭志霞, 刘任涛, 冯永宏, 王文帆, 蒋嘉瑜. 不同降水对荒漠灌丛土壤理化性质和地表植被分布的影响. 水土保持通报, 2021, 41(1): 56-65. |
| [31] |
黄永祥, 成克武, 杨文云, 周彬. 滇中地区滇石栎灌丛植被恢复改造技术研究. 安徽农业科学, 2008, 36(35): 15456-15459. |
| [32] |
梁凤玉. 山西省天然灌丛植被群落类型. 山西林业科技, 2006, 35(2): 28-31. |
| [33] |
系统与进化植物学国家重点实验室中国科学院植物研究所. 植物智--中国植物+物种信息系统. 2019.
|
| [34] |
胡绍庆. 灌木与观赏竹. 北京: 中国林业出版社, 2011.
|
| [35] |
Daryanto S, Eldridge D J, Throop H L. Managing semi-arid woodlands for carbon storage: grazing and shrub effects on above- and belowground carbon. Agriculture, Ecosystems & Environment, 2013, 169: 1-11. |
| [36] |
宋美娟. 放牧甘肃马鹿与牦牛藏羊对东祁连山杯腺柳灌丛植被和土壤的影响[D]. 甘肃农业大学, 2021.
|
| [37] |
Noble J C. Delicate and Noxious Scrub: CSIRO Studies on Native Tree and ShrubProliferation in the Semi-Arid Woodlands of Eastern Australia. CSIRO Publishing, 1998.
|
| [38] |
Li S L, Yu F H, Werger M J A, Dong M, Ramula S, Zuidema P A. Understanding the effects of a new grazing policy: the impact of seasonal grazing on shrub demography in the Inner Mongolian steppe. Journal of Applied Ecology, 2013, 50(6): 1377-1386. DOI:10.1111/1365-2664.12159 |
| [39] |
Downing B H. Goat and sheep grazing in shrub-infested semi-arid woodlands of new South Wales. The Rangeland Journal, 1986, 8(2): 140. DOI:10.1071/RJ9860140 |
| [40] |
Fujiki D, Kishimoto Y, Sakata H. Assessing decline in physical structure of deciduous hardwood forest stands under Sika Deer grazing using shrub-layer vegetation cover. Journal of Forest Research, 2010, 15(2): 140-144. DOI:10.1007/s10310-009-0172-2 |
| [41] |
Garkotj S C, Singh S. Biomass, productivity and nutrient cycling in alpine Rhododendron community of Central Himalaya. Oecologia Montana, 1992, 1: 21-32. |
| [42] |
张婕, 刘玉洁, 潘韬, 封志明, 杨艳昭, 葛全胜. 自然资源资产负债表编制中生态损益核算. 自然资源学报, 2020, 35(4): 755-766. |
| [43] |
王兵, 魏江生, 胡文. 中国灌木林-经济林-竹林的生态系统服务功能评估. 生态学报, 2011, 31(7): 1936-1945. |
| [44] |
Sturm M, Racine C, Tape K. Increasing shrub abundance in the Arctic. Nature, 2001, 411(6837): 546-547. DOI:10.1038/35079180 |
| [45] |
中国农业科学院北京畜牧兽医研究所. 草原健康状况评价. 中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2008, 16. |
| [46] |
Xu M R, Zhang J, Li Z H, Mo Y. Attribution analysis and multi-scenario prediction of NDVI drivers in the Xilin Gol grassland, China. Journal of Arid Land, 2022, 14(9): 941-961. DOI:10.1007/s40333-022-0032-x |
| [47] |
闫宝龙, 吕世杰, 王忠武, 韩国栋. 草地灌丛化成因及其对生态系统的影响研究进展. 中国草地学报, 2019, 41(2): 95-101. |
| [48] |
安琪琪. 灌丛化对黄土高原草地植物多样性和生物量的影响[D]. 杨凌: 西北农林科技大学, 2021.
|
| [49] |
柴永福, 许金石, 刘鸿雁, 刘全儒, 郑成洋, 康慕谊, 梁存柱, 王仁卿, 高贤明, 张峰, 石福臣, 刘晓, 岳明. 华北地区主要灌丛群落物种组成及系统发育结构特征. 植物生态学报, 2019, 43(9): 793-805. |
| [50] |
张晓龙, 邓清月, 秦浩, 史利江, 苏迎庆, 张殷波, 牛俊杰. 不同海拔梯度灌丛草甸群落多样性的分布特征——以五台山亚高山-高山带南坡为例. 生态环境学报, 2020, 29(4): 657-664. |
| [51] |
屈兴乐, 方江平. 围栏封育对退化灌丛草地群落土壤特性和植被的影响. 北方园艺, 2019(3): 109-115. |
| [52] |
胡会峰, 王志恒, 刘国华, 傅伯杰. 中国主要灌丛植被碳储量. 植物生态学报, 2006, 30(4): 539-544. DOI:10.3321/j.issn:1005-264X.2006.04.001 |
| [53] |
张蔷, 李家湘, 徐文婷, 熊高明, 谢宗强. 中国亚热带山地杜鹃灌丛生物量分配及其碳密度估算. 植物生态学报, 2017, 41(1): 43-52. |
| [54] |
王希华, 闫恩荣, 严晓, 王良衍. 中国东部常绿阔叶林退化群落分析及恢复重建研究的一些问题. 生态学报, 2005, 25(7): 1796-1803. DOI:10.3321/j.issn:1000-0933.2005.07.037 |
| [55] |
董丹, 倪健. 利用CASA模型模拟西南喀斯特植被净第一性生产力. 生态学报, 2011, 31(7): 1855-1866. |
| [56] |
庞学勇, 向双, 赵文强, 李丹丹, 刘庆. 西南亚高山次生灌丛促进建群树种更新恢复技术. 应用与环境生物学报, 2021, 27(3): 667-676. |
| [57] |
姜凤岐, 杨瑞英, 薛才, 林鹤鸣. "三北"地区天然灌丛改造利用途径的研究. 生态学杂志, 1989, 8(4): 16-19. |
| [58] |
贾希洋, 周静静, 宿婷婷, 周瑶, 陈佳宝, 马红彬, 马静利, 王晓芳. 平茬密度对荒漠草原人工柠条林间生境的影响. 生态学报, 2020, 40(12): 4126-4136. |
| [59] |
王震, 张利文, 虞毅, 杨永华, 朝格吉乐, 高健斌, 高慧敏, 何淑清. 平茬高度对四合木生长及生理特性的影响. 生态学报, 2013, 33(22): 7078-7087. |
| [60] |
段广东, 裴志永, 郝少荣, 乔敬伟, 庞国辉, 王凯, 刘英琦. 毛乌素沙地沙柳人工林最优平茬周期的确定. 科学技术与工程, 2019, 19(7): 65-69. |
| [61] |
刘剑荣, 杨磊, 卫伟, 张钦弟. 半干旱黄土区柠条灌丛不同植被管理方式下的土壤水分. 草业科学, 2021, 38(8): 1439-1450. |
| [62] |
谢宗强, 江明喜. 三峡地区石灰岩灌丛植被特征及其合理利用. 植物学通报, 1995, 30(S2): 85-89. |
| [63] |
刘国荣, 松树奇, 刘国良, 王新颖, 张建英, 扈延成. 禁牧与放牧管理下灌丛草地植被变化. 内蒙古草业, 2005, 17(2): 41-45. |
| [64] |
马文明, 刘超文, 周青平, 邓增卓玛, 唐思洪, 迪力亚尔·莫合塔尔, 侯晨. 高寒草地灌丛化对土壤团聚体生态化学计量学及酶活性的影响. 草业学报, 2022, 31(1): 57-68. |
| [65] |
Eldridge D J, Maestre F T, Maltez-Mouro S, Bowker M A. A global database of shrub encroachment effects on ecosystem structure and functioning. Ecology, 2012, 93(11): 2499. DOI:10.1890/12-0749.1 |
| [66] |
Knapp A K, Briggs J M, Collins S L, Archer S R, Bret-Harte M S, Ewers B E, Peters D P, Young D R, Shaver G R, Pendall E, Cleary M B. Shrub encroachment in North American grasslands: shifts in growth form dominance rapidly alters control of ecosystem carbon inputs. Global Change Biology, 2008, 14(3): 615-623. DOI:10.1111/j.1365-2486.2007.01512.x |
| [67] |
Van Auken O W. Causes and consequences of woody plant encroachment into western North American grasslands. Journal of Environmental Management, 2009, 90(10): 2931-2942. DOI:10.1016/j.jenvman.2009.04.023 |
| [68] |
Miller R F, Rose J A. Fire history and western juniper encroachment in sagebrush steppe. Journal of Range Management, 1999, 52(6): 550-559. DOI:10.2307/4003623 |
| [69] |
Sanz-Elorza M, Dana E D, González A, Sobrino E. Changes in the high-mountain vegetation of the central Iberian peninsula as a probable sign of global warming. Annals of Botany, 2003, 92(2): 273-280. DOI:10.1093/aob/mcg130 |
| [70] |
Booth C A, Sanchez-Bayo F, King G W. Establishment of woody weeds in western new South Wales. 2. growth and competitive potential. The Rangeland Journal, 1996, 18(1): 80-98. DOI:10.1071/RJ9960080 |
| [71] |
Ratajczak Z, Nippert J B, Hartman J C, Ocheltree T W. Positive feedbacks amplify rates of woody encroachment in mesic tallgrass prairie. Ecosphere, 2011, 2(11): art121. |
| [72] |
Morgan J A, Milchunas D G, LeCain D R, West M, Mosier A R. Carbon dioxide enrichment alters plant community structure and accelerates shrub growth in the shortgrass steppe. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(37): 14724-14729. |
| [73] |
Brantley S T, Young D R. Contribution of sunflecks is minimal in expanding shrub thickets compared to temperate forest. Ecology, 2009, 90(4): 1021-1029. DOI:10.1890/08-0725.1 |
| [74] |
Báez S, Collins S L. Shrub invasion decreases diversity and alters community stability in northern Chihuahuan Desert plant communities. PLoS One, 2008, 3(6): e2332. DOI:10.1371/journal.pone.0002332 |
| [75] |
Howard K S C, Eldridge D J, Soliveres S. Positive effects of shrubs on plant species diversity do not change along a gradient in grazing pressure in an arid shrubland. Basic and Applied Ecology, 2012, 13(2): 159-168. DOI:10.1016/j.baae.2012.02.008 |
| [76] |
Blaum N, Rossmanith E, Popp A, Jeltsch F. Shrub encroachment affects mammalian carnivore abundance and species richness in semiarid rangelands. Acta Oecologica, 2007, 31(1): 86-92. DOI:10.1016/j.actao.2006.10.004 |
| [77] |
Li X Y, Zhang S Y, Peng H Y, Hu X, Ma Y J. Soil water and temperature dynamics in shrub-encroached grasslands and climatic implications: results from Inner Mongolia steppe ecosystem of North China. Agricultural and Forest Meteorology, 2013, 171/172: 20-30. DOI:10.1016/j.agrformet.2012.11.001 |
| [78] |
Ratajczak Z, Nippert J B, Collins S L. Woody encroachment decreases diversity across North American grasslands and savannas. Ecology, 2012, 93(4): 697-703. DOI:10.1890/11-1199.1 |
| [79] |
D'Odorico P, Fuentes J D, Pockman W T, Collins S L, He Y F, Medeiros J S, DeWekker S, Litvak M E. Positive feedback between microclimate and shrub encroachment in the northern Chihuahuan Desert. Ecosphere, 2010, 1(6): 1-11. |
| [80] |
Li H, Zhang J H, Hu H F, Chen L Y, Zhu Y K, Shen H H, Fang J Y. Shift in soil microbial communities with shrub encroachment in Inner Mongolia grasslands, China. European Journal of Soil Biology, 2017, 79: 40-47. DOI:10.1016/j.ejsobi.2017.02.004 |
| [81] |
Kerley G I H, Knight M H, de Kock M. Desertification of subtropical thicket in the Eastern Cape, South Africa: are there alternatives?. Environmental Monitoring and Assessment, 1995, 37(1): 211-230. |
| [82] |
Alofs K M, Fowler N L. Habitat fragmentation caused by woody plant encroachment inhibits the spread of an invasive grass. Journal of Applied Ecology, 2010, 47(2): 338-347. DOI:10.1111/j.1365-2664.2010.01785.x |
| [83] |
Brown J H, Valone T J, Curtin C G. Reorganization of an arid ecosystem in response to recent climate?change. Proceedings of the National Academy of Sciences of the United States of America, 1997, 94(18): 9729-9733. |
| [84] |
Castro H, Freitas H. Above-ground biomass and productivity in the Montado: from herbaceous to shrub dominated communities. Journal of Arid Environments, 2009, 73(4/5): 506-511. |
| [85] |
D'Odorico P, Okin G S, Bestelmeyer B T. A synthetic review of feedbacks and drivers of shrub encroachment in arid grasslands. Ecohydrology, 2012, 5(5): 520-530. DOI:10.1002/eco.259 |
| [86] |
Derner J D, Tischler C R, Polley H W, Johnson H B. Seedling growth of two honey mesquite varieties under CO2Enrichment. Rangeland Ecology & Management, 2005, 58(3): 292-298. |
| [87] |
Chen L Y, Li H, Zhang P J, Zhao X, Zhou L H, Liu T Y, Hu H F, Bai Y F, Shen H H, Fang J Y. Climate and native grassland vegetation as drivers of the community structures of shrub-encroached grasslands in Inner Mongolia, China. Landscape Ecology, 2015, 30(9): 1627-1641. DOI:10.1007/s10980-014-0044-9 |
| [88] |
Zavaleta E S, Kettley L S. Ecosystem change along a woody invasion chronosequence in a California grassland. Journal of Arid Environments, 2006, 66(2): 290-306. |
| [89] |
Tews J, Esther A, Milton S J, Jeltsch F. Linking a population model with an ecosystem model: assessing the impact of land use and climate change on savanna shrub cover dynamics. Ecological Modelling, 2006, 195(3/4): 219-228. |
| [90] |
Peng H Y, Li X Y, Li G Y, Zhang Z H, Zhang S Y, Li L, Zhao G Q, Jiang Z Y, Ma Y J. Shrub encroachment with increasing anthropogenic disturbance in the semiarid Inner Mongolian grasslands of China. CATENA, 2013, 109: 39-48. |
| [91] |
Maestre F T, Bowker M A, Puche M D, Hinojosa M B, Martínez I, García-Palacios P, Castillo A P, Soliveres S, Luzuriaga A L, Sánchez A M, Carreira J A, Gallardo A, Escudero A. Shrub encroachment can reverse desertification in semi-arid Mediterranean grasslands. Ecology Letters, 2009, 12(9): 930-941. |
| [92] |
Liu S S, Zhou L H, Li H, Zhao X A, Yang Y H, Zhu Y K, Hu H F, Chen L Y, Zhang P J, Shen H H, Fang J Y. Shrub encroachment decreases soil inorganic carbon stocks in Mongolian grasslands. Journal of Ecology, 2020, 108(2): 678-686. |
| [93] |
武锦辉. 基于多源遥感数据的物种多样性及陆表植被恢复力监测研究. 测绘学报, 2022, 51(8): 1826. |
| [94] |
刘佳丽, 范建容, 张茜彧, 杨超, 徐富宝, 张晓雪, 梁博. 高寒草地生长季/非生长季植被盖度遥感反演. 草业学报, 2021, 30(9): 15-26. |
| [95] |
杜中曼, 马文明, 周青平, 陈红, 邓增卓玛, 刘金秋. 基于遥感技术的植被识别方法研究进展. 生态科学, 2022, 41(6): 222-229. |
| [96] |
杨天垚, 邱建秀, 肖国安. 华北农业干旱监测与冬小麦估产研究. 生态学报, 2023, 43(5): 1936-1947. |
| [97] |
Zhang Y, Fang J N, Smith W K, Wang X A, Gentine P, Scott R L, Migliavacca M, Jeong S, Litvak M, Zhou S. Satellite solar-induced chlorophyll fluorescence tracks physiological drought stress development during 2020 southwest US drought. Global Change Biology, 2023, 29(12): 3395-3408. |
| [98] |
Picek L, Šulc M, Patel Y, Matas J. Plant recognition by AI: deep neural nets, transformers, and kNN in deep embeddings. Frontiers in Plant Science, 2022, 13: 787527. |
| [99] |
巫磊, 吴文挺. GEE平台下结合滤波算法和植被物候特征的互花米草遥感提取最优时间窗口确定. 地球信息科学学报, 2023, 25(3): 606-624. |
| [100] |
Yang T, Zhang G L, Li Y Z, Fan J W, Sun D F, Wang J, Di Y Y, You N S, Liu R Q, Zhang Q, Doughty R B. Satellite observed rapid green fodder expansion in northeastern Tibetan Plateau from 2010 to 2019. International Journal of Applied Earth Observation and Geoinformation, 2021, 102: 102394. |
| [101] |
李慧杰, 王兵, 牛香, 梁咏亮, 李静尧. 机器学习技术在生态学中的应用进展. 生态学杂志, 2023, 1-11. |
| [102] |
刘庆, 陈冬东. 中国典型灌丛研究: 格局与过程. 植物生态学报, 2022, 46(11): 1317-1320. |
| [103] |
Li X J, Zheng J G, Li X R, Tan H J, Pan Y X, Wei Y P. Responses of runoff, sedimentation, and induced nutrient loss to vegetation change in the Tengger Desert, Northern China. Soil Research, 2013, 51(2): 124-132. |
| [104] |
李愈哲, 樊江文, 张良侠, 翟俊, 刘革非, 李佳. 不同土地利用方式对典型温性草原群落物种组成和多样性以及生产力的影响. 草业学报, 2013, 22(1): 1-9. |
| [105] |
房福金, 肖金兰, 王东. 氮添加对川西高寒灌丛凋落枝化学计量特征及养分归还的影响. 生态学报, 2023, 43(7): 2927-2937. |
| [106] |
王小军, 刘光旭, 相爱存, 肖彤. 江西省油茶综合生产潜力与资源利用效率评估. 应用生态学报, 2020, 31(4): 1175-1184. |