 2024, Vol. 44
2024, Vol. 44文章信息
- 孔茹洁, 卫洁, 陈清, 赵英, 冯星星
- KONG Rujie, WEI Jie, CHEN Qing, ZHAO Ying, FENG Xingxing
- 土壤水分及其垂直分布对内蒙古草原冰草和冷蒿竞争的影响
- Effects of soil water and its vertical distribution on competition between Agropyron cristatum and Artemisia frigida in the Inner Mongolia grassland
- 生态学报. 2024, 44(6): 2495-2503
- Acta Ecologica Sinica. 2024, 44(6): 2495-2503
- http://dx.doi.org/10.20103/j.stxb.202304040678
-
文章历史
- 收稿日期: 2023-04-04
- 网络出版日期: 2023-12-09





2. 天津师范大学地理与环境科学学院, 天津 300387;
3. 鲁东大学资源与环境工程学院, 烟台 264025
2. College of Geographic and Environmental Sciences, Tianjin Normal University, Tianjin 300387, China;
3. College of Resources and Environmental Engineering, Ludong University, Yantai 264025, China
草地灌丛化作为退化草原的一种类型在全球范围内广泛发生, 使得国民经济和生态安全受到威胁[1]。近年来, 由于全球气候变化以及过度放牧等原因, 内蒙古典型草原植被退化严重, 出现了禾草群落向半灌木冷蒿(Artemisia frigida)群落的逆向演替[2], 相反, 围封恢复3—4年后, 出现冰草(Agropyron cristatum)突然成倍增加取代冷蒿变为优势种的“跃变”现象[3—4]。冰草群落取代冷蒿群落对于退化典型草原植被的恢复有重要的意义, 但是关于冰草-冷蒿的演替和竞争机制的研究鲜见报道。
水分是退化草原生产力和植被恢复的主要限制因子[5], 土壤水分的垂直分布情况, 是决定植被结构的关键因素[6]。降水是内蒙古草原土壤水分的主要来源, 6—8月份降水集中, >10mm的降水能增加0—30cm层土壤水分, 且土壤水分的响应随土层深度的增加而递减[7], 一般呈现出上湿下干的空间分布格局。然而, 随放牧强度的增加, 牲畜对土壤的践踏加剧, 草原土壤表层紧实度增加, 容重上升, 0—20cm土层土壤含水量逐渐下降[8], 而深层土壤含水量变化规律与表层恰恰相反[9], 放牧会破坏上层植被根系, 根系吸水能力减弱, 上层土壤水分向下运移, 使得放牧区下层土壤含水量相对升高[10], 土壤水分呈现出上干下湿的空间分布格局。此外, 持续干旱在草原频繁发生[11—12], 土壤水分呈现出上干下干的空间分布格局。草原土壤水分的垂直变化必然对草原植物种群变化产生影响。
由于禾草与灌木根系分布深浅的不同, 对土壤水分的空间利用有所差异。目前, 关于禾草-灌木水分利用策略有了一些研究[13—15]。干旱半干旱区域, 植物水分利用方式普遍符合两层水利用模式, 即浅根草本植物主要利用不稳定的表层土壤水, 深根系木本植物主要利用稳定的深层土壤水[16]。Sala[17]在巴塔哥尼亚草原的试验表明, 禾本科植物主要吸收土壤上层水分, 而灌木主要吸收土壤下层水分。鄂尔多斯高原草地研究表明, 沙米(Agriophyllum squarrosum)、本氏针茅(Stipa bungeana)等草本植物主要利用0—20cm层土壤水分, 籽蒿(Artemisia sphaerocephala)、油蒿(Artemisia ordosica)等灌木植物主要利用50cm以下土壤水分[18]。也有研究发现在湿润条件下浅根草本植物和深根灌木共同依赖浅层土壤水, 而在干旱条件下, 灌木开始转为深层吸水模式, 依赖深层土壤水[19—20]。因此, 在干旱条件下, 深层土壤水分具有相对优势时, 灌木的竞争力可能高于禾草。如科尔沁沙地和科罗拉多高原研究表明:干旱时, 盐蒿(Artemisia halodendron)、小叶锦鸡儿(Caragana microphylla)等灌木加强了对深层土壤水的利用率[21], 而浅根植物长毛落芒草(Oryzopsis hymenoides)因水分胁迫而死亡[22]。
内蒙古草原半灌木冷蒿为深根植物, 其主根入土可达100cm左右, 侧根发达, 在浅层土壤中长有大量的匍匐不定根[23]。冰草是典型的根茎型多年生禾本科草本植物, 属于浅根性植物, 其根量集中分布在25cm以上[24]。由于根系分布深浅不同, 内蒙古草原土壤水分的垂直变化可能会影响冰草和冷蒿的竞争格局。因此, 我们假设上层土壤水分充足时, 冰草的竞争力超过冷蒿;上层土壤干旱下层土壤相对湿润时, 冷蒿比冰草具有竞争优势。目前, 土壤含水量及其垂直分配对冰草-冷蒿竞争影响的研究鲜见报道。在我国草地灌丛化日趋严重的环境背景下, 研究土壤水分及其垂直分布对冰草-冷蒿竞争的影响, 对理解内蒙古退化草原冷蒿群落恢复演替机制有重要的理论和实践意义。
1 材料与方法本研究在内蒙古典型草原锡林浩特市周边锡林郭勒退化冷蒿草原进行, 该地气候类型属中温带半干旱大陆性气候, 冬季寒冷干燥, 夏季温暖湿润。该区域年平均温度2.65℃, 年平均降水量100—400mm, 主要集中在4—9月[25]。近年来, 锡林郭勒草原退化日趋严重, 草地灌丛化趋势明显, 本研究选择于2021年5月至2021年9月在内蒙古锡林浩特市温室大棚进行。
1.1 试验设计试验采用温室土培的方法, 试验设2因素, 即不同土壤深度水分处理和不同竞争序列种植方式。水分处理设3个水平:a.上干下湿:上层(0—30cm)干旱, 下层(30—60cm)湿润;b.上湿下干:上层湿润, 下层干旱;c.上干下干:上下层土壤均干旱。冷蒿对干旱胁迫的响应表明, 土壤含水量为12%时, 冷蒿可以正常生长, 当土壤含水量降低至6%以下时, 冷蒿则停止生长[26]。孙建等[27]对内蒙古典型草原生长季土壤水分监测数据表明, 5cm和20cm层土壤含水量对降雨量响应敏感, 波动范围在3.4%—32.2%之间。根据前人经验以及本课题组前期草原野外试验土壤含水量实际变化情况, 设置干旱处理土壤含水量为5%—10%, 湿润处理土壤含水量为25%—30%。种植方式设3个水平竞争序列:a.冰草单种, b.冷蒿单种, c.冰草和冷蒿1:1混种。单种处理设3次重复, 混种处理为防止变异设置5次重复, 共设33个栽培装置。
1.2 植株的移栽及培养2021年4月底, 挖取典型草原退化冷蒿样地土壤, 土壤分两层(0—30cm, 30—60cm)采集, 在室温下风干, 过3mm筛去除植物根系带回温室, 留作盆栽试验用。2021年5月生长季初, 在野外样地选取生长相对均匀的冷蒿、冰草斑块, 用高20cm直径30cm的铁皮圆圈柱和半圆圈柱打入土壤中, 挖取冰草和冷蒿斑块的原状土柱(深20cm带根系的土壤原状草块), 分别取冷蒿和冰草斑块各9个圆柱(单种用)和15个半圆柱(混种用), 带回温室准备移植栽培。
将前期采集并风干后的草原土壤分上下两层分装到上下层栽培桶中, 上层透明培养桶直径30cm, 高度38cm;下层培养桶直径35cm, 高度30cm。为尽量模拟野外土壤分布情况, 上层栽培桶装野外挖取的0—30cm的风干土壤, 下层栽培桶装30—60cm风干土壤。参考前人相关试验方法[28—29], 为使植物根系从上桶顺利长至下桶, 在上层培养桶底部均匀打12个直径为1.5cm的孔洞。上下桶中间设3cm厚蛭石隔水层, 阻断上下层土壤毛管水。下层培养桶装16kg过筛后的干土, 加水5kg;上层桶装入风干土约10kg, 并压平加水3kg, 之后将修整好的野外挖取的冷蒿和冰草带根土块移栽到上层桶中, 再加水约2kg, 使上下栽培桶土壤含水量均达到30%左右。
试验开始的前15d, 每天早晨7时用TDR 100土壤水分测量仪测量各栽培桶上下层土壤含水量, 探针长度选择12cm, 含水量每低至20%浇水, 保证上、下层土壤含水量维持在25%—30%之间。每周浇一次营养液, 保证移栽的幼苗成活并正常生长。培养过程中冰草斑块、冷蒿斑块单种和混种处理中出现其他杂草都要及时拔出。15d后, 观察到幼苗根系穿透隔水层, 根系分布于上下栽培桶时, 开始进行不同深度水分调节处理。采用土壤水分测量仪TDR 100每天测定土壤含水量, 湿润处理低至25%、干旱处理低至5%时浇水500g, 维持各处理土壤含水量在设定区间内。实验在通风的温室进行, 只进行遮蔽降水处理, 光照温度与野外实际状况相近。
1.3 数据测定及样品采集植物生长期末9月底进行植株的收获, 将地上部植株分物种齐土面剪下, 分别装入标记好的信封袋中, 带回实验室进行称重, 然后放入烘箱65℃烘干48h后再称干重, 测定地上生物量。
根系样品的采集采用网袋冲洗法。上下栽培桶带根系土壤装入孔径30—40目的尼龙网袋中, 通过人工抖动将部分土壤筛出, 如干旱处理的土壤很容易筛出来。然后将附着根系和少量土壤的网袋泡在装满水的塑料大桶中, 泡软后将剩余土壤揉出网袋, 多次清洗, 直到网袋中只留下根系。之后通过用水盆再次漂洗的方法初步将根系分为死根(漂在水上)和活根(沉在水下), 同时将根系中的砂粒和石头清除。清洗干净后再采用人为分根方法, 通过颜色和韧性等特征, 将根样进一步区分为活根、死根, 分别装入密封袋, 并放入冰箱冷藏待测。其中活根系样品采用根系扫描的方法测量根系长度, 将活根系放在根盘中, 倒入蒸馏水并用镊子将根系分散开, 以避免根系的重叠, 采用根系扫描仪Expression 11000XL进行扫描, 保存图像并用WinRhizo图像分析软件统计根长。最后, 将活根和死根样品装进信封袋中, 放入烘箱65℃烘干48h后称重, 计算地下总生物量。
1.4 数据处理比根长、相对竞争强度(Relative competition intensity, RCI)和竞争攻击力系数(Aggressivity, A)[30]的计算方法如下:
比根长=活根系长度/活根系生物量(m/g)。
相对竞争强度(RCI)和竞争攻击力系数(A), 以冰草、冷蒿的地上生物量为依据, 评价不同处理下两物种的竞争能力。
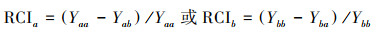
|

|
式中, a和b分别为冰草与冷蒿, Yaa和Ybb分别为单种时冰草与冷蒿的生物量, Yab和Yba分别为混种时冰草与冷蒿的生物量, p和q分别为混种方式下冰草与冷蒿的比例。若0<RCI<1则物种间存在竞争, 且RCI值越大, 该物种的竞争能力越弱;若RCI<0则另一物种的存在能促进该物种的生长。因为Aab=-Aba, 故仅计算冰草的竞争攻击力系数Aab即可。若Aab>0则冰草的竞争能力超过冷蒿, 若Aab<0则冷蒿的竞争能力超过冰草。
本研究中测定的所有数据使用Microsoft Excel 2019进行整理以及基础分析。使用SPSS 25软件进行单因素方差分析(ANOVA), 以分析不同水分处理以及种植方式对植物地上、地下生物量、根长、比根长以及相对竞争强度、竞争攻击力系数的影响, 采用邓肯法多重比较进一步探讨各影响因素中不同处理之间平均值的差异性, 同时采用t检验比较相对竞争强度、竞争攻击力系数与0之间的差异, 显著水平均为0.05。使用Origin 2019软件进行绘图。
2 结果与分析 2.1 不同水分处理和种植方式对冰草和冷蒿地上生物量以及种间竞争的影响 2.1.1 不同水分处理和种植方式对冰草和冷蒿地上生物量的影响冰草地上生物量受土壤水分显著影响, 无论是冰草单种还是冰草-冷蒿混种处理, 湿润处理(上干下湿、上湿下干)的冰草地上生物量显著高于上干下干处理。相反, 冷蒿地上生物量受水分影响不显著, 无论是冷蒿单种还是冰草-冷蒿混种处理, 湿润处理的冷蒿地上生物量与上干下干处理差异不显著。无论单种还是混种, 上干下湿和上湿下干处理间地上总生物量和单物种生物量差异均不显著(图 1, 2)。

|
| 图 1 不同水分处理和种植方式地上总生物量 Fig. 1 Total aboveground biomass of different water treatments and planting methods 相同小写字母表示同一种植方式下, 不同水分处理之间差异不显著(P>0.05) |

|
| 图 2 混种处理冰草与冷蒿的地上生物量 Fig. 2 Aboveground biomass of Agropyron cristatum and Artemisia frigida under intercropping treatment 相同小写字母表示混种条件下, 同一物种不同水分处理之间差异不显著(P>0.05) |
不同水分处理下, 冰草与冷蒿的相对竞争强度均显著大于0, 表明混种时冰草和冷蒿之间存在显著的种间竞争关系。冰草的相对竞争强度湿润处理小于上干下干处理, 表明在湿润处理下冰草的竞争能力比上干下干处理时更强。而冷蒿在不同水分处理间相对竞争强度无显著差异, 表明不同水分处理冷蒿的竞争能力没有明显变化(表 1)。
| 处理类别 Processing category |
冰草的相对竞争强度RCIa The relative competition intensity of Agropyron cristatum RCIa |
冷蒿的相对竞争强度RCIb The relative competition intensity of Artemisia frigida RCIb |
冰草的竞争攻击力系数Aab The aggressivity of Agropyron cristatum Aab |
| 上干下湿Drought-wet | 0.55ab* | 0.60a* | 0.09a* |
| 上湿下干Wet-drought | 0.47b* | 0.56a* | 0.19a |
| 上干下干Drought-drought | 0.64a* | 0.52a* | -0.24b* |
| RCI:相对竞争强度Relative competition intensity;A:竞争攻击力系数Aggressivity;不同小写字母表示不同水分处理之间差异显著(P<0.05), *表示RCI、A与0相比差异显著(P<0.05) | |||
湿润处理冰草的竞争攻击力系数均大于0, 表明湿润处理冰草的竞争力强于冷蒿, 在资源利用上处于优势地位。上干下干处理冰草的竞争攻击力系数小于0, 表明干旱处理冷蒿的竞争力强于冰草。
2.2 不同水分处理和种植方式对冰草和冷蒿地下生物量以及根系形态的影响 2.2.1 不同水分处理和种植方式对冰草和冷蒿地下生物量的影响各处理植物根系主要分布在0—30cm层, 占总根系生物量的79%—98%(表 2)。土壤水分变化显著影响冰草的上、下土层的地下总生物量, 而对冷蒿没有显著影响。上层(0—30cm)土壤, 与上层干旱(上干下干、上干下湿)处理相比, 单种条件下上湿下干处理冰草地下总生物量显著增加, 而冷蒿单种、冰草-冷蒿混种处理的地下生物量没有发生显著变化(表 2);上层土壤不同种植方式相比, 冷蒿单种、冰草-冷蒿混种的地下生物量显著高于冰草单种处理。下层(30—60cm)土壤, 水分变化仍然显著影响冰草的地下总生物量, 而对冷蒿没有显著影响。与上干下干处理相比, 湿润处理冰草单种地下生物量显著增加, 而湿润处理冷蒿单种、冰草-冷蒿混种处理地下生物量没有发生显著变化;下层30—60cm土壤, 不同种植方式相比, 冰草平均地下生物量显著高于冷蒿单种、冰草-冷蒿混种处理(表 2)。
| 土层深度/cm Soil depth |
处理类别 Processing category |
冰草单种Agropyron cristatum monoculture | 冷蒿单种 Artemisia frigida monoculture |
冰草-冷蒿混种 Intercropping of Agropyron cristatum and Artemisia frigida |
平均 Average |
| 0—30 | 上干下湿 | 16.53±0.64bB | 23.59±3.33aAB | 23.45±1.29aA | 21.19±1.38b |
| 上湿下干 | 25.18±0.39aB | 28.53±2.64aAB | 29.52±2.52aA | 27.75±1.37a | |
| 上干下干 | 18.57±1.67bB | 23.86±2.53aAB | 27.92±0.98aA | 23.45±1.50ab | |
| 平均 | 20.09±1.41B | 25.33±1.64A | 26.97±1.22A | 24.13±1.46 | |
| 30—60 | 上干下湿 | 3.70 ±0.17aA | 1.75±0.33aB | 1.88±0.32aB | 2.44±0.31a |
| 上湿下干 | 2.64±0.51abA | 1.57±0.25aA | 1.81±0.44aA | 2.01±0.26ab | |
| 上干下干 | 1.82±0.36bA | 1.09±0.26aA | 1.04±0.15aA | 1.32±0.17b | |
| 平均 | 2.72±0.33A | 1.47±0.17B | 1.58±0.21B | 1.92±0.27 | |
| 相同小写字母表示同一种植方式下, 不同水分处理之间差异不显著(P>0.05);相同大写字母表示同一水分处理下, 不同种植方式处理之间差异不显著(P>0.05) | |||||
植物根系长度受土壤水分影响显著, 湿润处理的平均根长显著高于干旱处理(表 3)。上层0—30cm土壤中, 冰草、冷蒿单种条件下, 上层湿润处理(上湿下干)根系长度显著高于上干下干处理;冰草-冷蒿混种条件下不同水分处理之间根长差异不显著。下层30—60cm土壤中, 湿润处理植物根长高于干旱处理。
| 土层深度/cm Soil depth |
处理类别 Processing category |
冰草单种 Agropyron cristatum monoculture |
冷蒿单种 Artemisia frigida monoculture |
冰草-冷蒿混种 Intercropping of Agropyron cristatum and Artemisia frigida |
平均 Average |
| 0—30 | 上干下湿 | 1873±89aA | 864±21bB | 1634±211aA | 1457±130b |
| 上湿下干 | 2131±228aA | 1711±85aB | 2179±233aA | 2007±110a | |
| 上干下干 | 1332±79bA | 728±210bB | 1542±222aA | 1201±105b | |
| 平均 | 1779±139A | 1101±167B | 1785±140A | 1555±169 | |
| 30—60 | 上干下湿 | 531±114aA | 253±33aB | 334±53aAB | 373±43a |
| 上湿下干 | 460±50aA | 267±64aA | 298±59aA | 341±31ab | |
| 上干下干 | 327±36aA | 158±19aB | 133±16bB | 206±32b | |
| 平均 | 439±48A | 226±27B | 255±34B | 307±43 |
不同种植方式相比, 上层0—30cm土壤, 冷蒿单种根长最低, 冰草单种、冰草-冷蒿混种处理的根长显著高于冷蒿单种处理, 约为冷蒿单种处理的1.2—2.2倍。下层30—60cm土壤, 冷蒿单种根长依然最低, 冰草单种处理根长最高, 冰草单种的根长约为冷蒿单种、冰草-冷蒿混种处理的1.5—2.3倍(表 3)。
2.2.3 不同水分处理和种植方式对冰草和冷蒿比根长的影响土壤水分变化显著影响上层土壤植物的比根长。上层0—30cm土壤, 单种条件下与上层湿润处理相比, 上干下干处理冰草的比根长下降不明显, 而冷蒿的比根长显著下降(表 4);上层土壤不同种植方式相比, 冰草单种的比根长显著高于冷蒿单种(表 4)。下层(30—60cm)土壤, 不同水分处理及不同种植方式植物比根长均无显著差异。
| 土层深度/cm Soil depth |
处理类别 Processing category |
冰草单种 Agropyron cristatum monoculture |
冷蒿单种 Artemisia frigida monoculture |
冰草-冷蒿混种 Intercropping of Agropyron cristatum and Artemisia frigida |
平均 Average |
| 0—30 | 上干下湿 | 128±0aA | 47±5bC | 92±8aB | 90±10a |
| 上湿下干 | 97±7abA | 68±21aB | 86±7aAB | 84±5a | |
| 上干下干 | 80±11bA | 34±6bB | 77±11aA | 64±8a | |
| 平均 | 102±8A | 50±5B | 85±5AB | 79±5 | |
| 30—60 | 上干下湿 | 143±27aA | 149±17aA | 180±6aA | 157±10a |
| 上湿下干 | 179±15aA | 172±31aA | 171±11aA | 174±9a | |
| 上干下干 | 186±21aA | 153±16aA | 135±11aA | 157±10a | |
| 平均 | 170±13A | 158±12A | 162±7A | 163±6 |
冰草和冷蒿对土壤水分变化的响应不同, 湿润处理冰草单种和混种的地上生物量显著高于上干下干处理, 而冷蒿地上生物量没有受土壤干湿水分变化的显著影响(图 1, 2)。湿润的土壤环境中冰草竞争力强于冷蒿, 干旱的土壤环境中冷蒿的竞争力强于冰草(表 1)。这与黄土高原半干旱区的研究结果相似, 与半灌木兴安胡枝子(Lespedeza davurica)相比, 白羊草(Bothriochloa ischaemum)的生物量随土壤水分减少显著降低, 对土壤水分有效性的增加更敏感[31]。荒漠草原也发现, 与蒙古冰草(Agropyron mongolicum)相比, 柠条锦鸡儿(Caragana korshinskii)在土壤干旱环境下死亡率较低, 竞争性更强[32]。适宜水分条件下冰草的日均耗水量分别是中度、重度干旱的1.46和2.26倍, 属于高耗水草种, 充足的水分能够促进冰草的生长, 适宜水分(田间持水量的75%)条件下冰草的株高和叶面积明显高于干旱条件(田间持水量40%—55%)下的株高和叶面积[33]。而持续干旱的环境下冰草的生长高度降低, 生长期缩短, 干物质积累量明显下降[34]。与冰草相比, 冷蒿更有利于在干旱环境中获得竞争优势。冷蒿具有较低的水分竞争系数, 尤其是干旱情况下更低, 它蒸腾耗水减少, 且可以牺牲光能利用效率, 保持低蒸腾、低光合等方式来适应干旱环境, 具有较高的抗旱性[35]。在研究水分对克氏针茅(Stipa krylovii)和冷蒿生殖生长的影响中发现, 与增水处理相比, 自然降水条件下冷蒿的生殖枝数量和重量反而更高[36]。土壤含水量为6%时, 冷蒿可以抵抗干旱胁迫对其造成的不利影响[26]。冷蒿繁殖旺盛, 生根萌蘖的再生生长能力强, 在草地退化的环境下依然能保持一定的生产力水平[37]。
3.2 土壤水分变化对冰草和冷蒿地下生物量和根系形态的影响植物根系具有高度可塑性, 可根据外界环境的改变调整根系形态结构以响应胁迫[38—39]。本研究观察到冰草的根系呈白色细长胡须状, 冷蒿根系呈褐色较粗较短, 上层0—30cm土壤冰草的根长、比根长显著高于冷蒿, 约为冷蒿1.6—2.1倍。下层土壤30—60cm层为新生根系, 冰草单种的地下根系生物量和根长也都显著高于冷蒿单种, 即使是在上干下干处理的土壤环境中(表 2, 3)。根系是影响水分竞争能力的重要指标之一, 资源受限时, 植物根系可以通过增加根系长度、比根长和根表面积等, 帮助植物获得更多的水分和养分资源[40—41]。通过建立禾本科牧草水分竞争能力的评价指标体系发现, 植物根长与牧草水分竞争能力显著相关, 是决定其水分竞争能力的重要指标之一[42]。黄土高原研究发现, 与湿润处理(75%田间持水量)相比, 多年生草本植物柳枝稷(Panicum virgatum L.)在干旱(35%田间持水量)条件下, 有增加比根长和降低根平均直径的趋势, 而多年生亚灌木兴安胡枝子则相反[43]。本研究中, 与上层湿润处理相比, 上干下干处理冰草的比根长没有显著下降, 而冷蒿比根长显著下降(表 4), 表明干旱条件下, 冰草可能通过不减少根系比根长来增强水分的吸收能力, 适应干旱环境。在内蒙古温室实验和宁夏荒漠草原的研究中也发现相似结果, 蒙古冰草通过增加根长和根尖数以应对水分减少状况[44—45]。然而, 干旱条件下, 尽管冰草根系形态做出响应, 比冷蒿有更高的根长和比根长值, 但其地上生物量仍然显著下降, 地上竞争力低于冷蒿。
前人研究认为冰草为浅根性植物, 根系集中分布在25cm以上土层[24], 半灌木冷蒿为深根植物, 其主根系可深入100cm左右土层中[23]。我们推测上层土壤水分增加后冰草竞争力高于冷蒿, 下层土壤水分增加后冷蒿竞争力高于冰草。然而本试验结果与假设不同, 上干下湿与上湿下干处理间, 冰草与冷蒿地上竞争力差异不显著。冰草新生根系不仅可以分布到30—60cm层土壤, 而且比冷蒿根系在下层30—60cm土壤中生长地更多更长。造成与假设不同的可能原因:1)本研究只是一年的试验结果, 而植物竞争的研究中多进行两年及两年以上实验[46—47]。冰草和冷蒿均为多年生的植物, 随着种植时间的延长, 研究结果可能会发生变化。2)温室培养和野外的环境条件不同。冰草和冷蒿均为多年生的植物, 野外环境下多种物种共存, 根系有多年的积累, 竞争效应受多种因素影响。而温室培养上下分层设计, 通过对上层培养桶底部打孔洞的方式, 使植物根系伸长至下层桶, 一定程度上改变了根系的空间分布和构型, 可能对结果产生一定影响。3)研究表明, 环境变化后, 植物地下部的竞争比地上部的竞争更加明显[48], 本试验由于采样时环境和技术条件限制, 没有将混种处理冷蒿和冰草的根系分开分析, 只是肉眼观察到混种处理, 冰草的根系胡须更密集更长。本研究只是利用地上生物量计算冰草和冷蒿的竞争系数, 无法比较冰草和冷蒿的地下竞争情况。未来的研究需要扩大试验时间尺度, 分物种测量根系指标, 更加深入地研究两种植物混种时地下竞争情况。
4 结论1) 与冷蒿相比, 冰草对水分变化更敏感。与干旱处理相比, 湿润条件下冰草的地上、地下生物量均显著增加, 冷蒿地上、地下生物量则无显著变化。
2) 湿润的土壤环境中, 冰草的竞争力大于冷蒿;干旱的土壤环境中, 冷蒿的竞争力大于冰草;上干下湿和上湿下干处理间, 冰草和冷蒿的竞争力差异不显著。
3) 干旱和湿润的土壤环境中, 冰草和冷蒿的根系均可分布在30—60cm土壤中, 且土壤30—60cm层冰草的根生物量和根长均显著高于冷蒿。冰草并非浅根植物, 深层土壤水分优势促进冷蒿竞争力高于冰草的试验假设有待未来研究进一步地验证。
| [1] |
蔡文涛, 来利明, 李贺祎, 周继华, 管天玉, 张晓龙, 高楠楠, 郑元润. 草地灌丛化研究进展. 应用与环境生物学报, 2016, 22(4): 531-537. |
| [2] |
梁秀华. 内蒙古草原退化与恢复演替机理的探讨. 中国畜牧兽医文摘, 2018, 34(6): 31-50. |
| [3] |
Chen Q, Wang Z L, Zou C B, Fan Y, Dittert K, Lin S. Legacy effects of historical grazing affect the response of vegetation dynamics to water and nitrogen addition in semi-arid steppe. Applied Vegetation Science, 2018, 21(2): 229-239. DOI:10.1111/avsc.12364 |
| [4] |
刘钟龄, 王炜, 郝敦元, 梁存柱. 内蒙古草原退化与恢复演替机理的探讨. 干旱区资源与环境, 2002, 16(1): 84-91. |
| [5] |
Chen Q, Hooper D U, Li H, Gong X Y, Peng F, Wang H, Dittert K, Lin S. Effects of resource addition on recovery of production and plant functional composition in degraded semiarid grasslands. Oecologia, 2017, 184(1): 13-24. DOI:10.1007/s00442-017-3834-3 |
| [6] |
Eggemeyer K D, Awada T, Harvey F E, Wedin D A, Zhou X H, Zanner C W. Seasonal changes in depth of water uptake for encroaching trees Juniperus virginiana and Pinus ponderosa and two dominant C4 grasses in a semiarid grassland. Tree Physiology, 2009, 29(2): 157-169. |
| [7] |
张存厚, 杨丽萍, 越昆, 刘朋涛, 张德龙. 锡林郭勒典型草原土壤水分对降水过程的响应. 干旱区资源与环境, 2022, 36(8): 133-139. |
| [8] |
郭振宁, 孙世贤, 擅建国, 郭月峰, 祁伟, 王涵, 张久飞. 放牧强度对荒漠草原土壤含水量及群落特征的影响. 中国草地学报, 2022, 44(6): 27-35. |
| [9] |
苏都, 红梅, 陈有君, 白佳君, 额尔顿奥其尔, 阿木日吉日嘎拉. 放牧强度对荒漠草原淡栗钙土水分、养分及生物量的影响. 内蒙古农业大学学报: 自然科学版, 2011, 32(3): 132-137. |
| [10] |
徐冉, 张圣微, 朱仲元, 庞文台. 放牧对典型草原区土壤水分及其对降雨响应规律的影响. 中国草地学报, 2019, 41(3): 59-66. |
| [11] |
Wang Q A, Zhao L, Wang M L, Wu J J, Zhou W, Zhang Q P, Deng M E. A random forest model for drought: monitoring and validation for grassland drought based on multi-source remote sensing data. Remote Sensing, 2022, 14(19): 4981. DOI:10.3390/rs14194981 |
| [12] |
Ru J Y, Wan S, Hui D F, Song J. Overcompensation of ecosystem productivity following sustained extreme drought in a semiarid grassland. Ecology, 2023, 104(4): 3997. DOI:10.1002/ecy.3997 |
| [13] |
Wei Y F, Fang J, Liu S, Zhao X Y, Li S G. Stable isotopic observation of water use sources of Pinus sylvestris var. mongolica in Horqin Sandy Land, China. Trees, 2013, 27(5): 1249-1260. DOI:10.1007/s00468-013-0873-1 |
| [14] |
Zhu Y J, Wang G J, Li R Q. Seasonal dynamics of water use strategy of two Salix shrubs in alpine sandy land, Tibetan Plateau. Plos One, 2016, 11(5): e0156586. DOI:10.1371/journal.pone.0156586 |
| [15] |
朱雅娟, 崔清国, 杜娟, 许素寒, 刘志兰. 毛乌素沙地三种灌木群落的水分利用过程. 生态学报, 2020, 40(13): 4470-4478. |
| [16] |
Ward D, Wiegand K, Getzin S. Walter's two-layer hypothesis revisited: back to the roots!. Oecologia, 2013, 172(3): 617-630. DOI:10.1007/s00442-012-2538-y |
| [17] |
Sala O E, Golluscio R A, Lauenroth W K, Soriano A. Resource partitioning between shrubs and grasses in the Patagonian steppe. Oecologia, 1989, 81(4): 501-505. DOI:10.1007/BF00378959 |
| [18] |
李巧燕, 来利明, 周继华, 姜联合, 杨柳, 易三桂, 孙清琳, 郑元润. 鄂尔多斯高原草地灌丛化不同阶段主要植物水分利用特征. 生态学杂志, 2019, 38(1): 89-96. |
| [19] |
Zou C B, Barnes P W, Archer S, McMurtry C R. Soil moisture redistribution as a mechanism of facilitation in savanna tree-shrub clusters. Oecologia, 2005, 145(1): 32-40. DOI:10.1007/s00442-005-0110-8 |
| [20] |
Nippert J B, Knapp A K. Soil water partitioning contributes to species coexistence in tallgrass prairie. Oikos, 2007, 116(6): 1017-1029. DOI:10.1111/j.0030-1299.2007.15630.x |
| [21] |
陈定帅, 董正武, 高磊, 陈效民, 彭新华, 司炳成, 赵英. 不同降水条件下科尔沁沙地小叶锦鸡儿和盐蒿的水分利用动态. 植物生态学报, 2017, 41(12): 1262-1272. |
| [22] |
Schwinning S, Starr B I, Ehleringer J R. Summer and winter drought in a cold desert ecosystem (Colorado Plateau) part Ⅰ: effects on soil water and plant water uptake. Journal of Arid Environments, 2005, 60(4): 547-566. DOI:10.1016/j.jaridenv.2004.07.003 |
| [23] |
占布拉, 陈世璜, 张昊, 曹亮晓, 布仁吉雅. 冷蒿的特性和生态地理分布的研究. 内蒙古农牧学院学报, 1999, 20(1): 6-12. |
| [24] |
陈世鐄. 内蒙古草原植物根系类型. 呼和浩特: 内蒙古人民出版社, 1987.
|
| [25] |
王爽, 李庆旭, 张彪. 锡林郭勒盟净初级生产力时空变化及其气候影响. 生态学杂志, 2021, 40(3): 825-834. |
| [26] |
吴建华. 冷蒿对干旱胁迫适应机制的研究[D]. 呼和浩特: 内蒙古农业大学, 2009.
|
| [27] |
孙建, 刘苗, 李胜功, 胡中民, 孙晓敏, 温学发, 张雷明. 内蒙古典型草原克氏针茅与冰草的生存策略. 生态学报, 2011, 31(8): 2148-2158. |
| [28] |
朱林, 王甜甜, 赵学琳, 祁亚淑, 许兴. 紫花苜蓿和斜茎黄耆水力提升作用及其对伴生植物的效应. 植物生态学报, 2020, 44(7): 752-762. |
| [29] |
Wan C G, Xu W W, Sosebee R E, Machado S, Archer T. Hydraulic lift in drought-tolerant and-susceptible maize hybrids. Plant and Soil, 2000, 219(1): 117-126. |
| [30] |
李晓雪, 贺燕燕, 杨文航, 王朝英, 燕江伟, 崔振, 李昌晓. 不同水分处理和密度配置对牛鞭草与狗牙根生长与种间竞争的影响. 生态学报, 2018, 38(9): 3046-3058. |
| [31] |
Xu W Z, Niu F, Kang F R, Xu B C, Deng X P, Palta J, Chen Y L. Biomass allocation and competitive ability of a semiarid perennial grass and a legume in mixtures under periodical soil water decreasing conditions. Agronomy, 2022, 12(10): 2256. DOI:10.3390/agronomy12102256 |
| [32] |
张振杰, 于露, 王红梅. 荒漠草原向灌丛转变过程两种优势植物定植土壤水分阈值特征. 生态学报, 2022, 42(19): 8061-8072. |
| [33] |
郭颖, 韩蕊莲, 梁宗锁. 土壤干旱对黄土高原4个乡土禾草生长及水分利用特性的影响. 草业学报, 2010, 19(2): 21-30. |
| [34] |
蒋菊芳, 王鹤龄, 魏育国, 丁文魁, 杨永龙, 任丽雯, 程倩, 杨华. 不同水分处理对荒漠刺蓬和冰草生长的影响. 干旱气象, 2016, 34(5): 847-851. |
| [35] |
崔骁勇, 陈佐忠, 杜占池. 半干旱草原主要植物光能和水分利用特征的研究. 草业学报, 2001, 10(2): 14-21. |
| [36] |
张昊, 李鑫, 姜凤和, 林国辉, 杜丽华. 水分对克氏针茅和冷蒿生殖生长的影响. 草地学报, 2005, 13(2): 106-110. |
| [37] |
杨持, 宝音陶格涛, 李良. 冷蒿种群在不同放牧强度胁迫下构件的变化规律. 生态学报, 2001, 21(3): 405-408. |
| [38] |
Henry A, Cal A J, Batoto T C, Torres R O, Serraj R. Root attributes affecting water uptake of rice (Oryza sativa) under drought. Journal of Experimental Botany, 2012, 63(13): 4751-4763. DOI:10.1093/jxb/ers150 |
| [39] |
张翠梅, 师尚礼, 吴芳. 干旱胁迫对不同抗旱性苜蓿品种根系生长及生理特性影响. 中国农业科学, 2018, 51(5): 868-882. |
| [40] |
鲍雅静, 曹明, 李政海, 郭鹏, 张靖, 秦洁. 羊草与大针茅根系构型对水分梯度响应的比较研究. 生态学报, 2019, 39(3): 1063-1070. |
| [41] |
Schippers P, Olff H. Biomass partitioning, architecture and turnover of six herbaceous species from habitats with different nutrient supply. Plant Ecology, 2000, 149(2): 219-231. DOI:10.1023/A:1026531420580 |
| [42] |
张国娟. 禾本科牧草水分竞争能力的评价指标体系建立[D]. 杨凌: 西北农林科技大学, 2010.
|
| [43] |
Wang S Q, Liu J B, Kang J Y, Xu B C, Chen Y L. Root growth and morphology of switchgrass (Panicum virgatum L.) and bush clover (Lespedeza davurica S.) in mixed plantation under varying soil water and phosphorus supply conditions. Pakistan Journal of Botany, 2021, 53(4): 1227-1237. |
| [44] |
马艳红, 刘旭婷, 张旭婷, 于肖夏, 姜超, 于卓. 干旱胁迫对蒙古冰草苗期根系特征及生理特性的影响. 种子, 2017, 36(11): 66-69. |
| [45] |
史佳梅. 异质生境下蒙古冰草表型性状及生理生态适应策略研究[D]. 银川: 宁夏大学, 2022.
|
| [46] |
蒋智林, 王五云, 雷桂生, 桂富荣, 刘万学, 李正跃. 紫茎泽兰与4种功能型草本植物根系生长特征和竞争效应. 应用生态学报, 2014, 25(10): 2833-2839. |
| [47] |
刘屹湘. 小麦/玉米间作体系竞争—恢复过程的根系互作机制[D]. 北京: 中国农业大学, 2015.
|
| [48] |
王政权, 王军邦, 孙志虎, 范志强, 韩有志. 水曲柳苗木地下竞争与地上竞争的定量研究. 生态学报, 2003, 23(8): 1512-1518. |