 2024, Vol. 44
2024, Vol. 44文章信息
- 韩志立, 张庆, 张署军, 曾嘉伟, 徐然亮, 程军回, 陶冶, 张元明, 尹本丰
- HAN Zhili, ZHANG Qing, ZHANG Shujun, ZENG Jiawei, XU Ranliang, CHENG Junhui, TAO Ye, ZHANG Yuanming, YIN Benfeng
- 温带荒漠灌丛和藓类结皮对土壤养分的贡献
- Contribution of shrubs and moss crusts to soil nutrient in a temperate desert
- 生态学报. 2024, 44(5): 1951-1961
- Acta Ecologica Sinica. 2024, 44(5): 1951-1961
- http://dx.doi.org/10.20103/j.stxb.202305080959
-
文章历史
- 收稿日期: 2023-05-08
- 网络出版日期: 2023-12-20





2. 新疆农业大学, 资源与环境学院, 乌鲁木齐 830052;
3. 中国科学院大学, 北京 100049;
4. 新疆师范大学, 生命科学学院, 乌鲁木齐 830017
2. College of Resources and Environment, Xinjiang Agricultural University, Urumqi 830052, China;
3. University of Chinese Academy of Sciences, Beijing 100049, China;
4. College of Life Sciences, Xinjiang Normal University, Urumqi 830017, China
土壤是植物赖以生存的物质基础, 能够为植物生长提供稳定的环境与营养条件。碳、氮和磷等养分是土壤中重要的营养元素, 其含量及有效性, 不仅影响着植物的生长发育、群落组成及生产力水平[1], 而且对维持区域生态系统的稳定具有重要作用[2]。此外, 土壤养分是反映土壤质量的重要指标, 直接反映出土壤现状以及于周围环境的耦合现状。在荒漠生态系统中, 植物的生长不仅受到水分的限制, 土壤养分贫瘠也是制约植物发育的重要因素[3]。土壤养分的供给与植物生长之间的平衡关系决定了荒漠生态环境演化的过程, 一旦这种平衡关系遭到破坏, 对脆弱的荒漠生态系统将会造成重大影响[4]。所以, 探明荒漠地表植被分布类型对土壤养分分布格局的影响对理解植被与土壤间的养分互馈机制具有重要意义。
荒漠生态系统占据了全球陆地表面积的45.4%, 生物土壤结皮是该区域重要的地表覆被类型, 在部分荒漠地区的盖度可达70%左右[5—6]。生物土壤结皮是由具有较强耐旱特性的蓝藻、地衣、苔藓等隐花植物与土壤表面颗粒胶结形成的复合体, 在防风固沙、生态系统恢复及养分循环中具有重要意义[7]。生物土壤结皮通过光合生物和固氮微生物对碳、氮的固定作用是荒漠生态系统土壤重要的碳氮来源[8]。随着生物土壤结皮的发育阶段的不同, 其对土壤养分的贡献也有显著差异。其中藓类结皮作为生物土壤结皮发育的高级阶段, 可显著影响土壤微环境, 有效增强土壤稳定性和抗风蚀能力, 改善土壤水分和养分循环, 为其他维管植物的生长发育创造有利条件[9]。相对于裸沙以及结皮的早中期发育阶段, 藓类结皮对碳、氮和磷相关养分及酶活性的提升具有更强的贡献[10]。
在微尺度荒漠景观下, 微生境的变化对土壤养分的空间分布具有重要影响。其中, 灌丛的斑块状分布塑造了荒漠生态系统两种重要微生境单元(灌丛下和裸露地)。灌丛的冠幅阴影、凋落物和根系活动可以改变土壤微环境, 凋落物输入加速了土壤的形成并增加了表土厚度, 同时也极大地影响了土壤理化性质[11—13]。此外, 灌丛的存在能够增加土壤细小颗粒, 有效防止表土粗化、促进养分的积累。灌丛根系的生长可以增大土壤孔隙度, 降低土壤容重, 有利于土壤气体交换, 为微生物生长繁殖创造适宜的生存环境, 从而促进微生物对凋落物的分解, 提高土壤养分的有效性[14]。总体而言, 灌丛的“冷岛”“湿岛”和“肥岛”效应对土壤养分具有明显的聚集效应, 这也为喜阴喜湿的藓类植物提供了定植的理想场所。尽管, 诸多研究分别对灌丛或藓类结皮对土壤养分的影响均做了大量研究, 但二者对荒漠区土壤养分的贡献程度还未量化。量化荒漠灌丛和藓类结皮对土壤养分的贡献能够帮助认识在荒漠生态系统养分循环过程的主要贡献者, 对荒漠生态系统稳定管理和生态系统修复提供物种选择依据。
基于此, 提出以下科学问题:荒漠典型灌丛和藓类结皮及二者的交互作用对土壤养分有何影响?二者对土壤养分变化的贡献度如何?为解答此科学问题, 选择中国北方典型温带荒漠古尔班通古特沙漠为研究区, 以优势灌丛膜果麻黄(Ephedra przewalskii)灌丛和生物土壤结皮发育的高级阶段藓类结皮为研究对象, 采集裸露地、自然状态灌丛下的裸沙与藓类结皮土壤样品, 对比分析不同微生境下土壤碳、氮、磷、钾相关养分的变化特征, 探究影响土壤养分变化的关键因子, 解析灌丛与藓类结皮这两大荒漠重要的景观单元对土壤养分空间异质性的贡献度大小。为荒漠生态系统土壤养分状况评估提供数据支撑, 为荒漠植物多样性和土壤稳定性的保护和管理提供重要参考。
1 材料与方法 1.1 研究区概况研究区位于中国新疆北部准噶尔盆地腹地的古尔班通古特沙漠(44.18°—46.33°N, 84.52°—90.00°E), 该沙漠是中国面积最大的固定、半固定沙漠, 海拔300—600 m, 属典型的温带大陆性气候, 年平均降水量70—150 mm, 年蒸发量达2000 mm以上, 夏季炎热, 极端高温可达到40 ℃以上[15]。与其他荒漠生态系统不同, 古尔班通古特沙漠冬季具有10—30 cm的稳定积雪, 占年降雨量的25%。线性沙垄和树枝状沙垄组成了该沙漠的基本景观特征, 植被主要由白梭梭(Haloxylon persicum)、梭梭(Haloxylon ammodendron)、膜果麻黄(Ephedra przewalskii)、沙拐枣(Calligonum mongolicum)等小灌乔木和较为丰富的草本植物组成。此外, 在沙垄中下部和垄间低地广泛分布着藻结皮、地衣结皮和藓类结皮, 其中藓类结皮在膜果麻黄、沙拐枣等灌丛下发育尤其良好。
1.2 实验设计与样品采集为了更好的了解灌木及藓类结皮覆盖对土壤养分的影响, 在古尔班通古特沙漠腹地(45.24°N, 87.60°E)选择土壤环境相对一致且藓类结皮发育良好的垄间低地, 设置50 m×50 m大样方。选取生境和冠幅大小相近(直径50—60 cm)的膜果麻黄灌丛, 并设置裸露地对照, 在此基础上设置了有藓类结皮覆盖与无藓类结皮覆盖(裸沙)2种地表覆被类型, 具体分为裸露地藓类结皮覆盖、裸露地裸沙、灌丛下藓类结皮覆盖和灌丛下裸沙4种微生境, 每种微生境设置5个2 m×2 m的小样方, 共20个样方。使用内径5 cm的土钻在灌丛遮荫面(距离灌丛根部10—20 cm)和裸露地(远离灌丛冠幅外围至少1 m)小心采集0—2 cm和2—6 cm层土壤样品。将采取的土壤样品装进自封袋内迅速带回实验室进行样品的初步处理, 首先过2 mm筛去除大的凋落物和结皮层的藓类植物残体, 随后将样品分为2份, 一份-20 ℃保存用以测定土壤速效氮、速效磷、速效钾含量;另一部分样品自然风干后用于土壤全碳、全氮、全磷、全钾、pH、电导率等指标的测定。
1.3 土壤理化性质分析土壤样品主要测定土壤中含水量、电导率、pH、全碳、全氮、全磷、全钾、速效氮、速效磷和速效钾含量。测定土壤含水量(SWC)采用称重法, 将新鲜土壤置于105 ℃烘箱, 持续烘干48 h至恒重后称重测定。pH值采用电位法, 在1∶2.5的水土比下进行测定。电导率(EC)采用交流电导法, 在1∶5的水土比下测定[16]。通过碳氮分析仪(Multi 3100C/N, Analytik Jena AG, 德国)采用盐酸(HCL)滴定-燃烧法测定土壤有机碳(SOC)。通过全自动流动分析仪(Bran Luebbe, AA3, 德国)分别测定使用浓硫酸、高氯酸和氢氟酸消解后的土壤全氮(TN)和全磷(TP)含量以及使用0.5 mol/L NaHCO3溶液提取后的土壤速效磷(AP)含量, 全氮采用凯氏定氮法, 全磷和速效磷采用钼锑抗比色法。通过碱解扩散法测定土壤速效氮(AN)含量;采用NaOH熔融-火焰分光光度法测定土壤全钾(TK)含量;采用1 mol/L NH4OAc提取后火焰光度法测定速效钾(AK)含量。
1.4 数据分析使用Microsoft Excel对数据进行初步处理, 用SPSS 25.0软件对数据进行正态性和方差齐性检验。对灌丛、藓类结皮覆盖、土层深度及三者交互作用对土壤环境因子(SWC、pH、EC)及养分的影响进行三因素方差分析。对灌丛下与裸露地的裸沙与藓类结皮土壤的养分数据进行单因素方差分析和多重比较, 利用Origin 2021进行作图。利用R语言软件"corrplot" 包对不同土层下土壤养分与环境因子进行相关性分析。最后对不同微生境对土壤养分的贡献度进行计算, 公式如下:
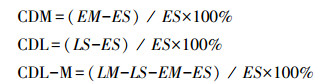
|
式中, CDM为藓类结皮贡献度, EM为裸露地藓类结皮覆盖, ES为裸露地裸沙, CDL为灌丛贡献度, LS为灌丛裸沙, CDL-M为灌丛与藓类结皮交互作用贡献度, LM为灌丛下藓类结皮覆盖
2 结果与分析 2.1 环境因子变化特征三因素方差分析表明(表 1), 灌丛、藓类结皮、土层深度对土壤含水量、pH均具有显著影响(P<0.05), 灌丛和藓类结皮对电导率影响及三者交互作用对土壤环境因子(土壤含水量、pH和电导率)的影响均不显著(P>0.05)。具体而言, 藓类结皮覆盖显著提高了土壤含水量(P<0.05), 降低了土壤pH。相对于裸露地, 灌丛下土壤含水量显著提高(P<0.05), pH和电导率变化不显著。从不同土层看, 随着土层深度的加深, 土壤含水量和pH显著上升, 而电导率则显著下降(P<0.05; 表 1;图 1)。
| 指标 Index |
灌丛 Shrub |
藓类结皮 Moss crust |
土层深度 Soil deep |
灌丛×藓类结皮 Shrub×Moss crust |
灌丛×土层深度 Shrub×Soil deep |
藓类结皮×土层深度 Moss crust×Soil deep |
灌丛×藓类结皮×土层深度 Shrub×Moss crust×Soil deep |
| SOC | 203.74** | 3.42 | 468.96** | 0.22 | 20.12** | 0.24 | 0.44 |
| TN | 77.80** | 0.13 | 189.93.** | 1.82 | 22.43** | 0.73 | 0.02 |
| TP | 4.83* | 1.68 | 13.90** | 14.97** | 3.95 | 2.15 | 3.26 |
| TK | 89.84** | 0.01 | 2.99 | 1.07 | 0.69 | 0.05 | 5.39* |
| AN | 45.37** | 6.09* | 240.86** | 62.68** | 29.36** | 2.22 | 5.85* |
| AP | 26.65** | 15.85** | 58.38** | 36.61** | 47.50** | 0.11 | 0.50 |
| AK | 146.30** | 16.23** | 14.19** | 3.03 | 0.04 | 0.07 | 3.00 |
| SWC | 43.93** | 5.42* | 247.85** | 28.27** | 0.34 | 0.34 | 0.06 |
| pH | 87.03** | 9.85** | 46.73** | 4.37* | 0.24 | 0.24 | 1.68 |
| EC | 2.83 | 0.30 | 160.29** | 0.03 | 1.10 | 0.11 | 0.34 |
| SOC:有机碳Organic carbon;TN:全氮Total nitrogen;TP:全磷Total phosphorus;TK:全钾Total potassium;AN:速效氮Available phosphorus;AP:速效磷Available phosphorus;AK:速效钾Available potassium;SWC:土壤含水量Soil water content;EC:电导率Electrical conductivity;* P<0.05, ** P<0.01 | |||||||

|
| 图 1 不同微生境下藓类结皮覆盖对土壤环境因子的影响(平均值±标准误, n=5) Fig. 1 Effects of moss crust coverage on soil environmental factors in different microhabitats (mean±standard error, n=5) E:裸露地Exposed areas;L:灌丛Shrub;S:裸沙Bare sand;M:藓类结皮覆盖Moss crust cover;不同大写字母和小写字母分别表示不同微生境间差异显著, *和**分别表示同一微生境不同土层存在显著差异(P<0.05)和极显著差异(P<0.01) |
就全量养分而言, 灌丛、土层深度、灌丛与土层深度的交互作用对有机碳和全氮均具有极显著影响(P<0.01), 灌丛、土层深度、灌丛与藓类结皮的交互作用对全磷具有显著影响(P<0.05), 灌丛、藓类结皮与土层深度三者的交互作用对有机碳、全氮和全磷影响不显著(P>0.05), 灌丛及灌丛、藓类结皮与土层深度三者的交互作用对全钾具有显著影响(P<0.05; 表 1)。具体而言, 藓类结皮覆盖显著提高了裸露地2—6 cm全氮、全磷含量(P<0.05), 而对有机碳、全钾影响不显著(图 2)。相较于裸露地, 灌丛显著提高了有机碳、全氮、全钾含量(P<0.05), 而降低了灌丛下藓类结皮覆盖全磷含量(图 2)。从不同土层来看, 随着土层深度的加深, 有机碳、全氮、全磷含量显著下降, 而对全钾的影响不显著(图 2)。

|
| 图 2 不同微生境下藓类结皮覆盖对土壤全量养分的影响(平均值±标准误, n=5) Fig. 2 Effects of moss crust coverage on soil total nutrient in different microhabitats (mean±standard error, n=5) |
由表 1可知, 灌丛、藓类结皮、土层深度对速效氮、速效磷和速效钾均具有显著影响, 三者交互作用对速效氮具有显著影响(P<0.05), 而对速效磷和速效钾的影响不显著(P>0.05)。具体而言, 藓类结皮覆盖提高了裸露地速效磷和速效钾的含量, 显著降低了速效氮含量(P<0.05);相对于裸露地, 藓类结皮覆盖显著提高了灌丛下2—6 cm速效氮、速效磷和速效钾的含量(P<0.05)。从不同土层看, 随着土层深度的加深, 灌丛下速效磷显著上升, 而速效氮和速效钾则显著下降(P<0.05; 图 3)。

|
| 图 3 不同微生境下藓类结皮覆盖对土壤速效养分的影响(平均值±标准误, n=5) Fig. 3 Effects of moss crust coverage on soil available nutrient in different microhabitats (mean±standard error, n=5) |
Pearson相关性分析表明(图 4), 不同土层养分(C、N、P、K)与环境因子(土壤含水量、pH、电导率)之间的关联程度不同。其中, 在0—2 cm土壤中, 有机碳、全氮、全钾、速效氮、速效钾与土壤含水量显著正相关, 而速效磷与土壤含水量显著负相关(P<0.05), 全磷与土壤含水量相关性不显著(P>0.05);有机碳、全氮、全钾和速效钾与pH显著正相关;有机碳、速效钾与电导率显著正相关(P<0.05)。在2—6 cm土壤中有机碳、速效氮和速效钾与土壤含水量显著正相关;有机碳、全钾、速效磷和速效钾与pH显著正相关;速效钾与电导率显著正相关(P<0.05)。随着土层深度下降, 土壤含水量与养分指标的相关性明显减弱, pH和电导率与各指标的相关性也有较大变化。

|
| 图 4 不同土层养分与环境因子间的相关性热图 Fig. 4 Heat map of correlation between nutrients and environmental factors in different soil layers SOC:有机碳Organic carbon;TN:全氮Total nitrogen;TP:全磷Total phosphorus;TK:全钾Total potassium;AN:速效氮Available phosphorus;AP:速效磷Available phosphorus;AK:速效钾Available potassium;SWC:土壤含水量Soil water content;EC:电导率Electrical conductivity;*P<0.05, **P<0.01, **P<0.001 |
贡献度分析结果表明, 对养分的贡献度整体呈现灌丛>藓类结皮>藓类结皮与灌丛交互作用的态势。其中, 藓类结皮覆盖对土壤含水量和速效氮的贡献为负效应, 分别为-18.98%和-28.46%;对有机碳、全氮和全磷的贡献为显著正效应(P<0.05), 分别为13.30%、6.16%、5.10%;对电导率、全钾、速效磷和速效钾贡献不显著(P>0.05)。灌丛对所有养分指标均表现为正贡献效应, 其中对pH、有机碳、全氮、全磷、全钾、速效磷和速效钾的贡献为显著正效应(P<0.05), 分别为5.41%、55.63%、67.65%、13.90%、4.87%、79.85%和60.03%, 而对土壤含水量、电导率和速效氮贡献效应不显著(P>0.05)。藓类结皮覆盖与灌丛的交互作用对土壤含水量和速效氮表现为显著正贡献效应分别为50.29%和79.23%, 对全氮和全磷的贡献为显著负效应(P<0.05), 分别为-16.92%和-15.03%, 而对pH、电导率、有机碳、全钾、速效磷和速效钾的贡献效应不显著(P>0.05)。

|
| 图 5 灌丛和藓类结皮及二者交互作用对土壤属性的贡献度分析 Fig. 5 Contribution of shrub and moss crusts and their interaction to soil properties 虚线为贡献度为0的界限, 左侧为负效应, 右侧为正效应; *和**分别表示贡献度相对于0的显著差异(P<0.05)和极显著差异(P<0.01) |
藓类结皮可以在荒漠地表形成一层粗糙的保护层, 由假根缠绕土壤表面颗粒, 创造一个稳定的土壤环境, 从而改善土壤的结构、水分和养分等条件[17]。本研究发现, 相较于裸沙, 裸露地藓类结皮覆盖增加了0—2 cm土壤含水量而降低了2—6 cm土壤含水量, 这表明与裸沙相比, 藓类结皮具有更强的持水能力, 但渗透性较低, 这与先前的研究结果保持一致[18—19]。可见, 藓类结皮在控制干旱区深层土壤湿度中可能具有重要作用[20]。土壤pH和电导率条件也因藓类结皮的覆盖而显著改善, 这得益于藓类结皮土壤相比于裸沙具有更高的风成积盐能力和阴阳离子交换能力[21]。此外, 本研究发现相较于裸沙, 藓类结皮覆盖显著降低了2—6 cm电导率。这主要是由于其增强了土壤团聚体的稳定性, 提高了微生物活性, 从而降低了土壤电导率。然而, 土壤养分与环境因子密切相关, 而藓类结皮对土壤环境因子的改善势必会影响到养分的动态变化。
在古尔班通古特沙漠膜果麻黄灌丛内外, 藓类结皮的覆盖显著促进提升了土壤碳、氮和钾养分的含量和有效性。这与过去关于生物土壤结皮对土壤养分存在积极影响的结果保持一致[22—23]。藓类结皮是干旱区、半干旱区重要的生物类群, 可以富集土壤养分, 调节了土壤养分的分解和矿化速率, 进而影响土壤的初级生产力和养分有效性[14]。同时, 藓类结皮土壤中含有丰富的固氮微生物, 能够固定大气中的N2转换为可被植物利用的无机氮, 提高土壤的氮含量[24]。此外, 藓类覆盖可以显著提高土壤微生物群落多样性和丰富度, 促进土壤养分的循环过程, 提高了土壤氮、磷和钾养分含量及有效性[25—26]。研究表明, 在荒漠生态系统中, 藓类结皮对于土壤理化性质的改善主要集中在表层土壤中, 而改善程度与是否有植被覆盖密切相关[27]。这可能是灌丛对于养分的贡献大于裸露地的主要原因。研究还发现, 裸露地藓类结皮覆盖显著提升了全磷和速效磷的含量, 然而灌丛下藓类结皮覆盖显著降低了全磷和速效磷的含量。这主要是由于以藓类结皮为主的生物土壤结皮在呼吸过程中能够通过分泌大量有机酸、螯合剂、H+等影响土壤磷的有效性, 从而使更多矿物结合磷转向可利用性磷[25], 促进灌丛对磷的吸收利用, 降低了土壤中磷的含量。然而, 相较于碳、氮和钾, 速效磷在2—6 cm土壤中的含量明显高于在0—2 cm土壤中的含量, 这主要是由于磷的移动性较差, 即使被微生物、根系分泌物活化后短时间内也难以迁移到土壤表层, 因此导致磷主要集中在下层土壤中。另外, 相较于维管植物, 藓类植物在表层土壤养分的获取上具有较大优势, 因其能够产生耐分解的凋落物, 降低了其矿化速率和凋落物的分解率, 减少到达维管植物根际周围的养分[28], 这可能是藓类结皮覆盖对速效氮产生负贡献效应的主要原因。
3.2 灌丛对土壤养分的影响灌丛是荒漠中重要的植物群落, 在干旱区, 灌木要比乔木和草本植物具有更强的适应性, 防风阻沙效果也更具优势[29]。在干旱半干旱地区, 灌木群落通过植被-土壤反馈机制改善了土壤温度、湿度、pH以及土壤结构, 提高了土壤养分积累和氮矿化速率, 对土壤生物(原生动物、微生物等)和隐花植物(地衣、苔藓等)的生存具有积极影响[30]。研究表明, 灌丛定植后可以增加土壤表层有机质的积累, 但也积累了大量的盐分, 使pH升高, 导致旱地土壤盐碱化加剧[31], 这在本研究中也得以证实。土壤pH在驱动土壤养分循环过程中具有重要意义, 可直接或间接的影响土壤质量状况, 并且可以通过控制特定微生物类群, 从而驱动土壤养分循环[32]。
水分是限制干旱和半干旱地区土壤养分状况的关键因素, 而适宜的水分能提高凋落物的分解速率和微生物活性, 从而增加土壤养分含量。本研究结果表明, 灌丛对于土壤养分的贡献大于藓类结皮覆盖。导致这一结果的主要原因是由于灌丛在生长发育过程中对其自身凋落物和降尘的不断截获和沉积, 使枯落物和有机质在灌丛下累积, 形成明显的“肥岛效应”。先前对于不同地区不同种类的灌丛下土壤研究发现, 相较于裸露地, 灌丛具有更强的资源富集性[33—34], 而灌丛死亡后富集效应逐渐降低。此外, 灌丛根系的生长有利于增大土壤孔隙, 使得养分的转化和迁移变的更加容易, 而根系分泌物也有助于提高土壤微生物和土壤酶的活性[26], 这也是灌丛下土壤碳、氮、磷、钾含量比裸露地高的主要原因。可见, 灌丛的形成及其对养分的富集作用改变了土壤养分的空间分配, 减缓了植物对于养分需求的限制作用, 提高了植物对养分的利用效率[35]。此外, 研究表明, 灌丛对于养分的富集效应主要集中在表层土壤[36]。并且王湘等[37]对柴达木盆地白刺灌丛养分富集特征研究也发现, 白刺灌丛对于养分的富集效应主要发生在0—10 cm。除磷外, 其余土壤养分均随土层深度的增加呈减少趋势, 这与本文得出的结果相似, 因为灌丛广泛的根系可以将土壤养分从深层转移到浅层, 降低了深层土壤养分的含量。
3.3 灌丛与生物土壤结皮的交互效应对土壤养分的影响在干燥的荒漠生态系统中, 灌丛的定植为藓类结皮在冠下的发育提供了稳定的微环境, 而藓类结皮形成的表层则会影响到水分的入渗与保留[38]。藓类结皮和维管植物之间的共生或竞争关系在有限土壤水资源的利用中具有重要的意义[39]。本文结果表明, 灌丛与藓类结皮的交互作用对土壤含水量的贡献呈现显著正效应。这可能是由于藓类结皮将水分截流在表层, 而灌丛根系对于灌丛下藓类结皮的扰动和植物根系促进水分在根系通道的优先流动机制, 提高水分向土壤的入渗量和深度, 并降低蒸发速率[40—41]。此外, 水分也是改善土壤养分矿化潜力的重要影响因素。
灌丛与藓类结皮是干旱区、半干旱区有机质的主要贡献者。本文结果表明, 其单一对于土壤全量氮磷的贡献基本呈现为正效应, 对速效氮为负效应, 二者的交互效应则呈现相反的结果。这可能是由于在灌丛与藓类结皮共生的微环境中, 灌丛与藓类植物根系会改善土壤团聚体结构, 提升了土壤有效孔隙及气体交换, 增加土壤微生物和脲酶活性, 从而间接促进了有机氮的净矿化速率[42]。并且, 灌丛根系与藓类植物促进了土壤微生物活性而暂时增加了微生物对土壤中碳、氮和磷的消耗, 并进一步驱动土壤微生物与植物生长发育之间产生养分竞争, 而藓类植物在竞争中比维管植物更具有优势, 使更多营养物质被植物所吸收利用, 从而对全量养分造成负面影响。另外, 苔藓植物作为富磷植物, 自身偏好对磷的吸收[43], 在灌丛与藓类结皮共生的微环境中, 维管植物、藓类植物以及微生物对磷素的竞争可能抑制了磷元素对养分的归还, 这也可能是对土壤磷含量及有效性造成负面影响的重要原因。
藓类结皮与荒漠灌木对土壤养分的影响在不同的研究区已经做出大量研究, 本文的结果同样证实了二者作为荒漠生态系统的主要植被类群的重要地位。虽然从养分贡献这一单一的角度看, 灌木比藓类具有更强的富集能力, 但这并不代表藓类结皮的作用不强。在荒漠流沙表面, 藓类结皮是首先定植的先锋植物, 为土壤的稳定和后续维管植物的生长打下了基础。可见, 在荒漠化修复的过程中, 藓类结皮与灌木的选择应该在生态系统不同的植被演替阶段进行充分考虑。值得注意的是, 本文的研究证实, 二者的交互作用对土壤水分与养分的影响并未呈现叠加的效果, 甚至与单一类群作用下呈现相反的趋势。这强调了在后续的研究中, 考虑物种间相互影响的重要性, 并且为野外调查与样品采集工作的微生境选择提供一定的数据参考。
4 结论在古尔班通古特沙漠, 藓类结皮与灌丛镶嵌分布所形成的多种微生境, 在一定程度上塑造了表层土壤养分的空间异质性格局。结果表明, 在单一类群下(灌丛与结皮独立存在), 灌丛的存在与藓类结皮的覆盖差异性促进了土壤碳、氮、磷和钾相关养分的积累, 其中以灌丛对养分的42.54%贡献度远大于藓类结皮的2.43%贡献度。而在灌丛与藓类结皮共存的微生境中, 二者的交互作用对土壤养分则呈现负效应(-1.91%), 特别是显著抑制了全氮和全磷养分含量。值得注意的是, 灌丛与藓类结皮的交互作用极大的促进了土壤的氮矿化, 但在二者的单一群落中却显示为抑制或无影响, 这可能与土壤水分的变化密切相关。此外, 仅2—6 cm的土层深度下降便显著降低了土壤含水量与碳、氮、磷和钾各养分指标的关联度, 且与pH的相关性也发生巨大变化。本研究定量化的阐明了荒漠藓类结皮与灌丛这两大类群对于表层各土壤养分指标的贡献程度, 并且再次强调了水分在影响荒漠生态系统土壤微生物活性及养分循环方面的重要意义。
| [1] |
Aguiar M R, Sala O E. Patch structure, dynamics and implications for the functioning of arid ecosystems. Trends in Ecology & Evolution, 1999, 14(7): 273-277. |
| [2] |
张珂, 苏永中, 王婷, 刘婷娜. 荒漠绿洲区不同种植年限人工梭梭林土壤化学计量特征. 生态学报, 2016, 36(11): 3235-3243. |
| [3] |
Delgado-Baquerizo M, Maestre F T, Gallardo A, Bowker M A, Wallenstein M D, Quero J L, Ochoa V, Gozalo B, García-Gómez M, Soliveres S, García-Palacios P, Berdugo M, Valencia E, Escolar C, Arredondo T, Barraza-Zepeda C, Bran D, Carreira J A, Chaieb M, Conceição A A, Derak M, Eldridge D J, Escudero A, Espinosa C I, Gaitán J, Gatica M G, Gómez-González S, Guzman E, Gutiérrez J R, Florentino A, Hepper E, Hernández R M, Huber-Sannwald E, Jankju M, Liu J S, Mau R L, Miriti M, Monerris J, Naseri K, Noumi Z, Polo V, Prina A, Pucheta E, Ramírez E, Ramírez-Collantes D A, Romão R, Tighe M, Torres D, Torres-Díaz C, Ungar E D, Val J, Wamiti W, Wang D L, Zaady E. Decoupling of soil nutrient cycles as a function of aridity in global drylands. Nature, 2013, 502(7473): 672-676. DOI:10.1038/nature12670 |
| [4] |
刘进辉, 王雪芹, 马洋. 沙漠绿洲过渡带柽柳灌丛沙堆-丘间地系统土壤养分空间异质性. 生态学报, 2016, 36(4): 979-990. |
| [5] |
Zhang Y M, Chen J, Wang L, Wang X Q, Gu Z H. The spatial distribution patterns of biological soil crusts in the Gurbantunggut Desert, Northern Xinjiang, China. Journal of Arid Environments, 2007, 68(4): 599-610. DOI:10.1016/j.jaridenv.2006.06.012 |
| [6] |
Bowker M A, Reed S C, Maestre F T, Eldridge D J. Biocrusts: the living skin of the earth. Plant and Soil, 2018, 429(1): 1-7. |
| [7] |
Weber B, Belnap J, Büdel B, Antoninka A J, Barger N N, Chaudhary V B, Darrouzet-Nardi A, Eldridge D J, Faist A M, Ferrenberg S, Havrilla C A, Huber-Sannwald E, Malam Issa O, Maestre F T, Reed S C, Rodriguez-Caballero E, Tucker C, Young K E, Zhang Y M, Zhao Y G, Zhou X B, Bowker M A. What is a biocrust? A refined, contemporary definition for a broadening research community. Biological Reviews, 2022, 97(5): 1768-1785. DOI:10.1111/brv.12862 |
| [8] |
郭琦, 卜崇峰, 李宜坪, 鞠孟辰, 李亚红, 韦应欣. 区域尺度生物结皮下伏土壤养分的空间分布特征——以毛乌素沙地为例. 土壤学报, 2022, 59(3): 699-707. |
| [9] |
Delgado-Baquerizo M, Morillas L, Maestre F T, Gallardo A. Biocrusts control the nitrogen dynamics and microbial functional diversity of semi-arid soils in response to nutrient additions. Plant and Soil, 2013, 372(1): 643-654. |
| [10] |
Zhao Y, Xu M, Belnap J. Potential nitrogen fixation activity of different aged biological soil crusts from rehabilitated grasslands of the hilly Loess Plateau, China. Journal of Arid Environments, 2010, 74(10): 1186-1191. DOI:10.1016/j.jaridenv.2010.04.006 |
| [11] |
Bowker M A, Belnap J, Davidson D W, Phillips S L. Evidence for micronutrient limitation of biological soil crusts: importance to arid-lands restoration. Ecological Applications, 2005, 15(6): 1941-1951. DOI:10.1890/04-1959 |
| [12] |
秦宁强, 赵允格. 生物土壤结皮对雨滴动能的响应及削减作用. 应用生态学报, 2011, 22(9): 2259-2264. |
| [13] |
Xiao B, Sun F H, Hu K L, Kidron G J. Biocrusts reduce surface soil infiltrability and impede soil water infiltration under tension and ponding conditions in dryland ecosystem. Journal of Hydrology, 2019, 568: 792-802. DOI:10.1016/j.jhydrol.2018.11.051 |
| [14] |
Chamizo S, Cantón Y, Miralles I, Domingo F. Biological soil crust development affects physicochemical characteristics of soil surface in semiarid ecosystems. Soil Biology and Biochemistry, 2012, 49: 96-105. DOI:10.1016/j.soilbio.2012.02.017 |
| [15] |
周宏飞, 李彦, 汤英, 周宝佳, 徐宏伟. 古尔班通古特沙漠的积雪及雪融水储存特征. 干旱区研究, 2009, 26(3): 312-317. |
| [16] |
Bhattacharyya R, Rabbi S M F, Zhang Y Q, Young I M, Jones A R, Dennis P G, Menzies N W, Kopittke P M, Dalal R C. Soil organic carbon is significantly associated with the pore geometry, microbial diversity and enzyme activity of the macro-aggregates under different land uses. Science of the Total Environment, 2021, 778: 146286. DOI:10.1016/j.scitotenv.2021.146286 |
| [17] |
Nevins C J, Strauss S L, Inglett P W. Biological soil crusts enhance moisture and nutrients in the upper rooting zone of sandy soil agroecosystems. Journal of Plant Nutrition and Soil Science, 2020, 183(5): 615-626. DOI:10.1002/jpln.202000218 |
| [18] |
Sun F H, Xiao B, Kidron G J. Towards the influences of three types of biocrusts on soil water in drylands: insights from horizontal infiltration and soil water retention. Geoderma, 2022, 428: 116136. DOI:10.1016/j.geoderma.2022.116136 |
| [19] |
岳艳鹏, 成龙, 孙迎涛, 庞营军, 吴波, 石麟, 何金军, 贾晓红. 毛乌素沙地生物结皮覆盖区土壤水分收支变化特征. 应用生态学报, 2022, 33(7): 1861-1870. |
| [20] |
Xiao B, Hu K L. Moss-dominated biocrusts decrease soil moisture and result in the degradation of artificially planted shrubs under semiarid climate. Geoderma, 2017, 291: 47-54. DOI:10.1016/j.geoderma.2017.01.009 |
| [21] |
Chen X H, Duan Z H. Impacts of soil crusts on soil physicochemical characteristics in different rainfall zones of the arid and semi-arid desert regions of Northern China. Environmental Earth Sciences, 2015, 73(7): 3335-3347. DOI:10.1007/s12665-014-3622-x |
| [22] |
Bowker M A, Mau R L, Maestre F T, Escolar C, Castillo-Monroy A P. Functional profiles reveal unique ecological roles of various biological soil crust organisms. Functional Ecology, 2011, 25(4): 787-795. DOI:10.1111/j.1365-2435.2011.01835.x |
| [23] |
Delgado-Baquerizo M, Maestre F T, Eldridge D J, Bowker M A, Ochoa V, Gozalo B, Berdugo M, Val J, Singh B K. Biocrust-forming mosses mitigate the negative impacts of increasing aridity on ecosystem multifunctionality in drylands. The New Phytologist, 2016, 209(4): 1540-1552. DOI:10.1111/nph.13688 |
| [24] |
Elbert W, Weber B, Burrows S, Steinkamp J, Büdel B, Andreae M O, Pöschl U. Contribution of cryptogamic covers to the global cycles of carbon and nitrogen. Nature Geoscience, 2012, 5(7): 459-462. DOI:10.1038/ngeo1486 |
| [25] |
Baumann K, Siebers M, Kruse J, Eckhardt K U, Hu Y F, Michalik D, Siebers N, Kar G, Karsten U, Leinweber P. Biological soil crusts as key player in biogeochemical P cycling during pedogenesis of sandy substrate. Geoderma, 2019, 338: 145-158. DOI:10.1016/j.geoderma.2018.11.034 |
| [26] |
Zhao K, Zhang B C, Li J N, Li B, Wu Z F. The autotrophic community across developmental stages of biocrusts in the Gurbantunggut Desert. Geoderma, 2021, 388: 114927. DOI:10.1016/j.geoderma.2021.114927 |
| [27] |
周小泉, 刘政鸿, 杨永胜, 苑森朋. 毛乌素沙地三种植被下苔藓结皮的土壤理化效应. 水土保持研究, 2014, 21(6): 340-344. |
| [28] |
卜兆君, 陈旭, 姜丽红, 李鸿凯, 赵红艳. 苔藓植物相互作用的研究进展. 应用生态学报, 2009, 20(2): 460-466. |
| [29] |
唐艳, 刘连友, 屈志强, 胡霞, 郭兰兰, 吕艳丽, 曹恒武, 贾振杰, 杨岩岩. 植物阻沙能力研究进展. 中国沙漠, 2011, 31(1): 43-48. |
| [30] |
Ward D, Trinogga J, Wiegand K, du Toit J, Okubamichael D, Reinsch S, Schleicher J. Large shrubs increase soil nutrients in a semi-arid savanna. Geoderma, 2018, 310: 153-162. DOI:10.1016/j.geoderma.2017.09.023 |
| [31] |
Lu J N, Feng S, Wang S K, Zhang B L, Ning Z Y, Wang R X, Chen X P, Yu L L, Zhao H S, Lan D M, Zhao X Y. Patterns and driving mechanism of soil organic carbon, nitrogen, and phosphorus stoichiometry across Northern China's Desert-grassland transition zone. CATENA, 2023, 220: 106695. DOI:10.1016/j.catena.2022.106695 |
| [32] |
Delgado-Baquerizo M, Eldridge D J, Ochoa V, Gozalo B, Singh B K, Maestre F T. Soil microbial communities drive the resistance of ecosystem multifunctionality to global change in drylands across the globe. Ecology Letters, 2017, 20(10): 1295-1305. DOI:10.1111/ele.12826 |
| [33] |
Zaady E. Seasonal change and nitrogen cycling in a patchy Negev Desert: a review. Arid Land Research and Management, 2005, 19(2): 111-124. DOI:10.1080/15324980590916512 |
| [34] |
李艳琼, 邓湘雯, 易昌晏, 邓东华, 黄志宏, 项文化, 方晰, 景宜然. 湘西南喀斯特地区灌丛生态系统植物和土壤养分特征. 应用生态学报, 2016, 27(4): 1015-1023. |
| [35] |
Zhang J H, Tang Z Y, Wang W T, Zhang H F, Liu Y, Xin Y H, Zhao L J, Li H. Nutrient resorption responses of plant life forms to nitrogen addition in temperate shrublands. Ecosphere, 2022, 13(6): 1-12. |
| [36] |
何玉惠, 刘新平, 谢忠奎. 红砂灌丛对土壤和草本植物特征的影响. 生态学杂志, 2011, 30(11): 2432-2436. |
| [37] |
王湘, 焦菊英, 曹雪, 李建军, 白雷超, 孙晓晨. 柴达木盆地尕海湖区白刺灌丛沙堆剖面土壤养分的分布和富集特征. 应用生态学报, 2022, 33(3): 765-774. |
| [38] |
Chamizo S, Adessi A, Mugnai G, Simiani A, De Philippis R. Soil type and cyanobacteria species influence the macromolecular and chemical characteristics of the polysaccharidic matrix in induced biocrusts. Microbial Ecology, 2019, 78(2): 482-493. DOI:10.1007/s00248-018-1305-y |
| [39] |
Li X, Tian F, Jia R, Zhang Z, Liu L. Do biological soil crusts determine vegetation changes in sandy deserts? Implications for managing artificial vegetation. Hydrological Processes, 2010, 24(25): 3621-3630. DOI:10.1002/hyp.7791 |
| [40] |
Li S Z, Xiao H L, Cheng G D, Luo F, Liu L C. Mechanical disturbance of microbiotic crusts affects ecohydrological processes in a region of revegetation-fixed sand dunes. Arid Land Research and Management, 2006, 20(1): 61-77. DOI:10.1080/15324980500369467 |
| [41] |
Amir M M, Ilan S, Shlomo S, Hiam A G, Shimshon S, Eli Z. The dynamic of the eco-hydrological interrelations between shrubs and biocrusts in the Negev shrublands: Empiric assessments and perspectives for shrubland rehabilitation. CATENA, 2022, 214: 106296. |
| [42] |
Arróniz-Crespo M, Leake J R, Horton P, Phoenix G K. Bryophyte physiological responses to, and recovery from, long-term nitrogen deposition and phosphorus fertilisation in acidic grassland. The New Phytologist, 2008, 180(4): 864-874. |
| [43] |
郝占庆, 叶吉, 姜萍, 蔺菲. 长白山暗针叶林苔藓植物在养分循环中的作用. 应用生态学报, 2005, 16(12): 2263-2266. |