 2024, Vol. 44
2024, Vol. 44文章信息
- 寇钊阳, 李春越, 肖凤娇, 常顺, 王益, 党廷辉
- KOU Zhaoyang, LI Chunyue, XIAO Fengjiao, CHANG Shun, WANG Yi, DANG Tinghui
- 氮磷添加对黄土旱塬农田土壤线虫群落及能量结构的影响
- Effects of nitrogen and phosphorus addition on soil nematode communities and energy structure in Loess Plateau farmland
- 生态学报. 2024, 44(5): 1962-1971
- Acta Ecologica Sinica. 2024, 44(5): 1962-1971
- http://dx.doi.org/10.20103/j.stxb.202304210825
-
文章历史
- 收稿日期: 2023-04-21
- 网络出版日期: 2023-10-10

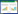




2. 中国科学院地球环境研究所, 西安 710061;
3. 西北农林科技大学水土保持研究所, 杨凌 712100
2. Institute of Earth Environment, Chinese Academy of Sciences, Xi'an 710061, China;
3. Institute of Soil and Water Conservation, Northwest Agricultural and Forestry University, Yangling 712100, China
土壤线虫由形态和功能多样化的群体组成[1], 其种类与数量非常丰富, 在地下食物网中占据多个生态位, 是陆地生态系统中主要的功能性种群之一[2], 在维持土壤生态系统稳定及促进物质循环和能量流动等方面发挥着重要作用[3—4]。线虫生态指数提供了线虫群落结构多样性和功能属性的分析, 从而为土壤生态系统的健康和功能提供了有价值的信息[5—6], 而代谢足迹则通过量化土壤线虫的碳分配模式, 使学者对线虫的能量利用、生长和呼吸过程的了解更加深入[4, 7]。这些指标的运用使我们能够全面掌握线虫在土壤生态系统中的作用, 为土壤生态系统的研究和管理提供了有力支持。
以前的研究已经强调了土壤线虫对养分添加的敏感反应[8—9]。一项meta分析[10]探究了氮富集对陆地生态系统土壤线虫群落的影响, 研究表明低氮添加增强了线虫的多样性, 而高氮添加则降低了线虫的多样性。另一项研究指出[11], 与施氮肥相比, 土壤线虫丰度对磷肥的响应并不明显, 由于土壤养分的增加改变了有机质的分解途径, 使其更偏向于食细菌线虫的分解, 从而减少了稀有物种线虫的数量。此外, 在对中国东北部地区进行的长期施肥农田实验中发现[12], 土壤中食细菌和真菌线虫随着土壤有机碳和全氮含量的增加而增加, 进而影响其代谢足迹, 值得注意的是, 这种环境变化对线虫代谢足迹的影响大于其丰度, 这些研究结果表明了土壤线虫对养分添加表现出不同的反应。
土壤微食物网内的能量流动代表了能量从初级生产者(植物和微生物)向更高营养级(捕食者和分解者)的转移[13], 了解能量流的动态对于评估土壤生态系统的功能和稳定性至关重要。营养物质的添加可以通过改变线虫营养群的丰度和组成来调节微食物网中的能量流动途径[14]。然而, 先前的研究中土壤微食物网能量结构对农业经营的响应在很大程度上仍然未知[10, 15]。因此, 本研究以黄土区长期氮磷添加的农田土壤为研究对象, 通过提取土壤中的线虫, 计算线虫各营养群间的代谢足迹和能量通量, 并与土壤理化性质进行相关性分析, 旨在提升我们对土壤生态系统动态的理解, 以实现农业管理的可持续性。
1 材料和方法 1.1 研究区概况本研究实验土壤取自黄土高原中南部的中国科学院长武生态试验站(35°12′N, 107°40′E), 该试验站海拔1200 m, 气候属于暖温带半湿润大陆性季风气候, 年均降水量584.1 mm, 无霜期171 d, 年均气温9.1℃, 属于典型的旱作农业区。该地区土壤类型为黑垆土, 冬小麦是主要的农作物。1984年布设试验时耕层土壤有机质含量6.50 g/kg, 全氮含量0.80 g/kg, pH为8.10。
1.2 试验设计长期定位施肥试验开始于1984年, 每个试验小区面积为4 m×8 m, 中间设置1 m的缓冲带。设6种施肥处理, 分别为:不施肥(CK);施N12肥, 氮肥量60 g m-2 a-1;施N24肥, 氮肥量120 g m-2 a-1;施P12肥, 磷肥量60 g m-2 a-1;施P24肥, 磷肥量120 g m-2 a-1;施N12P12肥, 氮肥和磷肥量均为60 g m-2 a-1。氮肥为尿素, 含氮量46.4%;磷肥为过磷酸钙, 含P2O5 46.0%。所有肥料在播种(2019年9月10日)前一次性撒入地表, 然后翻耕入土。
样品采集时间为2020年6月, 选择五点采样法进行一次性采样, 使用1 m取土钻采集0—20 cm处耕层土壤。将所采集土壤样品做好密封标记工作后, 及时带回实验室进行处理。剔除土壤中动物残体、碎小石子、植物枯枝、杂草及其他杂质。土壤过2 mm筛后, 取出约1/3土壤进行风干处理, 用于测定土壤全量指标;剩余新鲜土样在4 ℃冰箱保存, 进行无机氮, 速效磷和线虫的测定。
1.3 土壤理化性质的测定土壤理化性质测定参考《土壤农化分析》[16]。含水率(SMC)采用烘干法测定;pH采用PHS-3C型酸度计进行测定;全碳(TC)采用元素分析仪(Vario TOC, Elementar, Hanau, 德国)测定;全磷(TP)采用浓硫酸-高氯酸消煮钼锑抗比色法测定;速效磷(AP)采用碳酸氢钠浸提钼锑抗比色法测定;全氮(TN)采用凯氏定氮仪测定;硝态氮(NO3--N)和铵态氮(NH4+-N)采用硫酸钾溶液浸提-流动分析仪(Autoanalyzer 3, Bran-Luebbe, 德国)测定, 土壤理化性质测定结果见表 1。
| 处理 Treatments |
pH | 含水率 SMC/% |
全氮 TN/(g/kg) |
全磷 TP/(g/kg) |
全碳 TC/(g/kg) |
速效磷 AP/(mg/kg) |
铵态氮 NH4+-N/(mg/kg) |
硝态氮 NO3--N/(mg/kg) |
| CK | 8.22±0.03a | 19.95±0.14b | 0.85±0.02a | 0.73±0.00d | 17.49±0.28a | 7.37±0.46d | 0.29±0.04a | 11.13±0.08d |
| N12 | 8.20±0.06a | 16.78±0.21e | 0.72±0.06b | 0.67±0.01e | 16.85±0.43a | 2.53±0.15e | 0.32±0.04a | 13.50±0.29b |
| N24 | 7.92±0.02b | 18.03±0.04d | 0.90±0.05a | 0.69±0.02e | 16.00±0.49b | 4.30±0.70e | 0.23±0.04a | 25.11±0.18a |
| P12 | 8.15±0.09a | 18.97±0.12c | 0.85±0.02a | 1.06±0.01b | 17.04±0.28a | 42.80±0.69b | 0.22±0.01a | 11.13±0.11d |
| P24 | 8.23±0.05a | 15.66±0.07f | 0.76±0.01b | 1.32±0.02a | 16.17±0.04b | 66.77±2.55a | 0.38±0.18a | 10.88±0.04d |
| N12P12 | 8.22±0.05a | 20.59±0.06a | 0.75±0.05b | 0.96±0.04c | 16.96±0.28a | 23.27±1.89c | 0.33±0.10a | 11.46±0.12c |
| CK: 不施肥No fertilization;N: 氮肥Nitrogen fertilizer;P: 磷肥Phosphorus fertilizer;下标12, 24:施肥量分别为60和120 g m-2 a-1 The amount of fertilizer was 60 and 120 g m-2 a-1;SMC: 含水率Soil moisture content;TN: 全氮Total nitrogen;TP: 全磷Total phosphorus;TC: 全碳Total carbon;AP: 速效磷Available phosphorus;NH4+-N: 铵态氮Ammonium nitrogen;NO3--N: 硝态氮Nitrate nitrogen;同列不同字母表示不同处理间差异显著(P<0.05) | ||||||||
取50 g新鲜土样, 采用贝尔曼浅盘法[17]对土壤线虫进行分离提取并放置于解剖镜下计数, 最终折算成100 g干土中土壤线虫的数量。参照《中国土壤动物检索图鉴》[18]和Bongers[19]的分类图鉴对土壤线虫鉴定到属。根据线虫的形态学特征将其划分为4个营养类群[20]:食细菌类线虫(Ba), 食真菌类线虫(Fu), 捕食/杂食类线虫(Op)和植物寄生类线虫(Pp), 同时按照生活史策略把线虫划分成r策略者向k策略者过渡的5个类群, 记录相应的cp值[3]。
1.5 线虫生态指数计算Shannon多样性指数[21]:H=-∑Pi×ln(Pi);均匀度指数:J=H/ln(S);丰富度指数:SR=(S-1)lnN;线虫通路比值:NCR=B/(B+F)。公式中Pi为第i个分类单元中个体所占比例, S为鉴定分类单元的数量, N为鉴定线虫的个体数量, B为食细菌线虫数量, F为食真菌线虫数量[22]。成熟度指数:MI=∑vifi;植物寄生线虫成熟度指数:PPI=∑vif′i。其中vi为自由生活线虫按照生活史策略赋予的值(cp), fi和f′i分别为自由生活线虫和植食性线虫的科或属在总线虫中所占的比例[3, 23]。
富集指数:EI=100×[e/(e+b)], 用于评估食物网对可利用资源的响应。结构指数:SI=100×[s/(s+b)], 指示在干扰(胁迫)或生态恢复过程中土壤食物网结构的变化。式中e代表食物网中的富集成分, b代表基础成分, s代表结构成分[24]。
1.6 土壤线虫代谢足迹和能量结构 1.6.1 线虫代谢足迹的计算:利用 http://plpnemweb.ucdavis.edu/nemaplex/Ecology/nematode_weights.htm的公开数据计算线虫新鲜生物量(W, μg/100 g干土)和代谢足迹(NMF, μg/100 g)[4]。线虫的干重为鲜重的20%, 体内生物量碳的比例为干重的52%。计算公式如下:
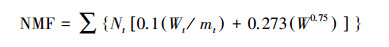
|
式中, Nt是指第t个属的个体数量, Wt和mt分别指第t个属的个体生物量和cp值。
1.6.2 线虫群落的能量通量计算:线虫的能量通量(F, μg C 100 g-1干土d-1)是生产和呼吸成分的总和[25]。根据文献[26], 假设土壤线虫的生命周期持续时间(以天为单位)可以近似为cp值的12倍, 使用0.058作为系数来估算每天的碳呼吸量(以μg为单位)。计算公式如下:

|
式中, Nt是指第t个属的个体数量, Wt和mt分别指第t个属的个体生物量和cp值。
1.6.3 能量流通数值的计算:假设食细菌, 食真菌和植食性线虫从资源中获取能量, 杂食/捕食线虫从其他营养类群的取食偏好建立五节点食物网[14]。首先计算了杂食捕食类线虫的能量通量(Fo)。因为这种能量代谢的计算不涉及更高营养水平的能量损失, 而是专门用于评估线虫的能量需求。随后, 计算了不同营养类群节点间的能量流动数值(Fi)。计算公式如下:
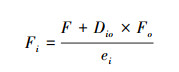
|
式中, F代表根据生产和呼吸计算的能量通量, Dio代表捕食杂食线虫对第i类营养类群的捕食偏好(根据丰度比例分配), Fo代表捕食杂食线虫的能量通量, 两者之积是指向更高营养级的能量损失。e代表营养级间的同化效率, 植食性线虫同化效率为0.25, 食细菌线虫为0.60, 食真菌线虫为0.38, 捕食杂食线虫为0.5[27—28]。
1.7 统计分析数据采用3次平行样本测定的平均值±标准误差表示, 利用SPSS 25.0进行统计分析, 对不同处理进行单因素方差分析和最小差异性显著(LSD), 显著水平为0.05, 使用Origin 2017绘图。采用Canoco 5软件, 以线虫丰度, 生态指数和各营养类群代谢足迹为响应变量, 以土壤理化因子为解释变量进行冗余分析。
2 结果与分析 2.1 土壤线虫丰度及群落结构不同氮磷添加处理平均每100 g干土可分离出线虫数量如图 1所示。与CK相比, 长期磷添加显著增加了线虫丰度, 其中P12处理最高, 为1147条/100 g干土, 其次是P24处理, 为1100条/100 g干土, 其余处理对线虫丰度影响不显著。

|
| 图 1 不同氮磷添加下土壤的线虫丰度 Fig. 1 Abundance of soil nematodes with different nitrogen and phosphorus additions CK: 不施肥No fertilization;N: 氮肥Nitrogen fertilizer;P: 磷肥Phosphorus fertilizer;下标12, 24:施肥量分别为60和120 g m-2 a-1 The amount of fertilizer was 60 and 120 g m-2 a-1;不同小写字母表示不同处理间具有显著差异(P<0.05) |
土壤线虫营养类群结构在不同氮磷添加下存在差异(图 2), 其中土壤植物寄生线虫(Pp)比例显著高于其他类群(50.7%—66.4%)。与CK相比, 氮磷添加提升了食真菌线虫(Fu)的比例, 增幅为P24(13.7%)>P12(11.0%)>N12(9.5%)>N12P12(2.9%)>N24(1.9%)。N12处理土壤食细菌线虫(Ba)占比最高(28.2%), 其余施肥处理降低了土壤食细菌线虫比例, N12P12处理杂食/捕食类线虫(Op)占比最高(7.6%)。

|
| 图 2 不同氮磷添加下土壤线虫各营养类群的比例 Fig. 2 Trophic taxa proportion of soil nematodes with different nitrogen and phosphorus additions Ba: 食细菌类线虫Bacterivores;Fu: 食真菌类线虫Fungivores;Pp:植物寄生类线虫Plant parasites;Op: 杂食/捕食类线虫Omnivores-predators |
由图 3可知, 在CK处理中未分离出cp5类群线虫, 而cp2的线虫类群在所有处理中占比最高(68.1%—84.0%)。不同氮磷添加处理后, cp1的线虫类群占比减少, 增加了cp5的线虫比例。

|
| 图 3 不同氮磷添加土壤线虫生活史类群特征 Fig. 3 Characterization of soil nematode life history taxa with different nitrogen and phosphorus additions cp: 殖民者-居住者Colonizer-persister;cp1-5:表示土壤线虫从殖民者到居住者类型过渡 |
土壤线虫作为生态环境受干扰程度的敏感性指标生物, 利用不同生态指数可以反映出土壤食物网情况和生态环境质量。如表 2所示, N12、P12和P24处理土壤线虫物种多样性指数(H)和均匀度指数(J)显著高于其他处理, 表明这三种处理土壤线虫种类丰富且群落结构更为稳定。线虫通路比值(NCR)属于间接评价土壤有机物分解途径的指标, 与CK相比, 氮磷添加显著提升了NCR指数, 表明土壤有机质的分解以细菌为主导。所有处理的土壤富集指数(EI)都小于50, 但结构指数(SI)大于50, 表明土壤养分状况较差但受干扰程度较小, 食物网处于结构化的状态, 其中N12P12处理显著提升了土壤SI指数。
| 生态指数 Ecological indices |
不同氮磷添加Different nitrogen and phosphorus additions | |||||
| CK | N12 | N24 | P12 | P24 | N12P12 | |
| Shannon多样性指数Shannon diversity index | 0.77±0.03b | 1.17±0.09a | 0.79±0.01b | 1.08±0.07a | 1.13±0.01a | 0.72±0.02b |
| 均匀度指数Evenness index | 0.21±0.01b | 0.32±0.02a | 0.22±0.00b | 0.29±0.02a | 0.31±0.00a | 0.20±0.00b |
| 丰富度指数Richness index | 7.33±0.06ab | 7.53±0.04a | 7.14±0.05b | 6.17±0.20c | 6.01±0.03c | 7.24±0.01ab |
| 植物寄生线虫成熟度指数 Plant parasites index |
1.79±0.04a | 1.35±0.03b | 1.88±0.17a | 1.70±0.13ab | 1.70±0.17ab | 2.11±0.14a |
| 成熟度指数Maturity index | 2.60±0.06a | 2.83±0.07a | 2.63±0.26a | 2.72±0.27a | 2.92±0.36a | 2.56±0.17a |
| 线路通路比值Nematode channel ratio | 0.51±0.00c | 0.71±0.03a | 0.65±0.00b | 0.64±0.07b | 0.62±0.01b | 0.65±0.03b |
| 富集指数Enrichment index | 28.28±2.63ab | 33.06±0.91a | 18.11±1.20b | 32.28±3.67a | 34.49±1.76a | 24.90±04.33ab |
| 结构指数Structure index | 55.04±5.04b | 50.72±4.86c | 55.81±5.37b | 56.00±4.90b | 50.08±3.81c | 66.10±3.86a |
| 同列不同字母表示不同处理间差异显著(P<0.05) | ||||||
线虫代谢足迹能够提供关于土壤食物网碳和能流的大小等信息。由图 4可知, 与CK相比, 两种磷添加处理显著提升了食真菌线虫和捕食杂食线虫的代谢足迹, 其中P12处理显著提升了食细菌线虫的代谢足迹, 其余处理对各营养类群的代谢足迹没有产生影响。

|
| 图 4 不同氮磷添加土壤各营养类群线虫代谢足迹 Fig. 4 Metabolic footprints of nematodes from various nutrient taxa in soils with different nitrogen and phosphorus additions 不同小写字母表示同一营养类群在不同处理间具有显著差异(P<0.05) |
以线虫丰度, 生态指数和不同营养类群的代谢足迹为响应变量, 以土壤理化性质为解释变量进行冗余分析(图 5), 结果表明, 两轴共解释了土壤线虫群落结构的61.85%。土壤中速效磷(AP)是解释度最高的环境因子(P<0.01), 解释度为48.10%, 其次为pH(3.3%), 全磷(TP, 3.2%), 含水率(SMC, 2.7%)和全碳(TC, 2.1%)。线虫丰度和代谢足迹与AP、TP呈正相关。土壤SMC和TC含量与SI、PPI呈正相关关系, 与H、MI、EI呈负相关关系。

|
| 图 5 土壤理化性质与线虫群落结构的冗余分析 Fig. 5 Redundancy analysis of soil physicochemical properties and nematode community structure SMC: 含水率Soil moisture content;TN: 全氮Total nitrogen;TP: 全磷Total phosphorus;TC: 全碳Total carbon;AP: 速效磷Available phosphorus;NO3--N: 硝态氮Nitrate nitrogen;H: Shannon多样性指数Shannon diversity index;PPI:植物寄生线虫成熟度指数Plant parasites index;MI: 成熟度指数Maturity index;EI:富集指数Enrichment index;SI:结构指数Structure index;蓝色箭头表示线虫丰度, 生态指数和各营养类群的代谢足迹, 红色箭头表示土壤理化因子 |
通过能量流动通道分析(图 6), 不同氮磷添加处理线虫群落内部的能量流动途径呈现一定的差异性。磷添加(P12和P24)处理的细菌通道、植物通道、真菌通道和捕食通道显著高于其他处理, 各营养类群的生物量也有相同的变化趋势。与CK相比, N12、N24和N12P12处理降低了捕食通道和植物通道, 提升了真菌通道, 增加了杂食/捕食类群的生物量, 但降低了植食性线虫的生物量。

|
| 图 6 不同氮磷添加线虫群落内部能量结构变化 Fig. 6 Changes in energy structure within nematode communities with different nitrogen and phosphorus additions 蓝色表示食细菌类群, 绿色表示植物寄生类群, 红色表示食真菌类群, 橙色表示杂食/捕食类群, 圆圈内部的数字代表各营养群的新鲜生物量(μg/100 g干土), 节点间的沿线数字代表能量流通数值(μg C 100 g-1干土d-1) |
氮磷添加对土壤线虫群落结构产生不同效应。本研究发现, 磷添加处理显著提高了线虫丰度和多样性, 增加了食真菌线虫的比例, 这与此前的研究结果相同[15, 29]。原因是由于该地区生态系统受到严格的磷限制[30], 磷的输入显著提升了土壤中速效磷含量, 为土壤生物提供足够的营养物质, 增加了线虫数量和活性[5]。农田生态系统相互作用网络中的另一个影响关系是施肥对地上和地下植物系统的影响, 以及与土壤动物的联系[31]。磷肥通常会增加该地区农作物的产量[30], 从而增加了根茬及根分泌物的投入, 因此增加了微生物碳源, 促进了细菌真菌等微生物的生长[29], 为食菌线虫提供更多的食物来源[15]。土壤理化性质也是影响土壤线虫丰度的重要驱动因素, 以往许多研究发现长期氮添加对土壤线虫群落产生负面影响[1, 15, 32], 土壤酸化会直接抑制植物和地下土壤动物群落的生长, 而NH4+则对植物寄生线虫有毒害作用[1]。然而氮诱导土壤酸化的影响取决于多种因素, 包括施肥强度、土壤缓冲能力和土地利用历史, 以及细菌和真菌群落对施肥的潜在土壤特异性反应[33]。因此, 与以往研究不同的是, 本研究中影响线虫丰度的主要环境因子为TP和AP, 这也导致了长期氮添加对土壤线虫丰度影响不显著。此外, 气候因子也可以调节氮对线虫多样性和丰度的影响。例如, 氮添加降低了寒冷地区线虫丰度, 但增加了温暖地区的丰度, 对线虫多样性的影响在干燥环境中是负面的, 但在潮湿环境中是积极的[10]。
与磷添加不同的是, 两种氮添加处理(N12和N24)对线虫群落结构产生不同效应。一项关于施氮肥强度对青藏高原草地土壤的研究发现[31], 线虫群落丰度和物种丰富度与施氮量呈驼峰关系, 峰值为60 g m-2 a-1, 这与本实验结果相同, 线虫的H、J和MI指数在N12处理下显著增加。对于氮转化和利用的动物群, 因为与其他土壤生物竞争氮有效性, 导致线虫群落的稳定性降低[34]。氮的输入为土壤生物提供了充足的氮, 线虫群落变得稳定且多样性逐渐增加, 氮过量时, 氮的可用性反而增加了NH4+和Al3+的毒性[5]。因此, N24处理对土壤线虫H、J和SR指数没有影响, 这可能是因为氮肥在该水平上的正负效应之间的权衡[13], 表明相对高施氮强度下的土壤条件对土壤线虫群落来说并不理想。
氮和磷是限制大多数生态系统生产力的最常见因素, 在农业实践和实验研究中, 氮肥和磷肥通常一起使用[5]。先前关于氮磷添加对土壤线虫群落的影响的研究显示了相互矛盾的结果。例如, 长期施氮磷肥降低了农田土壤线虫数量和多样性[22], 却对草原线虫营养类群没有影响[15]。本研究氮磷添加(N12P12)对线虫丰度没有产生显著影响, 但cp4和cp5的线虫类群相对丰度表现出联合效应大于单施氮肥或磷肥的趋势。cp4-5类群为k策略者, 一般来说, r策略者在不稳定环境中占主导地位, 而k策略者在适宜的生活环境中可以长时间稳定存在[3]。因此N12P12处理增强了生态系统的稳定性, 为线虫的生存创造了更适宜的环境, 同时较高的SI指数和较低的EI指数则表明该处理下食物网处于结构化状态[35]。
线虫代谢足迹是反映线虫代谢过程中用于生物量增长和呼吸的碳量, 也表征线虫对的生态系统功能和服务程度的贡献[4]。长期磷添加处理显著增强了食真菌和杂食捕食类线虫的代谢足迹, 而土壤中TP和AP含量也是影响代谢足迹的主要因素。这表明了随着生产系统外源养分的输入, 使得被捕食者生产和代谢活动增强的同时充分满足了捕食者的需求和维持了系统的代谢平衡[7]。土壤食物网内的资源和能量流动在很大程度上取决于土壤生物之间的摄食相互关系[14, 36]。本研究表明, 食细菌线虫到杂食捕食营养类群的能流数值比食真菌类线虫高, 因为与食真菌线虫相比, 食细菌线虫的食物资源相对容易获取, 而且由于真菌产物具有化学抗分解性, 使得微生物衍生的有机物在真菌分解通道中的降解可能较慢[37]。N12, N24和N12P12处理显著降低了食细菌和植物寄生线虫生物量, 但提高了捕食杂食线虫生物量。这是由于植物寄生线虫在食物网中作为被捕食者存在, 此外植物寄生线虫可以直接或间接诱导植物来源的碳释放到土壤中[37], 为更高营养级线虫提供充足的食物资源。
4 结论不同氮磷添加处理对黄土旱塬农田土壤线虫群落结构和能量流动特征产生不同影响。
(1) 与不施肥相比, 磷添加处理为线虫的生长提供丰富的资源, 提升了线虫丰度和多样性指数(H), 增强了食真菌和捕食杂食线虫的代谢足迹, 食物网内拥有更高的能量通量。
(2) N12处理显著增加了H、均匀度指数(J)和食真菌线虫能量通道, 而N24处理却没有影响, 表明高施肥量并不一定会产生积极的效果。
(3) 土壤中TP和AP含量是影响线虫丰度和代谢足迹的主要环境因子。
因此, 在黄土旱塬农田中, 建议添加磷肥, 控制氮肥施用量, 并注意维持适当的土壤磷含量, 从而支持线虫群落的健康发展。这些建议旨在优化土壤线虫群落结构, 促进土壤生态系统的健康和稳定性, 从而为农田管理提供科学依据。
| [1] |
Wang H L, Liu G C, Huang B B, Wang X C, Xing Y J, Wang Q G. Long-term nitrogen addition and precipitation reduction decrease soil nematode community diversity in a temperate forest. Applied Soil Ecology, 2021, 162: 103895. DOI:10.1016/j.apsoil.2021.103895 |
| [2] |
Yan D M, Yan D H, Song X S, Yu Z L, Peng D, Ting X, Weng B S. Community structure of soil nematodes under different drought conditions. Geoderma, 2018, 325: 110-116. DOI:10.1016/j.geoderma.2018.03.028 |
| [3] |
Bongers T. The maturity index: an ecological measure of environmental disturbance based on nematode species composition. Oecologia, 1990, 83(1): 14-19. DOI:10.1007/BF00324627 |
| [4] |
Ferris H. Form and function: metabolic footprints of nematodes in the soil food web. European Journal of Soil Biology, 2010, 46(2): 97-104. DOI:10.1016/j.ejsobi.2010.01.003 |
| [5] |
Zhao J, Wang F M, Li J, Zou B, Wang X L, Li Z A, Fu S L. Effects of experimental nitrogen and/or phosphorus additions on soil nematode communities in a secondary tropical forest. Soil Biology and Biochemistry, 2014, 75: 1-10. DOI:10.1016/j.soilbio.2014.03.019 |
| [6] |
张晓珂, 梁文举, 李琪. 我国土壤线虫生态学研究进展和展望. 生物多样性, 2018, 26(10): 1060-1073. DOI:10.17520/biods.2018082 |
| [7] |
程云云, 孙涛, 王清奎, 梁文举, 张晓珂. 模拟氮沉降对温带森林土壤线虫群落组成和代谢足迹的影响. 生态学报, 2018, 38(2): 475-484. |
| [8] |
李乌日吉木斯, 高欣梅, 张连奎, 木其勒, 特格希巴雅尔, Bayarmaa Gun-Aajav. 不同施氮量对大豆田土壤线虫群落的影响. 现代农业科技, 2022(21): 144-148, 157. |
| [9] |
吴少彬, 张萍, 余伯均, 喻佳欣, 雷应雪, 陈开惠, 杨礼通. 模拟氮沉降对土壤线虫群落的影响. 四川农业科技, 2022(9): 86-88. |
| [10] |
Xing W, Lu X M, Niu S L, Chen D M, Wang J S, Liu Y, Wang B X, Zhang S, Li Z L, Yao X J, Yu Q, Tian D S. Global patterns and drivers of soil nematodes in response to nitrogen enrichment. CATENA, 2022, 213: 106235. DOI:10.1016/j.catena.2022.106235 |
| [11] |
齐莎, 赵小蓉, 郑海霞, 林启美. 内蒙古典型草原连续5年施用氮磷肥土壤生物多样性的变化. 生态学报, 2010, 30(20): 5518-5526. |
| [12] |
Pan F, Han X, McLaughlin N B, Li C, Zhao D, Zhan L, Xu Y. Effect of long-term fertilization on free-living nematode community structure in Mollisols. Journal of Soil Science and Plant Nutrition, 2015, 15(1): 129-141. |
| [13] |
Zhao J, Zhang W, Wang K L, Song T Q, Du H. Responses of the soil nematode community to management of hybrid napiergrass: the trade-off between positive and negative effects. Applied Soil Ecology, 2014, 75: 134-144. DOI:10.1016/j.apsoil.2013.10.011 |
| [14] |
Wan B B, Hu Z K, Liu T, Yang Q, Li D M, Zhang C Z, Chen X Y, Hu F, Kardol P, Griffiths B S, Liu M Q. Organic amendments increase the flow uniformity of energy across nematode food webs. Soil Biology and Biochemistry, 2022, 170: 108695. DOI:10.1016/j.soilbio.2022.108695 |
| [15] |
张志委, 胡艳宇, 魏海伟, 侯双利, 殷江霞, 吕晓涛. 氮磷输入对过度放牧退化草原土壤线虫群落的影响. 应用生态学报, 2019, 30(11): 3903-3910. |
| [16] |
鲍士旦. 土壤农化分析(第三版). 北京: 中国农业出版社, 2000.
|
| [17] |
毛小芳, 李辉信, 陈小云, 胡锋. 土壤线虫三种分离方法效率比较. 生态学杂志, 2004, 23(3): 149-151. |
| [18] |
尹文英. 中国土壤动物检索图鉴. 北京: 科学出版社, 1998.
|
| [19] |
Bongers T. The nematodes of the Netherlands. Utrecht: Foundation Publisher of Royal Dutch Natural History Society, 1988.
|
| [20] |
Yeates G W, Bongers T, De Goede R G M, Freckman D W, Georgieva S S. Feeding habits in soil nematode families and genera-an outline for soil ecologists. Journal of Nematology, 1993, 25(3): 315-331. |
| [21] |
Shannon C E. The mathematical theory of communication. 1963. M D Computing: Computers in Medical Practice, 1997, 14(4): 306-317.
|
| [22] |
杨盼盼, 黄菁华, 张欣玥, 陈静, 赵世伟. 长期施肥对渭北旱塬麦田土壤线虫群落特征的影响. 干旱地区农业研究, 2022, 40(5): 242-251. |
| [23] |
Yeates G W. Nematodes as soil indicators: functional and biodiversity aspects. Biology and Fertility of Soils, 2003, 37(4): 199-210. |
| [24] |
Ferris H, Bongers T, de Goede R G M. A framework for soil food web diagnostics: extension of the nematode faunal analysis concept. Applied Soil Ecology, 2001, 18(1): 13-29. |
| [25] |
Bhusal D R, Tsiafouli M A, Sgardelis S P. Temperature-based bioclimatic parameters can predict nematode metabolic footprints. Oecologia, 2015, 179(1): 187-199. |
| [26] |
Liao X H, Fu S L, Zhao J. Altered energy dynamics of multitrophic groups modify the patterns of soil CO2 emissions in planted forest. Soil Biology and Biochemistry, 2023, 178: 108953. |
| [27] |
Jochum M, Barnes A D, Ott D, Lang B, Klarner B, Farajallah A, Scheu S, Brose U. Decreasing stoichiometric resource quality drives compensatory feeding across trophic levels in tropical litter invertebrate communities. The American Naturalist, 2017, 190(1): 131-143. |
| [28] |
Buchan D, Gebremikael M T, Ameloot N, Sleutel S, De Neve S. The effect of free-living nematodes on nitrogen mineralisation in undisturbed and disturbed soil cores. Soil Biology and Biochemistry, 2013, 60: 142-155. |
| [29] |
Chen X Y, Daniell T J, Neilson R, O'Flaherty V, Griffiths B S. Microbial and microfaunal communities in phosphorus limited, grazed grassland change composition but maintain homeostatic nutrient stoichiometry. Soil Biology and Biochemistry, 2014, 75: 94-101. |
| [30] |
高小峰, 闫本帅, 吴春晓, 王国梁. 长期施肥对黄土丘陵坡地农田土壤质量和谷子产量的影响. 干旱地区农业研究, 2021, 39(5): 76-83. |
| [31] |
Hu J, Chen G R, Hassan W M, Lan J B, Si W T, Wang W, Li G X, Du G Z. The impact of fertilization intensity on soil nematode communities in a Tibetan Plateau grassland ecosystem. Applied Soil Ecology, 2022, 170: 104258. |
| [32] |
Liang S W, Kou X C, Li Y B, Lü X T, Wang J K, Li Q. Soil nematode community composition and stability under different nitrogen additions in a semiarid grassland. Global Ecology and Conservation, 2020, 22: e00965. |
| [33] |
Ma X M, Zhou Z, Chen J, Xu H, Ma S H, Dippold M A, Kuzyakov Y. Long-term nitrogen and phosphorus fertilization reveals that phosphorus limitation shapes the microbial community composition and functions in tropical montane forest soil. Science of the Total Environment, 2023, 854: 158709. |
| [34] |
Wei C Z, Zheng H F, Li Q, Lü X T, Yu Q, Zhang H Y, Chen Q S, He N P, Kardol P, Liang W J, Han X G. Nitrogen addition regulates soil nematode community composition through ammonium suppression. PLoS One, 2012, 7(8): e43384. |
| [35] |
李钰飞, 李吉进, 许俊香, 刘本生, 乔玉辉, 孙钦平. 铜、锌污染梯度对自然林地和农田土壤线虫群落的影响. 土壤学报, 2020, 57(6): 1492-1503. |
| [36] |
Barnes A D, Jochum M, Mumme S, Haneda N F, Farajallah A, Widarto T H, Brose U. Consequences of tropical land use for multitrophic biodiversity and ecosystem functioning. Nature Communications, 2014, 5: 5351. |
| [37] |
Zhang Z Y, Zhang X K, Xu M G, Zhang S Q, Huang S M, Liang W J. Responses of soil micro-food web to long-term fertilization in a wheat-maize rotation system. Applied Soil Ecology, 2016, 98: 56-64. |