 2024, Vol. 44
2024, Vol. 44文章信息
- 牟英坤, 陈志飞, 周俊杰, 赖帅彬, 靳媛, 王绍妍, 徐炳成
- MOU Yingkun, CHEN Zhifei, ZHOU Junjie, LAI Shuaibin, JIN Yuan, WANG Shaoyan, XU Bingcheng
- 黄土丘陵区氮磷添加对草地群落优势种养分利用特征的影响
- Effects of nitrogen and phosphorus addition on nutrient use characteristics of grassland community dominant species in loess hilly-gully region
- 生态学报. 2024, 44(5): 2069-2080
- Acta Ecologica Sinica. 2024, 44(5): 2069-2080
- http://dx.doi.org/10.20103/j.stxb.202304170785
-
文章历史
- 收稿日期: 2023-04-17
- 网络出版日期: 2023-12-11





2. 西北农林科技大学黄土高原土壤侵蚀与旱地农业国家重点实验室, 杨凌 712100;
3. 贵州大学生命科学学院, 贵阳 550025
2. State Key Laboratory of Soil Erosion and Dryland Farming on the Loess Plateau, Northwest Agriculture and Forestry University, Yangling 712100, China;
3. College of Life Sciences, Guizhou University, Guiyang 550025, China
作为黄土丘陵区植被类型的主体, 草地在维持区域生态景观和发挥生态防护功能方面具有重要作用[1]。由于长期的土壤侵蚀, 该区土壤养分严重损失, 肥力下降, 草地群落生产力普遍受到土壤氮磷有效性的限制[2—5], 造成群落结构简单、生态功能低下, 退化后的生态恢复缓慢等问题[6]。自1999年黄土高原实施大范围退耕还林(草)政策与封山禁牧措施以来, 退化林草植被均得到了一定恢复, 生长状况明显改善, 但很多草地群落自我修复较慢[7]。通过施用外源氮磷肥, 提高土壤中氮磷养分含量, 可促进草地群落生产力的提高和草地群落结构的改善[8—10], 是草地管理的一项重要措施。但不同的氮磷施肥量配比效果有异, 因此, 探索合理的氮磷施肥量及其配比, 对促进草地群落恢复和生态功能提升具有重要意义。
养分重吸收是指植物将衰老组织中的部分养分转移到其他新生组织的过程, 是植物应对环境养分限制的重要策略之一[11]。氮磷化学计量特征是研究植物养分限制关系和不同物种养分利用状况的有效途径, 其中叶片氮磷比一定程度上能反映植物乃至植被生态系统养分限制状况[12—13]。研究表明, 适量的氮磷添加可提高土壤中氮磷养分的有效含量, 调整植物器官的氮磷比, 降低重吸收效率, 增加植物地上生物量[14—16];不当的氮磷输入则可能破坏植物与土壤间原有的养分平衡, 导致植物生长发育发生营养限制或加剧某种营养元素的限制, 使植物生长量减小[17]。草地群落中, 不同物种应对环境和养分变化的方式存在差异, 氮磷添加后其养分利用策略不同, 叶片氮磷比、氮磷重吸收效率和地上生物量响应存在很大差异[14], 这些将影响群落物种组成变化。因此, 研究氮磷添加后不同物种养分利用特征对针对性的进行草地恢复调控具有指导意义。
优势种在植物群落中占有重要地位, 是维持群落结构与功能稳定的关键[18]。半干旱地区植被对环境条件改变的响应, 很大程度上依赖于优势物种的反应和适应性[19]。植物相对生物量能够量化各物种在群落中的地位, 进而反映群落物种组成[20]。因此, 阐明氮磷添加下天然草地群落优势种养分利用特征和相对生物量的关系, 有助于从养分利用和循环角度, 探究群落结构和物种多样性变化的潜在机制。论文选取半干旱黄土丘陵区5种典型草地群落优势种, 即白羊草(Bothriochloa ischaemum)、长芒草(Stipa bungeana)、达乌里胡枝子(Lespedeza davurica)、铁杆蒿(Artemisia sacrorum)和猪毛蒿(Artemisia scoparia)为研究对象, 通过探究各优势种叶片氮磷比、养分重吸收和利用效率对氮磷添加的响应特征和差异, 分析其相对生物量与氮磷养分利用特征间的相关性, 以期揭示不同氮磷施肥水平下草地群落物种组成变化的潜在机制, 为该区草地管理中选择合理的氮磷配比提供理论依据。
1 材料与方法 1.1 研究区概况研究区位于陕西省延安市安塞区纸坊沟小流域(36°42′42″—36°46′28″N, 109°13′46″—109°16′03″E, 海拔1010—1431 m)。流域面积约8.27 km2, 流域内坡度在0°—65°之间, 年均降雨量为528.8 mm(1951—2016年), 其中生长季5—9月降雨量占年降雨量的82.2%。年均气温8.8℃, 无霜期约160 d。该区地处典型黄土丘陵沟壑区, 属温带半干旱大陆性季风气候;植被属温带灌丛草原, 区划上属半干旱森林草原带;天然植被以旱生草本植物为主, 主要优势种包括白羊草、铁杆蒿、达乌里胡枝子、长芒草和猪毛蒿等。
1.2 试验设计2018—2019年围封30 m × 50 m的退耕草地为试验样地, 样地坡向为北偏东36°, 坡度17°, 坡面落差8.77 m, 土壤全氮含量为0.60 g/kg, 土壤全磷含量为0.58 g/kg。试验过程中重点加强围栏管护, 防止放牧干扰。每年年初植物返青前全部刈割, 留茬2 cm。采用裂区试验设计, 主区设置4个氮添加水平, 分别为0(N0)、25(N25)、50(N50)和100 kg N hm-2 a-1(N100);副区设置4个磷添加水平, 分别为:0(P0)、20(P20)、40(P40)和80 kg P2O5 hm-2 a-1(P80), 每个处理设置三组重复。各氮磷添加分别在主区和副区采用随机区组布设, 以消除立地条件变化的影响。最低施氮标准(25 kg N hm-2 a-1)参考黄土丘陵区的平均年氮沉降量(21.76 kg N hm-2 a-1)[21], 并依次设置氮沉降量的2倍和4倍以评估未来氮沉降增加对黄土丘陵区草地群落的影响。最低施磷标准参考刘海威等[22]在纸坊沟流域退耕草地的施磷标准(21.7 kg P2O5 hm-2 a-1), 并依据氮添加的倍数依次设置2倍和4倍的施磷水平。样地中沿坡面设置三个10 m × 30 m的区组, 区组间由1.5 m未施肥缓冲区分隔, 每个区组沿坡面设置4个4 m × 4 m的主区, 主区间间隔1 m未施肥缓冲区, 主区再划分为4个2 m×2 m的副区。氮肥为硝酸铵钙[5Ca(NO3)2·NH4NO3·10H2O], 磷肥为重过磷酸钙[Ca(H2PO4)2·H2O]。施肥试验于2018年开始, 氮磷肥一年一次在雨前均匀撒入草地地表。2018年于6月4日完成, 6月5日降雨。2019年于5月21日完成, 5月22日和23日降雨。降雨过程中, 各试验小区均未产生坡面径流。
1.3 测定项目与方法 1.3.1 叶片氮磷含量、相对生物量和氮磷利用效率2019年7月23—28日, 在群落地上生物量达到峰值时, 随机选择每个副区内长势相同的各优势种植物20株, 用细红线标记, 从每株顶端取一个新近完全展开的成熟叶, 经80℃烘干48 h至恒重后称重。将烘干后的叶片在球磨机(MM-400, Retsch, 德国)中研磨成细粉, 以测定成熟叶片氮含量(MLN, mg/g)和磷含量(MLP, mg/g)。
2019年8月25—28日, 在每个处理副区设置一个1 m×1 m样方, 将样方内植物齐地面剪下, 按物种分类装袋后在实验室内经80℃烘干48 h至恒重后称重(SDM g/m2)。将各物种地上生物量占总生物量的百分比作为相对生物量(RB, %)[23]。
将全部样品粉碎后用H2SO4-H2O2消煮, 利用自动凯氏定氮法(Kjektec System 2300 Distilling Unit, Foss, 瑞典)进行各优势种地上部氮含量(SN, mg/g)的测定, 通过紫外分光光度计(UV-2600 spectrophotometer, 日本)利用钼锑比色法进行磷含量(SP, mg/g)的测定。将植物体内每单位的氮素或磷素所获得的地上生物量作为衡量植物氮磷利用效率的指标。物种氮利用效率(NUE, g生物量/g N)和物种磷利用效率(PUE, g生物量/g P)计算公式为:
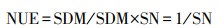
|
(1) |

|
(2) |
2019年9月20—10月20日, 每周对各优势种中红线标记的20株植物进行一次检查, 查看叶片是否全叶枯死, 在全叶枯死时认为叶片完全衰老, 进行采集[24]。经烘干处理后测定衰老叶片的氮含量(SLN, mg/g)和磷含量(SLP, mg/g), 根据植物对养分的重吸收效率和重吸收度来评估其重吸收能力。以衰老叶片中的养分含量表征氮和磷的重吸收度, 养分含量越低表明养分重吸收程度越高。养分重吸收效率为绿叶在叶片脱落前重吸收的氮磷的百分比。以相对养分重吸收效率表征植物的氮磷重吸收比例[25]。氮重吸收效率(NRE, %)、磷重吸收效率(PRE, %)和相对养分重吸收效率(RR, %)计算公式为:
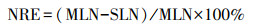
|
(3) |
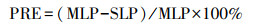
|
(4) |
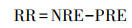
|
(5) |
MLN和MLP分别代表植株成熟叶片N、P含量, SLN和SLP分别代表植物衰老叶片N、P含量。
1.4 数据处理采用Excel 2019进行数据整理, 利用Genstat 22.0进行裂区试验设计方差分析(ANOVA), 以检验氮和磷的主效应、交互效应对优势种成熟叶和衰老叶氮磷比、氮和磷利用效率、氮和磷重吸收效率、相对养分重吸收效率以及相对生物量的影响。若氮、磷或氮磷交互效应显著(P<0.05), 则使用最小显著性差异值(LSD)来检验处理间的差异显著性。利用R 4.2.2对五个优势种叶片氮磷比、氮磷利用和重吸收效率、相对养分重吸收效率进行相关性分析, 对养分利用特征和相对生物量间关系进行主成分(PCA)分析。采用Origin 2021作图。
2 结果与分析 2.1 成熟与衰老叶片的氮磷比不同氮磷添加处理下, 平均成熟叶氮磷比最高的为达乌里胡枝子(17.76), 其次为长芒草(14.88)、白羊草(9.23)、猪毛蒿(8.90)和铁杆蒿(8.45)(图 1)。单施氮下, 白羊草和长芒草成熟叶氮磷比有增加趋势。磷添加显著降低了除猪毛蒿外四个优势种的成熟叶氮磷比(P<0.05)。氮磷添加仅对猪毛蒿成熟叶氮磷比的交互作用显著(P<0.05), 与磷配施显著降低了N50下猪毛蒿成熟叶氮磷比(P<0.05)(图 1)。

|
| 图 1 不同氮磷添加下五个优势种成熟叶片和衰老叶片的氮磷比 Fig. 1 Mature and senescent leaf N ∶ P ratio of the five dominant species under different N and P additions N0:0 kg N hm-2 a-1;N25:25 kg N hm-2 a-1;N50:50 kg N hm-2 a-1;N100:100 kg N hm-2 a-1;P0:0 kg P2O5 hm-2 a-1;P20:20 kg P2O5 hm-2 a-1;P40:40 kg P2O5 hm-2 a-1;P80:80 kg P2O5 hm-2 a-1;括号内数字为不同氮磷添加下最小显著差异值(LSD);* 表示 P<0.05, ** 表示P<0.01, *** 表示P<0.001, n.s. 表示差异不显著 |
不同氮磷添加处理下, 衰老叶氮磷比最高的为达乌里胡枝子(17.51), 其次为长芒草(15.07)、铁杆蒿(13.29)、白羊草(12.36)和猪毛蒿(8.90)(图 1)。单施氮显著提高了白羊草、长芒草和猪毛蒿衰老叶片氮磷比(P<0.05)。单施磷显著降低了白羊草、长芒草、达乌里胡枝子和铁杆蒿衰老叶氮磷比(P<0.05)。与磷配施显著降低了N50下猪毛蒿衰老叶氮磷比(P<0.05)。达乌里胡枝子和长芒草衰老叶氮磷比受到氮磷交互作用的显著影响(P<0.05)(图 1)。
2.2 叶片氮磷重吸收效率和相对重吸收效率不同氮磷添加处理下, 长芒草(43.15%)和白羊草(45.81%)的平均氮重吸收效率(NRE)显著高于猪毛蒿(38.14%)、铁杆蒿(30.44%)和达乌里胡枝子(22.55%)(图 2)。随着施氮量的增加, 猪毛蒿NRE逐渐增大。单施磷显著降低了达乌里胡枝子NRE(P<0.05)。铁杆蒿和猪毛蒿NRE受到氮磷交互作用的显著影响(P<0.05), 与磷配施显著降低了N25下铁杆蒿NRE,提高了N100下猪毛蒿NRE(P<0.05)(图 2)。

|
| 图 2 不同氮磷添加下五个优势种的氮重吸收效率(NRE)、磷重吸收效率(PRE) 及相对养分重吸收效率(RR) Fig. 2 Nitrogen resorption efficiency (NRE), phosphorus resorption efficiency (PRE) and relative nutrient resorption efficiency (RR) of the five dominant species under different N and P additions |
不同氮磷添加处理下, 白羊草、长芒草、铁杆蒿、猪毛蒿和达乌里胡枝子的平均磷重吸收效率(PRE) 分别为56.73%、52.47%、53.40%、45.00%和21.30%(图 2)。单施氮或单施磷均提高了长芒草的PRE。氮磷配施下, 白羊草PRE相比于单施磷显著增加(P<0.05)。P80与氮配施下, 白羊草PRE相较于单施氮显著增加(P<0.05)。与磷配施显著降低了N50和N100下达乌里胡枝子PRE(P<0.05)。单施N25下, 铁杆蒿PRE显著高于N25与磷配施(P<0.05)。
单施氮时, 白羊草的RR随施氮量的增加而逐渐降低, 而猪毛蒿的RR随施氮量的增加而逐渐增加(图 2)。单施磷时, 白羊草和长芒草的RR随磷添加水平的增加而逐渐降低。与P40和P80配施显著提高了单施氮下铁杆蒿RR(P<0.05)。
2.3 氮磷利用效率不同氮磷添加处理下, 白羊草、长芒草、铁杆蒿、达乌里胡枝子和猪毛蒿的平均氮利用效率(NUE)分别为114.95 g生物量/g N、78.95 g生物量/g N、46.32 g生物量/g N、42.47 g生物量/g N和38.78 g生物量/g N(图 3)。单施氮降低了白羊草、长芒草和铁杆蒿NUE。单施磷降低了白羊草、长芒草和达乌里胡枝子NUE。氮磷交互作用对达乌里胡枝子、铁杆蒿和长芒草NUE均有显著影响(P<0.05), N25和N50与磷配施下, 达乌里胡枝子NUE显著低于N25和N50单施(P<0.05)。氮磷配施下长芒草NUE显著高于单施氮(P<0.05)。

|
| 图 3 不同氮磷添加下五个优势种的叶片氮利用效率(NUE) 和磷利用效率(PUE) Fig. 3 Leaf nitrogen use efficiency (NUE) and phosphorus use efficiency (PUE) of the five dominant species under different N and P additions |
不同氮磷添加处理下, 白羊草、长芒草、达乌里胡枝子、猪毛蒿和铁杆蒿的平均磷利用效率(PUE) 分别为909.34 g生物量/g P、886.09 g生物量/g P、576.33 g生物量/g P、431.06 g生物量/g P和383.78 g生物量/g P(图 3)。单施氮显著提高了白羊草、铁杆蒿、长芒草和猪毛蒿的PUE, 而显著降低了达乌里胡枝子的PUE(P<0.05)。单施磷显著降低了五个优势种的PUE(P<0.05)。氮磷交互作用对白羊草和达乌里胡枝子PUE均有显著影响(P<0.05)。除白羊草N50与P20配施外, 与磷配施显著降低了单施氮下白羊草、达乌里胡枝子、长芒草和铁杆蒿的PUE (P<0.05)。
2.4 相对生物量单施磷显著降低了白羊草相对生物量, N25与磷配施下白羊草相对生物量较单施N25显著降低(P<0.05)(图 4)。单施磷显著提高了达乌里胡枝子相对生物量, 而单施氮则与之相反, 单施N25或N50下达乌里胡枝子相对生物量显著低于N25或N50与磷配施(P<0.05)。氮磷交互作用仅对铁杆蒿相对生物量有显著影响(P<0.05), 铁杆蒿相对生物量在N50下达到最高, 但与磷配施后显著降低(P<0.05)。单施N100下铁杆蒿相对生物量显著低于N100与磷配施(P<0.05)。猪毛蒿相对生物量随施肥水平的提高整体呈上升趋势(图 4)。

|
| 图 4 不同氮磷添加下五个优势种的相对生物量变化 Fig. 4 Relative biomass change of the five dominant species under different N and P additions 横坐标为氮磷添加配比;N0P0:对照;N0P20:P20单施;N0P40:P40单施;N0P80:P80单施;N25P0:N25单施;N25P20:N25与P20配施;N25P40:N25与P40配施;N25P80:N25与P80配施;N50P0:N50单施;N50P20:N50与P20配施;N50P40:N50与P40配施;N50P80:N50与P80配施;N100P0:N100单施;N100P20:N100与P20配施;N100P40:N100与P40配施;N100P80:N100与P80配施 |
五个优势种成熟叶氮磷比均与衰老叶氮磷比显著正相关(P<0.05), 磷重吸收效率与相对养分重吸收效率显著负相关(P<0.05)。白羊草、达乌里胡枝子和铁杆蒿中衰老叶氮磷比与相对养分重吸收效率呈显著负相关(P<0.05)。五个优势种的氮与磷重吸收效率间均呈正相关关系(图 5)。五个优势种的氮重吸收效率均与磷利用效率呈正相关关系。铁杆蒿、长芒草和猪毛蒿的氮磷利用效率间均表现为负相关, 而白羊草和达乌里胡枝子的表现为正相关。

|
| 图 5 不同氮磷添加下五个优势种叶片氮磷比和养分利用特征间的相关性 Fig. 5 Correlation between leaf N ∶ P ratio and utilization characteristics of the five dominant species under different N and P additions MLN ∶ P:成熟叶片氮磷比Mature leaf N ∶ P;SLN ∶ P:衰老叶片氮磷比Senescent leaf N ∶ P;NRE:氮重吸收效率N resorption efficiency;PRE:磷重吸收效率P resorption efficiency;RR:相对养分重吸收效率Relative nutrient resorption efficiency;NUE:氮利用效率N use efficiency;PUE:磷利用效率P use efficiency;图中正相关以蓝色表示, 负相关以红色表示, 颜色强度与相关系数成正比;*表示P<0.05, ** 表示P<0.01, ***表示P<0.001 |
采用主成分分析(PCA)来识别氮磷条件下不同物种各变量与相对生物量(RB)间的相关性(图 6)。不同氮磷添加处理下, 第一和第二排序轴对各变量变异的总解释量均超过50%, 将其作为主成分轴。单施氮或N50和N100与磷配施条件下, 相对生物量与养分利用特征(NRE、PRE、NUE、PUE)呈正相关, 与氮磷比和相对养分重吸收效率呈负相关。单施磷或N25与磷配施下, 相对生物量与氮磷比、相对养分重吸收效率和磷利用效率呈正相关, 与氮利用效率和氮磷重吸收效率呈负相关。单施氮条件下, 五种植物呈现五个独立的簇, 位于坐标系的不同位置, 说明五种植物对氮添加的响应方向和程度各不相同。单施磷时, 五种植物大致分为两个分离簇, 第一类为白羊草、长芒草、铁杆蒿和猪毛蒿, 第二类为达乌里胡枝子, 说明达乌里胡枝子对磷添加的响应与其他四种植物有明显差异。

|
| 图 6 五个优势种氮磷比、养分利用和相对生物量的主成分分析 Fig. 6 Principal components analysis (PCA) of N ∶ P, nutrient utilization and relative biomass of the five dominant species 箭头代表对应于单个变量的特征向量;RB:相对生物量Relative biomass |
不同物种叶片氮磷含量及其化学计量比存在显著差异, 其对氮磷添加的响应也具有特异性[12, 26]。本研究中, 豆科植物达乌里胡枝子叶片氮磷比最高, 其次是禾本科植物白羊草和长芒草, 菊科植物铁杆蒿和猪毛蒿叶片氮磷比较低[27](图 1)。氮添加对达乌里胡枝子无明显影响, 可能是其具有生物固氮作用, 或者其生长过程氮素需求较低, 施氮与否对其影响并不明显[26, 28]。与此相反的是, 白羊草和长芒草具有较高的氮磷比, 但本身不具备固氮能力, 因此对氮添加响应更敏感。除达乌里胡枝子外, 其他四种植物成熟叶氮磷比在中低氮添加水平下较高, 而在高氮添加水平下有下降趋势, 这是因为适度的外源氮添加提高了土壤中氮的有效性, 促进了植物对氮的吸收, 使氮磷比增加, 而过量氮添加会使土壤pH值降低, 从而影响植物养分尤其是磷的转移和利用, 导致植物叶片氮磷比有所下降[29]。
不同优势种地上生物量对氮磷添加的响应不同, 使其在地上总生物量中的占比发生改变。单施磷可以促进豆科植物结瘤并增强根瘤中固氮酶活性, 提高豆科植物的固氮能力[30], 因此达乌里胡枝子的地上生物量和相对生物量响应明显(图 4), 也表明黄土丘陵区天然草地中达乌里胡枝子的生长受到磷限制。单施氮或磷均提高了铁杆蒿和猪毛蒿的地上生物量和相对生物量, 说明两者均受到氮磷共限制。与铁杆蒿相比, 猪毛蒿随氮磷添加的差异更显著, 说明生活史短的植物生长速度和养分转换较快, 对养分添加更为敏感[31—32]。氮和磷添加均使白羊草和长芒草的相对生物量表现出下降趋势, 这可能是由于氮磷添加显著提高了达乌里胡枝子、铁杆蒿和猪毛蒿的相对生物量, 使其在群落中占据更多的生态位, 导致光和养分资源竞争增强, 白羊草和长芒草养分利用受限制, 导致其相对生物量有所降低。
N∶P阈值假说认为, 存在特定N∶P阈值来判定植物养分限制状况, 可将N∶P用于判定包含群落和物种水平的植物养分限制类型[14]。Koerselman和Meuleman提出[33], 当N∶P>16.0时, 植物生长受到磷限制;N∶P<14.0时受到氮限制;当N∶P比介于14.0和16.0间时, 植物生长不受氮磷限制, 或受到两者的共同限制。本研究中, 未施肥条件下白羊草、达乌里胡枝子、铁杆蒿和长芒草的氮磷比分别为10.85、24.63、10.73和19.39, 猪毛蒿不同氮磷添加水平下N∶P均小于16.0, 说明达乌里胡枝子和长芒草受到磷限制, 其他三个优势种受到氮限制。单施氮时, 白羊草和铁杆蒿氮磷比有增加趋势, 说明氮输入能够缓解氮限制[34];单施磷显著降低了白羊草、达乌里胡枝子、铁杆蒿和长芒草的氮磷比, 且最终四个优势种的氮磷比均小于14, 表明磷添加缓解了磷限制而加剧了氮限制[35];当氮磷配施时, 除猪毛蒿外的四个优势种氮磷比相比不施肥均显著降低, 表明氮对植物叶片N∶P的直接正效应弱于磷对其直接负效应[17]。
3.2 氮磷添加对植物养分利用的影响养分重吸收是植物一种重要的养分保护策略。对全球数据统计结果表明, 分别有62%的氮和65%的磷在落叶前被重新吸收到其他活组织中[36]。由于受基因、土壤营养物质等因素的影响, 不同植物物种对营养物质的重吸收具有差异。通常固氮植物的氮重吸收效率比非固氮植物要低, 禾本科植物的氮重吸收效率比豆科和杂类草要高, 而豆科植物的叶片氮重吸收效率与杂类草差异不明显, 或者比其他功能群植物低, 这是因为豆科草类可通过根瘤固定空气氮和土壤N库两个来源获取氮营养, 叶片重吸收效率较低, 而浅根系的禾本科草类更依赖于内部养分循环, 因而具有较高的养分重吸收效率[37]。本研究中, 达乌里胡枝子氮重吸收效率最低, 铁杆蒿和猪毛蒿居中, 白羊草和长芒草氮重吸收效率最高, 证实了上述结果。
相关性分析结果显示, 五种植物的成熟叶与衰老叶氮磷比之间、氮与磷重吸收效率之间都表现出相关性, 说明氮磷重吸收存在着耦合效应。植物的氮磷重吸收比例受到其本身内在氮磷比的制约, 在不同的养分添加条件下, 成熟叶片会按照一定比例再吸收衰老叶片中的氮磷, 以保持其氮磷比平衡[38]。氮磷重吸收间的耦合效应使得磷的重吸收效率在五个优势种间表现出与氮重吸收效率相同的大小差异, 即达乌里胡枝子的磷重吸收效率最低, 铁杆蒿和猪毛蒿居中, 白羊草和长芒草的磷重吸收效率最高。
3.3 氮磷添加水平对群落物种组成的影响不同物种对环境条件变化的响应和适应能力不同。物种间养分利用策略也存在较大差异, 这可能是群落中物种间功能互补和生态位分离的结果, 对于预测氮磷添加下群落结构的改变具有重要意义[26]。植物相对生物量反映了不同物种在群落中的地位和作用, 能够量化反映群落结构和物种组成的变化[20]。PCA分析结果显示, 植物相对生物量与其养分利用特征密切相关, 且随氮磷添加水平的不同而改变(图 6), 说明植物种间养分利用策略的权衡决定了其在群落中的生物量组成。本研究中, 单施磷或N25与磷配施下, 各物种相对生物量与氮磷比和磷利用效率呈正相关关系, 与氮利用效率和氮磷重吸收效率呈负相关关系。单施氮、N50和N100与磷配施下, 各物种相对生物量与氮磷利用效率和重吸收效率呈正相关, 与氮磷比呈负相关, 且与氮添加相比, 磷添加对植物氮磷利用影响更显著, 因此更易对物种组成产生影响。
白羊草和长芒草具有较高的氮磷比、氮磷重吸收效率与利用效率, 因此具有较高的内稳性[39]和适应养分贫瘠生境或因逆境导致的养分获取困难生境的能力[37], 更能充分利用比较有限的养分来与其它物种竞争, 较低的氮磷水平有益于其在种群中保持较高优势度。因此, 不施肥条件下, 白羊草和长芒草相对生物量最高。与之相反, 铁杆蒿和猪毛蒿氮磷比、氮磷重吸收效率和利用效率均较低, 内稳性较低, 更易受到外界环境的影响, 养分充足的环境更有益其获得竞争优势。因此, 高氮高磷添加水平下铁杆蒿和猪毛蒿相对生物量最高。达乌里胡枝子的氮磷比在三种功能群中最高, 这得益于其独特的固氮能力, 使其受外源氮供应的影响较小, 而对外源磷供应的响应更为敏感[26, 40], 低氮高磷的养分环境更有利于其占据生态位。因此, 低氮高磷添加水平下达乌里胡枝子相对生物量最高。
4 结论黄土丘陵区草地群落优势种的氮磷比、相对生物量、养分重吸收效率和养分利用效率对氮磷添加的响应存在明确的物种分异性, 氮与磷的重吸收存在耦合效应。不同氮磷添加水平下, 禾本科植物白羊草和长芒草的氮磷利用效率和重吸收效率最高, 豆科植物达乌里胡枝子叶片氮磷比最高, 而氮磷重吸收效率最低, 菊科植物铁杆蒿和猪毛蒿氮磷比、氮磷重吸收效率和利用效率均较低。各物种相对生物量随氮磷添加水平不同而改变, 且与氮添加相比, 磷添加对植物氮磷利用的影响更显著。各优势种通过改变相对生物量与养分利用特征间关系来应对土壤氮磷含量的变化。白羊草和长芒草内稳性较高, 通过增强氮磷比、氮磷利用和重吸收效率对相对生物量的正面影响来提高种间竞争力。与之相反, 猪毛蒿和铁杆蒿内稳性较低, 通过降低氮磷比、氮磷利用和重吸收效率对相对生物量的负面影响来获取竞争优势。达乌里胡枝子的生物固氮能力使其不易受到外源氮供应的影响, 并借助高氮磷比优势保持磷添加下在群落中的优势度。总之, 草地群落优势种对氮磷添加响应存在明确的种间差异性, 这是影响草地恢复和物种组成的重要因素, 在通过氮磷添加调控草地恢复过程中, 须考虑氮磷配比, 权衡群落各物种养分需求和物种组成及其关系, 以达到促进草地恢复、提升草地功能的目的。
| [1] |
白永飞, 赵玉金, 王扬, 周楷玲. 中国北方草地生态系统服务评估和功能区划助力生态安全屏障建设. 中国科学院院刊, 2020, 35(6): 675-689. |
| [2] |
王宝荣, 杨佳佳, 安韶山, 张海鑫, 白雪娟. 黄土丘陵区植被与地形特征对土壤和土壤微生物生物量生态化学计量特征的影响. 应用生态学报, 2018, 29(1): 247-259. |
| [3] |
Vitousek P M, Porder S, Houlton B Z, Chadwick O A. Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen-phosphorus interactions. Ecological Applications, 2010, 20(1): 5-15. DOI:10.1890/08-0127.1 |
| [4] |
孙天雨, 王雪, 李丹洋, 刘欣蕊, 王瑞丽, 张硕新. 黄土高原草地根系生物量沿环境梯度变化规律. 生态学报, 2021, 41(15): 6234-6244. |
| [5] |
袁梓裕, 张路, 廖李容, 王杰, 雷石龙, 刘国彬, 方怒放, 张超. 黄土高原草地植物多样性与群落稳定性的关系及其驱动因素. 生态学报, 2023, 43(1): 60-69. |
| [6] |
邹亚丽, 牛得草, 杨益, 文海燕, 傅华. 氮素添加对黄土高原典型草原土壤氮矿化的影响. 草地学报, 2014, 22(3): 461-468. |
| [7] |
王诗绮, 刘焱序, 李琰, 傅伯杰. 近20年黄土高原生态系统服务研究进展. 生态学报, 2023, 43(1): 26-37. |
| [8] |
曾德慧, 陈广生. 生态化学计量学: 复杂生命系统奥秘的探索. 植物生态学报, 2005, 29(6): 1007-1019. DOI:10.3321/j.issn:1005-264X.2005.06.018 |
| [9] |
Chen Z F, Zhou J J, Lai S B, Jian C X, Chen Y, Luo Y, Xu B C. Species differences in stoichiometric homeostasis affect grassland community stability under N and P addition. Environmental Science and Pollution Research, 2023, 30(22): 61913-61926. DOI:10.1007/s11356-023-26479-3 |
| [10] |
Chen Z F, Xiong P F, Zhou J J, Lai S B, Jian C X, Xu W Z, Xu B C. Effects of plant diversity on semiarid grassland stability depends on functional group composition and dynamics under N and P addition. Science of the Total Environment, 2021, 799: 149482. DOI:10.1016/j.scitotenv.2021.149482 |
| [11] |
苏波, 韩兴国, 黄建辉, 渠春梅. 植物的养分利用效率(NUE)及植物对养分胁迫环境的适应策略. 生态学报, 2000, 20(2): 335-343. DOI:10.3321/j.issn:1000-0933.2000.02.026 |
| [12] |
Güsewell S. N: P ratios in terrestrial plants: variation and functional significance. New Phytologist, 2004, 164(2): 243-266. DOI:10.1111/j.1469-8137.2004.01192.x |
| [13] |
罗芊芊, 周志春, 邓宗付, 喻龙华, 孙建军, 徐刚标. 南方红豆杉天然居群叶片的表型性状和氮磷化学计量特征的变异规律. 植物资源与环境学报, 2021, 30(1): 27-35. DOI:10.3969/j.issn.1674-7895.2021.01.04 |
| [14] |
田地, 严正兵, 方精云. 植物生态化学计量特征及其主要假说. 植物生态学报, 2021, 45(7): 682-713. |
| [15] |
Estiarte M, Campioli M, Mayol M, Penuelas J. Variability and limits of nitrogen and phosphorus resorption during foliar senescence. Plant Communications, 2023, 4(2): 100503. DOI:10.1016/j.xplc.2022.100503 |
| [16] |
Su Y, Ma X F, Le J J, Li K H, Han W X, Liu X J. Decoupling of nitrogen and phosphorus in dominant grass species in response to long-term nitrogen addition in an Alpine Grassland in Central Asia. Plant Ecology, 2021, 222(2): 261-274. DOI:10.1007/s11258-020-01103-3 |
| [17] |
刘艳, 陈梦娇, 郭童童, 崔靖, 李兰平, 梁德飞, 李永慧, 张振华, 朱小雪, 任飞. 多梯度氮磷添加对高寒草甸植物群落生物量与氮磷含量的影响. 草地学报, 2023, 31(3): 751-759. |
| [18] |
Sasaki T, Lauenroth W K. Dominant species, rather than diversity, regulates temporal stability of plant communities. Oecologia, 2011, 166(3): 761-768. DOI:10.1007/s00442-011-1916-1 |
| [19] |
赖帅彬, 徐升, 简春霞, 陈志飞, 周俊杰, 杨全, 王智, 徐炳成. 黄土丘陵区退耕草地群落优势种叶片光合生理对氮磷添加的响应. 生态学报, 2021, 41(13): 5454-5464. |
| [20] |
王洪义, 常继方, 王正文. 退化草地恢复过程中群落物种多样性及生产力对氮磷养分的响应. 中国农业科学, 2020, 53(13): 2604-2613. |
| [21] |
Liang T, Tong Y A, Xu W, Wei Y, Lin W, Pang Y, Liu F, Liu X J. Atmospheric nitrogen deposition in the Loess area of China. Atmospheric Pollution Research, 2016, 7(3): 447-453. DOI:10.1016/j.apr.2015.11.001 |
| [22] |
刘海威, 张少康, 焦峰. 氮磷添加对不同退耕年限草本植被群落及土壤化学计量特征的影响. 水土保持学报, 2017, 31(2): 333-338. |
| [23] |
Grime J P. Benefits of plant diversity to ecosystems: immediate, filter and founder effects. Journal of Ecology, 1998, 86(6): 902-910. DOI:10.1046/j.1365-2745.1998.00306.x |
| [24] |
刘道宏. 植物叶片的衰老. 植物生理学通讯, 1983, 19(2): 14-19. |
| [25] |
Peng H Y, Chen Y H, Yan Z B, Han W X. Stage-dependent stoichiometric homeostasis and responses of nutrient resorption in Amaranthus mangostanus to nitrogen and phosphorus addition. Scientific Reports, 2016, 6: 37219. DOI:10.1038/srep37219 |
| [26] |
高宗宝, 王洪义, 吕晓涛, 王正文. 氮磷添加对呼伦贝尔草甸草原4种优势植物根系和叶片C ∶ N ∶ P化学计量特征的影响. 生态学杂志, 2017, 36(1): 80-88. |
| [27] |
张文彦, 樊江文, 钟华平, 胡中民, 宋璐璐, 王宁. 中国典型草原优势植物功能群氮磷化学计量学特征研究. 草地学报, 2010, 18(4): 503-509. |
| [28] |
卢笑玥, 赵雪, 徐莉萍, 焦子怡, 王晨枫, 欧阳洪梁, 封伦, 邓健. 氮磷添加对黄土丘陵区撂荒草地优势植物功能性状的影响. 中国草地学报, 2023, 45(3): 49-59. |
| [29] |
陈继辉, 李炎朋, 熊雪, 成月, 孙逍. 氮磷添加对草地土壤酸度和化学计量学特征的影响. 草业科学, 2017, 34(5): 943-949. |
| [30] |
Ren F, Song W M, Chen L T, Mi Z R, Zhang Z H, Zhu W Y, Zhou H K, Cao G M, He J S. Phosphorus does not alleviate the negative effect of nitrogen enrichment on legume performance in an alpine grassland. Journal of Plant Ecology, 2017, 10(5): 822-830. |
| [31] |
戚智彦, 吕亚香, 刘伟, 孙佳美, 王璟, 潘庆民. 氮磷养分共同添加促进退化典型草原恢复的机制. 应用生态学报, 2023, 34(1): 75-82. |
| [32] |
侯东杰, 韩蓓蕾, 刘璐, 王忠武. 基于物种周转揭示放牧对荒漠草原植物多样性的影响. 草地学报, 2023, 31(4): 952-962. |
| [33] |
Koerselman W, Meuleman A F M. The vegetation N: P ratio: a new tool to detect the nature of nutrient limitation. The Journal of Applied Ecology, 1996, 33(6): 1441. DOI:10.2307/2404783 |
| [34] |
Neff J C, Townsend A R, Gleixner G, Lehman S J, Turnbull J, Bowman W D. Variable effects of nitrogen additions on the stability and turnover of soil carbon. Nature, 2002, 419(6910): 915-917. DOI:10.1038/nature01136 |
| [35] |
孙小妹, 陈菁菁, 李金霞, 李良, 韩国君, 陈年来. 高寒草甸植物化学计量比对磷添加的分层响应. 植物生态学报, 2018, 42(1): 78-85. |
| [36] |
江大龙, 徐侠, 阮宏华. 植物养分重吸收及其影响研究进展. 南京林业大学学报: 自然科学版, 2017, 41(1): 183-188. |
| [37] |
尤菲菲, 曾令强, 乔子楣, 刘敏国, 杨惠敏. 不同功能群草类植物叶片氮磷重吸收对施肥的响应. 中国草地学报, 2018, 40(3): 55-61. |
| [38] |
Lü X T, Reed S, Yu Q, He N P, Wang Z W, Han X G. Convergent responses of nitrogen and phosphorus resorption to nitrogen inputs in a semiarid grassland. Global Change Biology, 2013, 19(9): 2775-2784. DOI:10.1111/gcb.12235 |
| [39] |
Yu Q, Elser J J, He N P, Wu H H, Chen Q S, Zhang G M, Han X G. Stoichiometric homeostasis of vascular plants in the Inner Mongolia grassland. Oecologia, 2011, 166(1): 1-10. DOI:10.1007/s00442-010-1902-z |
| [40] |
李强, 黄迎新, 周道玮, 丛山. 土壤氮磷添加下豆科草本植物生物固氮与磷获取策略的权衡机制. 植物生态学报, 2021, 45(3): 286-297. |