 2024, Vol. 44
2024, Vol. 44文章信息
- 郭佩琴, 肖佳佳, 郭婉欣, 冷喜文, 孙丽君, 符辉, 袁桂香
- GUO Peiqin, XIAO Jiajia, GUO Wanxin, LENG Xiwen, SUN Lijun, FU Hui, YUAN Guixiang
- 沉水植物性状对不同氨氮脉冲的响应差异
- Plant trait changes of submerged macrophytes in response to ammonium pulses
- 生态学报. 2024, 44(5): 1985-1995
- Acta Ecologica Sinica. 2024, 44(5): 1985-1995
- http://dx.doi.org/10.20103/j.stxb.202304060690
-
文章历史
- 收稿日期: 2023-04-06
- 网络出版日期: 2023-10-25

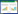




受全球气候变化和人类活动加剧的影响, 未来降水强度和频率发生改变, 极端降水事件增加[1], 这将改变自然界营养物质对植物的供应状态, 促进水溶性营养物质含量增加[2—3]。营养物质含量在短时间内增加的现象称为“营养脉冲”, 营养脉冲的频率和幅度受到降水强度和频率的影响, 营养脉冲现象已在干旱、半干旱生态系统中受到广泛关注[4]。营养的脉冲式变化可以影响植物的生长代谢, 改变物种关系, 影响生态系统功能[5]。在淡水生态系统中, 受湖滨带农业面源污染、旅游开发等影响, 降水对农田、道路、生活区地面等冲刷, 使得大量的营养物质进入湖泊[6], 改变湖泊营养物质状态[7]。然而, 目前有关营养物质的脉冲式变化对沉水植物影响的研究还鲜有报道。
沉水植物是湖泊生态系统中主要生产者之一[8], 它能降低水体营养物质浓度、抑制颗粒物质的再悬浮, 增加水体中的溶解氧含量, 为其它生命体提供食物和避难场所[9], 对维持水生态系统的多样性和稳定性起着重要作用[10—11]。氮是沉水植物生长所需的必要营养元素[12]。氨氮可通过降水径流冲刷、渗透等方式进入水体, 研究发现降水径流会导致水生生态系统中氨氮含量短时间内显著增加[13]。例如, 任玉芬等[14]发现降雨使城市径流中TN含量达到6—8mg/L, 张静等[15]发现首场降雨中的氨氮平均浓度最高达到21mg/L, 是水体氨氮污染的重要来源, 钱海铭等[16]发现强降雨引起的城市点面源污染、污水管网渗漏等造成河流氨氮水平增加三倍左右。沉水植物对氨氮的吸收缺少下调机制, 在高氨氮环境中它们会倾向于吸收超过生长所需要的氮量[17]。氨氮浓度较高时沉水植物的生长代谢受到抑制作用, 导致植物出现氨中毒, 具体表现为叶片发黄、根形态的改变、光合效率的降低以及细胞离子的不平衡和成分的改变等[18]。恒定氨氮浓度对沉水植物生理形态影响已有深入报道, 但沉水植物对氨氮脉冲式变化(浓度×频率)的生理形态响应机制仍不明确。
沉水植物既可以通过叶子从水体中吸收营养, 也可以通过根系从底质中吸收营养满足生长需求[19]。不同底质的理化性质和营养含量存在差异, 对沉水植物生长代谢和生物量分配产生不同的影响[20]。砂土底质具有营养低和透气好等特点, 其中生长的植物通常根系粗壮发达, 以提高对营养物质的吸收, 最大限度的弥补底质营养短缺[21]。较砂土相比, 黏土具有营养丰富、微生物多样等特点, 其中生长的沉水植物具有较高的生物量、株高和叶片数[22—23]。鉴于沉水植物对营养的双重吸收途径(即根和叶子吸收), 推测不同底质类型中的沉水植物对水体氨氮脉冲的响应存在差异。
选取湖泊两种常见优势沉水植物苦草(Vallisneria natans)与黑藻(Hydrilla verticillata)作为研究对象, 它们分别为多年生莲座型和冠层型草本植物[24], 具有生长速度快、环境适应能力强等特点, 在我国长江流域湖泊群中分布广泛[25]。相比黑藻, 苦草根系发达, 根系营养吸收能力强, 同时苦草光饱和点更低, 耐弱光胁迫能力更强[26], 这促进其具有更深的分布水深和更高的群落优势度[27]。研究探讨不同底质(黏土和砂土)、不同氨氮脉冲模式(对照, 高浓度低频率、低浓度高频率)和处理阶段(氨氮脉冲阶段和解除脉冲阶段)对苦草和黑藻生长代谢的影响, 旨在为全球变化背景下湖泊生态系统沉水植物的管理提供参考。我们推测:(1)氨氮脉冲抑制沉水植物的生物量积累, 改变生长形态和生理特征, 不同脉冲模式对植物的影响程度不同;(2)底质类型会影响沉水植物对氨氮脉冲的响应, 营养贫瘠的砂土中沉水植物响应更为敏感。
1 材料与方法 1.1 实验环境与材料实验于2022年5—7月进行, 地点位于湖南农业大学实验基地的透明遮雨棚内(28°11′10″N, 113°4′7″E), 该地区属亚热带季风气候, 气候温和, 雨热同期, 年平均气温16.8—17.3℃, 降雨主要集中在春夏两季。为模拟湖泊中沉水植物自然生长的光照水平, 在实验系统上加盖了一层透光率为50%的遮阳网, 实验系统中50%自然光照大约相当于中营养湖泊1.0m水深以内的水底光照水平[28]。试验所用苦草和黑藻为基地多年培养种苗。选择长势良好且大小一致的植株, 苦草保留7—8片叶片, 地上部分长10cm, 根长5cm;黑藻取15cm长的顶端(无根系), 上端10cm的叶子保留, 下端5cm叶子去掉。将选取的种苗均匀种植于装有黏土底质的实验桶中(直径:90cm, 高度:80cm, 水深60cm), 光照控制为50%, 驯化培养两周。
1.2 实验设计与采样实验选择苦草和黑藻, 并设置两种不同底质(黏土和砂土)。同时设置两个时间阶段, 即氨氮脉冲阶段和解除脉冲阶段。在氨氮脉冲阶段设置了两种脉冲模式(P1、P2)和对照组(CK)。P1(低浓度×高频率)根据实验大桶中水体积加入氯化铵溶液至水体氨氮浓度为3.5mg/L, 每三天半加入一次, 持续时间28d, 一共加8次;P2(高浓度×低频率)根据实验大桶中水体积加入氯化铵溶液至水体氨氮浓度为7.0mg/L, 每7天加入一次, 持续时间28d, 一共加4次, P1和P2组中加入的氨氮总量一致;CK组不添加氨氮。脉冲解除阶段:氨氮脉冲阶段结束后, 停止添加氨氮溶液, 将所有实验桶内的水更换一遍, 并维持植物正常生长28d, 观测其生长情况(图 1)。实验的每个处理组合均重复6次。实验所需底质黏土采自湖南农业大学实验基地内的池塘, 底质中总氮、总磷、总有机质初始含量分别为1.6mg/g、0.86mg/g和19.95mg/g。实验所需砂土为用自来水反复清洗后的细砂。

|
| 图 1 实验设计图解 Fig. 1 Graphical illustration of the experimental design CK:对照组, 不添加氨氮;P1:低浓度×高频率组, 每三天半添加一次氨氮, 一共加8次;P2:高浓度×低频率组, 每7天添加一次氨氮, 一共加4次 |
驯化培养后植株按照上述实验设计均匀种植于实验小桶(直径:26cm, 高度:18cm, 底泥厚度:12cm)。苦草为每小桶种植2株, 共72小桶(2底质×2时间阶段×3脉冲处理×6重复), 总计144株。黑藻为每小桶种植4株, 共72小桶, 总计288株。为使两种沉水植物的初始单位面积的生物量尽量一致, 根据苦草和黑藻植株的初始规格, 每小桶分别种植了2株和4株, 该种植密度与自然水体中两种植物的生物量密度一致[29]。将种有植物的实验小桶放于500L大桶(直径:90cm, 高度:80cm, 水深60cm), 每个大桶中放置4小桶植物, 对应2种不同底质处理和2个处理时间阶段, 每种植株共18大桶(即3种脉冲模式, 每种脉冲模式6个重复)。实验持续时间为56d, 实验过程中共取样两次, 第28d时取第一次样, 第56d时取第二次。
在整个实验期间, 每个大桶中添加5g缓释肥(Osmocote Plus, 15N-10P-12K+2MgO+TE, ICL)满足植物所需的基本营养, 每周随机调整每个大桶里4个小桶的位置, 用软毛刷轻轻刷去叶片表面附着藻类, 实验期间每两周采集每个大桶中的水样进行水质测定, 并于正午测定水体温度。实验期间水体温度范围为25.6—32.8℃。水体中TN、NH4+-N(对照组)、NO3--N、TP和PO4--P含量分别为(0.622±0.007)mg/L、(0.009±0.002)mg/L、(0.471±0.003)mg/L、(0.063±0.002)mg/L和(0.026±0.004)mg/L。
1.3 测定指标及方法(1) 形态指标测定:植株从种植小桶中小心完整取出, 用自来水冲洗干净后, 再用纯净水冲洗2遍, 用天平和刻度尺对每株植物进行生长参数测量, 即单株生物量(g)、地上部分生物量(g)、地下部分生物量(g)、分株数、叶数、株高(cm)和根长(cm), 计算地上与地下生物量的比值(AG∶BG)。株高是指从茎的基部到最长叶的顶端的距离;根长为根的最大长度。
(2) 生理指标测定:样品在60℃烘箱中烘至恒重, 然后磨成粉末状, 测定植物淀粉、可溶性总糖(SC)和游离氨基酸(FAA)含量, 分别采用碘-碘化钾法、蒽酮法和茚三酮法[30]。具体操作为取0.5g左右的研磨好的样品, 用8mL 80%的乙醇提取2次(第一次提取用5mL;第二次3mL), 在80℃的水浴锅中提取20分钟, 再用离心机4200转离心13min后收集上清液, 用活性炭脱色后用滤膜(φ20mm)过滤, 滤液用于测定SC和FAA含量。提取的残渣过夜烘干后加入30%的高氯酸继续萃取90min, 每隔30min摇匀一次, 4200转离心13min, 收集上清液用于测定淀粉含量。
1.4 数据分析统计分析前用Shapiro-Wilk′s进行正态性检验, 用Levene′s方法进行方差齐性检验, 对于不满足方差条件检验的变量进行对数转换。用SPSS 20软件对植物形态和生理性状进行一般线性模型分析。采用Duncan′s多重比较检验各生长性状在不同处理下的差异显著性(P < 0.05)。图中数值表示均值±标准误(SE)。
2 研究结果 2.1 氨氮脉冲对苦草生长形态和生理性状的影响 2.1.1 氨氮脉冲对苦草生长形态的影响在氨氮脉冲阶段, 脉冲模式对苦草生物量和株高有显著影响, 底质类型对苦草AG∶BG有显著影响(表 1, P < 0.05);在解除脉冲阶段, 脉冲模式对苦草的形态性状无显著影响, 底质类型对苦草生物量、根长和AG∶BG有显著影响(表 2, P < 0.05)。
| 植物性状 Plant traits |
脉冲Pulse | 底质Substrate | 脉冲×底质Pulse×Substrate | ||||||||
| df | F | P | df | F | P | df | F | P | |||
| 单株生物量Plant biomass | 2 | 7.324 | 0.003 | 1 | 1.409 | 0.245 | 2 | 3.127 | 0.059 | ||
| 株高Plant height | 2 | 6.524 | 0.005 | 1 | 2.033 | 0.165 | 2 | 1.221 | 0.310 | ||
| 根长Root length | 2 | 0.690 | 0.510 | 1 | 0.370 | 0.548 | 2 | 0.013 | 0.987 | ||
| 地上/地下生物量AG∶BG | 2 | 1.320 | 0.283 | 1 | 10.672 | 0.003 | 2 | 1.483 | 0.244 | ||
| 游离氨基酸FAA | 2 | 4.766 | 0.017 | 1 | 1.316 | 0.261 | 2 | 1.191 | 0.319 | ||
| 可溶性总糖SC | 2 | 3.713 | 0.037 | 1 | 0.002 | 0.963 | 2 | 1.212 | 0.313 | ||
| 淀粉Starch | 2 | 4.899 | 0.015 | 1 | 6.319 | 0.018 | 2 | 0.307 | 0.738 | ||
| AG∶BG:地上/地下生物量Above ground and below ground biomass ratio;FAA:游离氨基酸Free amino acid;SC:可溶性总糖Soluble carbohydrate | |||||||||||
| 植物性状 Plant traits |
脉冲Pulse | 底质Substrate | 脉冲×底质Pulse×Substrate | ||||||||
| df | F | P | df | F | P | df | F | P | |||
| 单株生物量Plant biomass | 2 | 1.989 | 0.156 | 1 | 44.494 | <0.001 | 2 | 0.074 | 0.929 | ||
| 株高Plant height | 2 | 3.222 | 0.055 | 1 | 2.043 | 0.164 | 2 | 1.143 | 0.333 | ||
| 根长Root length | 2 | 2.402 | 0.109 | 1 | 4.834 | 0.036 | 2 | 0.651 | 0.529 | ||
| 地上/地下生物量AG∶BG | 2 | 2.670 | 0.087 | 1 | 30.995 | <0.001 | 2 | 0.100 | 0.905 | ||
| 游离氨基酸FAA | 2 | 25.110 | <0.001 | 1 | 13.347 | 0.001 | 2 | 7.008 | 0.003 | ||
| 可溶性总糖SC | 2 | 22.174 | <0.001 | 1 | 151.053 | <0.001 | 2 | 10.186 | <0.001 | ||
| 淀粉Starch | 2 | 5.924 | 0.007 | 1 | 18.244 | <0.001 | 2 | 0.265 | 0.769 | ||
在脉冲阶段, 黏土P2组生物量和株高显著低于对照组, 砂土P1组生物量和株高显著低于对照组。在解除脉冲阶段, 三种脉冲模式之间的生物量和株高无显著差异(图 2)。另外, 除P1组的AG∶BG外, 黏土和砂土之间的苦草生物量和AG∶BG在脉冲阶段无显著差异, 而在解除脉冲阶段黏土的苦草生物量和AG∶BG显著大于砂土。不同脉冲模式、底质类型和实验阶段对苦草根长无显著影响。结果表明氨氮脉冲会抑制苦草生物量积累, 黏土底质时高浓度脉冲抑制作用强。氨氮脉冲的对生长形态的抑制作用在解除后1个月基本得到恢复。

|
| 图 2 氨氮脉冲模式、底质类型和实验阶段对苦草形态性状的影响 Fig. 2 Effects of ammonium pulse patterns, substrate types and experimental phases on morphological traits of V. natans 不同字母表示处理组存在显著差异(P < 0.05) |
在氨氮脉冲阶段, 脉冲模式对苦草的FAA、SC和淀粉含量有显著影响, 底质类型对淀粉有显著影响(表 1, P < 0.05);在解除脉冲阶段, 脉冲模式和底质阶段均对苦草的FAA、SC和淀粉含量有显著影响, 脉冲模式和底质类型的交互作用对FAA和SC有显著影响(表 2, P < 0.05)。
对于FAA含量, 在脉冲阶段和解除脉冲阶段, 砂土底质中P1和P2组显著增加, 而黏土底质在不同脉冲处理下的FAA含量差异较小。对于SC含量, 氨氮脉冲阶段砂土底质P1和P2组显著低于对照组;解除脉冲阶段P1和P2组高于对照组(图 3)。对于淀粉, 脉冲阶段P1和P2组与对照组无显著差异;解除脉冲阶段砂土P2组低于对照组(图 3)。结果表明脉冲阶段氨氮添加促进FAA积累, 降低了SC, 这种变化在砂土底质的植物中更为明显, 同时氨氮添加促进了淀粉积累。在解除脉冲一个月后苦草代谢物质含量仍与对照组存在差异, 说明氨氮脉冲对苦草生理性状影响持续长久, 这种影响对砂土底质中苦草更为强烈而持久。

|
| 图 3 不同氨氮脉冲模式、底质类型与实验阶段下苦草的碳氮代谢物含量(平均值±标准误, n=6) Fig. 3 Contents (mean±SE, n=6) of carbon and nitrogen metabolites of V. natans under different ammonium pulse patterns, substrate types and experimental phases |
在氨氮脉冲阶段, 不同脉冲模式对黑藻生物量和根长有显著影响(表 3, P < 0.05);在解除脉冲阶段, 脉冲模式对黑藻株高与AG∶BG有显著影响, 底质类型、脉冲模式与底质类型的交互作用对黑藻AG∶BG有显著影响(表 4, P < 0.05)。
| 植物性状 Plant traits |
脉冲Pulse | 底质Substrate | 脉冲×底质Pulse×Substrate | ||||||||
| df | F | P | df | F | P | df | F | P | |||
| 单株生物量Plant biomass | 2 | 3.581 | 0.042 | 1 | 2.487 | 0.127 | 2 | 0.340 | 0.715 | ||
| 株高Plant height | 2 | 2.286 | 0.122 | 1 | 3.236 | 0.084 | 2 | 0.370 | 0.694 | ||
| 根长Root length | 2 | 3.615 | 0.041 | 1 | 0.518 | 0.478 | 2 | 0.246 | 0.784 | ||
| 地上/地下生物量AG∶BG | 2 | 1.390 | 0.267 | 1 | 0.000 | 0.989 | 2 | 1.498 | 0.242 | ||
| 游离氨基酸FAA | 2 | 17.653 | <0.001 | 1 | 7.067 | 0.013 | 2 | 8.541 | 0.001 | ||
| 可溶性总糖SC | 2 | 4.497 | 0.021 | 1 | 2.108 | 0.158 | 2 | 0.143 | 0.867 | ||
| 淀粉Starch | 2 | 0.290 | 0.751 | 1 | 0.780 | 0.385 | 2 | 0.897 | 0.420 | ||
| 植物性状 Plant traits |
脉冲Pulse | 底质Substrate | 脉冲×底质Pulse×Substrate | ||||||||
| df | F | P | df | F | P | df | F | P | |||
| 单株生物量Plant biomass | 2 | 1.700 | 0.203 | 1 | 0.245 | 0.625 | 2 | 2.658 | 0.090 | ||
| 株高Plant height | 2 | 25.398 | <0.001 | 1 | 2.573 | 0.121 | 2 | 0.508 | 0.608 | ||
| 根长Root length | 2 | 2.006 | 0.156 | 1 | 2.709 | 0.112 | 2 | 0.920 | 0.412 | ||
| 地上/地下生物量AG∶BG | 2 | 4.532 | 0.021 | 1 | 50.836 | <0.001 | 2 | 4.632 | 0.019 | ||
| 游离氨基酸FAA | 2 | 2.891 | 0.074 | 1 | 0.033 | 0.857 | 2 | 0.465 | 0.633 | ||
| 可溶性总糖SC | 2 | 0.380 | 0.688 | 1 | 1.947 | 0.175 | 2 | 21.653 | <0.001 | ||
| 淀粉Starch | 2 | 10.550 | <0.001 | 1 | 2.426 | 0.132 | 2 | 1.891 | 0.172 | ||
氨氮脉冲阶段, 较对照组和P1组相比, P2组对黑藻生物量、株高和根长的抑制作用最大, 而对AG∶BG无显著影响。解除脉冲阶段, 除砂土中黑藻生物量, P1和P2组黑藻生物量和株高较对照组下降, 不同脉冲和底质类型对根长无显著影响, 砂土中黑藻AG∶BG显著低于黏土(图 4)。表明高浓度低频率的氨氮脉冲对黑藻形态性状影响最大;在脉冲解除一个月后氨氮仍显著抑制形态生长, 这种抑制作用对黏土中黑藻更为明显。

|
| 图 4 氨氮脉冲模式、底质类型和实验阶段对黑藻形态性状的影响 Fig. 4 Effects of ammonium pulse patterns, substrate types and experimental phases on morphological traits of H. verticillata |
在氨氮脉冲阶段, 脉冲模式对黑藻的FAA和SC含量有显著影响, 底质类型、脉冲模式与底质类型的交互作用对黑藻FAA含量有显著影响(表 3, P < 0.05);在解除脉冲阶段, 脉冲模式对黑藻淀粉含量有显著影响, 脉冲模式与底质类型的交互作用对SC有显著影响(表 4, P < 0.05)。
脉冲阶段, 脉冲处理显著增加了黏土中黑藻FAA含量, 砂土中变化不显著(图 5);P2处理使SC含量显著增加。解除脉冲阶段, FAA含量较脉冲阶段显著降低, 不同脉冲模式下FAA含量无显著差异;黏土中黑藻SC含量在P2处理最大, 而砂土中在P2处理最小。除解除脉冲阶段的砂土中黑藻淀粉含量外, 脉冲模式和底质类型处理对黑藻淀粉含量无显著影响。表明氨氮脉冲促进了FAA和SC积累, 其中P2的影响更大。解除脉冲后, 积累的FAA得到较快恢复, 而SC代谢失衡。

|
| 图 5 不同氨氮脉冲模式、底质类型与实验阶段下黑藻的碳氮代谢物含量(平均值±标准误, n=6) Fig. 5 Contents (mean±SE, n=6) of carbon and nitrogen metabolites of H. verticillata under different ammonium pulse patterns, substrate types and experimental phases |
氨氮是沉水植物正常生长发育必不可少的营养元素, 而水体中的氨氮浓度超过一定水平后, 会产生胁迫作用, 对沉水植物的生长造成抑制, 甚至死亡[31]。研究发现, 与较低浓度高频率相比, 高浓度低频率氨氮脉冲对苦草和黑藻生物量和株高等形态性状的抑制作用更强, 可能是高浓度氨氮对苦草和黑藻产生了更强的胁迫作用。Yuan等[32]对常见3种沉水植物研究也发现高浓度低频率的氨氮脉冲对植物性状连接性的影响更大。通常, 为了缓解氨氮在植物体内的毒性, 大部分植物会通过将氨氮转换成游离氨基酸和有机胺类来减少其在植物体内的积累或者将其转移出植物细胞, 这两个过程分别需要植物碳代谢中间产物提供碳骨架和能量[33—35]。前人研究发现氨氮适当添加可以促进金鱼藻和轮叶黑藻的生长, 而当浓度超过4mg/L时开始抑制生长[36—37]。这与本研究结果是一致的。在氨氮脉冲解除后的一个月, 苦草和黑藻的生物量与对照组无显著差异, 即植株形态生长基本得到恢复。这个结果可能是由于前期脉冲阶段水体中氨氮浓度过高抑制了植物生长, 当脉冲解除后, 水体浓度恢复正常水平, 植物生长得到恢复。
较形态性状相比, 发现苦草和黑藻响应氨氮脉冲的生理性状的变化更为强烈。如砂土中苦草FAA显著增加, SC显著降低;黑藻高氨氮脉冲下FAA和SC显著增加。可溶性碳水化合物是连接光合作用, 呼吸作用和碳水化合物储备的重要中间代谢产物[38], 也是植物氨氮同化所需的底物[39]。通常氨氮胁迫环境下沉水植物体内的游离氨基酸含量增加, 可溶性碳水化合物、淀粉和可溶性蛋白质含量减少[40], 因此SC和FAA的变化能够反映植物生理性状的胁迫状态。研究结果证明两种沉水植物碳和氮代谢的失衡在氨氮脉冲解除一个月后仍没有得到恢复, 说明氨氮脉冲对沉水植物生理性状的影响较为持久。这可能改变沉水植物对营养物质的吸收利用效率, 以及应对外界环境波动时的能量供应效率, 使沉水植物对环境变化更加敏感和脆弱[41]。研究发现沉水植物穗状狐尾藻较高的淀粉和SC储备量有利于其对氮的吸收利用[40], 碳水化合物存储量会影响植物的生存和生长以及受到干扰后的恢复能力[42]。因此, 除了形态性状, 较多的关注沉水植物生理性状对环境波动的响应将有助于我们深入理解沉水植物的生态适应性。
不同物种对氨氮脉冲的响应存在差异。在解除脉冲后, 苦草FAA和SC含量与对照组仍存在显著差异;而黑藻FAA含量恢复至对照组水平, 而SC含量和砂土中黑藻淀粉含量与对照组仍存在显著差异。表明氨氮在苦草中的代谢周转较慢。苦草作为莲座型沉水植物, 具有呼吸速率慢、碳代谢较为保守等特点[43], 使氨氮向FAA转化过程中碳骨架供应较慢, 从而降低了氨氮的周转。相反, 黑藻中FAA含量在解除氨氮脉冲后快速降低, 表明FAA在黑藻中的代谢周转较快。已有研究发现黑藻较苦草具有较大的光合速率和呼吸速率[44], 较快的碳周转为氨氮向FAA转化提供了充足的能量和碳骨架, 而氮的快速周转却打破了植物体内碳的代谢平衡。
较黏土中苦草相比, 砂土中苦草FAA和SC含量变化更大。苦草作为莲座型沉水植物, 根系发达, 根系为其营养吸收的重要途径[45—46]。黏土能够提供较为丰富的营养物质满足苦草生长需求, 相对砂土可能具有更高的代谢周转速率[47]。而砂土中苦草营养物质缺乏, 氨氮脉冲下, 使得原本处于营养饥饿状态的苦草大量吸收了氨氮, 致使其打破了原有的碳氮代谢平衡。朱正杰[48]通过研究水体氨氮浓度与两种营养条件(高:0.11%TN、低:0.04%TN)的底质对沉水植物生长的影响, 发现底质营养的高低不改变苦草对氨氮的耐受浓度。这与研究结果不同, 可能是因为本研究中两种底质类型营养差异较大。相比苦草, 黑藻FAA和SC含量在不同底质类型下的差异较小。这可能是因为黑藻作为冠层型沉水植物, 叶子是其营养吸收的重要途径, 根系营养吸收作用减弱, 在水体中主要起到对植株的固定作用[49], 因此不同底质类型对黑藻生理性状差异影响较小。由此, 可以认为, 沉水植物响应氨氮脉冲的差异与不同物种及其生长的底质类型密切相关, 这种响应差异可能会影响沉水植物的相互作用关系, 进而影响沉水植被群落结构组成和生态功能发挥。
4 结论本文以两种常见的沉水植物苦草和黑藻为研究对象, 分析了这两种植物在不同底质类型下对不同氨氮脉冲模式和处理阶段的植株形态和生理性状响应。研究发现高浓度低频率的氨氮脉冲模式对沉水植物生长的抑制作用更明显, 表明极端强降水事件引起的高浓度氨氮脉冲会对水生态系统造成强大的冲击。相比形态性状, 生理性状对氨氮脉冲响应更快、更敏感且更持久, 因此在全球气候变化背景下, 关注植物生理性状响应显得尤为重要。鉴于植物本身代谢周转速率和适宜底质类型等存在差异, 在氨氮脉冲时, 其对氨氮的吸收利用效率存在差异, 这可能改变沉水植被群落结构的动态组成, 进而影响水生态系统功能发挥。本研究强调了全球气候变化背景下降水强度和频率变化对沉水植物造成的潜在影响。然而, 降水引起水体环境的变化相当复杂, 如光照、磷供给等变化, 这些综合因素对水生植物乃至水生态系统的影响仍有待进一步评估。
| [1] |
Kimutai J, New M, Wolski P, Otto F. Attribution of the human influence on heavy rainfall associated with flooding events during the 2012, 2016, and 2018 March-April-May seasons in Kenya. Weather and Climate Extremes, 2022, 38: 100529. DOI:10.1016/j.wace.2022.100529 |
| [2] |
Kohzu A, Matsuzaki S S, Komuro S, Komatsu K, Takamura N, Nakagawa M, Imai A, Fukushima T. Identifying the true drivers of abrupt changes in ecosystem state with a focus on time lags: Extreme precipitation can determine water quality in shallow lakes. Science of The Total Environment, 2023, 881: 163097. DOI:10.1016/j.scitotenv.2023.163097 |
| [3] |
倪婉敏, 朱蕊, 张建英. 大气氮湿沉降对青山湖富营养化的影响. 环境化学, 2012, 31(5): 631-635. |
| [4] |
谭晓瑶. 氨氮脉冲对沉水植物生长和碳氮代谢的影响[D]. 长沙: 湖南农业大学, 2020.
|
| [5] |
Chesson P, Gebauer R L E, Schwinning S, Huntly N, Wiegand K, Ernest M S K, Sher A, Novoplansky A, Weltzin J F. Resource pulses, species interactions, and diversity maintenance in arid and semi-arid environments. Oecologia, 2004, 141(2): 236-253. DOI:10.1007/s00442-004-1551-1 |
| [6] |
Tan X Y, Yuan G X, Fu H, Peng H, Ge D B, Lou Q, Zhong J Y. Effects of ammonium pulse on the growth of three submerged macrophytes. PLoS One, 2019, 14(7): e0219161. DOI:10.1371/journal.pone.0219161 |
| [7] |
陈洁, 许海, 詹旭, 许笛, 朱广伟, 朱梦圆, 季鹏飞, 康丽娟. 沉积物参与下氮磷脉冲式输入对太湖水体营养盐浓度和藻类生长的影响. 环境科学, 2020, 41(6): 2671-2678. |
| [8] |
Wang X W, Wang Y J, Yao W P, Shangguan L F, Zhang X B, Jin Q J, Cong X, Qian P, Xu Y C. Improving the efficacy of different life-form macrophytes in phytoremediation of artificial eutrophic water by combined planting. Environmental Science and Pollution Research, 2023, 30(25): 67621-67633. DOI:10.1007/s11356-023-27238-0 |
| [9] |
姚程, 胡小贞, 姜霞, 陈俊伊, 王坤. 太湖贡湖湾人工湖滨带水生植物恢复及其富营养化控制. 湖泊科学, 2021, 33(6): 1626-1638. |
| [10] |
郭雅倩, 薛建辉, 吴永波, 周之栋, 华建峰. 沉水植物对富营养化水体的净化作用及修复技术研究进展. 植物资源与环境学报, 2020, 29(3): 58-68. |
| [11] |
Dong B L, Qin B Q, Gao G, Cai X L. Submerged macrophyte communities and the controlling factors in large, shallow Lake Taihu (China): Sediment distribution and water depth. Journal of Great Lakes Research, 2014, 40(3): 646-655. DOI:10.1016/j.jglr.2014.04.007 |
| [12] |
王天佑. 贵州草海沉积物-植物碳氮磷时空分布特征[D]. 贵阳: 贵州师范大学, 2021.
|
| [13] |
Qin H J, Diao M H, Zhang Z Y, Visser P M, Zhang Y Y, Wang Y, Yan S H. Responses of phytoremediation in urban wastewater with water hyacinths to extreme precipitation. Journal of Environmental Management, 2020, 271: 110948. DOI:10.1016/j.jenvman.2020.110948 |
| [14] |
任玉芬, 王效科, 欧阳志云, 侯培强. 北京城市典型下垫面降雨径流污染初始冲刷效应分析. 环境科学, 2013, 34(1): 373-378. |
| [15] |
张静, 周玉文, 王中正, 刘春, 赵树旗, 李伟. 北方城市天然降雨中氨氮污染特征研究. 环境科学与技术, 2016, 39(6): 129-134. |
| [16] |
钱海铭, 张运林, 李娜, 王玮佳, 孙晓, 张毅博, 施坤, 冯胜, 高阳辉. 典型降雨过程中河流饮用水源地水质高频监测研究. 生态环境学报, 2023, 32(3): 579-589. |
| [17] |
Cao T, Xie P, Ni L Y, Wu A P, Zhang M, Wu S K, Smolders A J P. The role of NH4+ toxicity in the decline of the submersed macrophyte Vallisneria natans in lakes of the Yangtze River basin, China. Marine and Freshwater Research, 2007, 58(6): 581-587. DOI:10.1071/MF06090 |
| [18] |
Cao T, Xie P, Ni L Y, Zhang M, Xu J. Carbon and nitrogen metabolism of an eutrophication tolerative macrophyte, Potamogeton crispus, under NH4+ stress and low light availability. Environmental and Experimental Botany, 2009, 66(1): 74-78. DOI:10.1016/j.envexpbot.2008.10.004 |
| [19] |
Lin Q, Gu B, Hong J. Tracking uptake of submerged macrophytes (Ceratophyllum demersum)—Derived nitrogen by cattail (Typha angustifolia) using nitrogen stable isotope enrichments. Ecological Engineering, 2017, 99: 114-1188. DOI:10.1016/j.ecoleng.2016.11.027 |
| [20] |
Xie Y H, An S Q, Yao X, Xiao K Y, Zhang C. Short-time response in root morphology of Vallisneria natans to sediment type and water-column nutrient. Aquatic Botany, 2005, 81(1): 85-96. DOI:10.1016/j.aquabot.2004.12.001 |
| [21] |
Li Z Q, Kong L Y, Yang L F, Zhang M, Cao T, Xu J, Wang Z X, Lei Y. Effect of substrate grain size on the growth and morphology of the submersed macrophyte Vallisneria natans L. Limnologica, 2012, 42(1): 81-85. DOI:10.1016/j.limno.2011.09.003 |
| [22] |
Xu W W, Hu W P, Deng J C, Zhu J G, Zhou N N, Liu X. Impacts of water depth and substrate type on Vallisneria natans at wave-exposed and sheltered sites in a eutrophic large lake. Ecological Engineering, 2016, 97: 344-354. DOI:10.1016/j.ecoleng.2016.10.029 |
| [23] |
王磊, 胡效卿, 张卓伦, 高雨轩, 张新厚, 王国祥. 不同水深和基质下苦草(Vallisneria natans)的生理生态适应策略. 生态学杂志, 2021, 40(8): 2421-2430. |
| [24] |
陈磊, 叶其刚, 潘丽珠, 徐立铭, 黄宏文. 长江中下游湖泊两种混生苦草属植物生活史特征与共存分布格局. 植物生态学报, 2008, 32(1): 106-113. |
| [25] |
符辉, 袁桂香, 曹特, 倪乐意, 张霄林. 水深梯度对苦草(Vallisneria natans)克隆生长与觅食行为的影响. 湖泊科学, 2012, 24(5): 705-711. |
| [26] |
朱丹婷, 李铭红, 乔宁宁. 正交试验法分析环境因子对苦草生长的影响. 生态学报, 2010, 30(23): 6451-6459. |
| [27] |
李启升. 水深和氮负荷对沉水植物生长及其种间竞争关系的影响[D]. 上海: 上海海洋大学, 2019.
|
| [28] |
Yuan G X, Fu H, Zhong J Y, Lou Q, Ni L Y, Cao T. Growth and C/N metabolism of three submersed macrophytes in response to water depths. Environmental and Experimental Botany, 2016, 122: 94-99. DOI:10.1016/j.envexpbot.2015.09.009 |
| [29] |
Yuan G X, Cao T, Fu H, Ni L Y, Zhang X L, Li W, Song X, Xie P, Jeppesen E. Linking carbon and nitrogen metabolism to depth distribution of submersed macrophytes using high ammonium dosing tests and a lake survey. Freshwater Biology, 2013, 58(12): 2532-2540. DOI:10.1111/fwb.12230 |
| [30] |
Zhong C, Cao X C, Bai Z G, Zhang J H, Zhu L F, Huang J L, Jin Q Y. Nitrogen metabolism correlates with the acclimation of photosynthesis to short-term water stress in rice (Oryza sativa L.). Plant Physiology and Biochemistry, 2018, 125: 52-62. DOI:10.1016/j.plaphy.2018.01.024 |
| [31] |
王华, 逄勇, 刘申宝, 马璇. 沉水植物生长影响因子研究进展. 生态学报, 2008(8): 3958-3968. |
| [32] |
Yuan G X, Tan X Y, Guo P Q, Xing K, Chen Z L, Li D B, Yu S Z, Peng H, Li W, Fu H, Jeppesen E. Linking trait network to growth performance of submerged macrophytes in response to ammonium pulse. Water Research, 2023, 229: 119403. DOI:10.1016/j.watres.2022.119403 |
| [33] |
Zhang M, Cao T, Ni L Y, Xie P, Li Z Q. Carbon, nitrogen and antioxidant enzyme responses of Potamogeton crispus to both low light and high nutrient stresses. Environmental and Experimental Botany, 2010, 68(1): 44-50. DOI:10.1016/j.envexpbot.2009.09.003 |
| [34] |
Britto D T, Kronzucker H J. NH4+ toxicity in higher plants: a critical review. Journal of Plant Physiology, 2002, 159(6): 567-584. DOI:10.1078/0176-1617-0774 |
| [35] |
Zhang M, Wang Z Q, Xu J, Liu Y Q, Ni L Y, Cao T, Xie P. Ammonium, microcystins, and hypoxia of blooms in eutrophic water cause oxidative stress and C-N imbalance in submersed and floating-leaved aquatic plants in Lake Taihu, China. Chemosphere, 2011, 82(3): 329-339. DOI:10.1016/j.chemosphere.2010.10.038 |
| [36] |
周金波, 金树权, 包薇红, 罗艳, 胡杨. 不同浓度氨氮对4种沉水植物的生长影响比较研究. 农业资源与环境学报, 2018, 35(1): 74-81. |
| [37] |
颜昌宙, 曾阿妍, 金相灿, 赵景柱, 许秋瑾, 王兴民. 不同浓度氨氮对轮叶黑藻的生理影响. 生态学报, 2007, 27(3): 1050-1055. |
| [38] |
卢姣姣. 高铵、低光与硫化物等多因子胁迫典型沉水植物——苦草的驱动规律模拟研究[D]. 南昌: 南昌大学, 2018.
|
| [39] |
Li H J, Cao T, Ni L Y. Effects of ammonium on growth, nitrogen and carbohydrate metabolism of Potamogeton maackianus A. Benn. Fundamental and Applied Limnology, 2007, 170(2): 141-148. DOI:10.1127/1863-9135/2007/0170-0141 |
| [40] |
Yuan G X, Fu H, Zhong J Y, Cao T, Ni L Y, Zhu T S, Li W, Song X. Nitrogen/carbon metabolism in response to NH4+ pulse for two submersed macrophytes. Aquatic Botany, 2015, 121: 76-82. DOI:10.1016/j.aquabot.2014.12.001 |
| [41] |
常诏峰. 高原沉水植物对铵盐的生理响应研究[D]. 拉萨: 西藏大学, 2021.
|
| [42] |
Rao Q Y, Su H J, Ruan L W, Xia W L, Deng X W, Wang L T, Xu P K, Shen H, Chen J, Xie P. Phosphorus enrichment affects trait network topologies and the growth of submerged macrophytes. Environmental Pollution, 2022, 292(Pt A): 118331. |
| [43] |
Chen J F, Cao T, Zhang X L, Xi Y L, Ni L Y, Jeppesen E. Differential photosynthetic and morphological adaptations to low light affect depth distribution of two submersed macrophytes in lakes. Scientific Reports, 2016, 6: 34028. DOI:10.1038/srep34028 |
| [44] |
苏文华, 张光飞, 张云孙, 肖衡, 夏峰. 5种沉水植物的光合特征. 水生生物学报, 2004, 28(4): 391-395. DOI:10.3321/j.issn:1000-3207.2004.04.010 |
| [45] |
何文凯. 富营养化水体中沉水植被恢复重建影响因子研究——底泥特性对沉水植物生长的影响[D]. 武汉: 武汉大学, 2017.
|
| [46] |
Schuurkes J A A R, Kok C J, Den Hartog C. Ammonium and nitrate uptake by aquatic plants from poorly buffered and acidified waters. Aquatic Botany, 1986, 24(2): 131-146. DOI:10.1016/0304-3770(86)90093-8 |
| [47] |
Li F, Zhu L L, Xie Y H, Jiang L, Chen X S, Deng Z M, Pan B H. Colonization by fragments of the submerged macrophyte Myriophyllum spicatum under different sediment type and density conditions. Scientific Reports, 2015, 5: 11821. DOI:10.1038/srep11821 |
| [48] |
朱正杰. 氨氮与环境因子对沉水植物苦草的综合影响[D]. 南京: 南京大学, 2016.
|
| [49] |
Wang S R, Jin X C, Zhao H C, Wu F C. Phosphate biosorption characteristics of a submerged macrophyte Hydrilla verticillata. Aquatic Botany, 2008, 89(1): 23-26. DOI:10.1016/j.aquabot.2008.01.006 |