 2024, Vol. 44
2024, Vol. 44文章信息
- 李悦, 赵家乐, 樊晓丽, 郑强, 战杜鹃, 陈玉娟, 林植华
- LI Yue, ZHAO Jiale, FAN Xiaoli, ZHENG Qiang, ZHAN Dujuan, CHEN Yujuan, LIN Zhihua
- 端黑萤和边褐端黑萤两性异形和雌性个体生育力
- Sexual dimorphism and female reproductive outputs of the Abscondita chinensis and Abscondita terminalis
- 生态学报. 2024, 44(5): 2127-2135
- Acta Ecologica Sinica. 2024, 44(5): 2127-2135
- http://dx.doi.org/10.20103/j.stxb.202303040399
-
文章历史
- 收稿日期: 2023-03-04
- 网络出版日期: 2023-12-11





2. 丽水学院生态学院, 丽水 323000
2. College of Ecology, Lishui University, Lishui 323000, China
两性异形在动物界普遍存在, 是指动物的个体大小、体色、局部特征等方面的两性差异[1]。其中个体大小的两性异形有三种类型, 分别为雌性偏大、雄性偏大和雌雄大小相似[2]。哺乳动物主要为雄性偏大的两性异形, 如北象海豹(Mirounga angustirostris)、北海狗(Callorhinus ursinus)[3—4], 有少数物种也报道了雌性偏大的两性异形, 如大食蚁兽(Myrmecophaga tridactyla)[5]。蝙蝠以雌性偏大的两性异形为主, 也有少数物种雄性偏大, 如锤头果蝠(Hypsignathus monstrosus)[6]。大多数鸟类的两性异形类型与婚配制度有关, 婚配制度为一夫一妻制或一夫多妻制, 表现为雄性偏大的两性异形, 如草地鹨属(Sturnella);当物种的婚配制度为一妻多夫制, 表现为雌性偏大的两性异形;但滨海鸟类除环颈鸻(Charadrius alexandrines)以外的以一妻多夫为婚配制度的鸟类表现为雌性偏大的两性异形[7—8]。在爬行动物中, 蜥蜴类主要是雄性偏大的两性异形, 如宽头石龙子(Eumeces laticeps)、西方鞭尾蜥(Cnemidophorus tigris)[9—10]。蛇类以雌性偏大的两性异形为主[11], 一项对8个科374种蛇的研究报道, 有243种蛇类属于雌性偏大的两性异形, 124种蛇属于雄性偏大的两性异形, 7种蛇类雌雄大小相似[12]。龟鳖类两性异形的种间差异较大, 一项对9个科135种海龟的研究表明, 半水生和陆栖种类是雄性偏大的两性异形, 水生种类为雌性偏大的两性异形[13]。与蜥蜴类和蛇类相比, 龟鳖类雌雄体型差异程度更大[14]。鳄类两性异形的研究较少, 多数鳄类是雄性偏大的两性异形, 如鳄科(Crocodylids)、短吻鳄科(Alligatorids), 但是少数体型较小的种类, 如侏儒鳄(Osteolaemus tetraspis)是雌性偏大的两性异形[2, 15]。两栖类以雌性偏大的两性异形为主, 如无尾目中90%的种类和有尾目中61%的种类, 蚓螈目中雌雄大小相似的种类占多数[16]。不同类型两性异形也同样存在于鱼类中, 雌性偏大的种类如食蚊鱼(Gambusia affinis);雄性偏大的种类如凯利贝(Lamprologus callipterus)、胡子鲶(Clarias fuscus);雌雄大小相似的种类如温州厚唇鱼(Acrossocheilus wenchowensis)[17—20]。无脊椎动物昆虫纲主要为雌性偏大的两性异形[21—22], 一项研究报道, 昆虫纲每个目中72%—95%的物种表现出雌性偏大的两性异形, 一个例外是蜻蜓目, 只有27%的物种雌性偏大[23]。
雌性偏大的两性异形与生育力选择、性成熟时间、移动性假说、吉塞林-赖斯小雄性假说有关。根据生育力选择, 雌性偏大有利于提高繁殖输出, 如花鱼骨(Hemibarbus maculatus)、芒果切叶象甲虫(Deporaus marginatus)[24—26]。两栖动物体型大小与性成熟时间有关, 雄性的性成熟时间早于雌性, 故雄性将更多的能量分配给精子的形成和求偶鸣叫, 限制了雄性体型生长, 如北仑姬蛙(Microhyla beilunensis)[27—28]。移动性假说表明, 较小的雄性在争夺配偶时更受青睐, 后肢占身体的比例更大、身体更小的雄性更加灵活, 活动范围更大, 有更高的受精成功率, 较小的雄性从运动性能中获益, 如沙螽(Deinacrida rugosa)[29]、叶虫(Phyllium philippinicum)[30]。吉塞林-赖斯小雄性假说表明了觅食与寻找配偶之间的时间分配权衡, 体型较小的雄性需要较少的能量支持其活动, 所以进食的时间较少, 因此有较多时间寻找配偶[30]。这种假说已在水黾(Aquarium paludum)得到证实, 较小的雄性动物交配成功率更高[31]。雄性偏大的两性异形与性选择有关, 体型较大的雄性具有更高抢夺资源、求偶、占有领地等方面的适合度, 追逐配偶和繁殖的成功率较高, 在雄性竞争中不断强化[32—33], 例如牙买加变色蜥(Anolis garmani)[34]。
萤火虫属于昆虫纲鞘翅目萤科, 分布在温带、亚热带和热带地区, 已知全世界共有2000多种, 分别属于8个亚科92属和亚属[35], 腹部末端的发光器能发黄绿色光[36]。萤火虫对栖息地的环境质量要求较高, 故可作为生态环境质量的指示生物[37]。端黑萤(Abscondita chinensis)和边褐端黑萤(Abscondita terminalis)隶属于萤科棘手萤属, 鞘翅为橙黄色, 其末端黑色, 雄萤有2节发光器, 雌萤仅有1节发光器[38]。端黑萤和边褐端黑萤的外部形态相似, 其鉴定方法如下:端黑萤的雄萤的第5腹节整个腹板为黑色, 雌萤为不相连的对称黑斑, 而边褐端黑萤的雄萤和雌萤的第3、4和5腹节两侧均有不相连的逐渐变大的对称黑斑[39]。端黑萤多发生在森林中, 边褐端黑萤多发生在稻田[39]。当前国内外萤火虫的研究主要集中在种群分布、生活史、闪光模式和行为, 以及线粒体基因组等内容[37, 40—41], 本文通过检测端黑萤和边褐端黑萤繁殖期形态特征的两性异形和雌性个体生育力, 来评估同属相似物种的两性异形程度和个体生育力的种间差异及相关性, 从而探讨出两性异形的进化和选择压力, 为萤火虫的形态、繁殖生态学提供基础数据。
1 材料与方法 1.1 实验动物的采集2022年7月2日—8月7日, 从浙江省丽水市13个地点(采样点经纬度信息见图 1)捕获端黑萤(N=340)和边褐端黑萤(N=126), 带回实验室进行形态测定。

|
| 图 1 采样点经纬度信息 Fig. 1 Information on latitude and longitude of sampling points |
根据腹部发光器鉴定性别, 雄萤的腹部末端具有2节乳白色发光器, 第一节为长椭圆形, 第二节为半圆形, 雌萤只有一节发光器。用Sony DSC-T100数码相机拍摄以厘米纸作为标尺的萤火虫背面和腹面形态, 在电脑上运用ImageJ 1.44p软件测定如下指标:背面的前胸背板宽(前胸背板末端两侧之间的距离, ± 0.001 mm)和鞘翅长(右鞘翅中间侧的长度);腹面的全长(从复眼前端到躯干末端之间的距离)、复眼宽(左右两只复眼的最大宽度)、触角长(右侧触角基部到末端之间的距离)、胸长(前足与后足基部之间距离)、腹长(从第一腹节到身体末端的距离)和腹宽(第6腹节的宽度, 腹部最宽之处的距离);雌萤和雄萤的发光器面积。用Sartorius分析天平分别称取体重(± 0.001 g)。
1.3 雌萤的生育力将野外采集的雌萤放入有泥土和潮湿苔藓覆盖的培养皿中, 待雌萤自行产卵后, 对产在苔藓上的卵逐个计数, 同时用毛笔转移至有刻度尺标签的白纸上拍照, 利用相同方法测量每只雌萤的6—30枚卵, 取卵径平均值。若在第3天苔藓中未发现雌萤产卵, 经解剖发现体内无卵时, 认定为雌萤已在野外产卵。
1.4 数据分析数据的统计分析用Statistic 5.0统计软件包完成。数据在作进一步统计分析前进行正态性和方差同质性检验。经检验, 数据无需转换可直接进行参数统计。用两因素方差分析(Two-way ANOVA)、单因素协方差分析(One-way ANCOVA)、两因素协方差分析(Two-way ANCOVA)及后续的Tukey′s检验、主成分分析、线性回归等处理和比较相应的数据, 比较矫正平均值前检验斜率的均一性。文中涉及的非参数统计为G-检验。两性异形指数(SDI)[42]计算公式如下: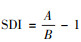
共检测端黑萤个体340只, 其中雌性个体97只, 雄性个体243只, 雌性个体的数量显著少于雄性个体的数量(G-检验, G=64.78, df=1, P<0.001)。共检测边褐端黑萤个体126只, 其中雌性个体22只, 雄性个体104只, 雌性个体的数量显著小于雄性个体的数量(G-检验, G=57.97, df=1, P<0.001)。
端黑萤和边褐端黑萤个体大小和形态特征的基本描述性统计见表 1。Two-way ANOVA显示, 端黑萤的全长显著小于边褐端黑萤的全长, 雌性个体显著大于雄性个体, 物种与性别的相互作用对全长影响不显著(表 1)。以全长为协变量的Two-way ANCOVA显示, 端黑萤的前胸背板宽、鞘翅长、复眼宽、胸长、腹宽、发光器面积及体重均显著小于边褐端黑萤, 腹长显著大于边褐端黑萤, 触角长物种间差异不显著;雌性的前胸背板宽、腹长、腹宽及体重均显著大于雄性, 鞘翅长、复眼宽、胸长、触角长及发光器面积均显著小于雄性个体;物种和性别的交互作用对前胸背板宽、复眼宽、腹长、触角长及发光器面积影响显著, 对其余的鞘翅长、胸长、腹宽及体重影响不显著(表 1)。根据全长数据, 端黑萤的两性异形指数为0.144, 边褐端黑萤的两性异形指数为0.091。
| 形态特征 Morphological characteristics |
端黑萤 Abscondita chinensis |
边褐端黑萤 Abscondita terminalis |
F值和显著水平 F-values and significant levels |
||||||
| 雌性成体 Adult females (n=97) |
雄性成体 Adult males (n=243) |
雌性成体 Adult females (n=22) |
雄性成体 Adult males (n=104) |
物种 Species |
性别 Sex |
交互作用 Interaction |
|||
| 全长/mm Total length |
9.83±0.10 7.40—11.99 |
8.59±0.04 6.84—10.60 |
10.88±0.23 8.53—13.32 |
9.97±0.06 8.29—11.22 |
F1, 460= 153.35** P<0.001, ch<te |
F1, 460= 118.35** P<0.001, F>M |
F1, 460= 2.92 P=0.088 |
||
| 前胸背板宽/mm Pronotum width |
2.65±0.02 2.20—3.21 |
2.26±0.01 1.85—3.05 |
2.92±0.05 2.45—3.32 |
2.68±0.02 2.22—3.09 |
F1, 457= 76.09** P<0.001, ch<te |
F1, 457= 67.29** P<0.001, F>M |
F1, 457= 5.90* P<0.016 |
||
| 鞘翅长/mm Elytron length |
7.09±0.05 5.97—8.52 |
6.51±0.03 5.27—7.97 |
8.06±0.13 6.58—9.10 |
7.47±0.03 6.22—8.14 |
F1, 457= 98.09** P<0.001, ch<te |
F1, 457= 15.26** P<0.001, F<M |
F1, 457= 2.06 P=0.152 |
||
| 复眼宽/mm Compound eye width |
1.56±0.01 1.32—1.85 |
1.93±0.01 1.47—2.31 |
1.73±0.03 1.44—1.92 |
2.22±0.01 1.91—2.45 |
F1, 459= 68.71** P<0.001, ch<te |
F1, 459= 1131.88** P<0.001, F<M |
F1, 459= 11.70** P<0.001 |
||
| 胸长/mm Thorax length |
4.24±0.03 3.39—4.98 |
3.72±0.02 2.16—4.82 |
4.80±0.09 3.90—5.50 |
4.37±0.02 3.68—4.90 |
F1, 456= 44.07** P<0.001, ch<te |
F1, 456= 25.06** P<0.001, F<M |
F1, 456= 0.05 P=0.817 |
||
| 腹长/mm Abdomen length |
4.99±0.07 3.61—6.52 |
3.95±0.03 3.03—5.12 |
5.09±0.14 3.81—6.67 |
4.40±0.04 3.30—5.20 |
F1, 457= 83.10** P<0.001, ch>te |
F1, 457= 43.65** P<0.001, F>M |
F1, 457= 4.64* P<0.032 |
||
| 腹宽/mm Abdomen width |
2.80±0.02 2.26—3.37 |
2.33±0.01 1.69—3.13 |
3.25±0.07 2.46—3.94 |
2.79±0.02 2.28—3.17 |
F1, 458= 113.35** P<0.001, ch<te |
F1, 458= 154.64** P<0.001, F>M |
F1, 458= 0.56 P=0.459 |
||
| 触角长/mm Antenna length |
3.69±0.04 2.62—4.40 |
3.62±0.04 1.89—4.77 |
3.91±0.12 2.80—4.62 |
4.18±0.05 2.87—5.04 |
F1, 449= 1.74 P=0.188 |
F1, 449= 25.19** P<0.001, F<M |
F1, 449= 4.02* P<0.045 |
||
| 发光器面积/mm2 Photogenic organ area |
1.54±0.03 1.00—2.51 |
3.51±0.03 2.44—5.44 |
1.97±0.07 1.35—2.50 |
4.47±0.05 2.97—5.79 |
F1, 457= 16.03** P<0.001, ch<te |
F1, 457= 2335.61** P<0.001, F<M |
F1, 457= 17.25** P<0.001 |
||
| 体重/g Body weight |
0.02±0.00 0.01-0.04 |
0.01±0.00 0.00-0.03 |
0.02±0.00 0.01-0.04 |
0.02±0.00 0.01-0.03 |
F1, 452= 19.92** P<0.001, ch<te |
F1, 452= 46.95** P<0.001, F>M |
F1, 452= 0.04 P=0.849 |
||
| 用平均值±标准误(范围)表示描述性统计值, 表中显示Two-way ANOVA(全长)或ANCOVA(其余形态变量以全长为协变量)的F值和显著性水平, α设为0.05, 显著性差异P<0.05用*表示, P<0.001用* *表示。ch:端黑萤Abscondita chinensis;te:边褐端黑萤Abscondita terminalis;F:成年雌体Female;M:成年雄体male | |||||||||
9个形态特征变量的主成分分析(Eigen value≥1)发现, 前2个主成分共解释56.8%的变异(表 2)。复眼宽、发光器面积在第一主成分有较高的负负载系数, 腹长有较高的正负载系数(解释31.3%变异);前胸背板宽、鞘翅长在第二主成分有较高的负负载系数(解释25.5%变异(表 2)。
| 形态特征 Morphological characteristics |
负载系数Factor loading | |
| 第一主成分 First principal component (PC1) |
第二主成分 Second principal component (PC2) |
|
| 前胸背板宽Pronotum width/mm | 0.204135 | -0.767411 |
| 鞘翅长Elytron length/mm | -0.052370 | -0.821274 |
| 复眼宽Compound eye width/mm | -0.899181 | -0.208476 |
| 胸长Thorax length/mm | -0.009641 | -0.602586 |
| 腹长Abdomen length/mm | 0.711982 | 0.101442 |
| 腹部宽Abdomen width/mm | 0.271278 | -0.676369 |
| 触角长Antenna length/mm | -0.389657 | -0.312313 |
| 发光面积Photogenicorgan area/mm2 | -0.884645 | -0.074417 |
| 体重Body weight/g | 0.668291 | -0.233267 |
| 解释变异Variance explained | 31.3% | 25.5% |
| 用变量与全长的回归剩余值去除大小差异的影响 | ||
物种间和雌雄两性间在第一和第二的分值差异显著性见表 3。Two-way ANOVA显示, 端黑萤在第一主成分和第二主成分的分值均显著大于边褐端黑萤;雌性个体在第一主成分显著大于雄性个体, 在第二主成分差异不显著;两因素的相互作用对分值的影响均不显著。以第一和第二主成分分值分别为X和Y轴, 端黑萤和边褐端黑萤的雌雄两性的空间位置见图 2。
| 主成分分析的分轴 Axes of Principal component analysis |
物种 Species |
性别 Sex |
交互作用 Interaction |
| 第一主成分 First principal component (PC1) |
F1, 440= 29.50** P<0.001, ch>te |
F1, 440= 90.94** P<0.001, F>M |
F1, 440= 3.82 P =0.051 |
| 第二主成分 Second principal component (PC2) |
F1, 440= 50.64** P<0.001, ch>te |
F1, 440= 1.67 P=0.197 |
F1, 440= 0.01 P=0.908 |

|
| 图 2 端黑萤和边褐端黑萤两性成体在由9个形态特征变量区分出的第一和第二主成分轴上的空间位置 Fig. 2 The spatial positions of Abscondita chinensis and Abscondita terminalis adults on the first and second principal component axes distinguished by 9 morphological characteristic variables 个体大小差异的影响用相应变量与全长的回归剩余值去除; 放大实心符号代表前二个分轴上分值的平均值 |
本实验检测的端黑萤雌性个体97只, 31只已产, 共收集66窝。窝卵数在10—81粒之间, 平均为28.9粒。回归分析表明, 端黑萤的窝卵数与全长(r2= 0.28, F1, 65=25.47, P<0.001)和体重(r2= 0.29, F1, 63= 26.23, P<0.001)回归关系显著(图 3, 图 4)。

|
| 图 3 端黑萤和边褐端黑萤的窝卵数与母体全长之间的回归关系 Fig. 3 The linear regression of clutch on total length of maternal Abscondita chinensis and Abscondita terminalis 端黑萤: 圆点、空心、虚线; 边褐端黑萤: 圆点、实心、实线 |

|
| 图 4 端黑萤和边褐端黑萤的窝卵数与母体体重之间的回归关系 Fig. 4 The linear regression of clutch on body mass of maternal Abscondita chinensis and Abscondita terminalis |
本实验检测的边褐端黑萤雌性个体22只, 4只已产, 共收集18窝。窝卵数在6—38粒之间, 平均为18粒。回归分析表明, 边褐端黑萤的窝卵数与全长(r2= 0.41, F1, 16= 10.94, P<0.004)和体重(r2= 0.61, F1, 16= 24.85, P<0.001)回归关系显著(图 3, 图 4)。
One-way ANCOVA及后续的矫正平均值Tukey′s检验显示, 特定全长的端黑萤的窝卵数显著大于边褐端黑萤(F1, 82= 25.56, P<0.001)。
3 讨论 3.1 雌萤体长大于雄萤是生育力选择、能量分配、运动综合影响的结果雌性偏大的两性异形是昆虫纲个体大小两性异形的主要方式[43], 本研究结果表明, 浙江丽水地区的端黑萤和边褐端黑萤两性异形方式也属于上述类型。雄性偏大的两性异形通常归因于性选择, 即雄性之间通过格斗竞争配偶和资源, 雌性偏大的两性异形通常归因于自然选择倾向于增加生育力和繁殖输出[44]。在形态检测过程中, 未发现雄萤存在与格斗有关的角、毒腺等结构, 野外调查时也未发现雄萤参与护卵、抚育后代的行为, 这排除了性选择影响雄萤向大个体方向进化的趋势。萤火虫的繁殖潜力很大程度上取决于交配次数, 野外捕捉的端黑萤雌雄性比约为1 ∶ 2.51, 边褐端黑萤雌雄性比约为1 ∶ 4.73, 雌萤数量远少于雄萤, 在自然环境中, 雌萤面临更大的生存压力, 如更易于被天敌捕食, 这可以说明两物种通过提高繁殖场所雄性个体的密度进而提高交配成功率, 因此克服体型小所导致的劣势。
在昆虫中, 体型大小通常被视为雌性个体生育力的重要决定因素, 对不同目、科的许多昆虫物种的研究表明, 昆虫的生育力与成虫大小呈显著的正相关[45]。根据生育力选择, 物种的生育力取决于雌性个体的体型大小, 体型较大的雌性个体通常拥有更多的卵巢, 产卵量更多, 同时摄食和吸收营养物质更快, 具有更多的营养物质, 通过分配这些额外的营养物质也可以影响后代的数量或质量[46]。本研究显示, 雌萤的腹部长度和宽度显著大于雄萤, 因此雌萤的腹腔容量更大, 这为生育力选择解释端黑萤和边褐端黑萤大个体雌性和大的腹腔容量的形成提供可能。
从成虫的能量分配来看, 雄萤偏小可能与萤火虫的觅食与寻找配偶之间的能量分配权衡有关, 较小的雄萤需要较少的能量支持其活动, 进食时间较少, 因此将更多的能量用于寻找配偶[47]。从运动的灵活性来看, 雄萤的复眼宽、触角长和鞘翅长等与感觉和运动有关的局部形态比雌萤大, 为雄萤在空中飞行寻找雌萤提供了优势;此外, 体型较小的雄萤承担的运动成本较少, 灵活性高, 每晚活动范围更大, 具有更高的交配成功率, 在争夺配偶时更具有优势[47]。从运动的稳定性来看, 首先, 在空中飞行的昆虫, 悬停或缓慢向前飞行时, 需要在空中保持稳定, 更大和更重的体型会产生更多的空气阻力, 翼载荷(体重和翼面积比值)升高, 不利于昆虫的飞行[48]。其次, 为了安全附着在树叶上, 昆虫必须克服冲击力, 体型较小的雄萤体重相对较轻, 在空中降落后能够更快地稳定身体角度, 着陆时易于抓住树枝或树叶, 体型较大的雄萤飞行能力相对较差, 也更容易从光滑的物体表面掉落, 一旦坠落, 可能被捕食者发现, 若重新返回树冠也会消耗额外的能量, 甚至失去配偶[49—50]。因此, 研究发现有助于阐明相对较小的雄萤在争夺竞争交配系统中的重复进化, 明确地将运动视为昆虫的形态、灵活性和交配成功之间的中间步骤。
3.2 端黑萤和边褐端黑萤的两性异形差异程度与生育力成正相关目前为止, 对动物种内个体的两性异形与生育力的关系的分析较多[19, 24], 对种间两性异形的比较分析较少[51]。由于不同物种的体型大小存在显著的差异, 因此, 在种间比较时, 更关注的是两性异形指数的种间差异与物种的生育力(窝卵数)的关系。本研究的数据表明, 端黑萤的两性异形指数为0.144, 边褐端黑萤的两性异形指数为0.091, 端黑萤的两性异形指数比边褐端黑萤约大50%, 端黑萤的全长显著小于边褐端黑萤, 而窝卵数显著大于边褐端黑萤, 本研究推测, 同属相似种端黑萤和边褐端黑萤种间两性异形的差异程度与窝卵数呈正相关。
综上所述, 端黑萤和边褐端黑萤均属于雌性偏大的两性异形, 是生育力选择、能量分配和运动综合影响的结果。雌萤腹腔容量等关键局部特征的增大是对生育力选择的适应。通过种间比较得出, 物种的生育力与两性异形差异程度成正相关。
| [1] |
Temeles E J, Pan I L, Brennan J L, Horwitt J N. Evidence for ecological causation of sexual dimorphism in a hummingbird. Science, 2000, 289(5478): 441-443. DOI:10.1126/science.289.5478.441 |
| [2] |
廖灏泓, 徐峰, 杨维康. 两栖爬行动物两性大小异形研究进展. 生态学杂志, 2013, 32(11): 3082-3093. |
| [3] |
Moore S L, Wilson K. Parasites as a viability cost of sexual selection in natural populations of mammals. Science, 2002, 297(5589): 2015-2018. DOI:10.1126/science.1074196 |
| [4] |
Janicke T, Fromonteil S. Sexual selection and sexual size dimorphism in animals. Biology Letters, 2021, 17(9): 20210251. DOI:10.1098/rsbl.2021.0251 |
| [5] |
Naples V L. Morphology, evolution and function of feeding in the giant anteater (Myrmecophaga tridactyla). Journal of Zoology, 1999, 249(1): 19-41. DOI:10.1111/j.1469-7998.1999.tb01057.x |
| [6] |
吴慧, 江廷磊, 冯江. 蝙蝠体型性别二态性研究现状与展望. 兽类学报, 2014, 34(3): 313-322. |
| [7] |
Székely T, Lislevand T, Figuerola J. Sexual size dimorphism in birds. Wilson Bulletin, 2007, 89(4): 619-620. |
| [8] |
Jehl J, Murray B G. The evolution of normal and reverse sexual size dimorphism in shorebirds and other birds. Current Ornithology, 1986, 1-86. |
| [9] |
Vitt L J, Cooper W E Jr. The evolution of sexual dimorphism in the skink Eumeces laticeps: an example of sexual selection. Canadian Journal of Zoology, 1985, 63(5): 995-1002. DOI:10.1139/z85-148 |
| [10] |
Anderson R A, Vitt L J. Sexual selection versus alternative causes of sexual dimorphism in teiid lizards. Oecologia, 1990, 84(2): 145-157. DOI:10.1007/BF00318265 |
| [11] |
Ji X A, Xie Y Y, Sun P Y, Zheng X Z. Sexual dimorphism and female reproduction in a viviparous snake, Elaphe rufodorsata. Journal of Herpetology, 1997, 31(3): 420. DOI:10.2307/1565671 |
| [12] |
Shine R. Sexual size dimorphism in snakes revisited. Copeia, 1994, 1994(2): 326. DOI:10.2307/1446982 |
| [13] |
Gosnell J S, Rivera G, Blob R. A phylogenetic analysis of sexual size dimorphism in turtles. Herpetologica, 2009, 65(1): 70-81. DOI:10.1655/07-057R2.1 |
| [14] |
Agha M, Ennen J R, Nowakowski A J, Lovich J E, Sweat S C, Todd B D. Macroecological patterns of sexual size dimorphism in turtles of the world. Journal of Evolutionary Biology, 2018, 31(3): 336-345. DOI:10.1111/jeb.13223 |
| [15] |
Luiselli L, Akani G C, Capizzi D. Is there any interspecific competition between dwarf crocodiles (Osteolaemus tetraspis) and Nile monitors (Varanus niloticus ornatus) in the swamps of central Africa? A study from south-eastern Nigeria. Journal of Zoology, 1999, 247(1): 127-131. DOI:10.1111/j.1469-7998.1999.tb00200.x |
| [16] |
Shine R. Sexual selection and sexual dimorphism in the Amphibia. Copeia, 1979, 1979(2): 297. DOI:10.2307/1443418 |
| [17] |
樊晓丽, 林植华, 胡雄光, 雷焕宗, 李香. 卵胎生入侵种食蚊鱼的两性异形和雌性繁殖输出. 生态学报, 2016, 36(9): 2497-2504. |
| [18] |
樊晓丽, 林植华, 丁先龙, 朱吉峰. 鲶鱼和胡子鲶的两性异形与雌性个体生育力. 生态学报, 2014, 34(3): 555-563. |
| [19] |
Takahashi T, Ota K, Kohda M, Hori M. Some evidence for different ecological pressures that constrain male and female body size. Hydrobiologia, 2012, 684(1): 35-44. DOI:10.1007/s10750-011-0961-4 |
| [20] |
徐德钦, 林植华, 雷焕宗. 温州厚唇鱼形态特征的两性异形和雌性个体生育力. 上海交通大学学报: 农业科学版, 2006, 24(4): 335-340. |
| [21] |
Boisseau R P, Ero M M, Makai S, Bonneau L J G, Emlen D J. Sexual dimorphism divergence between sister species is associated with a switch in habitat use and mating system in thorny devil stick insects. Behavioural Processes, 2020, 181: 104263. DOI:10.1016/j.beproc.2020.104263 |
| [22] |
Bank S, Bradler S. A second view on the evolution of flight in stick and leaf insects (Phasmatodea). BMC Ecology and Evolution, 2022, 22(1): 62. DOI:10.1186/s12862-022-02018-5 |
| [23] |
Stillwell R C, Blanckenhorn W U, Teder T, Davidowitz G, Fox C W. Sex differences in phenotypic plasticity affect variation in sexual size dimorphism in insects: from physiology to evolution. Annual Review of Entomology, 2010, 55: 227-245. DOI:10.1146/annurev-ento-112408-085500 |
| [24] |
林植华, 雷焕宗, 林植云, 华和亮. 花的两性异形和雌体繁殖输出. 上海交通大学学报: 农业科学版, 2005, 23(3): 284-288. DOI:10.3969/j.issn.1671-9964.2005.03.016 |
| [25] |
柏天琦, 解德宏, 陈于福, 张翠仙, 王永芬, 尼章光, 王美存, 章勇. 芒果切叶象甲虫体大小和形状的两性异形分析. 植物保护, 2022, 48(2): 188-195, 200. |
| [26] |
De Block M, Stoks R. Adaptive sex-specific life history plasticity to temperature and photoperiod in a damselfly. Journal of Evolutionary Biology, 2003, 16(5): 986-995. DOI:10.1046/j.1420-9101.2003.00581.x |
| [27] |
Cadeddu G, Giacoma C, Castellano S. Sexual size dimorphism in the Tyrrhenian tree frog: a life-history perspective. Journal of Zoology, 2012, 286(4): 285-292. DOI:10.1111/j.1469-7998.2011.00878.x |
| [28] |
陈智强, 钟俊杰, 冯磊, 郑伟成, 丁国骅, 吴延庆, 林植华. 浙江九龙山发现北仑姬蛙新种群的两性异形. 动物学杂志, 2020, 55(2): 178-188. |
| [29] |
Herberstein M E, Painting C J, Holwell G I. Scramble competition polygyny in terrestrial arthropods. Advances in the Study of Behavior. Amsterdam: Elsevier, 2017: 237-295.
|
| [30] |
Boisseau Romain P, Büscher Thies H, Klawitter Lexi J, Gorb Stanislav N, Emlen Douglas J, Tobalske Bret W. Multi-modal locomotor costs favor smaller males in a sexually dimorphic leaf-mimicking insect. BMC Ecology and Evolution, 2022, 22(1): 39. DOI:10.1186/s12862-022-01993-z |
| [31] |
Blanckenhorn W U, Preziosi R F, Fairbairn D J. Time and energy constraints and the evolution of sexual size dimorphism-to eat or to mate?. Evolutionary Ecology, 9(4): 369-381. DOI:10.1007/BF01237760 |
| [32] |
Pyron M, Pitcher T E, Jacquemin S J. Evolution of mating systems and sexual size dimorphism in North American cyprinids. Behavioral Ecology and Sociobiology, 2013, 67(5): 747-756. DOI:10.1007/s00265-013-1498-5 |
| [33] |
Kelly C D. Sexual selection on size and shape in Japanese beetles (Popillia japonica). Behavioral Ecology, 2020, 31(4): 1073-1083. DOI:10.1093/beheco/araa054 |
| [34] |
Trivers R L. Sexual selection and resource-accruing abilities in Anolis garmani. Evolution, 1976, 30(2): 253. |
| [35] |
McDermott F A. The taxonomy of the Lampyridae (Coleoptera). Transactions of the American Entomological Society, 1964, 90(1): 1-72. |
| [36] |
何华, 刘玉升, 王振鹏, 张秀波. 台湾地区萤火虫资源及其开发利用情况. 华东昆虫学报, 2007, 16(2): 156-158. |
| [37] |
王郡明, 梁醒财, 罗佑珍. 萤火虫生物学特性及其应用研究. 云南农业大学学报, 2006, 21(5): 576-580. DOI:10.3969/j.issn.1004-390X.2006.05.006 |
| [38] |
Ballantyne L, Fu X H, Lambkin C, Jeng M L, Faust L, Wijekoon M C D, Li D Q, Zhu T. Studies on South-east Asian fireflies: Abscondita, a new genus with details of life history, flashing patterns and behaviour of Abs. chinensis (L.) and Abs. terminalis (Olivier) (Coleoptera: Lampyridae: Luciolinae). Zootaxa, 2013, 3721: 1-48. DOI:10.11646/zootaxa.3721.1.1 |
| [39] |
付新华. 中国萤火虫生态图鉴. 北京: 商务印书馆, 2014: 92-93, 102-103.
|
| [40] |
Wang J L, Fu X H. The complete mitochondrial genome of the firefly, Abscondita chinensis (Coleoptera: Lampyridae). Mitochondrial DNA Part B, 2019, 4(1): 1599-1600. DOI:10.1080/23802359.2018.1551090 |
| [41] |
Sarfati R, Hayes J C, Peleg O. Self-organization in natural swarms of Photinus carolinus synchronous fireflies. Science Advances, 2021, 7(28): eabg9259. DOI:10.1126/sciadv.abg9259 |
| [42] |
Walker L A, Holwell G I. Static allometries do not reflect evolutionary allometry in exaggerated weaponry of male New Zealand sheetweb spiders (Cambridgea spp.). Journal of Evolutionary Biology, 2022, 35(11): 1524-1536. DOI:10.1111/jeb.14100 |
| [43] |
Okutaner A Y, Sarikaya A D. Sexual dimorphism of the pronotum in Dorcadion micans J. Thomson, 1867 (Coleoptera: Cerambycidae) using geometric morphometrics. Journal of Anatolian Environmental and Animal Sciences, 2021, 6(1): 88-91. DOI:10.35229/jaes.824342 |
| [44] |
Monroe M J, South S H, Alonzo S H. The evolution of fecundity is associated with female body size but not female‐biased sexual size dimorphism among frogs. Journal of Evolutionary Biology, 2015, 28(10): 1793-1803. DOI:10.1111/jeb.12695 |
| [45] |
Ogur E. Fecundity and body size-fecundity relations of Palaeolecanium bituberculatum (Targ. And Tozz.) (hemiptera: Coccidae) on apple trees. Pakistan Journal of Agricultural Sciences, 2018, 55(2): 381-387. DOI:10.21162/PAKJAS/18.5017 |
| [46] |
Akman O, Whitman D. Analysis of body size and fecundity in a grasshopper. Journal of Orthoptera Research, 2008, 17(2): 249-257. DOI:10.1665/1082-6467-17.2.249 |
| [47] |
Reiss M J. The allometry of growth and reproduction. Comparative Biochemistry and Physiology Part A: Physiology, 1990, 96(1): 242. |
| [48] |
Biewener A A, Patek S N. Animal locomotion. Second edition. Oxford: Oxford University Press, 2018.
|
| [49] |
Dudley R, Byrnes G, Yanoviak S P, Borrell B, Brown R M, McGuire J A. Gliding and the functional origins of flight: biomechanical novelty or necessity?. Annual Review of Ecology, Evolution, and Systematics, 2007, 38: 179-201. DOI:10.1146/annurev.ecolsys.37.091305.110014 |
| [50] |
Zeng Y, Chang S W, Williams J Y, Nguyen L Y N, Tang J A, Naing G, Kazi C, Dudley R. Canopy parkour: movement ecology of post-hatch dispersal in a gliding nymphal stick insect (Extatosoma tiaratum). Journal of Experimental Biology, 2020, 223(19): jeb226266. |
| [51] |
寿鹿, 杜卫国, 舒霖. 金线侧褶蛙和泽陆蛙的两性异形与生育力. 生态学报, 2005, 25(4): 664-668. DOI:10.3321/j.issn:1000-0933.2005.04.002 |