 2024, Vol. 44
2024, Vol. 44文章信息
- 杜雨霜, 吴刘萍, 陈杰, 区余端
- DU Yushuang, WU Liuping, CHEN Jie, OU Yuduan
- 崩岗生态修复不同人工林林下入侵植物和本土植物对群落稳定性的影响
- Effects of understory invasive plants and native plants on community stability in Benggang ecological restoration of different artificial forests
- 生态学报. 2024, 44(4): 1588-1600
- Acta Ecologica Sinica. 2024, 44(4): 1588-1600
- http://dx.doi.org/10.20103/j.stxb.202304010641
-
文章历史
- 收稿日期: 2023-04-01
- 网络出版日期: 2023-09-27

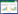




崩岗主要发育在华南和东南热带和亚热带湿润季风气候区丘陵坡地[1], 是在水力和重力作用下, 边坡土壤和石体崩塌, 植被毁坏的侵蚀现象[2]。近年来对于崩岗地区早期的修复多采用基于自然的解决方案来进行生态修复, 如通过不同树种来大量营造人工林, 并根据树种特征来对林下植物进行合理配置, 以此能更快达到较好的近自然修复效果[3—5]。崩岗地区常用的修复树种有包括桉树、杉树、松树、茶树和麻竹等, 而不同树种因为不同特征, 在成林过程中会形成不同的林下植物群落[6]。
林下植物是森林生态系统中重要的组成部分, 通常包括本土植物和外来植物, 其中外来植物可能会与本土植物产生激烈竞争从而成为入侵植物, 占据更多的生态位, 降低群落物种多样性, 对种间关系产生负面影响[7—8]。物种多样性反映了不同物种及物种组合对不同环境的适应力[9]。生态位是物种在群落中占据的位置及功能与地位, 反映物种利用资源的能力, 能作为筛选先锋植物的重要依据[7]。种间关系是物种在空间上的竞争、共存或随机关系, 体现物种之间的稳定性, 可应用于植物群落配置过程中[10]。植物的多样性、生态位和种间关系是现代生态学的热门话题[11—12], 许多学者对林下植物进行多角度研究以此来探究生态修复的效果[13—14], 但对于崩岗区林下植物的研究较少。
雷州半岛地处于属于粤西剥蚀台地, 暴雨气候和人类活动等造成严重的水土流失, 其中崩岗侵蚀占水土流失的20%, 位于全广东省之首, 严重破坏生态系统稳定, 恶化环境[15]。本研究以雷州半岛为研究区域, 选择崩岗区内4种不同类型的人工林进行调查, 旨在:(1)探讨林下入侵植物和本土植物的多样性和生态位特征来比较不同林分抵抗林下入侵植物的能力;(2)以生态位特征和种间联结为基础, 探讨合适的林下植物群落配置。
1 研究方法 1.1 研究地概况研究地为广东三岭山国家森林公园(109°30′—110°55′E, 20°12′—21°35′N), 地处雷州半岛, 总面积738.79 hm2。气候属热带北缘海洋性季风气候, 阳光充足, 历年平均气温22.8—23.5℃, 年均降雨量1396.3—1759.4 mm。公园为崩岗地貌, 侵蚀强度剧烈, 且高于自然沟蚀阶段侵蚀强度的上限[15]。土壤类型为砖红壤和砂砾土, 成土母岩为花岗岩、砂页岩, 土层中厚, 含砂量较大, 土壤较松散。植被类型以人工单一林为主, 主要有桉树林、松树林和相思林等[15—16]。
1.2 样地设置于2019年7月, 在广东三岭山国家森林公园的湿地松、尾叶桉、大叶相思和樟树等4种林分中, 分别设置10个20 m×20 m的样方, 调查每个样地的乔木种类、树高、胸径、叶面积指数、郁闭度, 在每个样方的四角和中心设置5个2 m×2 m小样方, 调查林下灌木和草本植物的物种、多度和盖度。2013年3月—5月进行人工种植4种林分的幼苗, 林下植物任其自然生长。根据中国外来入侵物种信息系统和《中国入侵植物名录》来确定外来入侵植物。仅研究已产生危害的外来入侵植物, 即入侵等级为前三级的入侵植物[17]。
1.3 数据分析与处理重要值表示植物物种在群落中的相对重要性, 选取各林分下重要值排名前十的不同物种进行分析研究[18]。重要值是反映某个物种在森林群落中的作用和地位的综合数量指标[19]
重要值=相对多度+相对盖度+相对频度
式中相对多度=某种植物的个体数/样方内全部个体总数×100%;相对盖度=某种植物的盖度/样方内全部植物的盖度和×100%;相对频度=某种植物的频度/所有物种的总频度×100%。
群落多样性[21]采用Patrick丰富度指数(Pa)、ShannonWiener指数(H′)、Simpson优势度指数(R)和Pielou均匀度指数(E)测定。
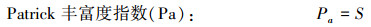
|
(1) |
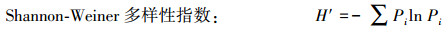
|
(2) |

|
(3) |

|
(4) |
式中, S为群落中的物种总数目, Pi为样方内第i个物种的个体数占所有物种个体数的比例, ni为样方内i个物种的个体数, N为样方内所有物种的个体总数。
生态位宽度体现物种利用资源多样性的能力, 生态位宽度越大其利用资源的能力越强, 在群落中更占优势地位[20]。运用Levins生态位宽度指数计算五种林分下重要值前十的物种生态位宽度, 公式如下:
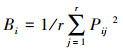
|
其中,
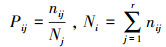
|
式中, Bi是种i 的生态位宽度; Pij是种i 在第j 个资源位的重要值与它在全部资源水平的重要值的比值; nij为种i 在资源位j上的重要值; r为资源位总位数(即样方数)。
生态位重叠度反应群落中两个物种利用相同资源的能力, 生态位重叠度高, 可能会导致一些物种之间的相互作用更加冲突(如竞争和排斥), 相反重叠度较低, 物种间的相互作用较小, 资源利用差异化大[21]。采用Pianka生态位重叠指数[22]来计算林下入侵植物和本土植物之间的重叠关系, 计算公式如下:
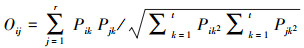
|
式中, Oij为种i和种j的生态位重叠度指数, Pik和Pjk分别种i和种j在资源梯度级k的重要值, r是样方数量。Oij取值范围为0—1, Oij> 0.6和Oij> 0.3分别表示物种i和物种j显著重叠和重叠有意义。
生态响应是植物在与环境资源综合作用下彼此竞争表现出发展性或衰退性结果, 反映物种对生态资源的反应速率[18]。生态响应的公式如下:
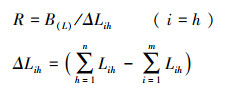
|
式中, 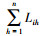
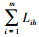
种间联结是指群落物种之间在空间上的关联性, 采用基于物种有无的Ochiai指数(OI)以及基于物种数量特征的Pearson指数(rp)[23]来计算林下入侵植物和本土植物的联结程度, 能反应两者之间的竞争、共存或随机的关系, 公式如下。
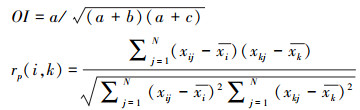
|
式中, a为2个物种同时出现的样方数, b、c分别为2个物种单独出现的样方数;OI指数的值域为[0, 1], 值越高, 2个物种同时出现的概率越大, 但不一定表示种间的正关联程度高[23];N为总样方数, xij和xkj分别是物种i和物种k在样方j中的重要值;xi和xk分别是j个样方中物种i和k的重要值的平均值;rp指数的值域为[-1, 1], 表示种间联结从负联结到正联结[24]。
应用SPSS 26软件对所获数据进行处理和相关统计分析。采用Origin 2021软件绘图。
2 结果与分析 2.1 林下植物群落组成和多样性4种林分中尾叶桉林的平均树高和平均胸径均最大, 其次是大叶相思林, 湿地松林和樟树林相近且最小, 说明尾叶桉的生长速度比其他3种树种都快。平均密度大小分别为大叶相思林>湿地松林>尾叶桉林>樟树林, 郁闭度大小分别为大叶相思林>樟树林>尾叶桉林>湿地松林, 说明大叶相思林最为密集且郁闭(表 1)。
| 林分 Stand |
平均树高 Average height/m |
平均胸径 Average DBH/cm |
林分密度 Stand density/ (plant/hm2) |
郁闭度 Canopy density |
| 湿地松Pinus elliottii | 4.92±0.78c | 7.47±1.45a | 7940±2201a | 0.54±0.11c |
| 尾叶桉Eucalyptus urophylla | 9.97±1.12a | 8.34±2.38a | 5080±2581b | 0.56±0.10b |
| 大叶相思Acacia auriculaeformis | 9.91±0.94b | 7.63±0.70b | 8790±3208a | 0.83±0.03a |
| 樟树Cinnamomum camphora | 4.17±0.78d | 7.49±1.78b | 3950±940b | 0.62±0.09c |
4种林分下共有植物104种, 隶51科92属, 本土和外来植物分别有93种和11种, 其中菊科(Asteraceae)种类最多, 有33种, 其次是禾本科(Poaceae)24种。湿地松林下本土植物为46种;尾叶桉林次之, 有33种;樟树林有24种;大叶相思林最少仅14种。而各林分下入侵植物种类分别为尾叶桉(4种) < 大叶相思林(6种) < 樟树林(7种)=湿地松林(7种)。
单因素方差分析结果表示, 不同林分下入侵植物的Patrick丰富度(F=11.204, P < 0.005)、Pielou均匀度(F=4.384, P < 0.005)、Simpson优势度(F=7.926, P < 0.005)和Shannon-Wiener多样性指数(F=9.205, P < 0.005)有显著性差异, 本土植物的Patrick丰富度(F=11.719, P < 0.005)、Pielou均匀度(F=6.804, P < 0.005)、Simpson优势度(F=3.251, P < 0.005)有显著性差异, 而Shannon-Wiener多样性指数(F=1.799, P=0.165)无显著差异。
由图 1可知, 尾叶桉林的林下入侵植物的Patrick丰富度、Pielou均匀度、Simpson优势度和Shannon-Wiener多样性指数均小于其他三种林分。在林下本土植物中, 湿地松林、尾叶桉林和樟树林的林下本土植物Simpson优势度和Shannon-Wiener多样性指数无明显差距。樟树林下本土植物Pielou均匀度最大, 其次是尾叶桉林, 而湿地松和大叶相思林指数较小。本土植物丰富度指数为湿地松林最大, 其次是尾叶桉林, 大叶相思林和樟树林近似且最小。

|
| 图 1 不同林分林下入侵植物和本土植物的多样性 Fig. 1 Diversity of understory invasive and native plants of different stands |
在重要值排名前十的林下植物中, 本土植物的数量分别为:尾叶桉林(7种)>湿地松林(6种)>大叶相思林(5种)>樟树林(4种), 入侵植物的数量顺序则相反(表 2)。其中尾叶桉林和湿地松林重要值排名前三的均为本土植物, 而大叶相思林和樟树林仅有一种本土植物。重要值较大的本土植物有芒萁、芒、筒轴茅等, 入侵植物有红毛草、假臭草等, 以上植物在群落中处于优势地位。生态位宽度较大的本土植物有芒萁、芒、桃金娘等, 入侵植物有假臭草、红毛草、阔叶丰花草等, 以上植物在群落中的资源利用能力更佳。其中尾叶桉林生态位宽度排名前三的均为本土植物, 湿地松林有两种本土植物, 而大叶相思林和樟树林仅有一种本土植物。
| 科 Family |
属 Genus |
物种 Species |
缩写 Abbreviation |
入侵等级 Invasion level |
| 禾本科Poaceae | 芒属 | 芒Miscanthus Sinensis | Misi | 本土植物 |
| 芒属 | 五节芒Miscanthus Floridulus | Mifl | 本土植物 | |
| 筒轴茅属 | 筒轴茅Rottboellia Cochinchinensis | Roco | 本土植物 | |
| 白茅属 | 白茅Imperata Cylindrica | Imcy | 本土植物 | |
| 稻属 | 冷水麻Oryza Sativa | Orsa | 1 | |
| 红毛草属 | 红毛草Rhynchelytrum Repens | Rhre | 2 | |
| 微甘菊属 | 微甘菊Mikania Micrantha | Mimi | 1 | |
| 菊科Asteraceae | 泽兰属 | 假臭草Praxelis Clematidea | Prcl | 1 |
| 泽兰属 | 飞机草Eupatorium Odoratum | Euod | 1 | |
| 鬼针草属 | 鬼针草Bidens Pilosa | Bipi | 1 | |
| 泽菊属 | 南美蟛蜞菊Sphagneticola Trilobata | Sptr | 1 | |
| 桃金娘科Myrtaceae | 桉属 | 桉树幼苗Eucalyptus Robusta | Euro | 本土植物 |
| 桃金娘属 | 桃金娘Rhodomyrtus Tomentosa | Rhto | 本土植物 | |
| 茜草科Rubiaceae | 九节属 | 蔓九节Psychotria Serpens | Psse | 本土植物 |
| 钮扣草属 | 阔叶丰花草Borreria Latifolia | Bola | 1 | |
| 芸香科Rutaceae | 花椒属 | 簕欓花椒Zanthoxylum Avicennae | Zaav | 本土植物 |
| 冬青科Aquifoliaceae | 冬青属 | 冬青Ilex Chinensis | Ilch | 本土植物 |
| 大戟科Euphorbiaceae | 野桐属 | 白楸Mallotus Paniculatus | Mapa | 本土植物 |
| 豆科Fabaceae | 含羞草属 | 含羞草Mimosa Pudica | Mipu | 2 |
| 野牡丹科Melastomataceae | 野牡丹属 | 野牡丹Melastoma Candidum | Meca | 本土植物 |
| 里白科Gleicheniaceae | 芒萁属 | 芒萁Dicranopteris Dichotoma | Didi | 本土植物 |
| 乌毛蕨科Blechnaceae | 乌毛蕨属 | 乌毛蕨Blechnum Orientale | Blor | 本土植物 |
| 莎草科Cyperaceae | 飘拂属 | 两歧飘拂草Fimbristylis Dichotoma | Fidi | 本土植物 |
| 入侵等级依据中国外来入侵物种信息系统划分 | ||||
尾叶桉林和湿地松林的林下本土植物种类多于入侵植物, 且重要值和生态位宽度排名前列的也是本土植物较多, 这表明这两种林分本土植物群落比入侵植物群落更占优势。而大叶相思林和樟树林则相反, 入侵植物群落更占优势。
| 林分 Stand |
物种 Species |
重要值 Important value |
生态位宽度 Niche width |
林分 Stand |
物种 Species |
重要值 Important value |
生态位宽度 Niche width |
|
| 湿地松 | Imcy | 0.456 | 0.543 | 大叶相思 | Misi | 0.886 | 0.758 | |
| Pinus elliottii | Misi | 0.420 | 0.641 | Acacia | Rhre | 0.856 | 0.711 | |
| Mifl | 0.377 | 0.486 | auriculaeformis | Bipi | 0.302 | 0.375 | ||
| Didi | 0.264 | 0.362 | Blor | 0.168 | 0.444 | |||
| Bipi | 0.263 | 0.440 | Mimi | 0.148 | 0.672 | |||
| Prcl | 0.188 | 0.546 | Zaav | 0.110 | 0.446 | |||
| Meca | 0.089 | 0.459 | Rhto | 0.097 | 0.280 | |||
| Euod | 0.081 | 0.211 | Ilch | 0.085 | 0.440 | |||
| Blor | 0.073 | 0.247 | Mipu | 0.084 | 0.402 | |||
| Mipu | 0.061 | 0.298 | Prcl | 0.069 | 0.173 | |||
| 尾叶桉 | Didi | 0.927 | 0.623 | 樟树 | Bola | 0.522 | 0.673 | |
| Eucalyptus | Roco | 0.646 | 0.449 | Cinnamomum | Prcl | 0.474 | 0.838 | |
| urophylla | Misi | 0.314 | 0.100 | camphora | Rhto | 0.306 | 0.608 | |
| Prcl | 0.256 | 0.366 | Roco | 0.222 | 0.507 | |||
| Euro | 0.176 | 0.496 | Bipi | 0.211 | 0.579 | |||
| Psse | 0.144 | 0.300 | Mimi | 0.147 | 0.282 | |||
| Bipi | 0.082 | 0.226 | Mapa | 0.111 | 0.310 | |||
| Orsa | 0.075 | 0.119 | Imcy | 0.077 | 0.100 | |||
| Euod | 0.068 | 0.367 | Imcy | 0.077 | 0.100 | |||
| Fidi | 0.050 | 0.100 | Sptr | 0.031 | 0.100 | |||
| 只列出重要值排前十位的植物;“Δ”为入侵植物 | ||||||||
由图 2可知4种林分下林下入侵植物和本土植物组成种对的重叠度指数有较大的差异性, 重叠度指数范围0—0.3的种对数占总种对数的比例分别为尾叶桉林(71.43%)>湿地松林(58.33%)>樟树林(50.00%)>大叶相思林(32.00%)。生态位重叠度在0.3—0.6和>0.6的范围中, 尾叶桉林的种对数均最小, 所占比例分别为23.81%和4.76%, 大叶相思林的种对数均最大, 所占比例分别为44.00%和24.00%。说明尾叶桉林的林下入侵植物与本土植物重叠程度最小。

|
| 图 2 不同林分下林下入侵植物和本土植物的生态位重叠度指数范围 Fig. 2 Ecological niche overlap index range of understory invasive plants and native species in different stands |
4种林分下发展型物种数湿地松林最多, 有6种;其次是尾叶桉林和樟树林, 均有5种;大叶相思林最少, 只有4种(表 4)。其中林下本土发展型植物数量分别为:湿地松林4种>尾叶桉林3种>樟树林2种>大叶相思林1种(表 4)。本土发展型植物越多, 说明本土植物在未来发展中相比于入侵植物更占优势, 因此湿地松林的未来发展趋势最佳, 其次是尾叶桉林, 大叶相思林最差。
| 林分 Stands |
物种 Species |
ΔLih | R | 物种 Species |
ΔLih | R |
| 湿地松 | Imcy | 0.101 | 5.371 | Prcl | 0.161 | 3.389 |
| Pinus elliottii | Mifl | 0.041 | 11.940 | Meca | 0.035 | 13.147 |
| Misi | 0.175 | 3.662 | Euod | -0.209 | -1.013 | |
| Didi | -0.059 | -6.164 | Blor | -0.157 | -1.576 | |
| Bipi | 0.033 | 13.423 | Mipu | -0.122 | -2.454 | |
| 尾叶桉 | Didi | 0.262 | 2.381 | Bipi | -0.092 | -2.462 |
| Eucalyptus urophylla | Roco | 0.175 | 2.564 | Euod | 0.111 | 3.321 |
| Prcl | 0.022 | 20.867 | Orsa | -0.198 | -0.603 | |
| Euro | 0.137 | 3.631 | Fidi | -0.111 | -0.903 | |
| Psse | -0.086 | -3.488 | Misi | -0.199 | -0.503 | |
| 大叶相思 | Rhre | 0.209 | 3.395 | Zaav | -0.045 | -8.831 |
| Acacia auriculaeformis | Misi | 0.286 | 2.650 | Rhto | -0.191 | -1.465 |
| Bipi | -0.091 | -4.106 | Ilch | -0.051 | -8.658 | |
| Blor | -0.035 | -12.701 | Mipu | -0.064 | -6.305 | |
| Mimi | 0.231 | 2.912 | Prcl | -0.249 | -0.696 | |
| 樟树 | Munu | -0.246 | -0.755 | Bipi | 0.156 | 3.712 |
| Cinnamomum camphora | Bola | 0.162 | 4.145 | Mimi | -0.112 | -2.523 |
| Prcl | 0.408 | 2.056 | Mapa | -0.138 | -2.239 | |
| Rhto | 0.095 | 6.402 | Imcy | -0.192 | -0.522 | |
| Roco | 0.036 | 14.049 | Sptr | -0.170 | -0.589 | |
| 只列出重要值排前十位的植物; R:生态响应速率;ΔLih:指种群i占用其他种群的资源总量与种群h被其他种群占用的资源总量之间的差 | ||||||
各林分中发展型植物的发展程度以湿地松-鬼针草、尾叶桉-假臭草、大叶相思-红毛草以及樟树-筒轴茅的最大, 可预见这些物种将在各自林分中逐渐取代其他衰退型植物(表 4);各林分中衰退型植物的衰退程度不大, 只有大叶相思-乌毛蕨、大叶相思-簕欓花椒有明显的衰退趋势, 说明除大叶相思林外, 其他3种林分发展型的平均速率大于衰退型, 林下植物群落处于正向发展(表 4)。
2.5 种间关联4种林分下入侵植物和本土植物的种间联结呈中下水平, OI种间联结指数都不大于0.5。尾叶桉林中种对联结指数大于0.4的仅假臭草-筒轴茅, 其余三种林分种对联结指数大于0.4的种对分别有湿地松林4对、大叶相思林8对、樟树林9对(图 3, 4, 5, 6)。这表明该4种林分下种间联结程度不大, 入侵植物和本土植物的种对同时出现的概率不高, 其中以尾叶桉林种对出现的概率最小。

|
| 图 3 湿地松林的林下植物之间的Ochiai指数(OI) Fig. 3 The Ochiai index (OI) of understory plants in Pinus elliottii stand Blor:乌毛蕨;Meca:野牡丹;Didi:芒萁;Mifl:五节芒;Misi:芒;Imcy:白茅;Bipi:鬼针草;Euod:飞机草;Mipu:含羞草 |

|
| 图 4 尾叶桉林的林下植物之间的Ochiai指数(OI) Fig. 4 The Ochiai index (OI) of understory plants in Eucalyptus urophylla stand Fidi:两歧飘拂草;Orsa:冷水麻;Psse:蔓九节;Euro:桉树幼苗;Roco:筒轴茅;Prcl:假臭草 |

|
| 图 5 大叶相思林的林下植物之间的Ochiai指数(OI) Fig. 5 The Ochiai index (OI) of understory plants in Acacia auriculaeformis stand Ilch:冬青;Rhto:桃金娘;Zaav:簕欓花椒;Rhre:红毛草;Mimi:微甘菊 |

|
| 图 6 樟树林的林下植物之间的Ochiai指数(OI) Fig. 6 The Ochiai index (OI) of understory plants in Cinnamomum camphora stand Sptr:南美蟛蜞菊;Bola:阔叶丰花草;Mapa:白楸 |
各林分下优势植物分别有45组种对, 其中显著正联结的种对数为大叶相思林5对>尾叶桉林3对>樟树林2对>湿地松林1对, 4种林分下呈负联结的种对数量均大于正联结(图 7—10)。上述说明4种林分林下群落均不稳定。

|
| 图 7 湿地松林的林下植物Pearson指数 Fig. 7 Pearson index of understory plants in Pinus elliottii stand *表示显著联结, **、***表示极显著联结; *P≤0.05;**P≤0.01; ***P≤0.001 |

|
| 图 8 尾叶桉林的林下植物Pearson指数 Fig. 8 Pearson index of understory plants in Eucalyptus urophylla stand *P≤0.05;**P≤0.01; ***P≤0.001 |

|
| 图 9 大叶相思林的林下植物Pearson指数 Fig. 9 Pearson index of understory plants in Acacia auriculaeformis stand *P≤0.05;**P≤0.01; ***P≤0.001 |

|
| 图 10 樟树林的林下植物Pearson指数 Fig. 10 Pearson index of understory plants in Cinnamomum camphora stand *P≤0.05;**P≤0.01; ***P≤0.001 |
本研究中尾叶桉林、樟树林和湿地松林的林下本土植物多样性指数相近, 大叶相思林则最小, 而林下入侵植物多样性指数最小是尾叶桉林, 樟树林则最大。这说明尾叶桉林在林下本土植物丰富茂盛的情况下, 入侵植物较少。林下植物的生长与光照透过率密不可分, 其中林分的郁闭度影响林下透光率, 郁闭度较小树种其林下光照较多, 林下植物的物种丰富度更高[25—27]。相比于高郁闭度的树种, 中等郁闭度的树种对林下植物多样性起着正向增加的作用, 将郁闭度控制在0.45—0.6能保持较大的林下植物多样性[28—30], 而本研究中桉树林的郁闭度为0.56±0.10, 处在该阈值中, 冠层疏密有致, 适合林下植物生长, 而大叶相思林的郁闭度为0.83±0.03, 冠层最为郁闭, 其林下植物种类最少。杨民胜、余雪等[31—33]学者认为桉树会使林下物种丧失, 多样性降低, 而周笛轩、张卫强、段文军等[34—36]学者认为桉树不会对林下植物多样性产生负面影响, 本研究与后者的研究结果一致, 桉树林的林下植物多样性并未显著低于其他人工林。本土植物多样性越大, 其群落内入侵植物能占据的生态位较小, 使入侵植物获得较少的资源, 在一定程度上有利于减少植物入侵[37—38]。尾叶桉林下的本土物种数量位居第二, 对降低外来植物成功入侵有一定的正向作用。而湿地松林的林下本土植物虽丰富度最高, 但入侵植物也最多, 这是因为其Pielou均匀度指数最小, 而较大的物种均匀性会提高群落生产力, 减少成功入侵的机会[39]。樟树林的林下入侵植物和本土植物多样性指数近似, 说明其入侵和本土植物在群落中竞争, 两者互不相让。大叶相思林下本土植物和入侵植物多样性指数均较小, 说明其林下植物本土植物和入侵植物种类均最小。
3.2 崩岗修复中林下植物的生态位特征尾叶桉林的林下入侵植物生态位宽度最小, 且入侵植物与本土植物的生态位重叠度也最小, 说明林下本土植物据有更高的资源利用能力, 入侵植物在群落中所占优势较小, 两者之间互相竞争不激烈。尾叶桉存在化感作用, 能对入侵植物产生一定的抑制作用[40]。同时尾叶桉林下有大量的芒萁, 其重要值和生态位宽度均最大, 芒萁据有生态筛效应, 强竞争性, 能使其他物种难以入侵[41]。湿地松林的林下入侵植物虽最多, 但生态位宽度不大, 林下本土植物更占优势, 而大叶相思和樟树林的林下入侵植物鬼针草、假臭草和红毛草的生态位宽度较大, 分布范围广, 对资源的利用高, 且具有化感作用, 对其他本土植物有着抑制作用[42—44]。
4种林分下林下植物生态位分化较为明显, 本土植物和入侵植物重叠度不算大, 这与郭朝丹等[18]的研究结果类似。崩岗地区不稳定的环境条件和人工林系统中人类的频繁干扰使得林下本土植物会聚集在局部适宜的环境下, 具有空缺的生态位可以接受其他种群, 易被外来物种入侵[45—46]。在入侵过程中, 本土植物为了能更好的生存, 在资源需求上产生了分化, 使得与入侵植物的生态位重叠度降低, 进而在一定程度上减少了种群间对资源的竞争[47]。生态位宽度较大的物种多数为发展型, 如入侵植物鬼针草和假臭草在多个林分中均为发展型, 且部分响应速率较大, 这意味着它们不仅现在是群落中的优势物种, 而且在未来群落演替中也处于优势地位[48]。对于为衰弱型且窄生态位的本土植物其在未来可能会被入侵植物进一步侵占生存空间和资源, 处于劣势地位[49]。因此从生态环境恢复的角度来看, 可通过人为干扰来限制群落中优势入侵植物的部分空间和物种数量, 适当增宽劣势本土生态位宽度, 使之成为发展型物种[50]。
3.3 崩岗修复中林下植物的种间联结群落中正联结种对间能互利互补, 提升群落的稳定性;负联结种对间则多出现竞争排斥, 降低群落的稳定性[51]。本研究中4种林分优势林下植物多为负联结, 说明物种间存在竞争, 群落还处于不稳定状态, 其中尾叶桉林的林下本土植物和入侵植物联结程度最低, 说明其竞争程度最小。种间联结在一定程度上能体现群落中物种的相互关系, 因此可以将种间联结运用于植物配置过程中, 王敉敉[52]、陈家龙等[53]的研究表明, 将正联结的植物进行配置组合, 物种间多生长健康且长势良好, 并且能充分利用环境资源。随着群落演替的进行, 种间联结的正联结比例和显著程度都会增加, 种间竞争减弱, 群落处于稳定状态[54]。胡文强等[55]对山顶矮林群落和刘益鹏等[56]研究表明, 群落内种间关系均呈显著正联结, 群落处于稳定的演替后期, 可见本研究中4种林分均处于演替前期, 这点也与胡贝娟等[57]研究结果一致。
3.4 崩岗修复中林下植物对群落稳定性影响4种林分的林下入侵植物入侵等级均为1级或2级, 分别为恶性入侵类和严重入侵类, 多位学者研究表明入侵植物会使群落处于不稳定状态, 与本土植物产生竞争关系[58—61]。孙成[62]、郑晓阳等[50]研究表明林下植物的发展性物种越多, 则植物群落越稳定。种间联结能体现出群落物种之间的相互影响, 正联结物种能避免不必要的竞争, 使得群落趋向稳定[63]。而群落的稳定性和种间联结等都会受入侵植物的影响。因此对于林下植物群落的构建, 先选择合适的先锋植物来迅速占据群落中较大的生态位, 之后再选取发展型或与先锋植物呈正联结的本土植物进行配置, 并抵抗高风险的外来入侵植物, 以此形成相对稳定的植物群落。如群落配置模式可优化为:湿地松-芒-白茅-乌毛蕨(图 11);尾叶桉-芒萁-筒轴茅-芒(图 12);大叶相思-芒-乌毛蕨-桃金娘(图 13);樟树-桃金娘-筒轴茅-白茅(图 14)。同时要通过必要的人工干预措施来控制鬼针草、假臭草和红毛草等入侵植物。植物配置情况如下:

|
| 图 11 湿地松林的林下植物配置 Fig. 11 Understory plant configuration under Pinus elliottii stand |

|
| 图 12 尾叶桉林的林下植物配置 Fig. 12 Understory plant configuration under Eucalyptus urophylla stand |

|
| 图 13 大叶相思林的林下植物配置 Fig. 13 Understory plant configuration under Acacia auriculaeformis stand |

|
| 图 14 樟树林的林下植物配置 Fig. 14 Understory plant configuration under Cinnamomum camphora stand |
在崩岗修复早期, 可用尾叶桉作为先锋树种, 快速成林, 其林下本土植物丰富同时入侵植物较少, 在后期则可加入其他乔木物种如湿地松、大叶相思和樟树等增加林分的多样性, 形成更近自然的修复模式, 并继续进行追踪研究。本研究仅对4种林分的林下植物进行分析, 下一步将从林分冠层、土壤理化性质等角度来进行更详细的探究, 为崩岗区的生态修复提供合理的依据。
| [1] |
刘希林. 全球视野下崩岗侵蚀地貌及其研究进展. 地理科学进展, 2018, 37(3): 342-351. |
| [2] |
文慧, 冯舒悦, 王军光, 倪世民, 蔡崇法. 南方红壤区不同崩岗侵蚀治理模式及评价指标体系研究. 中国水土保持, 2019(9): 40-44, 69. DOI:10.3969/j.issn.1000-0941.2019.09.016 |
| [3] |
倪晨. 花岗岩崩岗集水坡面土壤水分状况研究[D]. 武汉: 华中农业大学, 2016.
|
| [4] |
谢金波. 粤东崩岗侵蚀区生态修复应用实践. 科技创业家, 2012(14): 216. |
| [5] |
孙冬婧, 温远光, 罗应华, 李晓琼, 张万幸, 明安刚. 近自然化改造对杉木人工林物种多样性的影响. 林业科学研究, 2015, 28(2): 202-208. |
| [6] |
刘宾, 周静, 袁秋平. 浅析园林绿化乡土树种优势及利用策略. 环境与发展, 2020, 32(7): 175-176. |
| [7] |
王世雄, 张广奇, 何跃军. 外来植物入侵对喀斯特乡土物种多样性影响研究进展. 山地农业生物学报, 2023, 42(1): 40-45. |
| [8] |
洪思思, 缪崇崇, 方本基, 胡仁勇, 丁炳扬. 浙江省阔叶丰花草入侵群落物种多样性、生态位及种间联结研究. 武汉植物学研究, 2008, 26(5): 501-508. |
| [9] |
张光明, 谢寿昌. 生态位概念演变与展望. 生态学杂志, 1997, 16(6): 46-51. |
| [10] |
胡相明, 程积民, 万惠娥. 黄土丘陵区人工林下草本层植物的结构特征. 水土保持通报, 2006, 26(3): 41-45. |
| [11] |
Schellenberger Costa D, Gerschlauer F, Kiese R, Fischer M, Kleyer M, Hemp A. Plant niche breadths along environmental gradients and their relationship to plant functional traits. Diversity and Distributions, 2018, 24(12): 1869-1882. DOI:10.1111/ddi.12815 |
| [12] |
Turnbull L A, Isbell F, Purves D W, Loreau M, Hector A. Understanding the value of plant diversity for ecosystem functioning through niche theory. Proceedings Biological Sciences, 2016, 283(1844): 20160536. |
| [13] |
刘尧尧, 辜彬, 王丽. 北川震后植被恢复工程植物群落物种多样性及优势种生态位. 生态学杂志, 2019, 38(2): 309-320. |
| [14] |
珊丹, 邢恩德, 荣浩, 刘艳萍, 梁占岐. 草原矿区排土场不同植被配置类型生态恢复. 生态学杂志, 2019, 38(2): 336-342. |
| [15] |
匡耀求, 孙大中. 雷州半岛第四纪台地区的崩岗侵蚀地貌──一种后果严重的水土流失现象. 热带地理, 1998, 18(1): 42-49. DOI:10.3969/j.issn.1001-5221.1998.01.008 |
| [16] |
王喜平. 广东三岭山国家森林公园景观林建设及其生态价值评估. 防护林科技, 2007(5): 55-56, 104. DOI:10.3969/j.issn.1005-5215.2007.05.023 |
| [17] |
闫小玲, 刘全儒, 寿海洋, 曾宪锋, 张勇, 陈丽, 刘演, 马海英, 齐淑艳, 马金双. 中国外来入侵植物的等级划分与地理分布格局分析. 生物多样性, 2014, 22(5): 667-676. |
| [18] |
郭朝丹, 赵彩云, 李飞飞, 李俊生. 天然林和人工林外来入侵和本地植物对比研究: 以弄岗国家级自然保护区为例. 生物多样性, 2022, 30(4): 45-57. |
| [19] |
王育松, 上官铁梁. 关于重要值计算方法的若干问题. 山西大学学报: 自然科学版, 2010, 33(2): 312-316. |
| [20] |
魏志琴, 李旭光, 郝云庆. 珍稀濒危植物群落主要种群生态位特征研究. 西南农业大学学报: 自然科学版, 2004, 26(1): 1-4. |
| [21] |
Tsafack N, Wang X P, Xie Y Z, Fattorini S. Niche overlap and species co-occurrence patterns in carabid communities of the northern Chinese steppes. ZooKeys, 2021, 1044: 929-949. DOI:10.3897/zookeys.1044.62478 |
| [22] |
吴会峰, 宋丽娟, 杜峰, 郝文芳. 黄土丘陵区草地主要种群生态位及其环境解释. 应用生态学报, 2017, 28(11): 3494-3504. |
| [23] |
刘润红, 陈乐, 涂洪润, 梁士楚, 姜勇, 李月娟, 黄冬柳, 农娟丽. 桂林岩溶石山青冈群落灌木层主要物种生态位与种间联结. 生态学报, 2020, 40(6): 2057-2071. |
| [24] |
程占红, 牛莉芹. 五台山南台山地草甸种群对旅游干扰响应的识别. 应用与环境生物学报, 2008, 14(3): 324-327. DOI:10.3321/j.issn:1006-687X.2008.03.007 |
| [25] |
Mestre L, Toro-Manríquez M, Soler R, Huertas-Herrera A, Martínez-Pastur G, Lencinas M V. The influence of canopy-layer composition on understory plant diversity in southern temperate forests. Forest Ecosystems, 2017, 4: 6. DOI:10.1186/s40663-017-0093-z |
| [26] |
Zhang Y X, Liu T R, Guo J P, Tan Z J, Dong W, Wang H H. Changes in the understory diversity of secondary Pinus tabulaeformis forests are the result of stand density and soil properties. Global Ecology and Conservation, 2021, 28: e01628. DOI:10.1016/j.gecco.2021.e01628 |
| [27] |
姜祖扬, 覃业辉, 刘季, 文志, 郑华, 欧阳志云. 海拔和林分密度对热带雨林林下植物多样性的影响. 生态学杂志, 2023, 42(5): 1049-1055. |
| [28] |
Ren S Y, Ali A, Liu H M, Yuan Z Q, Yang Q S, Shen G C, Zhou S S, Wang X H. Response of community diversity and productivity to canopy gap disturbance in subtropical forests. Forest Ecology and Management, 2021, 502: 119740. DOI:10.1016/j.foreco.2021.119740 |
| [29] |
王依瑞, 王彦辉, 段文标, 李平平, 于澎涛, 甄理, 李志鑫, 尚会军. 黄土高原刺槐人工林郁闭度对林下植物多样性特征的影响. 应用生态学报, 2023, 34(2): 305-314. |
| [30] |
Linstädter A, Bora Z, Tolera A, Angassa A. Are trees of intermediate density more facilitative? Canopy effects of four East African legume trees. Applied Vegetation Science, 2016, 19(2): 291-303. DOI:10.1111/avsc.12218 |
| [31] |
杨民胜, 吴志华, 陈少雄. 桉树的生态效益及其生态林经营. 桉树科技, 2006, 23(1): 32-39. DOI:10.3969/j.issn.1674-3172.2006.01.007 |
| [32] |
余雪标, 钟罗生, 杨为东, 龙腾. 桉树人工林林下植被结构的研究. 热带作物学报, 1999, 20(1): 66-72. |
| [33] |
杨民胜, 吴志华, 张维耀. 我国环北部湾地区发展桉树的思考. 世界林业研究, 2016, 29(6): 64-69. |
| [34] |
周笛轩, 林永标, 汪雁佳, 刘占锋, 周丽霞. 南亚热带不同人工林生态系统服务功能评估. 生态环境学报, 2021, 30(5): 907-919. |
| [35] |
张卫强, 张卫华, 潘文, 陈伟光, 唐洪辉, 盘李军. 桉树林和针阔混交林对植物多样性的影响比较. 水土保持研究, 2014, 21(6): 122-128. |
| [36] |
段文军, 李达, 李冲. 不同林龄尾巨桉人工林林下植物和种子库多样性比较及影响因素分析. 中南林业科技大学学报, 2022, 42(12): 26-33, 81. |
| [37] |
Liao H X, Luo W B, Peng S L, Callaway R M. Plant diversity, soil biota and resistance to exotic invasion. Diversity and Distributions, 2015, 21(7): 826-835. DOI:10.1111/ddi.12319 |
| [38] |
刘艳杰, 黄伟, 杨强, 郑玉龙, 黎绍鹏, 吴昊, 鞠瑞亭, 孙燕, 丁建清. 近十年植物入侵生态学重要研究进展. 生物多样性, 2022, 30(10): 276-292. |
| [39] |
Brett Mattingly W, Hewlate R, Reynolds H L. Species evenness and invasion resistance of experimental grassland communities. Oikos, 2007, 116(7): 1164-1170. DOI:10.1111/j.0030-1299.2007.15406.x |
| [40] |
Chen P D, Hou Y P, Zhuge Y H, Wei W, Huang Q Q. The effects of soils from different forest types on the growth of the invasive plant Phytolacca americana. Forests, 2019, 10(6): 492. DOI:10.3390/f10060492 |
| [41] |
刘美洁, 孙中宇, 耿守保, 温美丽, 戴佳玲, 杨龙. 华南桉树人工林芒萁层片的生态筛效应. 热带地理, 2021, 41(6): 1338-1346. |
| [42] |
丁丹, 陈超. 红毛草(Rhynchelytrum repens)入侵特性、地理分布和风险评估. 杂草学报, 2016, 34(2): 29-33. DOI:10.3969/j.issn.1003-935X.2016.02.007 |
| [43] |
王桔红, 史生晶, 陈文, 李云, 崔现亮. 鬼针草与含羞草化感作用及其入侵性的研究. 草业学报, 2020, 29(4): 81-91. |
| [44] |
邓文爽, 张玉宁, 赖洁莹, 袁俏玲. 假臭草化感作用的研究进展. 广东化工, 2018, 45(2): 106-107. |
| [45] |
江焕, 张辉, 龙文兴, 方燕山, 符明期, 朱孔新. 金钟藤入侵群落的种间联结及生态位特征. 生物多样性, 2019, 27(4): 388-399. |
| [46] |
Antunes C, Pereira A J, Fernandes P, Ramos M, Ascensão L, Correia O, Máguas C. Understanding plant drought resistance in a Mediterranean coastal sand dune ecosystem: differences between native and exotic invasive species. Journal of Plant Ecology, 2018, 11(1): 26-38. DOI:10.1093/jpe/rtx014 |
| [47] |
陈彤, 刘文莉, 张崇邦, 王江. 加拿大一枝黄花入侵对本土植物群落动态的影响及其机制. 植物生态学报, 2012, 36(3): 253-261. |
| [48] |
陈铭, 黄林娟, 黄贵, 刘昕宇, 薛跃规. 广西大石围天坑群草本植物多样性及其生态位变化规律. 生态学报, 2023, 43(7): 2831-2844. |
| [49] |
王伟伟, 杨海龙, 贺康宁, 胡兴波, 王辉, 董梅, 谢增武. 祁连山青海云杉人工林草本层生态位及生态响应研究. 草地学报, 2012, 20(4): 626-630. |
| [50] |
郑晓阳, 赵冲, 刘青青, 冯玉超, 朱晨曦, 王正宁, 刘博. 成熟杉木人工林林下草本层生态位特征. 生态学杂志, 2018, 37(2): 332-338. |
| [51] |
张明霞, 王得祥, 康冰, 张岗岗, 刘璞, 杜焰玲, 于飞. 秦岭华山松天然次生林优势种群的种间联结性. 林业科学, 2015, 51(1): 12-21. |
| [52] |
王敉敉, 吴泽民. 苏州古典园林主要群植植物生态关系. 中国城市林业, 2007, 5(4): 27-29. |
| [53] |
陈家龙. 城市公园木本植物群落类型及树种相关性研究[D]. 合肥: 安徽农业大学, 2009.
|
| [54] |
O'Connor I, Aarssen L W. Species association patterns in abandoned sand Quarries. Vegetatio, 1988, 73(2): 101-109. |
| [55] |
胡文强, 黄世能, 李家湘, 周光益, 王旭. 南岭石坑崆山顶矮林乔木优势种群的种间关联性. 生态学杂志, 2013, 32(10): 2665-2671. |
| [56] |
刘益鹏, 叶兴状, 叶利奇, 陈绪辉, 郑世群, 陈世品, 张国防, 刘宝. 观光木群落优势树种生态位和种间联结. 应用生态学报, 2022, 33(10): 2670-2678. |
| [57] |
胡贝娟, 张钦弟, 张玲, 毕润成. 山西太岳山连翘群落优势种种间关系. 生态学杂志, 2013, 32(4): 845-851. |
| [58] |
钱宗亮, 向华, 王悦, 庞涛. 白花鬼针草入侵对本土植物群落主要物种种间联结和稳定性的影响. 四川林业科技, 2023, 44(2): 31-37. |
| [59] |
黄小荣, 庞世龙, 申文辉, 侯远瑞, 何峰. 广西喀斯特地区假臭草入侵群落的草本植物多样性及其影响因素. 应用生态学报, 2016, 27(3): 815-821. |
| [60] |
郭佳琦, 陈俊辰, 黄旬, 黄佳乐, 赵丽娅, 李兆华. 喜旱莲子草入侵群落主要物种生态位和种间联结研究. 生态环境学报, 2021, 30(8): 1607-1616. |
| [61] |
郭连金. 苏门白酒草对乡土植物群落种间联结性及稳定性的影响. 亚热带植物科学, 2011, 40(2): 18-23. |
| [62] |
孙成, 秦富仓, 包雪源, 李龙, 杨振奇, 董晓宇, 李艳. 砒砂岩区人工油松林下草本植物的生态位特征. 水土保持通报, 2021, 41(3): 81-86. |
| [63] |
涂洪润, 农娟丽, 朱军, 赵佳佳, 杨婉琳, 朱琪琪, 谢彦军, 刘润红. 桂林岩溶石山密花树群落主要物种的种间关联及群落稳定性. 生态学报, 2022, 42(09): 3688-3705. |