 2024, Vol. 44
2024, Vol. 44文章信息
- 林小乔, 许玲玲, 刘沛荣, 同小娟, 张静茹, 张劲松, 李俊, 孟平
- LIN Xiaoqiao, XU Lingling, LIU Peirong, TONG Xiaojuan, ZHANG Jingru, ZHANG Jinsong, LI Jun, MENG Ping
- 亚热带杉木径向生长对气候因子的响应
- Response of stem radial growth of Cunninghamia lanceolata to climatic variables in the subtropic region
- 生态学报. 2024, 44(4): 1633-1646
- Acta Ecologica Sinica. 2024, 44(4): 1633-1646
- http://dx.doi.org/10.20103/j.stxb.202212313715
-
文章历史
- 收稿日期: 2022-12-31
- 网络出版日期: 2023-12-01

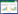




2. 中国林业科学研究院林业研究所, 北京 100091;
3. 中国科学院地理科学与资源研究所陆地水循环及地表过程重点实验室, 北京 100101;
4. 河南黄河小浪底关键带国家野外科学观测研究站, 济源 454650
2. Key Laboratory of Tree Breeding and Cultivation of State Forestry Administration, Research Institute of Forestry, Chinese Academy of Forestry, Beijing 100091, China;
3. Key Laboratory of Water Cycle and Related Land Surface Processes, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China;
4. Henan Xiaolangdi Earth Critical Zone National Research Station on the Middle Yellow River, Jiyuan 454650, China
受自然和人类活动的影响, 全球气候变暖和极端干旱事件发生的次数和频率增加, 使森林生长受到了限制, 甚至引发森林生态系统退化。在我国亚热带地区, 季节性干旱事件频发, 加之林木对干旱事件的响应具有遗留效应[1], 使环境制约因素更加复杂, 对林木造成了严重的损害和扰动。然而, 有关限制该地区林木生长的因素的关注远低于干旱和半干旱地区[2]。因此, 探讨亚热带地区树木径向生长对干旱事件的响应机制成为了必要。抵抗力、恢复力已被广泛用于评估树木应对干旱事件的能力, 有研究表明随着气候环境的变化, 林木的抵抗力和恢复力表现出明显的差异[3]。树木径向生长与增温引起的干旱分异现象极其显著[4]。研究发现, 夏季温度对林木径向生长表现出抑制作用, 在生理上表现为高温导致树木气孔关闭, 减弱林木的光合作用[5], 而林木生长速率降低是对干旱胁迫的一种适应策略[6]。当林木径向生长受到的干旱胁迫较严重时, 降水成为最主要的限制因子, 降水量较大的地区林木对干旱事件更加敏感, 主要表现在林木生长对干旱事件的抵抗力上[7], 干旱地区的树木比湿润地区的树木具有更差的抵抗力[3, 8—9]。还有学者发现, 随着海拔的变化林木对干旱事件的响应也不同[10], 低海拔地区树木生长比高海拔地区更容易受到夏季干旱的限制[11], 且随着海拔的上升林木对干旱事件的敏感性降低[12]。不同纬度的地区气候因子对林木生长的影响也存在一定的差异。纬度较高的地区降水较少, 林木受到干旱胁迫较严重。而在纬度较低或靠近海洋的地区水分较充足, 林木生长对气温的响应较明显, 对降水因子则不敏感[13]。
杉木(Cunninghamia lanceolata)广泛分布于我国亚热带地区[14], 亚热带地区降水和温度对于杉木生长的影响不同[15], 不同学者对其进行了系统研究[2, 16—17]。朱安明等[18]研究表明, 干旱会抑制杉木的生长。孟盛旺等[19]发现夏季的干旱会缩短杉木的生长季时间。在生长季末期, 温度升高会造成杉木净光合产物减少, 加剧干旱胁迫[20]。目前, 关于杉木年轮生长对气温升高响应的研究较多, 且这部分研究局限在年表和气候生长模型的建立上, 干旱事件对杉木人工林径向生长影响的研究相对较少, 特别是缺乏对干旱事件发生前后林木生长变化的研究。因此, 本文研究亚热带不同地区杉木人工林径向生长对气候变化的响应, 明确杉木对干旱事件的抵抗力、恢复力和弹性力[6], 揭示杉木在干旱事件下的响应策略, 为人工林应对气候变化提供参考, 并对森林生态系统生产力的估算提供数据支持。
1 材料与方法 1.1 研究区概况本研究选取的6个站点分别为亚热带地区的荆关、马鬃岭、分宜、将乐、东风、高峰, 其位置和分布如图 1所示。荆关林场(JG)位于河南省南阳市淅川县西南边陲(33°16′N, 111°4′E, 海拔710 m), 该林场属北亚热带和暖温带交汇地带, 年平均气温15.8℃, 高于同纬度其他地区。最低气温为-13.5℃, 最高气温40.3℃。无霜期约为228d。年降水量804.8 mm。土壤以黄棕壤为主, 呈中性偏弱碱性。

|
| 图 1 杉木人工林站点位置 Fig. 1 Locations of Cunninghamia lanceolata in this study 荆关: JG, 马鬃岭: MZL, 分宜: FY, 将乐: JL, 东风: DF, 高峰: GF |
马鬃岭自然保护区(MZL)位于安徽省六安市金寨县西南部(31°17′ N, 115°41′ E, 海拔1080 m)。该地区属亚热带湿润季风气候, 年均降水量1315 mm, 年平均气温14—15℃, 最冷月(1月)均温2.7℃, 极端低温-16.5℃, 最热月(7月)均温27.9℃, 最高温是40.1℃。土壤以山地棕壤为主, 土壤pH呈弱酸性, 有机质含量30 g/kg。
中国林业科学研究院亚热带林业实验中心山下实验林场(FY)位于江西省新余市分宜县西北部(27°44′N, 114°38′E, 海拔90 m)。该地区属亚热带季风气候, 年均降水量1640.9 mm, 年平均气温为17.5℃, 最冷月(1月)均温5.8℃, 极端低温-8.5℃, 最热月(7月)均温29.1℃, 最高温是40.7℃, 日照时数可达1535.3 h。土壤属于黄粘壤土, 土壤pH呈酸性。
将乐林场(JL)位于福建省三明市将乐县(26°41′ N, 117°26′ E, 海拔190 m)。该地区属于亚热带海洋性季风气候, 年均降水量1665 mm, 其中雨季(3—9月)占年降水量的79.8%, 年蒸发量1204 mm, 无霜期300 d。区域年均气温19.8℃, 该地区有明显的干湿季之分, 且干季和低温同期。1月最低月平均气温-4.9℃, 极端最低气温-7.1℃, 7月最高月平均气温36.3℃, 极端最高气温40℃。土壤类型以红壤为主, 腐殖质层较厚, pH值为5—6。
东风林场(DF)位于贵州省黔东南苗族侗族自治州黎平县(26°20′ N, 109°10′ E, 海拔490 m)。该地区气候属于中亚热带温暖湿润季风气候。年平均气温16.2℃, 7月平均温度25.9℃, 最高气温39.7℃, 1月平均温度4.6℃, 最低气温-3.4℃, 年平均日照时数为1317.9 h。年平均降水量1321.9 mm, 蒸发量为1000 mm, 无霜期年平均290 d。地貌类型属于典型的低山丘陵地貌区, 部分存在喀斯特地貌。土壤为红黄壤, pH值5.5—6.5。
高峰林场(GF)位于广西省贺州市昭平县(24°7′ N, 110°52′ E, 海拔520 m)。该地区属亚热带季风气候区, 夏长冬短, 春湿冬干, 具有典型的山区气候特征, 多年平均降水量1927 mm, 年平均气温19.8℃, 7月平均温度27.8℃, 最高气温39.7℃, 1月平均温度9.9℃, 最低气温2.5℃, 无霜期310 d以上。土壤类型为红壤和黄壤, pH值5.0—6.5, 土壤pH处于酸性呈酸性至弱酸性水平。
1.2 样品采集与年表建立2019年9—10月, 在6个杉木人工林进行样地调查和样本采集。每个样地随机选取23株杉木, 使用生长锥分别在每株树木胸高处向东西和南北两个方向垂直钻取两根树芯, 测量并记录其胸径和树高。采集完毕后, 在表面涂上树木生长愈合剂, 防止虫蚁蛀食。样品带回实验室后, 用胶带将树芯粘贴并固定于定制的木槽中, 置于通风处进行阴干。用装有低目砂纸的打磨机进行粗磨, 再用高目砂纸手工打磨, 直至杉木年轮清晰可见。用Lintab 6年轮分析仪和TASP-Win软件对每个样芯进行年轮宽度测定, 测量精度为0.01 mm。利用COFECHA软件进行交叉定年, 最终分别形成一组可以代表每个样点的宽度序列。使用“rwl.stats”函数计算原始树轮宽度序列和树轮宽度指数年表的基本统计量。由于树木的生长不仅受环境的影响, 而且还受到自身生理因子的控制, 本研究中样本树龄较短, 可能会导致年轮宽度上的低频变化, 因此使用“detrend.series”函数剔除大部分的低频变化[21], 去掉树木个体特有的前期生理条件对后期生长造成的连续性影响, 以达到去除与树龄相关的生长趋势的目的。用“rwi.stats”函数对树轮宽度指数序列进行公共区间分析。使用“chron”函数建立年表, 最终得到差值年表(Residual chronology, RES)[22]。
1.3 胸高断面积增长为了更深入地研究树木随气候变化的生长模式和趋势, 将原始数据的轮宽序列转化成胸高断面积增长(Basal area increment, BAI)进行研究。BAI是用来量化树木生长及生长趋势的变量, 其克服了衰老导致树木生长下降的问题以及数据转换带来的不可避免的偏差。其公式如下[23]:

|
(1) |
式中, Rn是n年的树盘半径, Rn-1是n-1年的半径。
1.4 气候数据获取和抗旱性指标的确定本研究使用的气候数据为样地附近的国家标准气象台站提供的数据进行插值得出。考虑到气候因子对树木年轮宽度的影响具有“滞后效应”, 选取1967—2019年的5个气候因子, 主要是上一年6月至当年10月期间的最高气温、最低气温、平均气温、降水量、相对湿度。干旱指数(standardized precipitation evaporation index, SPEI)数据来源于CRU TS 4.02全球气候数据库, 根据干旱指数等级划分, 将前一年10月至当年9月的平均SPEI<-1的年份设定为干旱年, 考虑到人工林年龄较短, 为避免幼龄效应和干旱重叠的影响, 选取了研究区树木生长过程中最近20年中同时发生的2次干旱事件, 分别是2004年和2011年, 干旱前和干旱后的时期均设定为3年。其中, 在2004年到2011年期间, 分宜在2007—2009年连续发生了干旱事件;将乐在2008年发生了干旱事件;东风在2006和2010年发生了干旱事件。
干旱时树木的生态弹性指标分别用抵抗力(Rt)、恢复力(Rc)、弹性力(Rs)来表示[24]:

|
(2) |

|
(3) |

|
(4) |
式中, Gd表示干旱事件发生时的平均树轮宽度, Gprev表示干旱事件发生前4年的平均树轮宽度, Gpost表示干旱事件发生后4年的平均树轮宽度。
抵抗力表示树木抵抗干旱事件的能力恢复力表示干旱事件发生后树木恢复生长的能力, 弹性力为干旱事件发生前后树木生长的差异, 即抵抗力与恢复力的比值[25]。其中干旱事件的发生需满足SPEI≤-1[26]。假设在任意抵抗力下树木的恢复弹力均等于1, 将其定义为完全恢复弹力标准线, 表示在干旱事件发生前后树木的生长可以完全恢复。通过比较树木生长的实际抵抗力、恢复力关系与完全恢复弹力标准线, 以探究树木实际恢复弹力。
1.5 统计分析采用Mann-Kendall(M-K)趋势检测法检验研究区杉木BAI的变化, 通过R语言的dplR扩展包建立年表, 综合分析树轮宽度差值年表与气象数据, 树木抵抗力、恢复力、弹性力与SPEI, 树木对干旱事件的实际恢复弹力与完全恢复弹力标准线三组数据间的关系, 利用SPSS进行相关性分析。使用Microsoft excel制作表格, 利用Origin-Pro2019软件进行绘图, 分别得出树木径向生长对气候和干旱事件的响应机制。
2 结果与分析 2.1 气候因子的变化本研究6个站点的年降水量都大于400 mm(图 2)。其中, 荆关大多数年份年降水量小于800 mm, 所以荆关属于半湿润地区, 其余5个研究区都属于湿润地区。总体来看, 高峰、将乐和分宜年降雨量比其它地方多, 东风、马鬃岭和荆关次之。年平均温度由高到低分别为高峰(20.1)、将乐(18.7)、分宜(17.8)、东风(16.6)、马鬃岭(16)、荆关(15.3)。6个研究区的年最高温度没有很大的差异。荆关、马鬃岭、分宜、将乐的年最低温都低于0, 荆关的年最低温大部分低于-10, 东风的年最低温在0左右, 高峰的年最低温都在0以上。将乐的年均相对湿度最大, 约在81%以上;东风、分宜和高峰次之, 在80%左右;荆关和马鬃岭的年均相对湿度较小, 分别在71%和75%左右。总体来看, 将乐、荆关的年均相对湿度整体呈下降趋势。荆关的年SPEI值介于重度干旱和轻度湿润之间(-1.5 < SPEI < 1.0), 1995、2004、2011和2014年的SPEI值最小, 小于-1;马鬃岭的年SPEI值介于重度干旱和中度湿润之间(-1.5 < SPEI < 1.5), 2004、2011年的SPEI值最小, 小于-1;东风的年SPEI值介于重度干旱和中度湿润之间(-2 < SPEI < 1.5), 2010、2011年的SPEI值最小, 在-1.5左右;高峰的年SPEI值介于中度干旱和重度湿润之间(-1.5 < SPEI < 2.0), 1989、1991、2004和2011年的SPEI值最小, 小于-1, 该4个研究区均无极端干旱和或极端湿润发生。而分宜在1998年存在极端湿润的发生, 2004和2008年的SPEI值最小, 小于-1.5, 达到重度干旱;将乐存在极端干旱和极端湿润的情况, 分别在2004年和1998年, 表明这两个研究区整体水分收支不平衡, 有明显偏干和偏湿的趋势, 且有一定的年份同步性。

|
| 图 2 6个研究区降水量、平均温度、最高温度、最低温度、相对湿度、标准化降水蒸散指数(SPEI)的年变化 Fig. 2 The trend of annual precipitation, annual average temperature, annual maximum temperature, annual minimum temperature, annual relative humidity, and SPEI at the six study areas SPEI: Standardized precipitation evaporation index |
本文选用树轮宽度差值年表(RES)用于分析, 差值年表具有低频变化较少、离散程度较低、逐年变化共性较高的特点, 能够代表采样点群体的年轮特征, 年表统计学参数包括:平均敏感度(MS)、样本间相关系数(RBAR)、信噪比(SNR)、样本总体代表性(EPS)(表 1)。6个研究区MS均大于0.15, 表明杉木径向生长的变化包含许多的气候信息且制约作用明显。RBAR越高代表数据质量越好[25], 各样点RBAR为0.206—0.348, 表明采样点杉木的年轮宽度变化的一致性较好。各年表的SNR均处于较高水平, 表明年表能够反映出气候高频变化[27]。EPS反映了年表的可靠性, 所有样本对群体的表达信号为0.935—0.970, 均在可信临界数值(0.85)以上, 说明样本所含气候信号能代表总体特征, 具有区域代表性。
| 站点 Site |
时间序列 Time series |
树龄 Stand age |
样本间平均相关系数 Mean inter-series correlation |
样本总体代表性 Expressed population signal |
信噪比 Signal to noise ratio |
平均敏感度 Mean sensitivity |
| 荆关 | 1976—2019 | 20—44 | 0.243 | 0.967 | 29.458 | 0.297 |
| 马鬃岭 | 1968—2019 | 17—52 | 0.213 | 0.951 | 19.572 | 0.235 |
| 分宜 | 1985—2019 | 20—35 | 0.232 | 0.949 | 18.509 | 0.278 |
| 将乐 | 1986—2019 | 23—34 | 0.358 | 0.935 | 34.294 | 0.224 |
| 东风 | 1989—2019 | 20—30 | 0.429 | 0.970 | 39.019 | 0.371 |
| 高峰 | 1990—2019 | 21—30 | 0.210 | 0.968 | 29.774 | 0.328 |
6个研究站点的杉木差值年表指数存在一定的差异(图 3)。荆关和将乐的杉木波动频率明显大于其他4个样地的波动频率, 其中荆关年表指数的最小值为0.47, 最大值为1.41, 将乐年表指数最小值为0.65, 最大值为1.41。马鬃岭、分宜、东风和高峰的杉木年表指数则随年份变化的趋势更加明显, 马鬃岭的年表指数在1970—1990整体呈下降趋势, 在1990年出现最低谷值, 为0.58, 随后上升并逐渐趋于平稳。分宜、东风和高峰的年表指数变化趋势相似, 先上升后在2002—2010年呈下降趋势, 2010后又逐渐上升。马鬃岭年表指数的最小值为0.58, 最大值为14.45;分宜年表值最小值为0.30, 最大值为1.35;东风年表值最小值为0.53, 最大值为1.61;高峰年表值最小值为0.63, 最大值为1.25, 东风的年表指数变化幅度最大。

|
| 图 3 不同站点杉木差值年表指数(RCI) Fig. 3 Residual chronology index (RCI) of Cunninghamia lanceolata at different sites |
通过分析6个研究区杉木BAI的变化(图 4), 发现6个研究区杉木BAI总体呈先上升后下降的趋势(α<0.05), 且均在前十几年甚至前几十年呈现出较大幅度的线性生长趋势, 说明杉木人工林在初期生长发育良好, 有研究显示未衰老以及未受到环境压力的树木, BAI将一直保持一种线性关系[28], 其中将乐、东风和高峰的生长速率相较于其他三个研究区更快, 说明不同的环境条件限制杉木径向生长速率。在生长后期, 6个研究区的BAI出现不同程度的衰退现象, 其中荆关的衰退程度较大。

|
| 图 4 胸高断面积增量(BAI) 变化 Fig. 4 Variations of basal area increment |
各研究站点RES年表与气候因子的相关关系分析结果表明(图 5)。荆关的RES年表与当年2、7月份的平均温度、上年9月以及当年5、6、11月的最低温度、当年9月的最高温度和当年6月的平均降雨量显著正相关(P < 0.05);与上年11月的最高温度极显著正相关(P < 0.01);与上年6月最高温度显著负相关(P < 0.05)。相对湿度和SPEI对RES则无显著影响。

|
| 图 5 差值年表对平均温度(Ta)、最低温度(Tmin)、最高温度(Tmax)、降雨(Pre)、相对湿度(RH) 和标准化降水蒸散指数(SPEI) 的响应相关性系数 Fig. 5 Pearson correlation coefficients between residual chronology and mean temperature (Ta), minimum temperature (Tmin), maximum temperature (Tmax), precipitation (Pre), relative humidity (RH), and standardized precipitation evapotranspiration index (SPEI) “*”表示P<0.05, “* *”表示P<0.01;C: 当年, P: 上一年 |
马鬃岭站点的RES年表与当年4月的平均降雨显著正相关(P < 0.05);与当年2月的最低温度、当年10月的最高温度和上年11月的平均降雨显著负相关(P < 0.05)。平均温度、相对湿度和SPEI对RSE影响不显著。
对于分宜站点, RES年表与当年3、4月的平均温度、当年3、9月的最低温度和上年7、8月的SPEI显著正相关(P < 0.05);与上年12月的平均温度极显著正相关(P < 0.01);与当年5月的最高温度和当年9月的平均降雨显著负相关(P < 0.05)。RSE年表与相对湿度无显著相关性。
将乐站点的RES年表与上年12月的平均温度、上年6、12月以及当年3月的最低温度、上年10月的最高温度、当年8月的平均降雨以及当年8月的相对湿度显著正相关(P < 0.05);与当年8月最高温度、上年9月平均降雨和当年2月相对湿度显著负相关(P < 0.05)。该站RSE年表与SPEI无显著相关关系。
在东风杉木人工林站点, RES年表与上年6、12月的平均温度、当年1月最低温度、上年8月平均降雨、上年7月以及当年6月的SPEI显著正相关(P < 0.05);与上年8—12月以及当年1—5月SPEI极显著正相关(P < 0.01)。最高温度和相对湿度对该站树木RES年表的影响则不显著。
在高峰站点, RES年表与当年1月平均温度、上年6月以及当年1月的最低温度、上年8月的平均降雨、上年7、11月以及当年1、6、7、9月的相对湿度、上年8、10月以及当年2、3、4、6月的SPEI显著正相关(P < 0.05);与当年4月的平均降雨、上年8—10月以及当年5、8、10月的相对湿度、当年5、7月的SPEI极显著正相关(P < 0.01)。RES年表上年9、11月的最高温度、当年11月的平均降雨显著负相关(P < 0.05), 与上年8月以及当年8月的平均温度、上年8月的最高温度极显著负相关(P < 0.01)。
2.5 树木径向生长对干旱事件的响应不同研究区杉木对干旱事件的响应存在差异性(图 6)。在2004年第一次干旱事件发生时, 不同研究区杉木对干旱的抵抗力由大到小分别为将乐、荆关、马鬃岭、分宜、高峰和东风, 其中将乐的抵抗力指数普遍大于1.0, 荆关和马鬃岭的抵抗力指数大部分都大于1.0, 其他3个研究区普遍都低于1.0;对干旱的恢复力较大的为分宜, 其次为马鬃岭和东风, 较小的为高峰、荆关和将乐, 其中马鬃岭、分宜和东风的恢复力指数大部分大于1.0, 其他3个区域普遍都低于1.0。对干旱的弹性力由大到小分别为马鬃岭、分宜、荆关、东风、将乐和高峰, 其中马鬃岭和分宜的弹性力指数部分大于1.0, 其他4个区域的弹性力指数普遍小于1.0。

|
| 图 6 杉木应对干旱事件的抵抗力、恢复力和弹性指数 Fig. 6 Resistance, recovery, and resilience of Cunninghamia lanceolata to drought events |
在2011年第二次干旱事件发生时, 不同研究区杉木对干旱的抵抗力由大到小分别为荆关、马鬃岭、分宜、东风、将乐和高峰, 其中荆关的抵抗力指数普遍大于1.0, 其他5个研究区的抵抗力指数大多数都小于1.0;对干旱的恢复力由大到小分别为将乐、东风、高峰、分宜、马鬃岭和荆关, 其中将乐的恢复力指数普遍大于1.0, 东风和高峰的恢复力指数部分大于1.0, 其他3个区域都大部分小于1.0。对干旱的弹性力最大为将乐, 次之东风、高峰、分宜和马鬃岭, 最小为荆关, 其中将乐的弹性力指数大部分大于1.0, 其他5个研究区普遍都低于1.0。
对比前后两次干旱事件的抵抗力、恢复力和弹性力, 荆关、分宜、东风和高峰的抵抗力增加, 马鬃岭和将乐的抵抗力减少;分宜、东风和高峰的恢复力增加, 荆关、马鬃岭和将乐的恢复减少;荆关、分宜、将乐、东风和高峰的弹性力增加;马鬃岭的弹性力减少。
从6个研究区抵抗力与恢复弹力的指标可看出(图 7), 两次干旱事件的实际恢复弹力和完全恢复弹力具有显著关联性。第一次干旱事件发生时关联性由高到低为:马鬃岭、分宜、将乐、高峰、荆关、东风, 关联性分别为0.68、0.61、0.50、0.38、0.29和0.25。第二次干旱事件发生时关联性由高到低分别为:分宜、将乐、荆关、高峰、东风、马鬃岭, 关联性分别为0.60、0.55、0.49、0.44、0.31、0.30。其中, 荆关第二次干旱事件的恢复弹力大于第一次恢复弹力, 马鬃岭第一次干旱事件的恢复弹力大于第二次的恢复弹力, 分宜第一次干旱事件的恢复弹力和第二次的恢复弹力都处于较高水平, 且无明显差异。将乐第二次干旱事件的恢复弹力大于第一次恢复弹力;东风第二次干旱事件的恢复弹力大于第一次恢复弹力;将乐第二次干旱事件的恢复弹力大于第一次恢复弹力。

|
| 图 7 树木对干旱事件完全恢复弹力标准线和实际恢复弹力 Fig. 7 The full resilience line and the actual resilience to the drought event |
六个研究区的杉木径向生长均与生长季的平均温度和降水量、上一年夏季的最低温度正相关。经过冬季的蛰伏后杉木逐渐开始生长, 这一时期随着气温的升高, 树木细胞分裂、生长加快, 充足的降水满足自身和树木蒸腾的需要, 林木进行光合作用制造大量有机质促进树木的生长[29]。因此, 温度和降水量的提高为处于生长季的杉木提供了必要的生长条件[30]。同时, 夏季温度对杉木径向生长表现出的抑制作用在六个研究区都得以体现, 这与很多学者研究得出的结论一致[31—32]。高温会引起树木气孔关闭, 减弱杉木的光合作用[5, 33]。南、中亚热带地区杉木径向生长与气候关系存在较大差异, 导致差异的原因在于不同研究区的杉木生长物候节律的差异。在中亚热带地区的荆关和马鬃岭站点, 冬季的最低温度低于-10℃, 当最低温度的升高时, 积雪的融化导致水分流失, 林木结束休眠开始生长时需要大量的水分, 水分的减少影响林木的早期生长[34]。到3—5月, 纬度较高的站点温度相对较低, 温度的升高有利于形成层的活化, 影响木质部生长起始和持续时间[35]。马鬃岭也属于纬度相对较高的地区, 但在该研究区温度的升高对杉木生长的积极影响并没有表现出显著水平, 这可能是马鬃岭的海拔比较高, 海拔高紫外线辐射严重, 高辐射会使树木的蒸散量增加, 温度与蒸散量的变化达到一个动态平衡[36]。在纬度较低的高峰站点, 杉木径向生长与上年生长季末期的温度呈负相关, 高峰夏季较长, 9、10月的温度较高, 高温会造成树木净光合产物减少, 降低形成层活性, 影响细胞分化, 加剧干旱胁迫[8, 37], 且生长季末期为种子成熟期, 温度升高积累的营养物质用于繁殖而不是用于来年的生长[38]。此外, 荆关和东风站点杉木径向生长与当年夏季温度具有正相关关系;将乐和高峰站点杉木径向生长则与上年6月的最低温度呈正相关。结合研究区的特点, 荆关属于北亚热带和暖温带交汇地带, 夏季温度较低, 而东风由于地理位置的优势, 夏无酷暑, 两个研究区的夏季温度不会过高, 温度在适宜范围内升高有利于林木生长[39—40]。与上一年6月最低温度正相关是因为最低温度的升高可以为下一个生长季提供了充足的营养物质[41]。其中, 荆关上年6月最高温度和当年最低温度的相反效应可能体现了水分的重要性, 即温度较高情况下, 降水不足会抑制树木生长[42—43]。荆关和将乐上年生长季末期的温度以及荆关、马鬃岭和分宜当年生长季末期的温度与杉木径向生长呈正相关, 说明在生长季末期温度升高有助于杉木继续进行生理活动[44—45], 这与周志强等[46]树木年轮的研究结果一致。
在降水方面, 6个研究区的径向生长与降水量的相关性较小, 且生长于不同纬度的杉木, 其径向生长对降雨的响应差异并不明显。在马鬃岭和高峰站点, 径向生长与当年4月的降水呈正相关。这是因为春季杉木逐渐进入生长期, 充足的降水可以较早地打破树木的休眠状态, 有利于树木的萌发与生长[47], 对杉木的生长的有促进作用。其他4个研究区径向生长与当年4月份降水关系并不显著。其中, 杉木径向生长与荆关、分宜当年春季温度相关性达到显著性水平, 说明该站点在春季限制树木生长的主要是温度。将乐、东风的降水较多, 在湿润的环境林木对降水的变化不敏感, 高峰虽然降水量大, 但其4月的平均温度高, 水分蒸发速度快, 充足的降水对杉木的生长更有利。荆关、将乐、东风和高峰径向生长均与夏季降水量具有正相关关系, 这是因为夏季研究区内温度较高, 高温加快了水分的蒸发, 充足的降水可以减弱水分胁迫, 有利于杉木形成宽轮[48]。在马鬃岭、分宜、将乐和高峰站点, 径向生长与秋季降水量呈负相关。秋季温度降低, 水分蒸发速度降低, 同时林木处于生长季后期, 对水分需求减少, 降水量的增加可能会引起土壤水分过多, 从而影响根的呼吸作用[49]。降水对杉木生理过程的直接作用以及影响温度与光照的间接作用同样适用于解释冬季相对湿度对杉木径向生长的负影响, 说明了温度和降水对树木的径向生长有复杂的交互作用[50]。在将乐和高峰站点, 杉木径向生长与生长季相对湿度呈正相关。在生长季, 较高的相对湿度对应适宜的温度和充足的水分, 可以促进树木的生长。
本研究发现, 分宜站点7、8月、东风站点上年7月到当年6月、高峰站点上年8、10月以及当年2—7月的SPEI与杉木径向生长始呈显著正相关关系。梁尔源等[51]的研究也发现, 树木生长与上一年7月至当年8月的干旱指数存在显著正相关关系。这说明随着温度的逐渐升高, 当降水量不足时, 干旱状况成为限制杉木径向生长的主要气候因子。树木在遭受干旱胁迫时, 会通过减小叶面积和关闭气孔的方式来减少水分蒸发, 但这种生理应对措施会导致林木光合作用的能力减弱[52—53]。
3.2 不同站点杉木径向生长对干旱事件响应的差异两次干旱事件发生后, 林木的弹性力大多数都小于1, 表明干旱事件几乎对林木都产生了负面影响。六个研究区的杉木年轮指数保持相对稳定, 且对干旱事件的抵抗力均大于0.6, 这表明杉木的径向生长并未受到严重的干旱事件干扰[54]。由于研究区地理位置的差异, 杉木径向生长对干旱事件的响应存在差异。在第一次干旱事件发生时, 将乐的抵抗力最大, 原因可能是将乐属于亚热带海洋性季风气候, 该气候对干旱胁迫具有积极的调节作用, 气温和降水条件较好, 有利于树木生长。同时, 杉木对干旱事件的抵抗力与恢复力在纬度的变化上也表现出明显的差异。在纬度较高的站点, 荆关和马鬃岭相较于分宜、高峰和东风的抵抗力较大, 有研究发现当干旱事件发生时, 潮湿环境下生长的树木的死亡率高于较干旱的环境, 长期生长在干旱环境的树木具备维持树木生理生长的适应能力[55]。随着纬度的降低, 分宜、高峰和东风的年平均气温和降水较高, 受到干旱胁迫更强。对多个树种的研究发现, 树木径向生长对干旱的抵抗力和恢复力呈现一种相互平衡的相反关系, 即抵抗力和恢复力通常呈显著负相关[56]。本文中除了马鬃岭和高峰以外其他研究区的抵抗力和恢复力均符合该规律。马鬃岭站点的海拔较高, 高海拔比低海拔有更充足的降水[57], 有效地缓冲干旱年份缺水的不利影响[58]。高峰的抵抗力、恢复力和弹性力都比较小, 这可能的原因是林木受到的干旱胁迫超过了最大忍受程度, 树木的生理机制受损将无法保持原有状态, 树木出现衰退现象[16, 59]。有研究发现, 较低的抵抗力的林木往往在干旱事件中有较高的死亡风险[60—61]。气候变暖导致干旱事件发生的强度、频率增加, 一般来说第二次干旱发生后, 树木的抵抗和恢复能力会下降。分宜、东风和高峰的抵抗力和恢复力相较于第一次干旱事件均得到提高, 其中分宜和东风在第一次干旱和第二次干旱期间还受到了连续干旱事件的影响, 这可能是树木在遭受干旱事件后适应了不利的生存条件, 从而产生了更强的抵抗力和恢复力[62—63]。在东风和高峰站点, 杉木的恢复弹力在两次干旱事件发生过程中比较小。这是因为在2004—2011年东风和高峰站的SPEI均较小, 干旱胁迫的压力累积效应明显, 导致树势降低, 恢复弹力也相对较小。在荆关和马鬃岭站点, 两次干旱事件发生时杉木的恢复弹力不同。这可能是因为纬度较高的站点受到干旱胁迫的影响较小, 第一次干旱事件发生对林木生长的影响未超过恢复弹力的阈值;当第二次干旱发生时, 干旱事件的累积效应则超过了恢复弹力的阈值, 树木生长下降。
4 结论6个研究点上杉木的生长对气候因子的响应表现出一定的相似性, 均与生长季的水热条件有关, 杉木年轮宽度指数与当年生长季平均温度和降水量、上一年夏季最低温度成正相关, 与当年夏季最高温度成负相关, 且对平均温度、最高温度和最低温度的响应比对平均降水的响应程度更显著。杉木BAI都呈先上升后下降的趋势, 且都受到了干旱事件的负面影响。不同研究点上的杉木对干旱的响应也表现出差异, 较低纬度研究点上的杉木受到干旱胁迫的影响较大, 温暖湿润环境下的杉木对干旱事件的抵抗力较小, 恢复力较高, 第二次干旱下杉木对干旱事件的应对能力更强。在未来干旱事件强度和频率可能增加的情形下, 需对亚热带不同水热条件的杉木干旱耐受性进行差异化评估, 并采取针对性解决方案, 防止杉木因干旱胁迫导致生长下降。除气候因子外, 杉木生长还受林分属性、地理因子、立地条件等复杂因素的影响, 区分并解析这些因子对杉木生长影响的相对重要性, 将更好地理解不同气候地理区杉木生长的限制因子和影响程度, 为杉木人工林精细化经营提供依据。
| [1] |
Huang M T, Wang X H, Keenan T F, Piao S L. Drought timing influences the legacy of tree growth recovery. Global Change Biology, 2018, 24(8): 3546-3559. DOI:10.1111/gcb.14294 |
| [2] |
Li Z S, Keyimu M, Fan Z, Wang X C. Climate sensitivity of conifer growth doesn't reveal distinct low-high dipole along the elevation gradient in the Wolong National Natural Reserve, SW China. Dendrochronologia, 2020, 61: 125702. DOI:10.1016/j.dendro.2020.125702 |
| [3] |
Gazol A, Ribas M, Gutiérrez E, Camarero J J. Aleppo pine forests from across Spain show drought-induced growth decline and partial recovery. Agricultural and Forest Meteorology, 2017, 232: 186-194. DOI:10.1016/j.agrformet.2016.08.014 |
| [4] |
赵志江, 郭文霞, 康东伟, 崔莉, 赵联军, 李俊清. 川西亚高山岷江冷杉和紫果云杉径向生长对气候因子的响应. 林业科学, 2019, 55(7): 1-16. |
| [5] |
Wehr R, Commane R, Munger J W, McManus J B, Nelson D D, Zahniser M S, Saleska S R, Wofsy S C. Dynamics of canopy stomatal conductance, transpiration, and evaporation in a temperate deciduous forest, validated by carbonyl sulfide uptake. Biogeosciences, 2017, 14(2): 389-401. DOI:10.5194/bg-14-389-2017 |
| [6] |
Liu X S, Nie Y Q, Wen F. Seasonal dynamics of stem radial increment of Pinus taiwanensis Hayata and its response to environmental factors in the Lushan Mountains, southeastern China. Forests, 2018, 9(7): 387. DOI:10.3390/f9070387 |
| [7] |
Zhang X, Fan Z F, Shi Z J, Pan L L, Kwon S, Yang X H, Liu Y S. Tree characteristics and drought severity modulate the growth resilience of natural Mongolian pine to extreme drought episodes. Science of the Total Environment, 2022, 830: 154742. DOI:10.1016/j.scitotenv.2022.154742 |
| [8] |
李夏榕, 陈怡歆, 陈静飞, 朱济友, 孙广鹏, 韦柳端, 张新娜, 徐程扬. 气候变化对北京近远郊地区油松径向生长影响的比较研究. 北京林业大学学报, 2022, 44(1): 19-28. |
| [9] |
Cavin L, Jump A S. Highest drought sensitivity and lowest resistance to growth suppression are found in the range core of the tree Fagus sylvatica L. not the equatorial range edge. Global Change Biology, 2017, 23(1): 362-379. DOI:10.1111/gcb.13366 |
| [10] |
Vitali V, Büntgen U, Bauhus J. Silver fir and Douglas fir are more tolerant to extreme droughts than Norway spruce in south-western Germany. Global Change Biology, 2017, 23(12): 5108-5119. DOI:10.1111/gcb.13774 |
| [11] |
Cai Q F, Liu Y. Two centuries temperature variations over subtropical southeast China inferred from Pinus taiwanensis Hayata tree-ring width. Climate Dynamics, 2017, 48(5): 1813-1825. |
| [12] |
刘兰娅, 勾晓华, 张芬, 尹定财, 王学佳, 夏敬清, 李芊, 杜苗苗. 升温对祁连山东部青海云杉径向生长的影响. 应用生态学报, 2021, 32(10): 3576-3584. |
| [13] |
王红桃, 包光. 秦岭及周边地区树木径向生长与气候因子的关系研究进展. 亚热带资源与环境学报, 2016, 11(1): 52-58. DOI:10.3969/j.issn.1673-7105.2016.01.008 |
| [14] |
Huang Y Q, Deng X W, Zhao Z H, Xiang W H, Yan W D, Ouyang S, Lei P F. Monthly radial growth model of Chinese fir (Cunninghamia lanceolata (lamb.) hook.), and the relationships between radial increment and climate factors. Forests, 2019, 10(9): 757. DOI:10.3390/f10090757 |
| [15] |
李建, 张国文, 刘阳. 基于灰色系统分析的井冈山杉木年轮生长的气候因子研究. 安徽农业科学, 2018, 46(22): 92-95. DOI:10.3969/j.issn.0517-6611.2018.22.028 |
| [16] |
张辉, 颜耀, 胡亚楠, 黄智军, 吴鹏飞, 马祥庆. 基于树轮气候相关分析法的杉木径向生长与气候因子关系. 福建农林大学学报: 自然科学版, 2020, 49(1): 59-66. |
| [17] |
曾伟, 江斌, 熊彩云, 肖复明, 张开珍. 江西省不同林龄杉木碳储量分配格局. 中国农学通报, 2015, 31(28): 1-5. DOI:10.11924/j.issn.1000-6850.casb15070038 |
| [18] |
朱安明, 段爱国, 张建国, 张雄清. 广西柳州杉木种源树轮宽度对温度与湿度的年际响应. 林业科学研究, 2017, 30(1): 53-62. |
| [19] |
孟盛旺, 杨风亭, 戴晓琴, 王辉民. 杉木径向生长动态及其对季节性干旱的响应. 应用生态学报, 2021, 32(10): 3521-3530. |
| [20] |
Rahman M H, Begum S, Nakaba S, Yamagishi Y, Kudo K, Nabeshima E, Nugroho W D, Oribe Y, Funada R. Relationship between the earlywood-to-latewood transition and changes in levels of stored starch around the cambium in locally heated stems of the evergreen conifer Chamaecyparis pisifera. Trees, 2016, 30(5): 1619-1631. DOI:10.1007/s00468-016-1395-4 |
| [21] |
黄蕴凯. 气候和竞争对莽山华南五针松径向生长的影响[D]. 长沙: 中南林业科技大学, 2021.
|
| [22] |
赵守栋, 江源, 焦亮, 王明昌, 张凌楠, 李文卿. ARSTAN程序和R语言dplR扩展包进行树轮年表分析的比较研究. 生态学报, 2015, 35(22): 7494-7502. |
| [23] |
Huang R, Zhu H F, Liu X H, Liang E Y, Grießinger J, Wu G J, Li X X, Bräuning A. Does increasing intrinsic water use efficiency (iWUE) stimulate tree growth at natural alpine timberline on the southeastern Tibetan Plateau?. Global and Planetary Change, 2017, 148: 217-226. DOI:10.1016/j.gloplacha.2016.11.017 |
| [24] |
Fang O Y, Zhang Q B. Tree resilience to drought increases in the Tibetan Plateau. Global Change Biology, 2018, 25(1): 245-253. |
| [25] |
Lloret F, Keeling E G, Sala A N. Components of tree resilience: effects of successive low-growth episodes in old ponderosa pine forests. Oikos, 2011, 120(12): 1909-1920. DOI:10.1111/j.1600-0706.2011.19372.x |
| [26] |
Serra-Maluquer X, Mencuccini M, Martínez-Vilalta J. Changes in tree resistance, recovery and resilience across three successive extreme droughts in the northeast Iberian Peninsula. Oecologia, 2018, 187(1): 343-354. DOI:10.1007/s00442-018-4118-2 |
| [27] |
方克艳, 杨保, 郑怀舟, 李颖俊, 周非飞, 董志鹏, 曹春福, 李大稳, 陈亚君. 树轮学研究方法及其在全球变化中的应用. 第四纪研究, 2015, 35(5): 1283-1293. |
| [28] |
Matías L, Linares J C, Sánchez-Miranda Á, Jump A S. Contrasting growth forecasts across the geographical range of Scots pine due to altitudinal and latitudinal differences in climatic sensitivity. Global Change Biology, 2017, 23(10): 4106-4116. DOI:10.1111/gcb.13627 |
| [29] |
Yang B, He M H, Shishov V, Tychkov I, Vaganov E A, Rossi S, Ljungqvist F, Grießinger J. New perspective on spring vegetation phenology and global climate change based on Tibetan Plateau tree-ring data. Proceedings of the National Academy of Sciences, 2017, 114(27): 6966-6971. DOI:10.1073/pnas.1616608114 |
| [30] |
杨镒如, 张茗珊, 张凌楠, 路强强, 洪艺雪, 刘晓宏. 秦岭中西部油松径向生长对气候因子的响应差异研究. 生态学报, 2022, 42(4): 1474-1486. |
| [31] |
包光, 刘治野, 刘娜, 吴买利. 呼伦贝尔沙地樟子松径向生长特征的VS模型模拟分析. 应用生态学报, 2021, 32(10): 3448-3458. |
| [32] |
Lyu L X, Zhang Q B, Pellatt M G, Büntgen U, Li M H, Cherubini P. Drought limitation on tree growth at the Northern Hemisphere's highest tree line. Dendrochronologia, 2019, 53: 40-47. DOI:10.1016/j.dendro.2018.11.006 |
| [33] |
张晴, 于瑞德, 郑宏伟, 杨美琳, 甘淼. 天山东部不同海拔西伯利亚落叶松对气候变暖的响应分析. 植物研究, 2018, 38(1): 14-25. |
| [34] |
Eriksson M. Methods of dendrochronology: applications in the environmental sciences. Forest Science, 1991, 37(2): 734-735. |
| [35] |
尚华明, 魏文寿, 袁玉江, 喻树龙, 张同文, 瓦合提·艾则买提, 李新建. 阿尔泰山南坡树轮宽度对气候变暖的响应. 生态学报, 2010, 30(9): 2246-2253. |
| [36] |
李书恒, 侯丽, 史阿荣, 陈兰, 朱显亮, 白红英. 基于Biome-BGC模型及树木年轮的太白红杉林生态系统对气候变化的响应研究. 生态学报, 2018, 38(20): 7435-7446. |
| [37] |
Vieira J, Carvalho A, Campelo F. Tree growth under climate change: evidence from xylogenesis timings and kinetics. Frontiers in Plant Science, 2020, 11: 90. DOI:10.3389/fpls.2020.00090 |
| [38] |
程瑞梅, 刘泽彬, 封晓辉, 肖文发. 气候变化对树木木质部生长影响的研究进展. 林业科学, 2015, 51(6): 147-154. |
| [39] |
Rossi S, Bordeleau A, Morin H, Houle D. The effects of N-enriched rain and warmer soil on the ectomycorrhizae of black spruce remain inconclusive in the short term. Annals of Forest Science, 2013, 70(8): 825-834. DOI:10.1007/s13595-013-0329-1 |
| [40] |
Zhang T W, Zhang R B, Yuan Y J, Gao Y Q, Wei W S, Diushen M, He Q, Shang H M, Wang J. Reconstructed precipitation on a centennial timescale from tree rings in the western Tien Shan Mountains, Central Asia. Quaternary International, 2015, 358: 58-67. DOI:10.1016/j.quaint.2014.10.054 |
| [41] |
余佳霖, 张卫国, 田昆, 松卫红, 李秋平, 杨荣, 张贇. 普达措国家公园海拔上限3个针叶树种径向生长对气候变化的响应. 北京林业大学学报, 2017, 39(1): 43-51. |
| [42] |
李越, 李胜利, 杨昌腾, 旷远文, 曹宗英. 南岭华南五针松树轮宽度对气候因子的响应. 亚热带资源与环境学报, 2016, 11(1): 26-31, 51. |
| [43] |
盖学瑞, 于大炮, 王守乐, 贾翔, 吴健, 周旺明, 周莉, 代力民. 树轮-气候"分异问题"形成机制的研究进展. 生态学杂志, 2017, 36(11): 3273-3280. |
| [44] |
赵志江, 谭留夷, 康东伟, 刘琪璟, 李俊清. 云南小中甸地区丽江云杉径向生长对气候变化的响应. 应用生态学报, 2012, 23(3): 603-609. |
| [45] |
Fan Z X, Bräuning A, Cao K F. Annual temperature reconstruction in the central Hengduan Mountains, China, as deduced from tree rings. Dendrochronologia, 2008, 26(2): 97-107. DOI:10.1016/j.dendro.2008.01.003 |
| [46] |
周志强, 刘彤, 胡林林, 王晓春. 穆棱东北红豆杉年轮-气候关系及其濒危机制. 生态学报, 2010, 30(9): 2304-2310. |
| [47] |
张贇, 尹定财, 田昆, 张卫国, 和荣华, 和文清, 孙江梅, 刘振亚. 玉龙雪山不同海拔丽江云杉径向生长对气候变异的响应. 植物生态学报, 2018, 42(6): 629-639. |
| [48] |
贾飞飞, 孙翠洋, 孙红月, 李鑫. 昌岭山两个优势树种径向生长对气候变化的响应. 生态学报, 2019, 39(17): 6332-6340. |
| [49] |
尚玉昌. 普通生态学 (3版). 北京: 北京大学出版社, 2010.
|
| [50] |
D'Arrigo R, Wilson R, Liepert B, Cherubini P. On the 'Divergence Problem' in Northern Forests: a review of the tree-ring evidence and possible causes. Global and Planetary Change, 2008, 60(3/4): 289-305. |
| [51] |
梁尔源, 邵雪梅, 刘鸿雁, Eckstein Dieter. 树轮所记录的公元1842年以来内蒙古东部浑善达克沙地PDSI的变化. 科学通报, 2007, 52(14): 1694-1699. DOI:10.3321/j.issn:0023-074x.2007.14.016 |
| [52] |
Zhang R B, Yuan Y J, Gou X H, Zhang T W, Zou C, Ji C R, Fan Z A, Qin L, Shang H M, Li X J. Intra-annual radial growth of Schrenk spruce (Picea schrenkiana Fisch. et Mey) and its response to climate on the northern slopes of the Tianshan Mountains. Dendrochronologia, 2016, 40: 36-42. DOI:10.1016/j.dendro.2016.06.002 |
| [53] |
Helman D, Lensky I M, Yakir D, Osem Y. Forests growing under dry conditions have higher hydrological resilience to drought than do more humid forests. Global Change Biology, 2017, 23(7): 2801-2817. DOI:10.1111/gcb.13551 |
| [54] |
阎弘, 孙滢洁, 刘滨辉. 竞争对红松树木的干旱适应性及生长衰退影响. 北京林业大学学报, 2022, 44(6): 1-9. |
| [55] |
Field C, Merino J, Mooney H A. Compromises between water-use efficiency and nitrogen-use efficiency in five species of California evergreens. Oecologia, 1983, 60(3): 384-389. DOI:10.1007/BF00376856 |
| [56] |
Schwarz J, Skiadaresis G, Kohler M, Kunz J, Schnabel F, Vitali V, Bauhus J. Quantifying growth responses of trees to drought-a critique of commonly used resilience indices and recommendations for future studies. Current Forestry Reports, 2020, 6(3): 185-200. DOI:10.1007/s40725-020-00119-2 |
| [57] |
Gao L L, Gou X H, Deng Y, Yang M X, Zhang F. Assessing the influences of tree species, elevation and climate on tree-ring growth in the Qilian Mountains of northwest China. Trees, 2017, 31(2): 393-404. DOI:10.1007/s00468-015-1294-0 |
| [58] |
徐贺年, 王江林, 彭小梅, 任子健. 青藏高原东北部祁连圆柏径向生长对不同类型干旱的响应. 应用生态学报, 2022, 33(8): 2097-2104. |
| [59] |
Allen C R, Angeler D G, Cumming G S, Folke C, Twidwell D, Uden D R. Quantifying spatial resilience. Journal of Applied Ecology, 2016, 53(3): 625-635. DOI:10.1111/1365-2664.12634 |
| [60] |
Hogg E H, Michaelian M, Hook T I, Undershultz M E. Recent climatic drying leads to age-independent growth reductions of white spruce stands in western Canada. Global Change Biology, 2017, 23(12): 5297-5308. DOI:10.1111/gcb.13795 |
| [61] |
DeSoto L, Cailleret M, Sterck F, Jansen S, Kramer K, Robert E M R, Aakala T, Amoroso M M, Bigler C, Camarero J J, Čufar K, Gea-Izquierdo G, Gillner S, Haavik L J, Hereş A M, Kane J M, Kharuk V I, Kitzberger T, Klein T, Levanič T, Linares J C, Mäkinen H, Oberhuber W, Papadopoulos A, Rohner B, Sangüesa-Barreda G, Stojanovic D B, Suárez M L, Villalba R, Martínez-Vilalta J. Low growth resilience to drought is related to future mortality risk in trees. Nature Communications, 2020, 11: 545. |
| [62] |
Mainali J, All J, Jha P K, Bhuju D R. Responses of montane forest to climate variability in the central Himalayas of Nepal. Mountain Research and Development, 2015, 35(1): 66-77. DOI:10.1659/MRD-JOURNAL-D-13-00101.1 |
| [63] |
张齐兵, 方欧娅, 吕利新. 青藏高原树木年轮生态学研究. 北京: 科学出版社, 2019.
|