 2024, Vol. 44
2024, Vol. 44文章信息
- 彭梦, 黄城晨, 李立杰, 余佳妮, 杨帆, 谭凤霞, 柴毅
- PENG Meng, HUANG Chengchen, LI Lijie, YU Jiani, YANG Fan, TAN Fengxia, CHAI Yi
- 长湖浮游植物优势种生态位特征及种间联结性分析
- Niche characteristics and interspecific association of dominant phytoplankton species in Lake Changhu
- 生态学报. 2024, 44(4): 1549-1563
- Acta Ecologica Sinica. 2024, 44(4): 1549-1563
- http://dx.doi.org/10.20103/j.stxb.202212023483
-
文章历史
- 收稿日期: 2022-12-02
- 网络出版日期: 2023-11-28

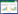




2. 长江大学农学院, 荆州 434025;
3. 长江大学动物科学学院, 荆州 434025
2. College of Agriculture, Yangtze University, Jingzhou 434025, China;
3. College of Animal Science, Yangtze University, Jingzhou 434025, China
生态位是指物种群落在特定的环境中利用资源的能力, 包括生态位宽度与重叠两方面[1]。种间联结性是指不同物种在空间分布上的相互关联性, 是揭示群落构造和演化程度的重要指标, 反映物种在不同生境中相互影响相互作用形成的有机联系及物种间相互吸引或相互排斥的性质[2]。生态位和种间联结是物种群落重要的数量指标, 对研究群落结构功能、特征、演替、种群进化等方面具有指导意义。生态位与种间联结理论共同解释了群落中物种的空间分布及其对资源的利用特征, 两者均会影响群落稳定性。目前, 生态位和种间联结的方法广泛运用在陆地生态系统中[3—5], 在水域生态系统的运用主要集中在鱼类[6—8]、底栖生物[9—11]方面, 在浮游植物的研究中运用相对较少, 仅在部分河流[12—15]、湖泊[14, 16]和水库[17—18]中有报道。
浮游植物是水域生态系统的重要组成部分, 其群落结构和动态变化影响着水体的稳定性和富营养化状况[19]。对浮游植物优势种生态位与种间联结性开展相关研究, 可以帮助了解浮游植物优势种对资源的利用情况及竞争机制和规律, 揭示浮游植物适应性特征与环境变化之间的关系, 为水域生态系统的修复和管理提供科学依据。
长湖是湖北省第三大湖泊, 也是长江中游重要的湿地生态系统[20]。20世纪90年代起, 大面积围栏养殖、面源污染、涉水工程等人类活动导致了长湖出现了鱼类的群落结构改变[21]和富营养化[22]等系列问题。2009年, 长湖获批建立鲌类国家级水产种质资源保护区;2015—2016年, 长湖拆除所有人工养殖设施, 实行退渔还湖政策;2018年, 长湖实施全面禁捕并持续开展鱼类人工增殖放流。本研究对长湖浮游植物优势种生态位特征和种间联结性进行综合分析, 阐明浮游植物群落结构现状, 揭示浮游植物优势种之间的依存和竞争关系, 对生态调控措施后的长湖水域生态系统现状进行评价, 有助于预测浮游植物群落结构变化趋势, 为长湖生态恢复及水环境综合治理提供科学依据, 也为富营养化湖泊治理提供基础数据, 以期实现可持续发展。
1 材料与方法 1.1 采样点设置长湖处于荆州、荆门、潜江三市交界处, 水域面积131km2, 平均水深2.1m。由西向东分为庙湖区、海子湖区、马洪台区和圆心湖区。其中, 庙湖区被堤坝拦截成静水区, 因此本次采样未设置采样点。海子湖区狭长, 马洪台区和圆心湖区湖面较为开阔。因此, 根据湖区形态及参考参考《湖泊富营养化调查规范》在长湖共设置了14个采样点(图 1)。其中海子湖区4个采样点(H1—H4), 马洪台区5个采样点(M1—M5), 圆心湖区5个采样点(Y1—Y5)。于2020年10月(秋)、2021年1月(冬)、2021年4月(春)和2021年7月(夏)分季节对长湖进行采样。

|
| 图 1 长湖浮游植物采样点分布 Fig. 1 Distribution of phytoplankton sampling sites in Lake Changhu |
浮游植物样本的采集参照《淡水浮游生物研究方法》进行[23]。然后参考《中国淡水藻类-系统、分类及生态》[24]及《中国内陆水域常见藻类图谱》[25]用0.1ml浮游生物计数框在光学显微镜(XSM-2.0, 宁波舜宇仪器有限公司, 宁波, 浙江)下进行种类鉴定和计数。
1.3 数据处理 1.3.1 优势种确定优势种根据McNaughton优势度指数(Y)确定, Y>0.02则为优势种, Y>0.1为绝对优势种[16]。
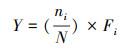
|
式中, ni为第i种在某次采样中各个采样点的总个体数, N为该次采样中各个采样点所有藻类的总个体数, Fi为第i种在各个采样点出现的频率。
1.3.2 生态位测定生态位测度指标包括生态位宽度和生态位重叠。采用Levins指数[26]计算优势种生态位宽度(Bi), 公式如下:
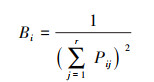
|
式中, Bi为第i种的生态位宽度, Pij为第i种在j个采样点出现的个体数占该种所有个体数的比例, r为采样点数。
采用Pianka[27]生态位重叠指数计算优势种之间的生态位重叠, 公式如下:
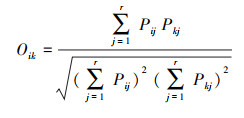
|
式中, k代表不同于物种i的另一种浮游植物;Oik的取值范围为[0, 1]。
1.3.3 浮游植物优势种总体关联强度要从整体上反映群落优势物种的关联性需要进行总体相关性检验, 采用Schluter[28]的方差比率法(Y), 公式如下:
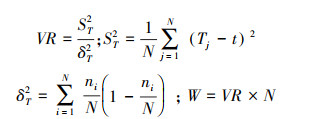
|
式中, S为物种总数, N为采样点总数;t为采样点中浮游植物优势种种数的平均数t(t=(T1+T2+⋯+Tn)/N);ni为物种i出现的采样点数目;Tj为采样点j内出现的优势种的种数。
当VR=1时, 符合所有物种无关联的零假设;当VR < 1时, 物种间总体为负关联; 当VR>1时, 物种间总体为正关联;统计量W=VR×N检验VR值偏离1的显著程度。若W>X0.05(N)2或W < X0.95(N)2, 则物种间总体关联显著(P < 0.05), 若W值落入X2分布的90%置信区间X0.05(N)2 < W < X0.05(N)2内, 则物种间总体关联不显著(P>0.05)。
1.3.4 优势种种对间关系(1) 种间联结性检验
种间联结性从定性的角度反映了物种间的共存概率, 会受到偶见种和伴生种的影响。
根据物种在采样点内是否存在, 将采样点和优势种数量数据矩阵转化为0, 1形式的二元数据矩阵。
基于2×2列联表, 采用Yates连续校正法[29], 进行X2卡方检验。公式如下:
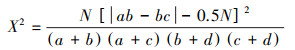
|
式中, N为采样点总数;a为两物种都出现的站位数;b、c分别为仅其中一个物种出现的站位数;d为两物种均未出现的站位数。X2 < 3.84(P>0.05), 表示物种间无显著关联;3.841≤X2 < 6.635(0.01≤P≤0.05), 表示表示物种间关联显著;X2>6.635(P < 0.01)则表示物种间关联性极显著。种对间正负关联取决于(ad-bc)值的正负。若(ad-bc)>0, 则种对间正联结;若(ad-bc) < 0, 则种对间负联结;若(ad-bc)=0, 则种对间无联结。在本研究中, 一些浮游植物优势种出现频率为100%时, 导致卡方检验中的分母为0, 不能准确反映该物种与其他物种间的种间联结关系。因此, 该物种应给一个加权值, 使得b、d值均为1[30]。
(2) 种间相关性检验
卡方检验定性表达了物种种间联结关系, 但是无法从定量角度反映种间相关性[31]。种间相关性的主要检验方法为Pearson相关系数(Pxy)和Spearman秩相关系数(ρ)[32]。公式如下:
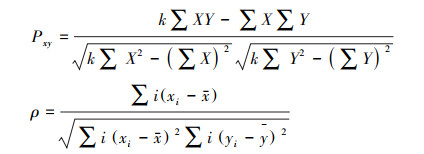
|
式中, X和Y分别为物种x和物种y在同一采样点中的密度;xi和yi分别为植物x和植物y在的同一采样点平均值。
1.4 数据分析使用Excel 2016软件计算浮游植物优势种;使用R4.1.3的spaa程序包计算生态位宽度、生态位重叠指数、种间总体关联强度以及种间联结性检验;psych包进行Pearson相关系数、Spearman秩相关系数计算;使用Excel 2016软件GraphPad Prism 8.0和CorelDRAW 2018绘图。
2 结果与分析 2.1 浮游植物优势种群落结构特征及生态位 2.1.1 浮游植物丰度时空变化及其优势种调查期间共鉴定出浮游植物8门、67属、78种。其中绿藻门出现最多为28属34种。其次是蓝藻门(13属17种)和硅藻门(15属16种), 其余各藻门种类较少。从种类数上来看, 长湖属于绿藻+蓝藻+硅藻为主的水体。
从时间上看, 浮游植物的丰度季节差异显著。浮游植物平均丰度表现为夏季>春季>秋季>冬季, 分别为(1.13±0.53)×107个/L、(7.63±1.62)×106个/L、(5.62±1.18)×106个/L和(3.37±0.88)×106个/L。按照湖泊藻类丰度评价指标[33], 夏季长湖属于中度富营养化状态, 其余三季均处于中度营养状态。秋季以隐藻门、绿藻门和硅藻门为主;冬季以硅藻门和隐藻门为主;春季以硅藻门和绿藻门为主;夏季以绿藻门和蓝藻门为主。绿藻门和硅藻门在三个季节(除冬季)均占据优势, 蓝藻门在夏季有大量繁殖的趋势, 有潜在的富营养化风险。
从空间分布上看, 除夏季外, 其余三季浮游植物丰度空间分布变化不大(如图 2)。秋季各采样点丰度变化范围3.4×106—7.3×106个/L, 丰度最高的采样点为Y5(7.33×106个/L)和Y3(7.19×106个/L);冬季采样点丰度变化范围为2.78×106—6.33×106个/L, Y3点丰度最高(6.33×106个/L);春季各采样点变化范围为5.50×106—10.80×106个/L, M1点丰度最高;夏季各采样点丰度波动较大(3.54×106—20.58×106个/L), H3(20.58×106个/L)和M4(20.10×106个/L)丰度均较大。从各湖区的丰度分布来看(如图 3), 仅夏季各湖区平均丰度变化显著(P < 0.05)。夏季海子湖区和马洪台区平均丰度显著高于圆心湖区。

|
| 图 2 长湖各采样点浮游植物丰度的时空变化 Fig. 2 Spatio-temporal variation of phytoplankton abundance at sampling sites in Lake Changhu |

|
| 图 3 长湖各湖区浮游植物丰度季节变化 Fig. 3 Seasonal variation of phytoplankton abundance in different areas of Lake Changhu |
根据优势度计算结果(Y≥0.02), 共鉴定出浮游植物优势种5门19种(表 1)。其中蓝藻门6种、绿藻门7种、硅藻门4种、裸藻门和隐藻门各1种。其中梅尼小环藻和四尾栅藻在全年占据优势地位。夏季共12个优势种, 蓝藻门6种, 占比50%。其中水华微囊藻和丝状蓝藻(螺旋鱼腥藻、巨颤藻和小席藻)处于优势地位, 然而在其他季节未见优势。隐藻门的啮蚀隐藻在秋季、冬季和春季均占据绝对优势地位(Y≥0.1);硅藻门的梅尼小环藻在冬季和春季占据绝对优势地位(Y≥0.1);绿藻门的四尾栅藻在秋季和春季占据绝对优势地位(Y≥0.1);绿藻门的小型色球藻和蓝藻门细小平裂藻在夏季占据绝对优势地位(Y≥0.1)。
| 门 Phylum |
优势种 Dominant species |
优势度 Degree of dominance | |||
| 秋 Autumn | 冬 Winter | 春 Spring | 夏 Summer | ||
| 硅藻门 Bacillariophyta | 颗粒直链藻 Melosira granulata | 0.084 | — | 0.028 | — |
| 梅尼小环藻 Cyclotella meneghiniana | 0.095 | 0.318* | 0.321* | 0.041 | |
| 肘状针杆藻 Synedra ulna | 0.036 | 0.055 | 0.022 | — | |
| 缘花舟形藻 Navicula radiosa | 0.021 | — | 0.024 | — | |
| 绿藻门 Chlorophyta | 顶锥十字藻 Crucigenia apiculata | 0.028 | 0.021 | 0.066 | |
| 镰型纤维藻 Ankistrodesmus falcatus | 0.041 | 0.052 | — | — | |
| 球衣藻 Chlamydomonas globosa | 0.023 | 0.028 | — | — | |
| 四尾栅藻 Scenedesmus quadricauda | 0.105* | 0.019 | 0.241* | 0.06 | |
| 小型色球藻 Chroococcus minor | 0.02 | — | 0.02 | 0.142* | |
| 螺旋弓形藻 Schroederia spiralis | — | 0.023 | — | 0.026 | |
| 四角十字藻 Crucigenia quadrata | — | — | — | 0.024 | |
| 蓝藻门 Cyanophyta | 细小平裂藻 Merismopedia minima | 0.021 | — | — | 0.155* |
| 巨颤藻 Oscillatoria princeps | — | — | — | 0.023 | |
| 水华微囊藻 Microcystis flos-aquqe | — | — | — | 0.026 | |
| 小席藻 Phormidium tenue | — | — | — | 0.021 | |
| 湖生卵囊藻 Oocystis lacustris | — | — | — | 0.039 | |
| 螺旋鱼腥藻 Anabaebna spirooides | — | — | — | 0.02 | |
| 隐藻门 Cryptophyta | 啮蚀隐藻 Cryptomonas erosa | 0.357* | 0.296* | 0.178* | |
| 裸藻门 Euglenophyta | 尖尾裸藻 Euglena oxyuris | 0.021 | 0.0201 | ||
| “—”表示该物种非本季节优势种;“*”表示该物种为本季节绝对优势种 | |||||
长湖全年浮游植物优势种生态位宽度如表 2所示。19个优势种生态位宽度变化范围为[1.46, 12.31]。其中, 冬季的啮蚀隐藻生态位最宽(12.31), 夏季的水华微囊藻生态位最窄(1.46)。梅尼小环藻、四尾栅藻四季均出现且平均生态位宽度大于7.0, 是长湖水域的第Ⅰ类优势种。该类优势种对资源的利用能力和生境适应能力较强。肘状针杆藻、顶锥十字藻、啮蚀隐藻为长湖水域第Ⅱ类优势种。这一类在三个季节均占优势(仅在高温的夏季或者寒冷的冬季不成为优势种), 且平均生态位宽度大于7, 生态位宽度次于第Ⅰ类。该类优势种对环境具有一定的选择性, 在环境适宜时能够充分利用环境资源。小型色球藻、颗粒直链藻、缘花舟形藻、镰型纤维藻、四尾栅藻、四角十字藻、细小平裂藻、尖尾裸藻, 球衣藻为长湖水域第Ⅲ类优势种。该类优势种虽然生态位宽度也较高, 但是对环境的适应能力较弱, 在特定的环境条件下能较好利用资源。湖生卵囊藻、巨颤藻、水华微囊藻、螺旋鱼腥藻、小席藻仅在某一个季节成为优势种, 为第Ⅳ优势种。该类优势种对环境因子依耐性较强, 水体生境的变化易对其产生影响。
| 优势种 Dominant species |
生态位宽度 niche width | |||
| 秋季 Autumn | 冬季 Winter | 春季 Spring | 夏季 Summer | |
| 颗粒直链藻 Melosira granulata | 10.99 | — | 7.34 | — |
| 梅尼小环藻 Cyclotella meneghiniana | 12.22 | 7.01 | 8.23 | 8.87 |
| 肘状针杆藻 Synedra ulna | 9.65 | 10.57 | 5.60 | — |
| 缘花舟形藻 Navicula radiosa | 7.47 | — | 5.43 | — |
| 顶锥十字藻 Crucigenia apiculata | 8.42 | 8.69 | 7.83 | |
| 镰型纤维藻 Ankistrodesmus falcatus | 10.05 | 7.35 | — | — |
| 球衣藻 Chlamydomonas globosa | 8.59 | 7.83 | — | — |
| 四尾栅藻 Scenedesmus quadricauda | 11.35 | 7.77 | 7.96 | 8.41 |
| 小型色球藻 Chroococcus minor | 7.32 | — | 5.18 | 7.39 |
| 螺旋弓形藻 Schroederia spiralis | — | 8.89 | — | 9.68 |
| 四角十字藻 Crucigenia quadrata | — | 7.83 | — | 6.11 |
| 细小平裂藻 Merismopedia minima | 9.61 | — | — | 7.50 |
| 巨颤藻 Oscillatoria princeps | — | — | — | 6.19 |
| 水华微囊藻 Microcystis flos-aquqe | — | — | — | 1.46 |
| 小席藻 Phormidium tenue | — | — | — | 3.48 |
| 湖生卵囊藻 Oocystis lacustris | — | — | — | 8.75 |
| 螺旋鱼腥藻 Anabaebna spirooides | — | — | — | 6.49 |
| 啮蚀隐藻 Cryptomonas erosa | 9.77 | 12.31 | 5.71 | — |
| 尖尾裸藻 Euglena oxyuris | — | 7.15 | 6.02 | — |
长湖各季节浮游植物优势种生态位重叠值如图 4所示, 秋季共有优势种种对55对, 优势种生态位重叠值在0.02—0.96之间。Wathne等认为生态位重叠值大于0.6, 表示种间生态位重叠较显著[34]。其中Oik>0.6的种对有44对, 占比80.00%, 表明秋季大多数种对间的重叠值较为显著;Oik>0.8的种对有17对, 占比30.91%。秋季Oik>0.9的有4对, 分别为颗粒直链藻-梅尼小环藻、肘状针杆藻-镰型纤维藻、细小平裂藻-镰型纤维藻和四尾栅藻-镰型纤维藻。冬季共有优势种种对28对, 其中Oik>0.6的种对有16对, 占比57.14%。Oik>0.8的种对仅2对, 占比7.14%, 分别为肘状针杆藻-啮蚀隐藻和球衣藻-镰型纤维藻。春季共有优势种种对36对, 其中Oik>0.6的种对有19对, 占比52.78%;Oik>0.8的种对有4对, 分别为小型色球藻-四尾栅藻、小型色球藻-尖尾裸藻、顶锥十字藻-四尾栅藻和四尾栅藻-尖尾裸藻。夏季共有优势种种对66对, 其中Oik>0.6的种对有22对, 占比33.33%, 和其他季节相比差异较大;Oik>0.8的种对共有6对, 占比9.09%。其中夏季特有优势种湖生卵囊藻与小型色球藻/顶锥十字藻/螺旋弓形藻间的Oik均大于0.8。综上, 生态位重叠值为秋季>冬季>春季>夏季。

|
| 图 4 长湖四季浮游植物优势种生态位重叠 Fig. 4 Niche overlap of dominant phytoplankton species in the four seasons of Lake Changhu M.g: 颗粒直链藻 Melosira granulate;C.m: 梅尼小环藻 Cyclotella meneghiniana;S.u: 肘状针杆藻 Synedra ulna;N.r: 缘花舟形藻 Navicula radiosa;C.a: 顶锥十字藻 Crucigenia apiculate;A.f: 镰型纤维藻 Ankistrodesmus falcatus;C.g: 球衣藻 Chlamydomonas globose;S.q: 四尾栅藻 Scenedesmus quadricauda;C.mi: 小型色球藻 Chroococcus minor;S.s: 螺旋弓形藻 Schroederia spiralis;C.q: 四角十字藻 Crucigenia quadrata;M.m: 细小平裂藻 Merismopedia minima;O.p: 巨颤藻 Oscillatoria princeps;M.f: 水华微囊藻 Microcystis flos-aquqe;P.t: 小席藻 Phormidium tenue;O.l: 湖生卵囊藻 Oocystis lacustris;A.s: 螺旋鱼腥藻 Anabaebna spirooides;C.e: 啮蚀隐藻 Cryptomonas erosa;E.o: 尖尾裸藻 Euglena oxyuris |
长湖浮游植物优势种总体联结性分析如表 3所示, 秋季和冬季总体方差比率VR>1, W落在X0.952(df)和X0.052(df)外, 表明秋季和冬季浮游植物优势种间总体呈现显著正联结。春季和夏季总体方差比率VR<1, W落在之间X0.952(df)和X0.052(df), 表明优势种间总体呈现不显著负联结。
| 季节 Seasons |
方差比率 Variance ratio test (VR) |
检验统计量 Statistic (W) |
df | X2的临界值 [X0.952(df), X0.052(df)] |
检验结果 Results |
| 秋季 Autumu | 1.564 | 21.897 | 9 | [3.33, 16.92] | 显著正相关 |
| 冬季 Winter | 1.070 | 14.979 | 6 | [1.64, 12.59] | 显著正相关 |
| 春季 Spring | 0.424 | 4.667 | 7 | [2.17, 14.07] | 不显著负相关 |
| 夏季 Summer | 0.739 | 10.348 | 10 | [3.94, 18.31] | 不显著负相关 |
四季X2检验结果如图 5和表 4所示。其中, 秋季浮游植物优势种共55个种对, 种群间正联结占据优势, 和方差比率法(VR)的检验结果一致。其中, 显著正联结的有4对, 分别为梅尼小环藻-缘花舟形藻、肘状针杆藻-镰型纤维藻、缘花舟形藻-小型色球藻和镰型纤维藻-球衣藻。冬季浮游植物优势种共28个种对, 种群间呈现中性关系。其中显著正联结有1对, 为球衣藻-镰型纤维藻。而其余种对具有相对独立的空间布局。春季共有36个种对, 种群间负联结占据优势, 和总体关联性结果较为一致。其中显著正联结1对, 为肘状针杆藻-小型色球藻。夏季共有66个浮游植物优势种种对, 种群间正联结占据主要地位。其中螺旋弓形藻与湖生卵囊藻和顶锥十字藻均呈现显著正相关关系。

|
| 图 5 长湖四季浮游植物优势种间的X2检验半矩阵 Fig. 5 Semi-matrix of X2 test among dominant phytoplankton species in each season of Lake Changhu |
| 季节 Season |
检验方法 Test method |
正相关 Positive correlations |
负相关 Negative correlations |
正负比 The ratio of positive to negative |
|||||
| 极显著 Extremely significant |
显著 Significant |
不显著 Not significant |
极显著 Extremely significant |
显著 Significant |
不显著 Not significant |
||||
| 秋季 Autumn | X2检验 | 0 | 4 | 25 | 0 | 0 | 26 | 1.12 | |
| Pearson 相关检验 | 2 | 6 | 30 | 0 | 1 | 16 | 2.24 | ||
| Spearman 相关检验 | 2 | 6 | 28 | 1 | 2 | 16 | 1.89 | ||
| 冬季 Winter | X2检验 | 0 | 1 | 13 | 0 | 0 | 14 | 1 | |
| Pearson 相关检验 | 1 | 1 | 11 | 0 | 0 | 15 | 0.87 | ||
| Spearman 相关检验 | 0 | 1 | 11 | 0 | 2 | 14 | 0.75 | ||
| 春季 Spring | X2检验 | 0 | 1 | 15 | 0 | 0 | 20 | 0.8 | |
| Pearson 相关检验 | 1 | 2 | 33 | 0 | 3 | 16 | 0.89 | ||
| Spearman 相关检验 | 0 | 4 | 13 | 0 | 3 | 16 | 0.89 | ||
| 夏季 Summer | X2检验 | 1 | 3 | 37 | 0 | 0 | 25 | 1.65 | |
| Pearson 相关检验 | 3 | 5 | 25 | 0 | 1 | 32 | 1 | ||
| Spearman 相关检验 | 0 | 5 | 40 | 0 | 2 | 19 | 2.14 | ||
| P < 0.05为显著相关;P < 0.01为极显著相关 | |||||||||
Pearson相关性分析结果如图 6和表 4所示, 秋季的55个优势种种对总体呈现正相关关系, 和总体方差比率的结果一致。其中显著相关和极显著相关的种对共计9对, 检测显著率为16.36%, 其余46个种对种间关系较为松散。冬季的28个种对总体略微呈现负相关, 这与X2检验的结果相异。其中显著相关和极显著相关的种对共计2对, 检测显著率为7.14%。春季的36个优势种种对总体略微呈现负相关。其中显著相关和极显著相关的种对共计6对, 检测显著率为16.67%。夏季的66个优势种种对总体呈现中性, 这一结果与X2检验中正联结占据优势的结果相异。其中显著相关和极显著相关的种对共计9对, 检测显著率为13.64%。四季中, 种间Pearson相关性不显著的种对占绝大部分, 种间相关性不强, 这同X2检验的结果相同。

|
| 图 6 长湖浮游植物四季优势种Pearson相关性检验 Fig. 6 Pearson correlation test of dominant phytoplankton species in four seasons of Lake Changhu “**”表示在0.01水平(双侧)上显著相关;“*”表示在0.05水平(双侧)上显著相关 |
Spearman相关性分析结果如图 7和表 4所示。秋季优势种种对整体呈现正相关关系, 和Pearson相关性结果一致。其中, 显著相关和极显著相关的种对共计11对, 检测显著率为20.00%, 其余44个种对种间关系较为松散。冬季的28个种对中, 总体略微呈现负相关, 这与X2检验的结果相异。其中, 显著相关和极显著相关的种对共计3对, 检测显著率为10.71%。春季的36个优势种种对总体略微呈现负相关。其中, 显著相关和极显著相关的种对共计7对, 检测显著率为19.44%。夏季的66个优势种种对总体呈现正相关关系, 这与Pearson的结果相异。其中, 显著相关和极显著相关的种对共计9对, 检测显著率为13.64%。四季中, Spearman相关性指数的显著率总体上高于Pearson相关性指数的显著率。种间Spearman相关性不显著的种对占绝大部分, 种间相关性不强, 这同X2检验的结果相同和Pearson相关性分析的结果一致。

|
| 图 7 长湖浮游植物四季优势种Spearman相关性检验 Fig. 7 Spearman correlation test of dominant phytoplankton species in the four seasons of Lake Changhu “**”表示在0.01水平(双侧)上显著相关;“*”表示在0.05水平(双侧)上显著相关 |
长湖浮游植物季节演替存在着显著的差异。由于高温和丰富的氮磷营养盐[35], 夏季浮游植物丰度最大。有研究表明隐藻主要存在于水温较低的春、冬季节, 水温为其生长的主要限制因子[36]。因此冬季隐藻门成为优势。Oliver等[37]的研究表明, 在较高浓度的氮磷营养盐的水体中, 蓝藻会占据优势地位。因此夏季的高温和丰富的氮磷营养盐使得蓝藻成为夏季优势门类。相关学者对太湖[36]、太平湖[38]、淀山湖[39]等长江流域湖泊的研究中有相似的发现。
从空间分布上看, 长湖夏季各湖区浮游植物丰度差异显著, 海子湖区和马洪台区丰度显著大于圆心湖区。其原因可能是夏季圆心湖区周围生长着较大面积的高等水生植物(荷叶和芦苇)等, 高等水生植物的化感作用对藻类生长起到了抑制作用[40];且圆心湖区水域面积大, 水体交换频繁, 净水能力更好。而海子湖区附近有沮口码头和村落, 人类活动频繁。水体搅动增加、生活污水排放等造成水体氮磷营养盐含量上升, 尤其是夏季水温高, 导致水体藻类大量生长。龚森森等[41]2017年对长湖的研究有相似的发现。
3.2 浮游植物优势种生态位生态位宽度反映了物种利用资源及适应环境的能力[42]。生态位宽度大的藻类可以影响藻类群落的群落结构及水环境状况[16]。因此监测生态位宽度较大的藻类的状况, 可判断出长湖水质本质的变化。梅尼小环藻、四尾栅藻的适宜生境均为富营养化水体[43]。其在长湖四季均为优势种且平均生态位宽度大于7.0, 表明长湖处于富营养化状态。且啮蚀隐藻为α中污带和多污带的指示种[44], 其在秋/冬/春三个季节均为长湖绝对优势种, 对环境的适应能力远超其他优势种, 表明长湖总体上属于中污染水体。这与龚森2020年对长湖夏、秋季的研究结果基本一致[41]。
生态位宽度较小的藻种, 它们种间相互竞争激烈且更趋向于生境特化, 通常可作为水域的指示种[45]。夏季的水华微囊藻和小席藻仅在夏季成为优势种且生态位宽度均小于5, 为狭生态位种。它们占据资源点位和出现的时间相对集中, 且对生境的适应能力较弱。王锦旗的研究表明, 水温是蓝藻门的丝状藻类生长的主要影响因子[46];宋婵媛研究发现栅藻分泌的化感物质对微囊藻有显著的抑制作用[47]。而四尾栅藻为长湖四季共有优势种。因此, 在温度和化感物质的影响下, 其他季节蓝藻门的水华微囊藻和小席藻退出优势地位。而水华微囊藻和小席藻与其他藻类的生态位重叠值均较小, 种间竞争较弱, 极易在夏季大量繁殖, 产生间断性的有毒蓝藻水华, 危害水体健康。
生态位宽度的变化能够反映物种可利用的资源量。可利用资源量较少时, 生态位宽度会增加, 使种群得到足够资源, 可利用资源量丰富时, 生态位宽度会变窄[29]。啮蚀隐藻在秋、冬、春三季均为绝对优势种, 其生态位宽度在春季显著低于秋冬季, 表明春季啮蚀隐藻可利用资源量有所增加。啮蚀隐藻作为富营养化代表物种[48], 表明春季长湖春季富营养化高于秋冬两季。这与浮游植物优势种丰度呈现出来的结果具有较好的一致性。
生态位重叠反映了物种之间利用资源或适应环境能力的相似程度。生态位重叠指数越大, 物种对相同资源的分享程度及适应环境的相似性就越大[49]。本文研究了不同季节的优势种间生态位重叠情况。总体而言, 四季的生态位均有一定的重叠值, 表明浮游植物群落总体上处于稳定状态, 没有形成明显的生境特化。而就生态位重叠值的显著率而言, 秋季>冬季>春季>夏季, 表明秋季优势种间生态学特性相似程度和对资源利用的相似程度为四季最高。然而生态位重叠不一定导致竞争, 竞争是在资源匮乏且生态位高度重叠条件下形成的[49]。长湖水体中氮磷营养盐丰富[41, 50—51], 在资源条件较为充足的条件下, 高的生态位重叠只表明存在微弱的竞争。这与肖艳梅和钟军第的研究结果相似[52—53]。生态位宽度较大(小)的物种与其他物种产生生态位重叠的概率也较大(小)[45]。本研究中梅尼小环藻和四尾栅藻在四季均出现, 且在秋季生态位宽度较大, 其与其他物种的生态重叠值也较高;而夏季水华微囊藻与小席藻的生态位宽度较小, 其与其他物种的生态位重叠值也较小, 这一定程度上导致夏季平均生态位重叠值较小。这与张零念等[54]对木本植物的研究结果一致。
3.3 浮游植物优势种的总体联结性和种间关联性种间总体联结性可反映群落演替进程和物种组成的稳定性[55]。长湖浮游植物优势种的总体联结性结果表明秋冬季节浮游植物群落总体趋向稳定, 环境条件变化不大的情况下, 群落结构不易发生大的波动;春季和夏季浮游植物群落结构虽然存在一定的联系, 但物种间未能形成稳定协调的关系, 以达到对资源的最佳利用状态, 群落整体不稳定, 易发生波动甚至退化, 表现出偶然性和随机性的独立分布格局[56]。
种间联结性是两个物种在空间分布上的联结性, 从定性的角度反映了物种间的相互影响和群落动态[57]。种间相关性是从定量的角度反映两个物种间线性关系和同时出现的可能性程度[58]。种间联结性和种间相关性的检验结果具有一定的一致性和差异性, 两者结合能更好地解释种间关系。
通过结果对比, 本研究发现X2检验、Pearson相关检验和Spearman秩相关检验的结果具有较好的一致性。三种分析方法的结果均显示夏季和秋季种间正联结占据优势, 冬季和春季负联结占据优势。一般来说, 处于演替早期的群落, 物种间的联结程度较低, 正负联结比值偏小, 群落处于不稳定阶段[59]。随着群落的演替更新, 种间联系也逐渐接近于正联结, 群落稳定性也逐渐增强, 群落向顶极方向演替[60]。即冬季和春季处于演替的初期, 夏季和秋季处于演替的中期。夏季和秋季群落总体较为稳定, 而冬季由于水温较低, 抑制了浮游植物体内酶的活性, 浮游植物生长代谢缓慢[61], 而部分藻类难以适应, 数量减少, 物种丰富度减低, 群落总体稳定性低于夏秋季节。春季水温升高, 浮游植物开始增殖;且长湖为草-藻型湖泊[62], 春季水生植物大量生长, 与藻类竞争氮磷等营养盐并分泌化感物质抑制藻类生长[63], 使得春季浮游植物群落稳定性较弱。且在长湖的185个优势种种对中, 三种检验方法的显著率较低, 表明优势种对间关联程度一般, 分布格局相对独立且种间关系较为松散。
种间联结性和种间相关性均能较好地反映一定时期内群落中物种间的相互关系。通常认为, 正联结的种对常表现为互利、互补关系, 负联结的种对更容易发生排斥与竞争[2]。综合三种检测方法显示:秋季肘状针杆藻和镰型纤维藻呈现显著正相关;冬季球衣藻和镰型纤维藻呈现显著正相关;夏季的顶锥十字藻与湖生卵囊藻和螺旋弓形藻呈现显著正相关。结合生态位重叠值的结果本研究发现, 这些物种的生态位重叠值均大于0.8, 表明物种间的正联结性越强, 对资源利用的相似性和生态位重叠的机会越大, 物种间越趋于协同进化。可能的解释是生境中资源丰富或者物种间在部分生态轴上形成了资源需求互补或互利关系, 且在某种程度上该相互作用的存在对一方或双方是有利的。这一规律在许多种间联结和生态位的研究中都得到证实[3, 64]。其中, 球衣藻/镰型纤维藻、顶锥十字藻/螺旋弓形藻这两个种对都为绿藻门的藻类, 其生态学特征较为相似, 资源利用策略较为接近, 出现的位点相对集中。肘状针杆藻/镰型纤维藻、顶锥十字藻/湖生卵囊藻这两个种对虽不属于同一个门, 但在浮游植物功能群理论里, 其适宜生境较为相似[65], 在资源充足的条件下, 种间趋于协同进化。
对比三种方法本研究也发现其结果也存在一定的差异。冬季两种定量分析(Pearson相关性检验和Spearman秩相关检验)结果中负联结的种对比X2检验更多。其原因可能是种间相关性以种对间的多度数据为基础, 可为群落中物种的消长规律提供分析依据, 所指示的物种相互关系与种间联结一致, 但灵敏度有一定区别。种间联结性分析中, 一般当两个种在样方中同时出现或消失时, 种间为正联结;但从定量的角度或许会因为一个种的数量增大而另一个种的数量较小而表现为负相关[66]。因此本研究中两种定量分析方法的结果负联结的对数较多。这与刘益鹏等的研究结果一致[67]。且两种定量检验结果在夏季则分别显示为中性和正相关占据优势。结果的差异可能是算法本身不同导致的。Pearson相关系数检验属于参数检验方法, 要求物种必须服从连续的正态分布, 然而自然界大多数物种均呈集群分布, 从而使得Pearson相关系数检验的结果存在一定局限性;而Spearman秩相关系数检验是一种非参数检验方法, 对物种的分布形式没有任何要求, 灵敏度更高[1]。与之对应的, 本研究中Spearman秩相关系数显著率更高也是其灵敏度更高的证明。这与朱利君、简敏菲等的研究结果一致[58, 68]。
此外, 种间联结性和相关性揭示的仅仅是种间和种内竞争的结果或现状, 并不能揭示导致该结果的生态学过程与机制以及各机制的相对贡献。因此未来应该继续对长湖进行长期监测, 并深入研究各生态因子对该群落稳定性的影响及其潜在机制, 并结合生理学、毒理学、分子生态学和遗传学等学科的先进方法与技术, 深入揭示种间关联性形成的生态学机制。
4 结论(1) 调查期间共鉴定出浮游植物8门、67属、78种。全年共鉴定出优势种19种, 其中, 梅尼小环藻和四尾栅藻在全年占据优势地位。蓝藻门的藻类在夏季有大量繁殖的趋势, 有潜在的有毒蓝藻水华的风险。
(2) 根据生态位理论将长湖优势种分为4类。广生态位的梅尼小环藻和四尾栅藻所指示状况和水体现状一致, 使用广生态位藻类评价水体现状具有可靠的生态学意义。四季的生态位均有一定重叠且秋季>冬季>春季>夏季, 表明浮游植物群落总体上处于稳定状态, 且在资源充足条件下未形成明显的竞争关系。
(3) 种间关联性结果显示:秋冬季节种间总体呈现显著正相关, 群落较稳定;春夏季节种间总体呈现不显著相关, 群落稳定的较弱。种间联结性和种间相关性的结果显示四季优势种种间总体呈现独立的分布格局, 冬季和春季负相关占据优势, 群落处于演替前期;夏季和秋季正相关占据优势, 群落处于演替中期。
| [1] |
Kylafis G, Loreau M. Niche construction in the light of niche theory. Ecology letters, 2011, 14(2): 82-90. DOI:10.1111/j.1461-0248.2010.01551.x |
| [2] |
徐满厚, 刘敏, 翟大彤, 刘彤. 植物种间联结研究内容与方法评述. 生态学报, 2016, 36(24): 8224-8233. |
| [3] |
陈佳卉, 卜元坤, 苏少峰, 黄维, 陈周娟, 李卫忠. 油松飞播林灌木层主要物种种间联结及其环境解释. 生态学杂志, 2021, 40(11): 3512-3522. |
| [4] |
刘润红, 姜勇, 常斌, 李娇凤, 荣春艳, 梁士楚, 杨瑞岸, 刘星童, 曾惠帆, 苏秀丽, 袁海莹, 傅桂焕, 吴燕慧. 漓江河岸带枫杨群落主要木本植物种间联结与相关分析. 生态学报, 2018, 38(19): 6881-6893. |
| [5] |
张乐满, 兰波, 张东升, 刘英杰, 张丽红, 许文锋, 段晨辉, 敦静怡, 刘正学. 三峡水库涪陵—奉节段消落带优势草本植物生态位与种间联结性研究. 生态学报, 2022, 42(8): 3228-3240. |
| [6] |
何雄波, 李波, 王锦溪, 易木荣, 康斌, 颜云榕. 不同时期北部湾日本带鱼营养生态位差异. 应用生态学报, 2021, 32(2): 683-690. |
| [7] |
李凡, 徐炳庆, 吕振波, 王田田. 莱州湾鱼类群落优势种生态位. 生态学报, 2018, 38(14): 5195-5205. |
| [8] |
刘坤, 俞存根, 郑基, 许永久, 颜文超, 韩磊, 刘惠, 孙蓓蓓, 戴冬旭, 于南京. 舟山群岛东侧海域春秋季主要鱼类空间生态位及其分化. 中国水产科学, 2021, 28(1): 100-111. |
| [9] |
刘好真, 李祥付, 刘懂, 李进京, 陈晨, 徐鹏, 孙元, 焦海峰, 王一农, 尤仲杰. 浙江象山东部海域夏季大型底栖动物优势种群和生态位. 海洋与湖沼, 2018, 49(4): 873-882. |
| [10] |
汤雁滨, 廖一波, 寿鹿, 曾江宁, 高爱根, 陈全震. 南麂列岛潮间带大型底栖动物群落优势种生态位. 生态学报, 2016, 36(2): 489-498. |
| [11] |
张苗苗, 王咏雪, 田阔, 田嘉琦, 孙鹏, 水柏年. 沿浦湾秋茄种植前后大型底栖动物生态位和功能群变化. 中国水产科学, 2019, 26(5): 949-958. |
| [12] |
李晓东, 潘成梅, 安瑞志, 巴桑. 西藏拉萨河中下游不同水文期浮游植物优势种生态位及种间联结性. 湖泊科学, 2023, 35(1): 118-130. |
| [13] |
李秋华, 马一明. 桐梓河浮游植物优势种生态位与种间联结性分析. 贵州师范大学学报(自然科学版), 2022, 40(2): 11-18. |
| [14] |
刘洋, 安瑞志, 王陈, 潘成梅, 巴桑. 西藏巴松错(湖)夏季浮游植物优势种生态位及其种间联结性. 水生态学杂志, 1-26. DOI:10.15928/j.1674-3075.202203140071 |
| [15] |
吴佳梦, 徐娜娜, 张文珺, 许丽, 胡婧, 朱旭辉. 浙江舟山定海护城河浮游植物优势种生态位与种间联结性季节性分析. 湖泊科学, 2019, 31(2): 429-439. |
| [16] |
杨文焕, 申涵, 周明利, 李卫平, 张生. 包头南海湖浮游植物优势种生态位及种间联结性季节分析. 中国环境科学, 2020, 40(1): 383-391. |
| [17] |
马一明, 李秋华, 潘少朴, 旷攀, 金爽. 贵州高原花溪水库浮游植物优势种生态位及种间联结性动态分析. 湖泊科学, 2021, 33(3): 785-796. |
| [18] |
陶敏, 岳兴建, 岳珊, 代丽娜, 韩文文, 王永明, 刘果, 李斌. 四川丘陵区水库浮游植物群落结构与蓝藻水华风险——基于优势种生态位与种间联结研究. 生态学报, 2021, 41(23): 9457-9469. |
| [19] |
王孟, 邬红娟, 马经安. 长江流域大型水库富营养化特征及成因分析. 长江流域资源与环境, 2004(5): 477-481. DOI:10.3969/j.issn.1004-8227.2004.05.015 |
| [20] |
郭坤, 彭婷, 罗静波, 杨德国, 何勇凤, 柴毅. 长湖浮游动物群落结构及其与环境因子的关系. 海洋与湖沼, 2017, 48(1): 40-49. |
| [21] |
何勇凤, 李昊成, 王旭歌, 朱永久, 杨德国. 长湖鱼类群落结构的时空变化. 长江流域资源与环境, 2016, 25(2): 265-273. |
| [22] |
何勇凤, 李昊成, 朱永久, 杨德国. 湖北长湖富营养化状况及时空变化(2012—2013年). 湖泊科学, 2015, 27(5): 853-864. |
| [23] |
章宗涉, 黄祥飞. 淡水浮游生物研究方法. 北京: 科学出版社, 1991.
|
| [24] |
胡鸿钧, 魏印心. 中国淡水藻类--系统、分类及生态. 北京: 科学出版社, 2006.
|
| [25] |
邓坚. 中国内陆水域常见藻类图谱. 武汉: 长江出版社, 2012.
|
| [26] |
Levins R. Evolution in changing environments: some theoretical explorations. Princeton University Press, 1968.
|
| [27] |
Pianka E R. The structure of lizard communities. Annual Review of Ecology and Systematics, 1973, 4: 53-74. DOI:10.1146/annurev.es.04.110173.000413 |
| [28] |
Schluter D. A Variance Test for Detecting Species Associations, with Some Example Applications. Ecology, 1984, 65(3): 998-1005. DOI:10.2307/1938071 |
| [29] |
张金屯. 数量生态学. 北京: 科学出版社, 2004.
|
| [30] |
王伯荪, 彭少麟. 南亚热带常绿阔叶林种间联结测定技术研究——Ⅰ.种间联结测式的探讨与修正. 植物生态学与地植物学丛刊, 1985(4): 274-285. |
| [31] |
段后浪, 赵安, 姚忠. 鄱阳湖区典型草洲植物种间关联性研究. 湿地科学, 2017, 15(1): 107-113. |
| [32] |
Bishara A J, Hittner J B. Testing the significance of a correlation with nonnormal data: comparison of Pearson, Spearman, transformation, and resampling approaches. Psychological Methods, 2012, 17(3): 399-417. DOI:10.1037/a0028087 |
| [33] |
苏新然, 于潘, 尤庆敏, 庞婉婷, 王全喜. 三峡库区浮游植物群落结构特征及水生态评价. 湖泊科学, 2023, 35(2): 493-507. |
| [34] |
Wathne J A, Haug T, Lydersen C. Prey preference and niche overlap of ringed seals Phoca hispida and harp seals P. groenlandica in the Barents Sea. Marine Ecology Progress Series, 2000, 194: 233-239. DOI:10.3354/meps194233 |
| [35] |
李滔, 章叶飞, 王海艳, 熊勤学, 张露, 习小杰, 陈如, 刘章勇, 朱建强, 杨军. 湖北长湖蓝藻季节动态及其驱动因子分析. 水生态学杂志, 1-9. DOI:10.15928/j.1674-3075.202204010095 |
| [36] |
夏莹霏, 胡晓东, 徐季雄, 李一平, 吴苏舒, 吴沛沛. 太湖浮游植物功能群季节演替特征及水质评价. 湖泊科学, 2019, 31(1): 134-146. |
| [37] |
Oliver R L, Ganf G G. Freshwater blooms. The ecology of cyanobacteria: their diversity in time and space. Dordrecht: Springer Netherlands, 2000: 149-194.
|
| [38] |
熊莲, 刘冬燕, 王俊莉, 吴明姝, 李东京. 安徽太平湖浮游植物群落结构. 湖泊科学, 2016, 28(5): 1066-1077. |
| [39] |
张亚克, 梁霞, 何池全, 黎彬, 方焰星, 张绚璇, 詹跃武. 淀山湖不同季节营养盐含量与藻类群落的相互关系. 湖泊科学, 2011, 23(5): 747-752. |
| [40] |
庄源益, 赵凡, 戴树桂, 金朝辉. 高等水生植物对藻类生长的克制效应. 环境科学进展, 1995(6): 44-49. |
| [41] |
龚森森, 李学梅, 吴嘉伟, 柴毅, 杨德国, 谭凤霞, 朱挺兵. 湖北长湖夏、秋季浮游生物功能群特征及主要影响因子. 水生生物学报, 2022, 46(5): 707-717. |
| [42] |
李兴, 李建茹, 李畅游. 内蒙古乌梁素海浮游植物优势种的生态位分析. 水生态学杂志, 2017, 38(6): 40-47. |
| [43] |
陈艳, 范明鋆, 李杲光, 李英臣, 于飞, 马剑敏. 氮浓度对小环藻、大型溞和金鱼藻相互作用的影响研究. 环境污染与防治, 2019, 41(12): 1427-1431, 1464. |
| [44] |
福迪. 藻类学. 罗迪安, 译. 上海: 上海科学技术出版社, 1980.
|
| [45] |
刘润红, 陈乐, 涂洪润, 梁士楚, 姜勇, 李月娟, 黄冬柳, 农娟丽. 桂林岩溶石山青冈群落灌木层主要物种生态位与种间联结. 生态学报, 2020, 40(6): 2057-2071. |
| [46] |
王锦旗, 宋玉芝, 薛艳. 气候变化诱导水体富营养化研究进展. 水资源保护, 2022, 38(4): 145-155. |
| [47] |
宋婵媛, 白芳, 李天丽, 宋立荣. 绿藻栅藻对微囊藻的抑制效应及评价. 水生生物学报, 2022, 46(12): 1916-1923. |
| [48] |
魏秘, 朱爱民, 王瑞, 胡菊香. 赤水河春季浮游植物群落结构变化及其优势种生态位分析. 水生态学杂志, 2022, 43(5): 49-58. |
| [49] |
赵永华, 雷瑞德, 何兴元, 贾夏. 秦岭锐齿栎林种群生态位特征研究. 应用生态学报, 2004(6): 913-918. |
| [50] |
龚森森, 吴嘉伟, 柴毅, 罗静波, 谭凤霞, 杨德国, 何勇凤. 长湖浮游动物群落结构特征及其季节变化. 水产科学, 2021, 40(3): 329-338. |
| [51] |
谭凤霞, 罗静波, 祁梅, 柴毅. 长湖水生植物多样性与生态位的季节性变化研究. 长江流域资源与环境, 2021, 30(5): 1121-1129. |
| [52] |
钟军弟, 李先琨, 叶铎, 向悟生, 吕仕洪, 张建亮. 广西木论国家级自然保护区铁榄群落优势种群的生态位研究. 植物资源与环境学报, 2009, 18(3): 38-43. |
| [53] |
肖艳梅, 解婧媛, 姚义鹏, 梁士楚, 高丽娜, 张惠, 莫适祯. 桂林岩溶石山常绿落叶阔叶混交林乔木层优势物种生态位研究. 生态学报, 2021, 41(20): 8159-8170. |
| [54] |
张零念, 朱贵青, 杨宽, 刘星月, 巩合德, 郑丽. 滇中云南杨梅灌丛主要木本植物生态位与种间联结. 植物生态学报, 2022, 46(11): 1400-1410. |
| [55] |
郭忠玲, 马元丹, 郑金萍, 刘万德, 金哲峰. 长白山落叶阔叶混交林的物种多样性、种群空间分布格局及种间关联性研究. 应用生态学报, 2004(11): 2013-2018. |
| [56] |
曹静, 白玉宏, 张钦弟, 毕润成. 山西太岳山接骨木群落优势种种间关联性分析. 生态学杂志, 2015, 34(5): 1313-1319. |
| [57] |
叶深, 杨蕊, 丁朋朋, 彭欣, 刘伟成. 浙江南部近海主要鱼类的时空生态位及种间联结性. 海洋学报, 2022, 44(1): 48-62. |
| [58] |
简敏菲, 刘琪, 朱笃, 游海. 九连山常绿阔叶林乔木优势种群的种间关联性分析. 植物生态学报, 2009, 33(4): 672-680. DOI:10.3773/j.issn.1005-264x.2009.04.005 |
| [59] |
张滋芳, 毕润成, 张钦弟, 许强. 珍稀濒危植物矮牡丹生存群落优势种种间联结性及群落稳定性. 应用与环境生物学报, 2019, 25(2): 291-299. |
| [60] |
张悦, 郭利平, 易雪梅, 曹伟, 王远遐, 吴培莉, 姬兰柱. 长白山北坡3个森林群落主要树种种间联结性. 生态学报, 2015, 35(1): 106-115. |
| [61] |
王三秀, 魏莱, 王爽, 陈玲, 黄清辉. 上海水源地毗邻湖库浮游植物群落结构的季节变化及其影响因子. 湖泊科学, 2022, 34(4): 1127-1139. |
| [62] |
章叶飞, 李滔, 张露, 胡琴, 刘章勇, 杨军. 长湖草/藻型湖区浮游植物群落特征及与环境因子的关系. 水生态学杂志, 1-16. DOI:10.15928/j.1674-3075.202202230048 |
| [63] |
朱丽萍, 孙丹焱, 林晓虎, 黄菊文, 李光明, 徐竟成. 沉水植物化感抑藻效应及作用模式研究进展. 应用化工, 2022, 51(8): 2337-2343. |
| [64] |
高浩杰, 袁佳, 高平仕. 舟山群岛次生林优势种群的生态位与种间联结. 热带亚热带植物学报, 2017, 25(1): 73-80. |
| [65] |
祁梅, 吴嘉伟, 龚森森, 柴毅, 罗静波, 谭凤霞, 杨军, 杨德国, 何勇凤. 长湖浮游植物功能群特征及其与环境因子的关系. 海洋与湖沼, 2020, 51(1): 75-84. |
| [66] |
周先叶, 王伯荪, 李鸣光, 陈章和. 黑石顶自然保护区森林次生演替过程中群落主要种的种间协变分析. 应用生态学报, 2004(3): 367-371. |
| [67] |
刘益鹏, 叶兴状, 叶利奇, 陈绪辉, 郑世群, 陈世品, 张国防, 刘宝. 观光木群落优势树种生态位和种间联结. 应用生态学报, 2022, 33(10): 2670-2678. |
| [68] |
朱利君, 苏智先, 王伟伟, 张亚爽. 卧龙自然保护区三江珙桐群落种间关系的数量分析. 生态学杂志, 2005(10): 1167-1171. |