 2024, Vol. 44
2024, Vol. 44文章信息
- 杨晓龙, 王晓丽, 唐文海, 吴振邦, 王一航, 田阔, 李宏亮, 唐峰, 张秀梅
- YANG Xiaolong, WANG Xiaoli, TANG Wenhai, WU Zhenbang, WANG Yihang, TIAN Kuo, LI Hongliang, TANG Feng, ZHANG Xiumei
- 两种生态型铜藻对不同氮源的吸收特征及生长响应
- The absorption characteristics and growth responses of two ecotypes of sargassum horneri to different nitrogen sources
- 生态学报. 2024, 44(24): 11450-11458
- Acta Ecologica Sinica. 2024, 44(24): 11450-11458
- http://dx.doi.org/10.20103/j.stxb.202403140530
-
文章历史
- 收稿日期: 2024-03-14
- 网络出版日期: 2024-09-09





2. 自然资源部第二海洋研究所, 杭州 310012;
3. 浙江丰宇海洋生物制品有限公司, 舟山 316104
2. Second Institute of Oceanography, Ministry of Natural Resources, Hangzhou 310012, China;
3. Zhejiang Fengyu Marine biological Products Co., LTD, Zhoushan 316104, China
大型海藻是近海生态系统初级生产力的主要贡献者。大型海藻场不仅为海洋生物提供了栖息、繁殖和避难的场所, 同时在缓解气候变化、净化水质、固碳增汇等方面具有重要的生态意义[1]。然而, 随着近年来海岸带城市化进程加速, 人类活动对海洋生态系统的影响日益增加, 特别是由全球海水升温、近岸水体富营养化程度加剧等一系列环境问题引发的藻类生态灾害事件频频发生, 成为影响近海生态环境以及制约海岸带经济可持续发展的重要威胁之一[2]。世界各地关于沿海地区漂浮藻灾害爆发的报道逐年增加, 其中由浒苔大量繁殖形成的“绿潮”[3]和由马尾藻大量繁殖形成的“金潮”[4]是典型的全球性漂浮藻灾害。
铜藻Sargassum horneri隶属于棕色藻门、褐藻纲、墨角藻目、马尾藻科、马尾藻属, 主要生长在低潮带或大干潮线下的礁石上, 是北太平洋沿海特有的暖温性底栖海藻, 在我国辽宁、浙江、福建、广东等地广泛分布[5]。铜藻通常营底栖生活, 因具备气囊等结构, 在固着器或者藻体断裂后会随海流进行长距离的漂移, 形成漂浮藻团。因此, 铜藻的生活史具有定生型和漂浮型两种不同的生态类型[6]。由漂浮铜藻形成的周期性、大规模“金潮”现象在日本、韩国沿海均有报道。近年来, 我国在东海和黄海海域均观测到大面积分布的漂流性铜藻, 并且其爆发频率、规模和致灾程度呈逐年上升趋势。例如, 受2016年底“金潮”灾害影响, 江苏沿海的紫菜养殖业经济损失达5亿元[7]。鉴于漂浮型铜藻所致的“金潮”灾害对近海生态系统及经济发展影响加剧, 深入开展铜藻分布现状、生长过程和爆发机制的研究, 对于防治和预警“金潮”灾害爆发具有重要意义。当前关于我国“金潮”起源的争议较大。一般认为漂浮型铜藻主要来源于定生型铜藻藻体断裂或假根脱落[8], 经风力和洋流作用在海平面汇集形成“金潮”;然而, Li等[9]基于线粒体DNA分析了黄海和东海海域定生型和漂浮型铜藻的种群结构和遗传连通性, 表明定生型和和漂浮型铜藻之间存在极少的基因交流, 我国近海的“金潮”爆发可能并非来源于近海的定生型铜藻。
氮是大型海藻最主要的限制性营养盐之一, 在参与藻体组织构建、细胞酶活、新陈代谢等生命活动中起到不可或缺的作用。铜藻在生长过程中从水体吸收的氮源包括溶解无机氮(DIN)和溶解有机氮(DON)。之前的研究重点关注了铜藻对海水中DIN的吸收和同化过程, 忽视了DON在铜藻生长过程中的作用机制。例如, 孙宏超等[10]开展了铜藻对DIN的吸附动力学特征研究, 表明氨氮作为优先氮源被铜藻吸收;李大鹏等[11]证实了铜藻在营养限制胁迫后对NH4-N的超补偿吸收效应。受陆源排放和海水养殖活动影响, 近岸海水中DON(如尿素)对大型藻潮爆发的影响逐渐引起国内外学者的关注。已有研究表明, 大型海藻对海水中DIN和DON具有不同的吸收途径, 当海水中的DIN含量较低时, DON可以作为大型海藻的替代氮源[12]。近年来我国近岸海域氮素水平的升高和金潮频发及其关联性逐渐引起国内外学者的关注, 但对不同生态型铜藻的氮源吸收动力学特征及其同化机制仍缺乏充分研究。
本研究以我国长江口嵊泗海域的两种不同生态型铜藻为研究对象, 通过室内培养实验比较分析定生型铜藻和漂浮型铜藻对海水中硝态氮(NO3-N)、氨态氮(NH4-N)和尿素氮(Urea-N)的吸收和同化过程, 解析DIN和DON对定生型和漂浮型铜藻生长和消亡的调控机制, 为我国东海长江口“金潮”的起源研究和防治提供科学支撑和数据参考。
1 材料与方法 1.1 实验样品采集及前处理实验用定生型铜藻样品和漂浮型铜藻样品于2023年4月分别采自浙江省嵊泗列岛枸杞岛潮间带及临近海域(30°41′—30°44′N, 122°45′—122°48′E)。采集时选取藻体完整、规格均一的健康鲜活藻体, 现场用海水清洗样品, 确保藻体无泥沙、附着藻类和动物等杂质。将样品低温(4℃)运输至浙江海洋大学海洋牧场与渔业碳汇生态功能创新实验室开展后续实验。
实验室内, 在恒温光照培养箱(GXZ-380B)开展两种生态型铜藻样品的氮饥饿培养实验。选取健康饱满、株高约为40 cm的铜藻植株, 此时铜藻处于快速生长期, 对氮源的吸收速率较高, 便于开展实验研究。用灭菌海水清洗, 并用1%的次氯酸钠溶液清洗1—2 min, 再用灭菌海水冲洗干净。选取藻体中段部位作为实验样本, 将两种生态型的铜藻样本按2 g/L的密度置于无氮源添加的改良型f/2培养溶液中暂养, 培养条件为温度(16±0.5)℃, 盐度(28.3±0.6)‰(参照样品采集时现场海水的温度和盐度), 光照强度5000 lux, 光照周期12 L: 12 D。为避免藻体携带的硅藻对实验结果的影响, 实验过程中在培养基中添加l mL/L的GeO2抑制硅藻生长。每天向培养基中补充除氮源以外的其他营养成分。氮饥饿实验连续培养48 h, 使铜藻藻体处于充分氮饥饿状态。
1.2 不同生态型铜藻对三种氮源的吸收动力学特征选取硝酸钠(NaNO3)、氯化铵(NH4Cl)和尿素(CO(NH2)2)三种氮源开展两种生态型铜藻的吸收动力学特征研究。根据预实验结果, 将三种氮源实验组分别设置6个浓度梯度, 每种氮源的f/2培养溶液中氮初始浓度分别为10 μmol/L、20 μmol/L、40 μmol/L、80 μmol/L、120 μmol/L、240 μmol/L。在不同处理条件中, 准确称取6.0 g相同部位的铜藻置于装有2 L培养液的培养瓶中进行培养, 另设置不添加铜藻样品的培养液作为对照组。培养条件参照1.1, 每组设置5个平行。在培养的第0 min、10 min、30 min、1 h、2 h、5 h、7 h、10 h、15 h和24 h测定培养液的氮浓度。采用锌-镉还原法、次溴酸盐氧化法和二乙酰一肟法分别测定培养液中硝态氮、氨态氮和尿素态氮的含量(μmol/L)。培养实验结束后, 准确称取藻体湿重(g)。
铜藻对不同氮源的吸收速率公式为:
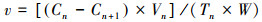
|
式中, v为吸收速率, Cn和Cn+1分别为第n次间隔取样的培养液N起始浓度和终末浓度(μmol/L), Vn为间隔取样时的培养液体积, Tn为每次取样的间隔时间(h), W为铜藻湿重(g)。
采用米氏动力学方程计算不同生态型铜藻对氮源的最大吸收速率(Vmax)和半饱合常数(Km):
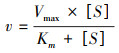
|
式中, Vmax为最大吸收速率(μmol L-1 h-1), Km为半饱合常数(μmol/L), [S]为每次间隔取样的培养液N起始浓度(μmol/L)。
1.3 不同氮源对两种生态型铜藻的生长影响根据铜藻的吸收动力学特征实验结果, 选取NH4-N、NO3-N和Urea-N作为不同N源, 比较不同氮源对两种生态型铜藻生长及其氮素同化效率的影响。在开展铜藻对氮源吸收动力学特征的预实验结果表明, 培养介质中氮源浓度为80 μmol/L时, 铜藻对氮素的吸收速率达到饱和, 氮素浓度不会影响铜藻对氮源的吸收速率。因此, 本研究将培养介质中的总氮浓度设置为80 μmol/L, 实验对照组中培养介质无氮添加, 实验周期为15天, 每个实验组设置3组平行, 其他实验条件参照1.2。分别在实验培养的第1、2、3、5、7、9、11、13天和15天称量藻体鲜重, 计算铜藻生长随培养时间的变化, 铜藻的生长采用相对生长速率表征, 计算公式如下:
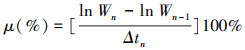
|
式中, SGR为铜藻相对生长率, Wn和Wn-1分别为培养时间段内前后铜藻的鲜重, Δtn为第n次采样的培养时间间隔。
在测定铜藻相对生长速率的同时, 于各阶段采集2.5 mL的培养液测定培养介质中三种氮源的浓度变化, 计算氮源在培养时间间隔Δtn的浓度变化率, 分别以δNH4-N、δNO3-N和δUrea-N表示。
培养实验结束后, 将铜藻烘干, 采用FlashSmart元素分析仪测定藻体的总N含量(mg/g)。
1.4 统计分析统计数据以平均值±标准误差(mean±SE)表示, 基于Excel 2010、SPSS 19.0及R软件4.2.1对数据进行统计分析并绘制图表。采用单因素方差和Duncan′s多重比较法分析不同处理组间铜藻的吸收动力学参数和铜藻藻体组织中总氮含量的差异显著性。根据Shapiro-Wilk检验判断铜藻相对生长速率和培养介质中氮源浓度变化率的数据分布正态性, 使用Pearson相关系数分析二者的相关性。上述分析均设置显著水平为P < 0.05。
2 结果与分析 2.1 不同生态型铜藻对三种氮源的吸收动力学特征在铜藻藻体培养的前2 h, NH4-N、NO3-N和Urea-N的浓度随培养时间呈线性下降, 2 h后不同氮源的浓度与培养时间呈非线性关系, 表明0—2 h的铜藻对培养介质中的氮吸收速率不变, 而2 h后铜藻对培养介质中的氮吸收速率随时间发生变化。因此, 本研究选取0—2 h作为研究两种生态型铜藻对三种氮源的吸收动力学特征时间程。
定生型和漂浮型铜藻对NH4-N的吸收动力学特征曲线见图 1。两种不同生态型的铜藻对NH4-N的吸收速率随培养介质中NH4-N的浓度升高而增加。当NH4-N浓度达到80 μmol/L时, 藻体对NH4-N的吸收速率趋于平缓。单因素方差分析结果表明, 当培养介质中NH4-N的浓度低于20 μmol/L时, 两种生态型的铜藻对NH4-N的吸收速率差异不显著(P=0.79);当培养介质中NH4-N的浓度高于40 μmol/L时, 定生型铜藻对NH4-N的吸收速率显著高于漂浮型铜藻(P < 0.05)。

|
| 图 1 不同生态型铜藻对氨态氮(NH4-N)、硝态氮(NO3-N)和尿素态氮(Urea-N)的吸收速率随氮源浓度的变化曲线 Fig. 1 Curves of NH4-N, NO3-N, and Urea-N uptake rates of different ecotypes of Sargassum horneri as a function of nitrogen source concentration |
两种不同生态型的铜藻对NO3-N的吸收动力学特征曲线见图 1。定生型和漂浮型铜藻对NO3-N的吸收速率随培养介质中NO3-N的浓度升高呈现上升趋势, 并在NO3-N浓度大于80 μmol/L时吸收速率逐渐达到平稳。两种生态型的铜藻对NO3-N的吸收速率在不同NO3-N浓度下均差异不显著(P>0.05)。
定生型和漂浮型铜藻对Urea-N的吸收动力学特征曲线与NH4-N的吸收曲线一致。随着培养介质中Urea-N的浓度升高, 两种生态型的铜藻对Urea-N的吸收速率增加并最终趋于平稳。当Urea-N浓度低于40 μmol/L时, 二者的吸收速率差异不显著(P>0.05);Urea-N浓度高于80 μmol/L时, 漂浮型铜藻的吸收速率显著高于定生型(P < 0.05)。
根据米氏方程计算获得不同生态型铜藻在三种氮源的最大吸收速率(Vmax)、半饱合常数(Km)及其比值(表 1)。不同氮源培养条件下, 铜藻的最大吸收速率和半饱合常数具有差异。其中, 铜藻对NH4-N和Urea-N的最大吸收速率差异不显著, 但显著高于NO3-N;铜藻对NO3-N的半饱合常数显著高于其他两种氮源。相同氮源培养条件下, 不同生态型铜藻的最大吸收速率和半饱合常数表现出较大差异。其中, 定生型铜藻对NH4-N的最大吸收速率和半饱合常数均显著高于漂浮型铜藻, 但漂浮型铜藻对Urea-N的最大吸收速率显著高于定生型铜藻;两种生态型铜藻对NO3-N的吸收动力学参数不具有差异显著性。定生型铜藻在NH4-N培养介质中的Vmax/Km显著大于漂浮型, 在NO3-N和Urea-N中不具有显著差异。
| 铜藻生态型
Ecotype of Sargassum horneri |
氮源形式
Form of nitrogen source |
Vmax/ (μmol L-1 h-1) |
Km/ (μmol/L) |
Vmax/Km | R2 |
| 定生型Stationary type | NH4-N | 39.86±4.15a | 34.25±3.99a | 1.16±0.12 a | 0.98 |
| 漂浮型Floating type | 32.28±3.54b | 26.23±5.28b | 1.25±0.20 b | 0.99 | |
| 定生型Stationary type | NO3-N | 29.97±2.79a | 43.38±4.16a | 0.72±0.11 a | 0.96 |
| 漂浮型Floating type | 31.49±3.69a | 41.66±9.47a | 0.74±0.06 a | 0.96 | |
| 定生型Stationary type | Urea-N | 34.54±5.14a | 31.33±8.24a | 1.12±0.11 a | 0.98 |
| 漂浮型Floating type | 38.25±4.78b | 34.88±5.23a | 1.06±0.14 a | 0.99 | |
| Vmax:最大吸收速率Maximum uptake rate;Km:半饱和常数Half-saturation constant; 不同小写字母表示不同生态型铜藻在相同氮源下具有显著差异(P < 0.05) | |||||
定生型和漂浮型铜藻在不同氮源下随培养时间的相对生长速率(μ)变化曲线如图 2。在NH4-N培养条件下, 两种生态型铜藻的相对生长速率较低, 并且随着培养时间的增加均呈现下降趋势。漂浮型铜藻在第9天开始腐烂, 并随着培养时间的增加腐烂程度加速。在NO3-N培养条件下, 两种生态型的铜藻藻体相对生长速率均呈现先上升后下降的趋势;漂浮型铜藻的相对生长速率高于定生型铜藻, 两种生态型铜藻在培养期间均未发现腐烂现象。在Urea-N作为氮源的培养条件下, 定生型铜藻的相对生长速率呈现先上升后下降的规律, 在培养第3天达到峰值((5.6± 0.15) g/d), 从第5天到第11天相对生长速率急剧下降, 此后处于平稳期。漂浮型铜藻在培养第1天的相对生长速率显著高于定生型铜藻(P < 0.05), 其后表现出持续下降的趋势, 藻体从第11天开始逐渐腐烂, 其后相对生长速率表现为负值。对照组藻体生物量除前2天有短暂增加外, 其后均表现为负增长。培养介质中NH4-N浓度变化速率(δNH4-N)和Urea-N浓度变化速率(δUrea-N)与铜藻相对生长速率均呈显著正相关(P < 0.05), NO3-N浓度变化速率(δNO3-N)同铜藻相对生长速率相关性不显著(图 3)。

|
| 图 2 不同氮源下两种生态型铜藻相对生长率随培养时间的变化 Fig. 2 Changes of relative growth rates of two ecotypes of Sargassum horneri with different nitrogen sources and culture time |

|
| 图 3 培养介质中三种氮源的浓度变化速率(δ) 与铜藻相对生长速率的相关性 Fig. 3 Correlation between the rate of concentration change of three nitrogen sources in culture medium and the relative growth rates of Sargassum horneri |
在NO3-N氮源介质中培养的定生型铜藻组织中的总氮含量显著升高, 而NH4-N和Urea-N氮源介质中培养的定生型铜藻组织中总氮含量未发生显著变化(图 4)。漂浮型铜藻经三种不同氮源介质培养后, 除NH4-N氮源介质中的藻体组织总氮含量未发生变化外, 其他两种氮源培养的铜藻组织中总氮含量均显著增加(P < 0.05), 在Urea-N氮源介质中生长的漂浮型铜藻总氮含量略高于在NO3-N氮源介质中生长的铜藻。

|
| 图 4 定生型铜藻和漂浮型铜藻在不同氮源介质培养前后藻体组织中总氮含量变化 Fig. 4 Changes of total nitrogen content in the tissues of stationary type and floating type of Sargassum horneri before and after culture in medium for different nitrogen sources |
开展大型藻类营养吸收动力学特征研究, 对解析大型藻类灾害爆发机制和调控大型藻类增养殖过程具有重要的生态学意义[13—14]。国内外学者前期对不同类型大型海藻的营养吸收动力学开展了较多研究[15—17], 并从形态学、生理学和基因调控等方面研究了大型海藻对氮磷等营养元素的吸收效率及其响应特征[18—20]。已有研究表明, 大型海藻对水体可溶性氮素表现出饱和吸收动力学的特征, 其对氮源的吸收速率和水体中相应的可溶性氮浓度符合米式动力学方程。米氏动力学参数Vmax表示底物浓度饱和时藻类的吸收速率, 在相同环境条件下, Vmax值越大表明藻类吸收氮素的潜力越大;Km表示当大型藻类的吸收速率为最大吸收速率一半时的底物浓度, Km值越大, 表明藻类对底物的亲和力越小[21]。但也有研究认为, Vmax/Km是表征大型藻类离子亲和力的优先指标, 因为Km值的计算主要依赖于Vmax[22—23]。
本研究基于米氏动力学方程比较分析了定生型和漂浮型两种生态型铜藻在NH4-N、NO3-N和Urea-N三种唯一氮源培养液中的氮素吸收动力学特征。结果表明, 铜藻对Urea-N与NH4-N的Km值和Vmax/Km差异不显著, 表明Urea-N和NH4-N对铜藻载体位点的亲和力相近, 而铜藻对NO3-N的Km值显著大于其他两种氮源, 表明铜藻对Urea-N和NH4-N的亲和力强于NO3-N, 这与大多数海藻的氮素吸收特征相似。孙宏超等[10]通过吸收实验初步分析了铜藻对无机磷、硅酸盐、亚硝酸盐和氨氮的吸收效率随时间的变化情况, 表明氨氮作为优先氮源被铜藻吸收, 上述结论同本研究的结果相一致。这是由于海藻对不同形态氮源的传递过程和同化机制不同所致[24]。在藻体吸收不同氮素后, NH4-N能够直接与藻体细胞的氨基酸结合, 有机氮源Urea-N被藻体吸收后, 可以被藻体内部的微生物快速分解释放出NH4-N被藻类同化利用[25], 而NO3-N需被硝酸还原酶还原成NH4-N后才能进行下一步同化, 该过程需要消耗大量的ATP和磷, 增加磷限制风险, 不利于海藻生长[26]。但也有研究表明, 在NH4-N与NO3-N共存时, 长茎葡萄蕨藻Caulerpa lentillifera更倾向于优先利用NO3-N, 表明大型海藻的营养吸收动力学特征受海藻种类、生长状态以及环境因素等影响[27]。
定生型铜藻对NH4-N的Vmax和Km均显著大于漂浮型铜藻, 表明定生型铜藻对NH4-N的吸收潜力较大, 但亲和力较低;而漂浮型铜藻对Urea-N的Vmax显著大于定生型铜藻, 表明漂浮型铜藻对Urea-N具有较强的吸收潜力。漂浮型铜藻通常分布在海面上, 随洋流营集群漂流生活或缠绕在贻贝等养殖筏架上, 而定生型铜藻主要分布于潮下带生境利用假根营固着生活。二者的生境环境存在较大差异, 可能导致两种生态型铜藻之间的基因交流发生阻隔, 造成生理和基因层面的差异[9]。已有研究表明, 为了适应海面生存环境, 漂浮型铜藻进化出一系列的适应机制, 从形态、超微结构、光合生理、生长速率以及电子传递速率等均异于定生型铜藻[6, 28—29], 具有更强的生长优势和光保护能力。本研究中, 随着培养液中NH4-N和Urea-N消耗殆尽, 漂浮型铜藻藻体开始腐烂, 而NO3-N培养液中的两种生态型铜藻均未发生腐烂现象, 表明NO3-N在漂浮型铜藻后期的存活和生长过程中起到积极作用。NO3-N组的藻体总氮含量显著高于同浓度的NH4-N和Urea-N组, 表明铜藻对NO3-N具有更强的储存能力。Gordillo等[30]发现大型海藻能够将吸收后未被转化利用的NO3-N储存于液泡中, 为海藻细胞后期生长时提供所需的大量N源, 这也解释了NO3-N组铜藻在培养后期仍能保持正向生长的原因。
有研究证实, 某些赤潮藻种和大型藻类能够优先利用Urea-N作为高效生长氮源, 如卡伦氏贝氏藻Karenia brevis对Urea-N的最大吸收速率显著高于NH4-N和NO3-N, 且同化为蛋白质的比例以Urea-N最高[31];微小原甲藻Prorocentrum minimum也会优先选择Urea-N作为氮源[32];Li等[26]结合室内培养实验和现场调查研究表明, 海水中有机氮(Urea-N和甘氨酸)含量增加是我国黄海南部海域绿潮爆发的可能原因之一。在本研究中, Urea-N组的漂浮型铜藻具有较高的初始相对生长速率, 其藻体组织中总氮含量显著提升, 而定生型铜藻未发生显著变化, 表明漂浮型铜藻对Urea-N具有较强的吸收和同化能力。尿素作为重要的氮肥和工业原料, 广泛运用在工农业上。研究表明, 浙江近岸海域的金潮主要在每年4月下旬爆发, 大面积的铜藻在浙江沿海集结并向北漂移至南黄海, 在5月下旬逐渐消亡[33]。根据王雪景等[34]和苏莹等[35]对东海长江口Urea-N水平及分布特征的调查结果表明, 长江口海域的Urea-N浓度在4月底出现最高值, 该时期也是东海赤潮和金潮的爆发期。赤潮的爆发会消耗大量无机氮, 导致长江口海域无机氮浓度下降, 推测水体中高浓度的Urea-N是维持金潮爆发的重要氮源。结合近年来沿海水域Urea-N浓度不断上升的现状, Urea-N对浮游植物和大型海藻的生长、消亡及群落演替等生态过程的作用和影响不容忽视。本研究结果表明, 漂浮型铜藻对Urea-N具有高效的同化作用, 客观反映了Urea-N对我国东海金潮频发的影响现状。因此, 深入探究不同生态型铜藻对氮源的利用和同化过程, 有助于解析我国近海藻类灾害爆发的形成机制。
4 结论研究重点关注了定生型铜藻和漂浮型铜藻对三种氮源(NH4-N、NO3-N和Urea-N)的吸收动力学特征及其同化作用研究, 解析了三种氮源对两种不同生态型铜藻生长过程的调控作用及其影响。定生型铜藻对NH4-N的吸收潜力较大, 但其亲和力较低;两种生态型铜藻虽然对NO3-N的吸收潜力和亲和力均较低, 但饱和浓度的NO3-N可以提升漂浮型铜藻的后期生长潜力, 而漂浮型铜藻对Urea-N的利用效率显著高于定生型铜藻, 表明海水中NO3-N和Urea-N的耦合作用可能是导致“金潮”爆发和持久性的主要因素之一。本研究明确了不同生态型铜藻对氮素的吸收特征和同化过程, 为我国“金潮”的防治和管理提供了理论基础。
| [1] |
章守宇, 刘书荣, 周曦杰, 汪振华, 王凯. 大型海藻生境的生态功能及其在海洋牧场应用中的探讨. 水产学报, 2019, 43(9): 2004-2014. |
| [2] |
于仁成, 吕颂辉, 齐雨藻, 周名江. 中国近海有害藻华研究现状与展望. 海洋与湖沼, 2020, 51(4): 768-788. |
| [3] |
王宗灵, 傅明珠, 肖洁, 张学雷, 宋伟. 黄海浒苔绿潮研究进展. 海洋学报, 2018, 40(2): 1-13. |
| [4] |
王丹, 姜亦飞, 王先桥, 王素芬, 何恩业, 张蕴斐. 我国马尾藻金潮生态动力学研究进展. 地球科学进展, 2021, 36(7): 753-762. |
| [5] |
毕远新, 章守宇, 王伟定, 吴祖立. 枸杞岛铜藻垂直分布格局及成因分析. 生态学报, 2014, 34(17): 4931-4937. DOI:10.5846/stxb201301030013 |
| [6] |
刘正一, 孙平, 秦松, 李晶晶, 庄龙传, 宋宛霖, 毕远新, 钟志海. 底栖铜藻和漂浮铜藻生长与光合生理的比较. 生态学杂志, 2021, 40(1): 76-83. |
| [7] |
邓邦平, 刘材材, 徐韧, 张昊飞, 张正龙. 南黄海绿潮对江苏海水养殖的影响及防控对策. 海洋开发与管理, 2015, 32(5): 103-106. DOI:10.3969/j.issn.1005-9857.2015.05.023 |
| [8] |
Yoshida T, Sawada T, Higaki M. Sargassum vegetation growing in the sea around Tsuyazaki, North Kyushu, Japan. Pacific Science, 1963, 17: 135-144. |
| [9] |
Li J J, Liu Z Y, Zhong Z H, Zhuang L C, Bi Y X, Qin S. Limited genetic connectivity among Sargassum horneri (Phaeophyceae) populations in the Chinese marginal seas despite their high dispersal capacity. Journal of Phycology, 2020, 56(4): 994-1005. DOI:10.1111/jpy.12990 |
| [10] |
孙宏超, 章守宇. 铜藻对几种无机盐的吸收动力学的初步研究. 上海水产大学学报, 2007, 16(6): 573-579. |
| [11] |
李大鹏, 马增岭, 李慧, 丁刚, 辛美丽, 吴海一, 郭文. 铜藻(Sargassum horneri)在营养限制胁迫后对NH4-N的超补偿吸收研究. 海洋与湖沼, 2018, 49(4): 904-909. |
| [12] |
Li H M, Zhang Y Y, Han X R, Shi X Y, Rivkin R B, Legendre L. Growth responses of Ulva prolifera to inorganic and organic nutrients: implications for macroalgal blooms in the southern Yellow Sea, China. Scientific Reports, 2016, 6: 26498. DOI:10.1038/srep26498 |
| [13] |
Pedersen M F, Borum J. Nutrient control of estuarine macroalgae: growth strategy and the balance between nitrogen requirements and uptake. Marine Ecology Progress Series, 1997, 161: 155-163. DOI:10.3354/meps161155 |
| [14] |
Thornber C S, DiMilla P, Nixon S W, McKinney R A. Natural and anthropogenic nitrogen uptake by bloom-forming macroalgae. Marine Pollution Bulletin, 2008, 56(2): 261-269. DOI:10.1016/j.marpolbul.2007.10.031 |
| [15] |
刘棋琴, 羊芃, 马明婕, 周丽亚, 李亚鹤, 杨锐. 温度对4种大型海藻氮磷吸收效率及光合生理特性的影响. 水生生物学报, 2018, 42(5): 1050-1056. |
| [16] |
Pérez-Mayorga D M, Ladah L, Zertuche-González J A, Leichter J, Filonov A, Lavín M. Nitrogen uptake and growth by the opportunistic macroalga Ulva lactuca (Linnaeus) during the internal tide. Journal of Experimental Marine Biology and Ecology, 2011, 406: 108-115. DOI:10.1016/j.jembe.2011.05.028 |
| [17] |
Perini V, Bracken M E S. Nitrogen availability limits phosphorus uptake in an intertidal macroalga. Oecologia, 2014, 175(2): 667-676. DOI:10.1007/s00442-014-2914-x |
| [18] |
韩婷婷, 齐占会, 黄洪辉, 梁庆洋. 3种氮源加富对半叶马尾藻幼苗生长和生化组成的影响. 渔业科学进展, 2018, 39(4): 159-166. |
| [19] |
Hurd C L, Harrison P J, Druehl L D. Effect of seawater velocity on inorganic nitrogen uptake by morphologically distinct forms of Macrocystis integrifolia from wave-sheltered and exposed sites. Marine Biology, 1996, 126(2): 205-214. DOI:10.1007/BF00347445 |
| [20] |
Liu X J, Wen J Y, Chen W Z, Du H. Physiological effects of nitrogen deficiency and recovery on the macroalga Gracilariopsis lemaneiformis (Rhodophyta). Journal of Phycology, 2019, 55(4): 830-839. DOI:10.1111/jpy.12862 |
| [21] |
刘静雯, 董双林. 海藻的营养代谢及其对主要营养盐的吸收动力学. 植物生理学通讯, 2001, 37(4): 325-330. |
| [22] |
Jaime B, Miguel S J, Dolores B M, Aranzazu R, Rocío G, Lazaro M, Manuel R J. The role of Cymodocea nodosa and Caulerpa prolifera meadows as nitrogen sinks in temperate coastal lagoons. Diversity, 2023, 15(2): 172. DOI:10.3390/d15020172 |
| [23] |
Narvarte B C V, Genovia T G T, Hinaloc L A R, Roleda M Y. Growth, nitrate uptake kinetics, and biofiltration potential of eucheumatoids with different thallus morphologies. Journal of Phycology, 2022, 58(1): 12-21. DOI:10.1111/jpy.13229 |
| [24] |
Hurd C L, Harrison P J, Bischof K, Lobban C S. Seaweed ecology and physiology. Cambridge, UK: Cambridge University Press, 2014.
|
| [25] |
Tamminen T, Irmisch A. Urea uptake kinetics of a midsummer planktonic community on the SW coast of Finland. Marine Ecology Progress Series, 1996, 130: 201-211. DOI:10.3354/meps130201 |
| [26] |
Li H M, Zhang Y Y, Han X R, Shi X Y, Rivkin R B, Legendre L. Growth responses of Ulva prolifera to inorganic and organic nutrients: implications for macroalgal blooms in the southern Yellow Sea, China. Scientific Reports, 2016, 6: 26498. DOI:10.1038/srep26498 |
| [27] |
Liu H T, Wang F, Wang Q H, Dong S L, Tian X L. A comparative study of the nutrient uptake and growth capacities of seaweeds Caulerpa lentillifera and Gracilaria lichenoides. Journal of Applied Phycology, 2016, 28(5): 3083-3089. DOI:10.1007/s10811-016-0858-8 |
| [28] |
张恒, 赵倩, 陈娟娟, 朱旗, 吴小凯, 骆其君, 杨锐, 陈海敏. 漂浮型和定生型铜藻中色素的液质联用分析. 核农学报, 2019, 33(6): 1173-1180. |
| [29] |
朱旗, 任继锐, 陈娟娟, 杨锐. 定生型和漂浮型铜藻的形态及超微结构观察. 生物学杂志, 2019, 36(3): 51-54. |
| [30] |
Gordillo F, Dring M J, Savidge G. Nitrate and phosphate uptake characteristics of three species of brown algae cultured at low salinity. Marine Ecology Progress Series, 2002, 234: 111-118. DOI:10.3354/meps234111 |
| [31] |
Sinclair G, Kamykowski D, Glibert P M. Growth, uptake, and assimilation of ammonium, nitrate, and urea, by three strains of Karenia brevis grown under low light. Harmful Algae, 2009, 8(5): 770-780. DOI:10.1016/j.hal.2009.02.006 |
| [32] |
Fan C, Glibert P M, Alexander J, Lomas M W. Characterization of urease activity in three marine phytoplankton species, Aureococcus anophagefferens, Prorocentrum minimum, and Thalassiosira weissflogii. Marine Biology, 2003, 142(5): 949-958. DOI:10.1007/s00227-003-1017-8 |
| [33] |
丁晓玮, 张建恒, 庄旻敏, 康新宇, 赵晓惠, 何培民, 刘书荣, 刘建芬, 文源, 沈辉, 钟佳明. 我国长江口及邻近海域铜藻生长和金潮分布变化特征. 海洋渔业, 2019, 41(2): 188-196. |
| [34] |
王雪景, 石晓勇, 赵玉庭, 张传松. 2011年春季东海赤潮高发区尿素分布特征及影响因素. 海洋学报, 2015, 37(2): 65-74. DOI:10.3969/j.issn.0253-4193.2015.02.007 |
| [35] |
苏莹, 姜聪聪, 石晓勇, 赵玉庭, 梁生康, 王修林. 春季东海赤潮高发区尿素的断面分布及影响因素. 海洋环境科学, 2015, 34(2): 171-175, 189. |