 2024, Vol. 44
2024, Vol. 44文章信息
- 谭凯, 温仲明, 郑诚, 袁浏欢, 吴沛桐, 王志鹏, 石长春, 马雅莉, 张艳
- TAN Kai, WEN Zhongming, ZHENG Cheng, YUAN Liuhuan, WU Peitong, WANG Zhipeng, SHI Changchun, MA Yali, ZHANG Yan
- 毛乌素沙地飞播植被演替植物功能型组成和土壤理化性质变化特征
- The variation characteristics of plant functional group composition and soil physical and chemical properties during the succession of aerial seeding vegetation in Mu Us Sandy Land
- 生态学报. 2024, 44(23): 10713-10723
- Acta Ecologica Sinica. 2024, 44(23): 10713-10723
- http://dx.doi.org/10.20103/j.stxb.202405051002
-
文章历史
- 收稿日期: 2024-05-05
- 网络出版日期: 2024-10-07

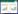




2. 西北农林科技大学水土保持研究所, 杨凌 712100;
3. 陕西省林业科学院, 西安 710028
2. Research Center of Soil and Water Conservation and Ecological Environment, Chinese Academy of Sciences and Ministry of Education, Yangling 712100, China;
3. Shaanxi Academy of Forestry Sciences, Xi'an 710028, China
毛乌素沙地位于黄土高原西北部, 长期以来, 由于自然干旱和人为因素的强烈干扰, 这里一直是中国土地荒漠化和沙化最严重的地区之一[1]。为抵御风沙侵害, 我国开展了大量的飞播造林等生态恢复工作, 毛乌素沙区人工植被面积逐年增加, 沙区植被盖度也由原来的不足10%增加至现在的20%—50%, 部分地区甚至超过50%[2]。围绕毛乌素沙地植被恢复, 众多学者对其开展了大量研究, 如毛乌素沙地飞播造林植被恢复特征[3]、土壤性质及酶活性变化[3—6]、土壤水分变化[7—8]、毛乌素沙地飞播造林不同恢复阶段植物群落物种多样性[9—10]和沙地植被恢复的固沙固碳效益[7, 11]等, 这些研究为理解毛乌素沙地植被恢复过程及其环境效应提供了重要依据, 但关于植被恢复演替过程中物种组成变化及其与土壤水分相互作用的研究仍为缺乏, 沙地飞播恢复成效的全面评估仍需开展深入的研究工作。
近年来, 一些研究发现, 毛乌素沙地植被恢复过程中水资源与植被间的供需矛盾加剧, 出现了土壤水分亏缺和植被群落退化现象[12—13], 加之土壤干燥、初始飞播植被进入生长衰败期等原因, 已治理的毛乌素沙地面临二次沙化风险[14]。那么当飞播植被开始衰退后, 群落组成结构如何发生改变?自然植被是否介入?群落演替方向是否具有某种确定性趋势?土壤水分究竟是被过度消耗还是恢复?这些问题都尚未解决, 而这些问题的解决需要对毛乌素沙地飞播植被的长时间序列演替过程进行系统的调查与分析。
为了深入了解植被演替过程中植物与环境变化的响应过程, 本研究在传统的物种组成调查的基础上, 引入植物功能性状的相关研究方法。植物功能性状是指与植物的发生、存活、生长和死亡过程高度相关的内在生理和外在形态特征[15]。植物个体具备调控自身特性的潜能, 借此以缓冲外在环境的压迫, 增强其环境适应性。演替环境在持续的筛选过程中, 塑造了植物多元化的生长、繁衍及自我防护机制等应对策略[16]。具有相似响应策略的同类型植物, 即植物功能型(Plant Functional Types, PFTs), 通常具有一套相似植物功能性状组合[17], 对同一环境变化产生相同响应, 对主要生态系统过程有相似作用[18]。以植物功能型为基础的方法侧重于研究许多不同亲缘关系的物种对同一生态环境压力的适应方式[19], 从而在一定程度上降低了群落复杂性。但在植物功能型分类时, 不同的研究人员会采取不同的分类方法、依据不同的分类角度进行划分, 诸如生长型(乔木、灌木、草本等)[20—21];分配对策(水分保持[22]、养分保持和繁殖等);代谢特征(阳性与荫性[23], C3与C4[24]和是否固氮[25]);生长速度(相对生长速度、养分吸收速度、光合速度[23]);种子大小、扩散方式、繁殖类型等, 但这些划分方法多为主观分类方法或演绎分类方法。对同一功能型内植物的性状差异往往无法得到充分反映。为此, 近年来, 一些研究开始基于植物功能性状的数量特征进行植物功能型划分(即数量分类方法[26]), 例如Diaz Barradas等[27]认为植物功能型应使用易于测量的植物性状进行分类, 这样更容易评估生态系统功能。因此, 本研究以毛乌素沙地4个不同年限飞播区的植物群落为研究对象, 采用群落加权平均性状值的方法建立植物性状与土壤环境的关系, 并基于植物性状的功能型划分方法和逐步回归模型, 探究毛乌素沙地飞播植被在不同演替时期的适应策略。旨在回答以下问题:(1)飞播植被演替过程中功能型组成有何变化, 不同功能型植物的适应策略有何不同;(2)群落演替方向是否具有某种确定性趋势;(3)飞播演替过程中土壤水分是否得到恢复。
1 材料与方法 1.1 研究区概况研究区位于陕西省榆林市榆阳区红石峡播区(38°20′50″N, 109°38′20″E), 平均海拔约1200m, 地处毛乌素沙地的东南边缘(图 1)。该区属温带半干旱大陆性季风气候, 春季多风、夏季干燥﹑秋季多雨、冬季寒冷, 日照充足。雨季较晚, 年降雨量变化较大, 年均气温8.3℃, 年均日照2743.3h, 年均降水量400mm左右, 主要集中在7—9月份。该地区植被稀疏, 地表覆盖物多为质地疏松的沙质土壤, 风沙活动强烈。植被以沙生草灌群落为主, 包括花棒(Corethrodendron scoparium)、柠条(Caragana korshinskii)、狗尾草(Setaria viridis)、阿尔泰狗娃花(Aster altaicus)、雾冰藜(Grubovia dasyphylla)、沙蒿(Artemisia desertorum)、青蒿(Artemisia caruifolia)、踏郎(Hedysarum fruticosum)、沙蓬(Agriophyllum squarrosum)、沙打旺(Astragalus laxmannii)。该区自1965年开始飞播灌草种实施生态恢复, 前后开展了4期飞播试验(1965年、1975年、1983年和2000年), 为研究提供了条件。

|
| 图 1 研究区分布图与采样点信息 Fig. 1 Distribution map of study area and information of sampling point |
采用时空置换的方法, 选取沙区不同飞播年限的草地和灌木地(表 1), 以长势良好的植物群落为研究对象, 研究飞播植被演替过程中植物功能型组成及植物性状的变化特征。为保证研究结果的可靠性, 本研究涉及的不同飞播年限的样地空间上相互靠近, 同一样点的气候条件相似;且每期飞播试验前均为无植被存在的沙地, 因此不同样点间初始土壤理化性质并无差异。每个演替序列都采用了多个重复, 以增加样本数量, 从而减少因主观因素而造成的误差。本研究于2023年8月份进行调查, 共选择了16个样地, 通过GPS采集了样地的经纬度、海拔、坡度和坡向, 详细记录了样地的基本环境和物种。根据研究区植被初始飞播年份, 演替时间序列大致分为20a、40a、50a和60a。通过查阅资料来确认各样地具体的飞播情况, 确保各样地的演替起点都是从实施飞播开始的。每个飞播年限选择3至5个样点采用典型样方法进行群落调查, 灌木和草本的样方大小分别为5m×5m和1m×1m, 每个样点重复5次。
| 飞播年份 Years of aerial seeding plot |
县区 County/district |
播区名称 Name |
作业面积 Operating area/hm2 |
带长 Belt length/m |
方位 Direction |
飞播物种 Species of aerial seeding plant |
| 1965 | 榆阳区 | 芹河东沙 | 70 | 2680 | 东南-西北(295°) | 花棒、踏郎、沙蒿 |
| 1975 | 榆阳区 | 白山海则 | 364 | 3470 | 东南-西北(340°) | 花棒、踏郎、沙蒿 |
| 1983 | 榆阳区 | 孟家湾东沙 | 422 | 4000 | 东南-西北(333°) | 花棒、踏郎、沙蒿 |
| 2000 | 榆阳区 | 孟家湾西沙 | 1320 | 5500 | 西南-东北(190°) | 花棒、踏郎、沙蒿 |
每个物种选择3株成熟、生长良好、大小一致的植株进行调查, 以获得平均株高。每个优势物种采集10到15片健康无病虫害且完全伸展的叶片, 装入写有编号的塑封袋防止叶片水分散失, 从而影响叶干物质含量数据的结果。叶片带回实验室后利用电子游标卡尺(精度为0.01mm)测量叶片厚度, 测量时要求统一避开叶脉, 否则实际叶厚度偏大, 每个物种测量10次, 最后取平均值得出叶片的厚度。用铁铲挖取每株植物的根部, 深度为20—30cm, 然后将附带的泥土和杂质清理干净, 尽可能地收集完整的根部。采集到的样品用自封袋密封并编号, 储存在车载冰箱中, 以备室内分析。当天采样结束后, 选择10片测完叶厚度的叶子, 用扫描仪(CanoScan LiDE400)扫描成图像(分辨率为300dpi), 然后放入60℃的烘箱中烘干、称重, 计算比叶面积;将另一部分叶片装入信封, 送至实验室在80℃恒温条件下烘干至恒重, 然后粉碎机粉碎并过60目筛, 以测定叶片的营养成分。将带回的植物根系用清水冲洗掉附着的泥土并去除根毛, 然后用直尺测量10条须根的长度, 精度为0.1cm。刚洗净的根用滤纸吸附掉多余的水分, 然后将其完全浸入装有水的量筒中约5s, 读取量筒中水增加的体积作为根体积(排水替代法);然后放入烘箱中烘干称重, 计算比根长;用粉碎机将烘干后的所有根样粉碎, 然后用60目筛子过筛, 以测定根系的营养成分。扫描的叶片图像通过Image J软件计算得到叶面积。烘干后的植物样品粉碎后通过H2SO4-H2O2法消煮, 用AA3型连续流动分析仪(德国Auto Analyzer3)测定叶氮含量和根氮含量;用钼锑抗比色法测定叶磷含量和根磷含量。
1.4 土壤性状测定土壤水分采样, 每25cm取样1次, 取样深度为1m, 采用烘干法(105℃, 24h)测定土壤含水量。土壤养分采取S形随机采样, 每个样点3个重复, 分0—10、10—20cm和20—40cm三个层次混合后进行分析。采集的新鲜土壤在实验室自然干燥, 过20目筛后粉碎送试验。土壤有机碳含量测定采用重铬酸钾容量法-外加热法, 土壤全氮用凯氏法消解, 土壤全磷含量用钼锑抗比色法测定。
1.5 基于植物性状的植物功能型划分方法植物的功能性状在植物的生长和生存及其对环境变化的反应中并不是单独起作用的, 而是相互密切相关的, 通过权衡与组合自身不同功能性状, 产生了不同的生活策略来应对生境变化。Grime[28]认为植物的生活史因其受到的环境胁迫(限制光合作用的因素和干扰)而有很大的差异, 在此理论的基础上, 植物生活对策可以分为三种, 即“C-S-R”三角对策。适应低干扰和低压力的“竞争型”, 适应低干扰高压力的“胁迫忍耐型”, 和适应高干扰和低压力的“干扰容忍型”[16]。本文挑选易于观测的12项叶片和根性状指标进行聚类分析, 该方法能够基于植物性状精准筛选出具有相似适应策略的植物功能型, 其划分依据较以往研究中主观判断划分植物功能型更具客观性和代表性。所选性状指标及其生态意义见表 2。
| 植物性状指标 Indicators of plant traits |
指示的生态含义 Ecological significance of the indicators |
| 株高 Height |
表征植物提前攫取外界资源的能力, 例如高大植物的叶片可以吸收更多的光照资源用光合作用, 种子传播距离更远。 |
| 叶厚度 Leaf thickness |
表征植物利用环境资源的能力(尤其是储存和吸收水分的能力)。 |
| 叶面积 Leaf area |
植物吸收和转化太阳辐射的能力取决于叶面积的大小, 与植物光合效率呈一定相关性。 |
| 比叶面积 Specific leaf area |
作为植物资源利用分类轴上对植物物种进行分类的最佳变量[29], 不仅能衡量植物物种相对生长速度[30], 还对植物维持内部水分平衡非常重要。 |
| 叶干物质含量 Leaf dry matter content |
表征植物获取外界资源的能力以及适应胁迫环境的水平, 叶片干物质含量越高, 抗外界干扰能力越强。 |
| 组织密度 Tissue density |
组织密度不仅能反映植物各部位生物量的积累[31], 还与植物组织的拉伸力和防御力具有一定相关关系[32]。 |
| 比根长 Specific root length |
比根长作为一项关键的根系性状, 可以看作是植物地下性状的比叶面积[33]。它能反映植物对水分和养分的潜在吸收效率[34]。是表征植物地下竞争力大小的重要形态指标[33]。 |
| 叶片、根系氮磷含量 Nitrogen and phosphorus content in leaves and roots |
氮、磷是植物生长发育的必需元素, 而植物器官中的氮、磷含量不仅能精准反映土壤肥力的大小[35], 还能反映植物具体的生长状况, 以及受养分条件胁迫的状态[33]。 |
本研究首先运用组间关联聚类法对采集到的所有物种进行探索性分类, 以12个植物功能性状(株高、叶厚度、叶面积、比叶面积、叶组织密度、叶干物质含量、叶氮含量、叶磷含量、比根长、根组织密度、根氮含量和根磷含量)为基础, 分析这些性状之间的差异。按照分类间距离越小表示类间相似性越大, 距离越大表示组间间相似性越小的原则, 将15个物种划分为不同的功能类型。聚类采用的是欧氏距离平方来衡量, 所有数据均使用Z得分数据转化法进行标准化[36]。群落水平的功能性状值由物种水平的功能性状值按其重要值加权得出。功能性状加权平均值的计算使用R语言中的“FD”软件包。数据的预处理、统计学分析和表格制作由Microsoft Excel2010软件完成, 聚类分析和回归分析是通过SPSS25.0软件完成, 绘制图像均通过R语言完成。
植物群落水平功能性状根据群落相对重要值进行计算:

|
(1) |
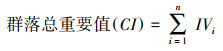
|
(2) |
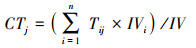
|
(3) |
式中, CTj代表群落或科属的j性状, Tij为群落中i 物种的j性状。
2 结果分析 2.1 毛乌素沙地植物功能型划分及其功能性状特征本研究采用组间关联聚类方法对采集和调查的物种进行探索性分类。在筛选过程中, 剔除了频率小于2的物种, 最终保留了15个植物物种。根据12项植物功能性状的之间差异, 毛乌素沙地常见物种被划分为不同的功能类型。聚类结果见树状图(图 2), 在虚线处根据相似性原则将其分为3类。结果显示, 第一类包含花棒, 踏郎, 角蒿, 牛皮消, 茵陈蒿, 青蒿, 沙蒿, 阿尔泰狗娃花, 柠条和沙打旺共10个物种, 其共同特征是较小的叶厚度、比叶面积、叶干物质含量和比根长, 同时表现为较大的株高、叶组织密度、叶氮含量、叶磷含量、根氮含量和根磷含量。第二类由沙蓬, 雾冰藜和狗尾草组成, 共3个物种, 这一功能型植物的特点是相对较小的株高、叶组织密度和叶干物质含量, 以及较大的比叶面积、叶磷含量、比根长和根组织密度。第三类则包括早熟禾与赖草两种植物, 这些植物的共同特性为拥有较大的叶厚度、叶组织密度、叶干物质含量和比根长, 但比叶面积、叶氮含量、叶磷含量、根组织密度、根氮含量和根磷含量相对较低。

|
| 图 2 毛乌素沙地飞播植被功能型分类及其性状特征差异 Fig. 2 Classification of plant functional types and differences in trait characteristics of aerial seeding vegetation in Mu Us Sandy Land PFT1:功能型Ⅰ植物; PFT2:功能型Ⅱ植物; PFT3:功能型Ⅲ植物; ****P<0.0001; ***P<0.001; ***P<0.01; *P<0.05 |
根据飞播植被演替恢复的群落特征和数量分布, 对不同时间序列(20a、40a、50a、60a)的功能型组成变化进行统计分析(表 3), 不同功能型的相对比重(%)为该类型物种数占群落总物种数的百分比。结果表明, 随演替序列延长, 物种丰富度(即物种数)是呈现增加的趋势, 尽管增加幅度不明显, 但对于毛乌素沙地恶劣的生境来说, 能够长期存活的物种数本就屈指可数。在演替过程中, 飞播植被与自然植被在演替早期的数量比例相对平衡, 分别占45%和55%。然而随演替时间的延长, 飞播植被的比例逐渐降低, 演替60年后仅占7%左右, 几乎被自然植被完全替代, 自然植被在演替后期占据了群落的绝对优势地位。在4个演替时期中, 功能型Ⅲ的物种数占全部物种数的百分比始终处于相对较低的水平, 由演替早期的9%降到演替后期(60a)不足2%;功能型Ⅰ由演替早期的52%降低到演替后期(60a)的28%, 功能型Ⅱ由38%上升至70%。这种变化说明:部分功能型Ⅲ的植物无法适应环境变化逐渐被其他类型植物替代, 而功能型Ⅰ在恢复初期占据群落中相对较大的优势, 然而整个演替期间其优势地位却呈现逐年下降的趋势。反观功能型Ⅱ随演替序列延长优势地位不断扩大, 在演替60年时占比达到最大值, 此时功能型Ⅱ植物数量占据群落内全部物种的70%。
| 演替年限 Time/a |
样点数 Number of sampling sites |
物种数 number of species |
飞播建群种 Dominant species |
飞播植被在群落中的数量比例/% Proportion of aerial seeding plants |
不同功能型组成/% Percentage of three PFTs |
||
| PFTⅠ | PFTⅡ | PFTⅢ | |||||
| 20 | 3 | 6.00±0.00 | 花棒、踏郎、沙蒿 | 45.12±10.46a | 52.49±17.48 | 37.72±19.18 | 9.79±2.20ab |
| 40 | 3 | 8.33±0.88 | 花棒、踏郎、沙蒿 | 28.32±4.88b | 38.13±4.59 | 44.14±3.91 | 11.62±4.81a |
| 50 | 5 | 7.60±0.51 | 花棒、踏郎、沙蒿 | 14.30±3.42bc | 32.02±6.90 | 66.90±6.43 | 3.08±1.63bc |
| 60 | 5 | 8.20±0.97 | 花棒、踏郎、沙蒿 | 7.36±2.96c | 28.19±13.50 | 70.07±13.08 | 1.74±0.87c |
| F | 1.547(3) | 10.116***(3) | 0.739(3) | 1.693(3) | 4.535*(3) | ||
| PFTⅠ: 功能型Ⅰ植物Plant functional typeⅠ; PFTⅡ: 功能型Ⅱ植物Plant functional typeⅡ; PFTⅢ: 功能型Ⅲ植物Plant functional typeⅢ; *** P<0.001; *** P<0.01; * P<0.05;括号内的数字为自由度 | |||||||
本研究共调查了16个样地, 并对不同演替序列的群落覆盖度、土壤水分和土壤养分进行了单因素方差分析。结果表明, 随演替进行, 群落盖度表现为先降低后升高的趋势。土壤水分在不同时间序列间差异显著(0.001<P<0.01), 土壤有机碳和土壤全氮变化较为显著(0.01<P<0.05), 土壤全磷和土壤碳氮比在不同时间序列间差异不显著(P>0.05)。表 4为飞播植被演替过程中群落盖度、土壤水分、土壤养分变化的单因素方差分析结果。结果表明, 在植被演替过程中, 除土壤全磷以外, 土壤有机碳、土壤全氮都随植被演替时间而呈增加趋势。土壤全磷在整个演替序列没有显著差异(P>0.05), 但其均值仅有0.15g/kg左右, 证明该地区的土壤全磷含量偏低。由于飞播后采取严禁放牧、围栏封育的政策, 基本上不存在放牧和砍伐现象, 植物群落演替变化的外部干扰可以忽略不计。因此, 本文假定在飞播植被演替过程中, 外部干扰是恒定不变的, 群落演替可以简单地看作是仅植物与环境相互作用的一个过程。
| 演替年限 Succession time/a |
样点数 Number of sample points |
群落盖度 Community coverage/% |
土壤水分 Soil moisture/% |
土壤有机碳 Soil organic carbon/(g/kg) |
土壤全氮 Soil total nitrogen/(g/kg) |
土壤碳氮比 Soil carbon and nitrogen ratio |
土壤全磷 Soil total phosphorus/(g/kg) |
| 20 | 3 | 20.00±12.65a | 1.77±0.86ab | 1.70±0.24ab | 0.11±0.01b | 15.48±0.85 | 0.16±0.04 |
| 40 | 3 | 12.70±4.75b | 2.43±1.32a | 1.52±0.20b | 0.10±0.02b | 14.76±1.00 | 0.15±0.02 |
| 50 | 5 | 20.52±7.77a | 1.16±0.52b | 2.15±0.76a | 0.14±0.04a | 15.35±0.69 | 0.15±0.02 |
| 60 | 5 | 21.20±4.35a | 1.93±1.01a | 1.90±0.50ab | 0.13±0.03ab | 15.10±1.64 | 0.15±0.01 |
| F | 3.924*(3) | 4.482***(3) | 3.010*(3) | 3.114*(3) | 0.723(3) | 0.088(3) | |
| *** P<0.001; *** P<0.01; * P<0.05;括号内的数字为自由度 | |||||||
通过双向逐步回归得到毛乌素沙地植物群落功能性状与土壤因子的最优回归方程(表 5)。结果表明, 土壤水分、土壤有机碳和土壤全磷含量是影响毛乌素沙地植物群落功能性状的主要土壤因子, 特别是对株高、叶厚度、比根长、根组织密度等植物群落功能性状有极显著的影响。除叶面积和根氮含量以外, 其余植物群落功能性状均能与土壤因子回归拟合。
| 性状 Traits |
逐步回归模型 Linear regression model |
相关系数 R2 |
P | F |
| 株高Height | H=35.585+63.998SOC-904.447TN | 0.504 | 0.001 | 13.138 |
| 叶厚Leaf thickness | LT=0.039+0.111SOC | 0.350 | 0.001 | 10.875 |
| 叶面积Leaf area | — | — | — | — |
| 比叶面积Specific leaf area | SLA=16.626+744.704SW | 0.243 | 0.030 | 4.875 |
| 叶组织密度Leaf tissue density | LD=4.977-0.791SOC | 0.222 | 0.048 | 4.035 |
| 叶干物质含量Leaf dry matter content | LDMC=0.555-1.482TP-2.596SW | 0.368 | 0.020 | 6.030 |
| 叶氮含量Leaf nitrogen content | LN=23.150-391.657SW+348.876TN-19.731SOC | 0.538 | 0.027 | 10.296 |
| 叶磷含量Leaf phosphorus conrent | LP=3.715+23.953SW-10.648TP | 0.477 | 0.003 | 11.361 |
| 比根长Specific root length | SRL=7726.278SW-52.901 | 0.409 | 0.000 | 15.423 |
| 根组织密度Root tissue density | RD=109.433SW-0.469 | 0.429 | 0.000 | 17.405 |
| 根氮含量Root nitrogen content | — | — | — | — |
| 根磷含量Root phosphorus content | RP=0.905-0.114SOC | 0.328 | 0.003 | 9.270 |
| SW: 土壤水分Soil water; SOC: 土壤有机碳Soil organic carbon; TN: 土壤全氮Soil total nitrogen; TP: 土壤全磷Soil total phosphorus; —表示模型模拟不显著(P>0.05) | ||||
对毛乌素沙地飞播植被演替过程中三种功能型组成变化进行分析(表 3), 结果表明, 在演替初期, 功能型Ⅰ胁迫-竞争型对策的植物占优势, 随着演替时间的延长, 其比例逐渐下降。而功能型Ⅱ竞争型对策的植物数量在整个演替序列都表现为持续增加, 至演替50年后取代功能型Ⅰ植物的优势地位。这是因为在演替初期, 种间竞争处于较低水平, 植物可利用资源相对丰富, 为植物生殖繁衍提供了可能。同时功能型Ⅰ植物植株高大, 光捕获能力较好, 长时间处于阳光照射下, 光合作用强烈, 植物叶片和根部含氮量较高, 表明植物生长良好, 容易建立含氮量较高的内环境, 以适应外部环境, 因此在演替初期具有相对更高的竞争力。但是随着飞播植被演替时间的延长, 生境中能量与资源分布的再分配一定程度上加剧了种间竞争[37], 为其他自然物种(沙蓬、雾冰藜)的入侵与定植提供了可能[30, 37]。演替过程中物种数量的增加导致植物对土壤水分、养分、光照等资源的竞争加剧, 养分消耗速度大于养分积累速度[38], 功能型Ⅰ植物因其比叶面积和比根长较小, 养分利用效率较低, 导致部分忍耐力和防御力较低的种无法适应竞争强烈、资源贫瘠的环境被淘汰。特别是, 功能型Ⅰ的一些豆科植物的生长受到磷供应量的严重制约, 而植物可吸收的土壤磷含量并没有明显增加(表 4), 这种限制对它们的生长和繁殖产生了不利影响。根据以往的研究发现, 植物比叶面积会随光照强度的降低而增加[39], 由于功能型Ⅱ的植株高度普遍较低, 光捕获能力较差, 且长期处于遮蔽状态, 只有依赖较大的比叶面积来增大叶片的捕光面积, 才能适应弱光环境。随着演替过程中土壤养分的增加, 为功能型Ⅱ植物提供了有利的外部环境, 这段时间植物充分利用养分迅速生长繁殖, 数量组成达到整个演替序列的峰值, 完全地占据了群落中的主导地位。Liu等人[40]对中国东部亚热带常绿阔叶林不同演替阶段的研究, 认为演替早期植物为适应光照利用率低、土壤温湿度多变的脆弱生境, 形成了一系列低生长速率的资源保守策略性状组合, 中后期植物为适应光照利用率低、生境竞争激烈的环境, 形成了较大比叶面积和比根长等逆向性状组合, 实现了高生长速率的资源获取策略。本研究中功能型Ⅱ植物也是凭借高生长速率的资源获取策略, 逐渐取代了群落中功能型Ⅰ植物的优势地位, 而功能型Ⅲ植物拥有较厚的叶和较高的叶干物质含量, 在各阶段所占比例逐渐下降, 演替后期逐渐淘汰的原因可能是由于有限的资源被过多投入到结构建设中, 而且比叶面积较小, 生长速率慢, 长时间处于叶氮、根氮、根磷含量很低的状态, 最终达不到植物生存的养分需求而死亡。而以花棒、踏郎、沙蒿为代表的先锋物种可能在早期阶段可利用沙地较为丰富的地下储水获得较好的生长, 但随着土壤水分的消耗, 必然反馈于植物生长过程, 一些低耗水的乡土物种进入群落, 这也是达尔文进化法则在群落演替过程的具体体现。因此, 本研究认为随着飞播植被演替的进行, 自然过程介入是群落趋于自然化发育的必然, 是植被演替过程中植被与土壤水分互馈作用的最终表现。。
3.2 毛乌素沙地飞播植被群落性状值与土壤因子响应毛乌素沙地有明显的旱季且温度高、地表蒸发大, 导致该区干旱现象严重, 土壤水分是制约植被正常生长的主要因子[41]。在干旱的黄土高原, 演替晚期群落的土壤水分高于演替初期群落, 随着时间推移, 土壤水分条件逐渐改善[42], 这与本研究发现的土壤水分变化结果一致。此外, 土壤养分变化规律也与李亚锦[43]的研究结果一致, 土壤中的SOC和TN含量均随演替逐渐增加, 说明飞播植被的恢复对这些养分含量具有明显的累积作用, 为自然植被取代飞播物种实现近自然化过程提供了外部条件。对毛乌素沙地飞播植被演替过程中群落功能性状值与土壤因子做回归分析(表 5), 表明土壤水分、土壤碳含量和土壤磷含量是影响该区域植物群落功能性状变化的主要土壤因子, 尤其是对株高、叶厚度、比根长、根组织密度有极显著的影响。其中土壤水分与比叶面积呈显著正相关, 随着演替过程中土壤水分条件得到改善, 植被更倾向于通过提高比叶面积来提高对光资源的利用效率, 采取快速生长的策略实现生存、繁殖的目的。这与刘可佳[44]对不同环境条件下植物比叶面积适应对策的研究结果一致, 可能是因为植物受到水分胁迫时, 它们会通过关闭气孔、减小比叶面积使细胞变得紧凑, 来达到减少水分向外扩散的目的[45—46]。而土壤水分充足的地区, 生长条件相对适宜, 环境中具有较高比叶面积的植物表现为更高的存活率。叶片干物质含量与土壤水分呈明显的负相关, 随着土壤水分的恢复, 植被的叶片干物质含量越小。这与王鑫[47]对黄土高原典型小流域草地群落功能性状的研究结果一致, 在土壤水分缺乏的地区, 植物增加叶片干物质含量是为了保存更多水分, 从而增加水分从叶片内部向叶片表面扩散的距离和阻力, 并通过降低比叶面积来减弱蒸腾作用[48]。通常情况下, 在土壤水分条件良好的生境, 群落内植物的比叶面积较大、叶干物质含量较低, 这在某种程度上反映出, 植物适应生存环境的过程与土壤水分的变化具有一定的规律性。同比叶面积变化性质类似, 叶干物质含量的下降表面更多的合成物质被用于生长构造, 植被趋向于采取快速生长策略来提高对资源的利用效率, 进而提高其生存适合度。总而言之, 随演替进行, 植物会根据生境的变化对其采取的性状策略进行调整, 即能够高效利用资源快速生长的获取型策略替代了低生长速率、注重投入防御的保守型策略。
4 结论研究发现, 毛乌素沙地飞播植被演替过程中土壤水分、土壤有机碳和土壤全磷是造成群落功能性状值差异的关键土壤因子。由于土壤水分、养分等因素的影响以及生物间的相互作用, 不同阶段的植物具有不同的功能型组成, 采用的环境适应策略也各不相同。随着植被演替时间延长, 能够高效利用资源快速生长的获取型策略代替了低生长速率、注重防御构造的保守型策略。当飞播种在群落中的数量骤减, 逐渐被沙蓬、雾冰藜为代表的自然物种所取代时, 表明飞播植被逐渐开始近自然化过程。飞播植被的恢复对群落养分含量具有明显的累积作用, 为沙蓬、雾冰藜等典型沙生植物的生长繁殖提供了外部条件。这说明飞播造林(草)能够促进群落的进展演替, 以土壤理化性质为介导实现了飞播植被演替过程中天然物种的更新。总之, 飞播植被退化并不会导致土壤地下水过度消耗, 反而其近自然化过程能带来良好的生态效益, 提高群落多样性和稳定性, 促进退化生态系统的修复, 为毛乌素沙地生态系统可持续恢复提供了理论依据。
| [1] |
韩雪莹, 杨光, 秦富仓, 贾光普, 凌侠, 高岗. 毛乌素沙地近30年沙漠化土地时空动态演变格局. 水土保持研究, 2019, 26(5): 144-150. |
| [2] |
裴艳武, 黄来明, 李荣磊, 邵明安, 张应龙. 毛乌素沙地东南缘人工林樟子松根系吸水来源与影响因素. 土壤学报, 2022, 59(5): 1336-1348. |
| [3] |
钱洲, 俞元春, 俞小鹏, 高捍东, 吕荣, 张文英. 毛乌素沙地飞播造林植被恢复特征及土壤性质变化. 中南林业科技大学学报, 2014, 34(4): 102-107. |
| [4] |
钱洲, 俞元春, 俞小鹏, 高捍东, 吕荣, 张文英. 毛乌素沙地飞播造林植被恢复过程土壤酶活性的变化. 水土保持研究, 2014, 21(6): 95-100. |
| [5] |
袁媛, 张鹤, 武杼华, 王钰, 毕博远, 韩凤朋. 植被恢复对毛乌素沙地土壤pH和养分含量的影响. 土壤通报, 2021, 52(1): 148-156. |
| [6] |
关其格, 浩必斯嘎拉图, 杨越, 韩永娇, 武智勇, 宋双双, 赵瑞. 毛乌素沙地南缘人工恢复植被对沙地土壤理化性质的影响. 安徽农业科学, 2013, 41(34): 13217-13220. |
| [7] |
王德富, 董凌勃, 李澳, 于志静, 邓蕾. 毛乌素沙地不同植被恢复类型的土壤碳水效应. 水土保持学报, 2024, 38(3): 101-110. |
| [8] |
白星雯, 布日古德, 洪光宇, 李炜, 邵飞舟, 张雷. 毛乌素沙地典型人工林土壤水分特征及其对降水的响应. 林草资源研究, 2023, 45(6): 52-60. |
| [9] |
姜晓燕, 高圣杰, 蒋燕, 田赟, 贾昕, 查天山. 毛乌素沙地植被不同恢复阶段植物群落物种多样性、功能多样性和系统发育多样性. 生物多样性, 2022, 30(5): 18-28. |
| [10] |
王妍, 吴波, 李小英, 卢琦. 毛乌素沙地油蒿群落不同演替阶段的物种多样性研究. 干旱区资源与环境, 2011, 25(2): 167-172. |
| [11] |
刘飞雄. 毛乌素沙地典型植被恢复模式生态效益研究. 现代园艺, 2015, 38(16): 176. |
| [12] |
高佳, 王文科, 赵明, 马稚桐, 侯昕悦, 李婉歆. 毛乌素沙地裸地与植被覆盖下非冻结期土壤水分时空分布特征. 水文地质工程地质, 2022, 49(6): 34-42. |
| [13] |
Feng X M, Fu B J, Piao S L, Wang S, Ciais P, Zeng Z Z, Lü Y H, Zeng Y, Li Y, Jiang X H, Wu B F. Revegetation in China's Loess Plateau is approaching sustainable water resource limits. Nature Climate Change, 2016, 6(11): 1019-1022. |
| [14] |
耿国彪. 警惕毛乌素沙地二次沙化. 绿色中国, 2022, 19(11): 24-29. |
| [15] |
Violle C, Navas M L, Vile D, Kazakou E, Fortunel C, Hummel I, Garnier E. Let the concept of trait be functional!. Oikos, 2007, 116(5): 882-892. |
| [16] |
郑颖, 温仲明, 宋光, 丁曼. 延河流域森林草原区不同植物功能型适应策略及功能型物种数量随退耕年限的变化. 生态学报, 2015, 35(17): 5834-5845. |
| [17] |
Woodward F I, Cramer W. Plant functional types and climatic change: Introduction. Journal of Vegetation Science, 1996, 7(3): 306-308. |
| [18] |
胡楠, 范玉龙, 丁圣彦, 卢训令. 伏牛山自然保护区森林生态系统乔木植物功能型分类. 植物生态学报, 2008, 32(5): 1104-1115. |
| [19] |
Diaz S, Cabido M. Vive la différence: plant functional diversity matters to ecosystem processes. Trends in Ecology & Evolution, 2001, 16(11): 646-655. |
| [20] |
Aguiar M R, Paruelo J M, Sala O E, Lauenroth W K. Ecosystem responses to changes in plant functional type composition: an example from the Patagonian steppe. Journal of Vegetation Science, 1996, 7(3): 381-390. |
| [21] |
Jobbágy E G, Paruelo J M, León R J C. Vegetation heterogeneity and diversity in flat and mountain landscapes of Patagonia (Argentina). Journal of Vegetation Science, 1996, 7(4): 599-608. |
| [22] |
Skarpe C. Plant functional types and climate in a southern African savanna. Journal of Vegetation Science, 1996, 7(3): 397-404. |
| [23] |
Box E O. Plant functional types and climate at the global scale. Journal of Vegetation Science, 1996, 7(3): 309-320. |
| [24] |
韩梅, 杨利民, 张永刚, 周广胜. 中国东北样带羊草群落C3和C4植物功能群生物量及其对环境变化的响应. 生态学报, 2006, 26(6): 1825-1832. |
| [25] |
Chapin F S III, Bret-Harte M S, Hobbie S E, Zhong H L. Plant functional types as predictors of transient responses of Arctic vegetation to global change. Journal of Vegetation Science, 1996, 7(3): 347-358. |
| [26] |
Petchey O L, Gaston K J. Functional diversity (FD), species richness and community composition. Ecology Letters, 2002, 5(3): 402-411. |
| [27] |
Díaz Barradas M C, Zunzunegui M, Tirado R, Ain-Lhout F, García Novo F. Plant functional types and ecosystem function in Mediterranean shrubland. Journal of Vegetation Science, 1999, 10(5): 709-716. |
| [28] |
Hodáňová D. Plant strategies and vegetation processes. Biologia Plantarum, 1981, 23(4): 254. |
| [29] |
宝乐, 刘艳红. 东灵山地区不同森林群落叶功能性状比较. 生态学报, 2009, 29(7): 3692-3703. |
| [30] |
郑淑霞, 上官周平. 不同功能型植物光合特性及其与叶氮含量、比叶重的关系. 生态学报, 2007, 27(1): 171-181. |
| [31] |
Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T L, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets V, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821-827. |
| [32] |
Craine J M, Froehle J, Tilman D G, Wedin D A, Wedin D A. The relationships among root and leaf traits of 76 grassland species and relative abundance along fertility and disturbance gradients. Oikos, 2001, 93(2): 274-285. |
| [33] |
Cornelissen J H C, Lavorel S, Garnier E, Díaz S, Buchmann N, Gurvich D E, Reich P B, ter Steege H, Morgan H D, van der Heijden M G A, Pausas J G, Poorter H. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Australian Journal of Botany, 2003, 51(4): 335. |
| [34] |
Turner I M. Sclerophylly: primarily protective?. Functional Ecology, 1994, 8(6): 669. |
| [35] |
Field C, Merino J, Mooney H A. Compromises between water-use efficiency and nitrogen-use efficiency in five species of California evergreens. Oecologia, 1983, 60(3): 384-389. |
| [36] |
郑颖. 延河流域植物叶片与细根功能性状的时空变化[D]. 杨凌: 西北农林科技大学, 2014.
|
| [37] |
傅致远, 姜宏, 王国强, 阿膺兰, 薛宝林, 王红旗. 半干旱草原区土壤性质对植物群落结构的影响. 生态学杂志, 2018, 37(3): 823-830. |
| [38] |
郭曼, 郑粉莉, 和文祥, 安韶山, 刘雨, 安娟. 黄土丘陵区不同退耕年限植被多样性变化及其与土壤养分和酶活性的关系. 土壤学报, 2010, 47(5): 979-986. |
| [39] |
Niinemets V, Kull O, Tenhunen J D. Within-canopy variation in the rate of development of photosynthetic capacity is proportional to integrated quantum flux density in temperate deciduous trees. Plant, Cell & Environment, 2004, 27(3): 293-313. |
| [40] |
Liu L B, Xia H J, Quan X H, Wang Y Q. Plant trait-based life strategies of overlapping species vary in different succession stages of subtropical forests, Eastern China. Frontiers in Ecology and Evolution, 2023, 10: 1103937. |
| [41] |
Jin Z, Li X R, Wang Y Q, Wang Y, Wang K B, Cui B L. Comparing watershed black locust afforestation and natural revegetation impacts on soil nitrogen on the Loess Plateau of China. Scientific Reports, 2016, 6: 25048. |
| [42] |
韦兰英, 上官周平. 黄土高原不同演替阶段草地植被细根垂直分布特征与土壤环境的关系. 生态学报, 2006, 26(11): 3740-3748. |
| [43] |
李亚锦, 郑景明, 王根柱, 周金星, 刘玉国, 哈文秀. 喀斯特区天然林不同演替阶段功能性状特征及其影响因素研究——以云南大黑山为例. 地球学报, 2021, 42(3): 397-406. |
| [44] |
刘可佳, 何念鹏, 侯继华. 中国温带典型森林植物比叶面积的空间格局及其影响因素. 生态学报, 2022, 42(3): 872-883. |
| [45] |
Galmés J, Ochogavía J M, Gago J, Roldán E J, Cifre J, Conesa M À. Leaf responses to drought stress in Mediterranean accessions of Solanum lycopersicum: anatomical adaptations in relation to gas exchange parameters. Plant, Cell & Environment, 2013, 36(5): 920-935. |
| [46] |
田俊霞, 魏丽萍, 何念鹏, 徐丽, 陈智, 侯继华. 温带针阔混交林叶片性状随树冠垂直高度的变化规律. 生态学报, 2018, 38(23): 8383-8391. |
| [47] |
王鑫, 杨磊, 赵倩, 张钦弟. 黄土高原典型小流域草地群落功能性状对土壤水分的响应. 生态学报, 2020, 40(8): 2691-2697. |
| [48] |
Maharjan S K, Poorter L, Holmgren M, Bongers F, Wieringa J J, Hawthorne W D. Plant functional traits and the distribution of West African rain forest trees along the rainfall gradient. Biotropica, 2011, 43(5): 552-561. |