 2024, Vol. 44
2024, Vol. 44文章信息
- 查苏娜, 吴桂林, 周璋, 吴建辉, 张涛, 农寿千, 陈德祥
- CHA Suna, WU Guilin, ZHOU Zhang, WU Jianhui, ZHANG Tao, NONG Shouqian, CHEN Dexiang
- 海南岛热带次生草地生物多样性变化的气候驱动效应
- The climate-driven effects on biodiversity changes in the tropical secondary grasslands of Hainan
- 生态学报. 2024, 44(23): 10864-10874
- Acta Ecologica Sinica. 2024, 44(23): 10864-10874
- http://dx.doi.org/10.20103/j.stxb.202401290262
-
文章历史
- 收稿日期: 2024-01-29
- 网络出版日期: 2024-08-29





2. 南京林业大学, 南京 210037;
3. 中国林业科学研究院热带林业研究所试验站, 尖峰 572542;
4. 海南省林业科学研究院(海南省红树林研究院), 海口 571100
2. Nanjing Forestry University, Nanjing 210037, China;
3. Experimental Station of Research Institute of Tropical Forestry, Chinese Academy of Forestry, Jianfeng 572542, China;
4. Hainan Academy of Forestry (Hainan Academy of Mangrove), Haikou 571100, China
次生草地主要是指受到自然因素和人类活动的破坏后, 经过自然恢复或人工干预形成的草地, 多分布于我国南方地区, 在维持生态平衡、提供碳储存、调节气候等方面发挥重要作用[1]。相比原生性草地, 次生草地生态系统的稳定性较差, 物种多样性较低[2], 但通过适当的管理措施, 其物种多样性可以得到显著的提升, 是进行生态系统恢复与物种多样性保护的重要研究对象[3—4]。
次生草地物种多样性受多种因素影响, 如放牧、刈割、火烧、耕作等人为干扰活动以及土壤、地形、气候等环境条件[5], 其中温度和降水因子尤为重要[6]。当次生草地处于恢复阶段时, 其群落结构与生态系统的稳定性较差, 水热因子将直接影响物种多样性[7]。已有研究显示, 年均温[8]和温度波动[9]显著影响草地物种多样性, 温暖湿润且气候稳定的环境, 水热条件良好, 资源利用率高, 为物种提供了更多的气候生态位[10], 可加速促进次生草地的恢复和物种多样性的增加。次生草地的群落结构相对简单, 对极端温度适应性较差, 随着温度的剧烈波动或者极端低温(高温)的发生, 物种多样性将迅速减少。众多学者发现, 极端温度会损害植物生理功能[11], 加剧土壤水分胁迫[12], 导致对温度敏感的物种大面积减少或死亡, 进而影响草地群落物种组成, 造成物种多样性的大幅度下降[13]。降水对次生草地物种多样性的影响在干旱和半干旱地区尤为明显[14]。例如, 在水分稀缺的温带草原[15]和非洲南部稀树草原[16], 长期的干扰导致草地土壤结构松散, 降水的增加可以显著提高土壤水分有效性, 减少水分亏缺导致的物种死亡, 有助于增加物种多样性。然而, 次生草地植被覆盖度较低, 极端降水事件常伴随强烈的风暴, 会导致土壤侵蚀[17], 加剧草地退化, 从而降低物种多样性。
草地物种多样性的关键性气候驱动因子可能随着纬度的变化而有所不同。大量的研究表明, 除温度外, 水分是影响北方高纬度次生草地物种多样性的另一重要气候因子[18—23]。在内蒙古[18]和藏北地区[19]发现年均降水量显著影响受干扰后的草地物种多样性;最湿季度降水量能够显著促进宁夏荒漠草原物种多样性[20];锡林郭勒[21]和新疆北部[22]农牧交错区的次生草地物种多样性均受降水因子影响。除降水量外, 草地物种多样性还受到降水频率的影响, 例如:在荒漠草原[23], 频繁的小降水事件显著促进物种多样性。
总之, 现有研究开展了大量内蒙古、新疆、西藏、青海以及甘肃等北方高纬度草地生态系统物种多样性与气候因子相关关系的研究[18—25], 而南方低纬度地区, 尤其是热带次生草地物种多样性与气候的关系尚不清楚。因此, 本文以海南岛热带次生草地为研究对象, 基于海南岛6个市县43个30 m×30 m草地样地的物种名称和数量, 分析海南岛主要热带次生草地物种组成;计算物种多样性指数, 结合温度和降水相关的19个气候因子, 探讨影响海南岛热带次生草地物种多样性的气候驱动因子, 了解其是否与以往研究中影响北方次生草地的气候驱动因子相同, 以期探索热带草地物种多样性与气候的关系, 并为热带次生草地生态系统的恢复提供重要参考依据。
1 材料与方法 1.1 研究区概况研究区位于海南岛, 地处我国大陆南端, 地理位置介于东经108°37—113°3′, 北纬18°10′—20°10′之间(图 1)。海南岛地势中间高, 四周低, 由山地、丘陵、台地、平原构成环形层状地貌, 梯级结构明显;海南岛地处热带北缘, 属典型的热带季风性气候, 年均气温为23—25℃, 最热月平均气温为28—30℃, 最冷月平均气温为17—20℃, 气温年较差在16—19℃[26];海南岛雨量充沛, 全岛年平均降水量为1600 mm, 干湿两季明显, 80%—90% 的雨量集中在雨季[27];海南岛降水分布不均匀, 东部多雨地区年降水量可达2400 mm以上, 而西部沿海少雨地区年降水量仅为1000 mm左右[28], 降水梯度明显。

|
| 图 1 海南岛概况与采样点 Fig. 1 Overview of Hainan Island and Sampling Sites |
目前在海南, 除了少数保持其原生特性的湿生性草地之外, 大部分草地受到一定程度的人类活动的干扰, 包括因开垦和改造、森林砍伐, 以及源自弃耕而形成的次生性草地[29]。海南岛草地面积较少, 主要分为两类, 地带性次生草丛和非地带性草丛, 主要分布在海南岛西部和北部沿海地区以及中部海拔较低的山坡和丘陵地带, 以中生型的多年生物种组成, 伴有少数乔木和灌木。地带性次生草丛分为禾草草丛和野蕉草丛, 以白茅、芒、红毛鸭嘴草、牛筋草、假臭草、飞机草、地毯草和含羞草等为主;非地带性草丛多为湿生草丛, 生长在海南岛沿海湿地、河流、湖泊、沼泽以及终年积水的低洼地等[30]。本次调查的草地样地均为受到农垦、采伐和轮牧等干扰后形成的地带性次生性草地。
1.3 实验设计和样品采集2022年1—4月在海南岛草地分布相对较多的文昌市、屯昌县、临高县、儋州市、昌江县、东方市设置43个30 m×30 m草地样地(图 1), 用GPS仪(Garmin12XL, Garmin Inc., USA)确定每个样地的经纬度。在每个样地四角、中心和样地周围20 m范围内布设6个样方(图 2), 按不同植被类型和植被密度布设不同面积的样方, 草甸为1 m×1 m样方, 灌草丛为2 m×2 m样方[31]。调查并记录样方内所有物种名称、数量和盖度, 计算物种多样性指数和重要值。

|
| 图 2 样方示意图 Fig. 2 Schematic diagram of the quadrat |
根据每个样地经纬度, 利用Rstudio 4.2.0(Rstudio Inc., Boston, MA, USA)软件中的“raster”程序包“extract”函数, 从Worldclim网站(https://www.worldclim.org/)中提取43个草地样地的19个温度和降水相关因子(表 1)。
| 生物气候变量 Bioclimatic variables |
描述 Description |
生物气候变量 Bioclimatic variables |
描述 Description |
|
| BIO1 | 年均气温 | BIO11 | 最冷季度平均温度 | |
| BIO2 | 昼夜温差月均值 | BIO12 | 年降水量 | |
| BIO3 | 等温性 | BIO13 | 最湿月份降水量 | |
| BIO4 | 温度季节性变化 | BIO14 | 最干月份降水量 | |
| BIO5 | 最暖月份最高温度 | BIO15 | 降水量季节性变化 | |
| BIO6 | 最冷月份最低温度 | BIO16 | 最湿季度降水量 | |
| BIO7 | 气温年较差 | BIO17 | 最干季度降水量 | |
| BIO8 | 最湿季度平均温度 | BIO18 | 最暖季度降水量 | |
| BIO9 | 最干季度平均温度 | BIO19 | 最冷季度降水量 | |
| BIO10 | 最暖季度平均温度 |
(1) 物种重要值计算
重要值(Importance Value)是一个综合考虑物种在某一生态系统中的相对多度、相对频度和相对盖度的指标, 用于评估某一物种在生态系统中的相对重要性。计算方法如下:

|
(2) 物种多样性计算
本文选取丰富度(R)、Simpson指数(D)和Shannon-Wiener指数(H)表征物种多样性, 对物种数量、物种丰度以及物种分布的均匀度进行综合评价, 计算方法如下:
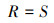
|
(1) |
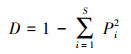
|
(2) |
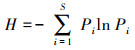
|
(3) |
式中, S为物种数目, Pi为此物种个体数占总个体数的比例[32]。采用Pearson相关性分析明确19个气候变量与物种多样性之间的关系。数据分析和制图分别由Origin 2021(OriginLab Inc., Northampton, MA, USA)、Rstudio 4.2.0(Rstudio Inc., Boston, MA, USA)和ArcGIS 10.7(ESRI Inc., Redlands, California, USA)完成。
2 结果与分析 2.1 海南岛热带次生草地植被物种组成经调查, 在海南岛43个草地样地中共包含131个物种, 隶属于106属、34科。表 2列出了重要值排名前十的科类, 禾本科(20属23种)、菊科(20属24种)和豆科(17属20种)是海南岛热带次生草地的主要组成科类, 重要值分别为35.966%、17.748%和13.521%, 三者之和占整体的67.235%。其次是茜草科(3属4种)、莎草科(4属9种)和锦葵科(4属6种), 其重要值分别为9.595%、7.531%和5.495%。
| 科名 Family name |
属的数量 Number of genera |
物种数量 Number of species |
重要值/% Important value |
| 禾本科Poaceae | 20 | 25 | 35.966 |
| 菊科Asteraceae | 20 | 24 | 17.748 |
| 豆科Fabaceae | 17 | 20 | 13.521 |
| 茜草科Rubiaceae | 3 | 4 | 9.595 |
| 莎草科Cyperaceae | 4 | 9 | 7.531 |
| 锦葵科Malvaceae | 4 | 6 | 5.495 |
| 大戟科Euphorbiaceae | 3 | 4 | 0.748 |
| 旋花科Convolvulaceae | 2 | 4 | 0.385 |
| 唇形科Lamiaceae | 3 | 3 | 0.387 |
| 马鞭草科Verbenaceae | 3 | 3 | 0.322 |
表 3为海南岛热带次生草地植被物种重要值排名前25的物种以及对应的相对频度、相对盖度和相对丰度。前十的物种分别为铺地黍、狗牙根、三点金、糙叶丰花草、地毯草、假臭草、紫马唐、香附子、赛葵和含羞草, 其重要值之和占整体的49.973%, 是构成海南岛热带次生草地植被群落的重要物种。其中铺地黍的相对频度和相对盖度最高, 分别为5.980%和10.629%;三点金数量最多, 相对多度为7.916%。海南岛热带次生草地以热带和亚热带物种为主, 在我国广东、广西、福建、云南、海南和台湾等地区广泛分布, 含丰花草和三点金等本地种, 假臭草和飞机草等少数外来入侵物种。
| 物种名称 Species name |
相对频度/% Relative frequency |
相对盖度/% Relative coverage |
相对多度/% Relative abundance |
重要值/% Important value |
| 铺地黍Panicum repens | 5.980 | 10.629 | 6.461 | 7.690 |
| 狗牙根Cynodon dactylon | 5.249 | 10.470 | 6.212 | 7.311 |
| 三点金Desmodium triflorum | 4.917 | 6.991 | 7.916 | 6.608 |
| 糙叶丰花草Spermacoce hispida | 4.983 | 6.116 | 6.248 | 5.782 |
| 地毯草Axonopus compressus | 2.791 | 6.041 | 4.420 | 4.417 |
| 假臭草Praxelis clematidea | 4.385 | 3.690 | 5.041 | 4.372 |
| 紫马唐Digitaria violascens | 3.522 | 5.008 | 3.159 | 3.896 |
| 香附子Cyperus rotundus | 2.658 | 3.369 | 5.218 | 3.748 |
| 赛葵Malvastrum coromandelianum | 4.651 | 2.529 | 2.751 | 3.310 |
| 含羞草Mimosa pudica | 3.389 | 2.697 | 2.432 | 2.839 |
| 夜香牛Vernonia cinerea | 2.193 | 1.204 | 4.455 | 2.617 |
| 龙爪茅Dactyloctenium aegyptium | 2.857 | 2.050 | 2.201 | 2.370 |
| 鬼针草Bidens pilosa | 1.595 | 3.029 | 1.757 | 2.127 |
| 丰花草Spermacoce pusilla | 2.259 | 1.572 | 1.899 | 1.910 |
| 白花蛇舌草Scleromitrion diffusum | 1.927 | 1.611 | 2.059 | 1.866 |
| 链荚豆Alysicarpus vaginalis | 2.193 | 1.683 | 1.438 | 1.771 |
| 野甘草Scoparia dulcis | 1.860 | 0.650 | 2.574 | 1.695 |
| 鸭跖草Commelina communis | 1.794 | 1.286 | 1.864 | 1.648 |
| 扁穗莎草Cyperus compressus | 2.193 | 1.307 | 1.384 | 1.628 |
| 红毛草Melinis repens | 0.997 | 2.436 | 1.331 | 1.588 |
| 长蒴母草Lindernia anagallis | 1.728 | 0.897 | 2.059 | 1.561 |
| 鼠曲草Pseudognaphalium affine | 1.528 | 0.925 | 1.970 | 1.475 |
| 鲫鱼草Eragrostis tenella | 1.728 | 1.261 | 0.958 | 1.316 |
| 羽芒菊Tridax procumbens | 1.993 | 0.614 | 1.083 | 1.230 |
| 小蓬草Erigeron canadensis | 1.395 | 0.686 | 0.923 | 1.001 |
由图 3可知, 海南岛热带次生草地丰富度与昼夜温差月均值(BIO2)、等温性(BIO3)和气温年较差(BIO7)之间存在显著正相关关系;其中BIO2对丰富度的影响极为显著(P=0.009), 可以解释丰富度15.5% 的变化原因;BIO3和BIO7与丰富度显著相关(P=0.039), 分别能够解释丰富度10%和9.8%的变化原因。BIO2、BIO3和BIO7均为温度波动相关因子, 三者之和能够解释海南岛热带次生草地物种丰富度35.3% 的变化原因;其它温度相关因子(BIO1、BIO4、BIO5、BIO6、BIO8、BIO9、BIO10、BIO11)与丰富度均无显著相关性(P>0.050)(表 4)。

|
| 图 3 丰富度与气候变量的线性拟合图 Fig. 3 Linear fit of richness and climate variables |
| 变量 Variables |
丰富度Richness | 变量 Variables |
丰富度Richness | |||||
| R2 | P | 相关系数 Correlation coefficient |
R2 | P | 相关系数 Correlation coefficient |
|||
| BIO1 | 0.009 | 0.551 | -0.094 | BIO11 | < 0.001 | 0.972 | -0.006 | |
| BIO2 | 0.155 | 0.009** | 0.394 | BIO12 | 0.003 | 0.733 | 0.054 | |
| BIO3 | 0.100 | 0.039* | 0.316 | BIO13 | < 0.001 | 0.865 | 0.027 | |
| BIO4 | 0.011 | 0.501 | -0.105 | BIO14 | 0.002 | 0.760 | 0.048 | |
| BIO5 | 0.042 | 0.188 | 0.205 | BIO15 | 0.003 | 0.715 | -0.057 | |
| BIO6 | 0.071 | 0.085 | -0.266 | BIO16 | 0.006 | 0.629 | 0.076 | |
| BIO7 | 0.098 | 0.041* | 0.312 | BIO17 | 0.003 | 0.736 | 0.053 | |
| BIO8 | 0.010 | 0.519 | -0.101 | BIO18 | < 0.001 | 0.982 | -0.003 | |
| BIO9 | 0.012 | 0.488 | 0.109 | BIO19 | 0.003 | 0.719 | 0.057 | |
| BIO10 | 0.052 | 0.142 | -0.228 | |||||
| *表示显著相关(P < 0.050), * *表示极显著相关(P < 0.010) | ||||||||
海南岛热带次生草地丰富度与降水相关因子(BIO12、BIO13、BIO14、BIO15、BIO16、BIO17、BIO18、BIO19)均无显著相关性(P>0.050)(表 4)。
2.2.2 Simpson指数与气候变量如图 4所示, 海南岛热带次生草地Simpson优势度指数与年均气温(BIO1)、最冷月份最低温度(BIO6)和最冷季度平均温度(BIO11)呈显著负相关关系。其中, BIO6与Simpson指数呈极显著相关关系, P值为0.008, R2为0.159;BIO1和BIO11与Simpson指数呈显著相关关系, P值分别为0.021和0.045, R2分别为0.123和0.095;而其它温度相关因子(BIO2、BIO3、BIO4、BIO5、BIO7、BIO8、BIO9、BIO10)与Simpson指数之间并不存在显著相关性(P>0.050)(表 5)。

|
| 图 4 Simpson指数与气候变量的拟合曲线 Fig. 4 Fit curve between Simpson index and climate variables |
| 变量 Variables |
Simpson指数Simpson index | 变量 Variables |
Simpson指数Simpson index | |||||
| R2 | P | 相关系数 Correlation coefficient |
R2 | P | 相关系数 Correlation coefficient |
|||
| BIO1 | 0.123 | 0.021* | -0.351 | BIO11 | 0.095 | 0.045* | -0.307 | |
| BIO2 | 0.023 | 0.328 | 0.153 | BIO12 | 0.033 | 0.241 | 0.183 | |
| BIO3 | < 0.001 | 0.970 | 0.006 | BIO13 | 0.029 | 0.277 | 0.169 | |
| BIO4 | 0.020 | 0.361 | 0.143 | BIO14 | 0.025 | 0.312 | 0.158 | |
| BIO5 | < 0.001 | 0.955 | -0.009 | BIO15 | 0.034 | 0.240 | -0.183 | |
| BIO6 | 0.159 | 0.008** | -0.398 | BIO16 | 0.043 | 0.183 | 0.207 | |
| BIO7 | 0.085 | 0.058 | 0.292 | BIO17 | 0.027 | 0.290 | 0.165 | |
| BIO8 | 0.009 | 0.537 | 0.097 | BIO18 | 0.027 | 0.295 | 0.163 | |
| BIO9 | 0.034 | 0.236 | -0.184 | BIO19 | 0.028 | 0.282 | 0.168 | |
| BIO10 | 0.086 | 0.056 | -0.293 | |||||
| *表示显著相关(P < 0.050), * *表示极显著相关(P < 0.010) | ||||||||
降水相关因子(BIO12、BIO13、BIO14、BIO15、BIO16、BIO17、BIO18、BIO19)与海南岛热带次生草地Simpson指数均无显著相关性(P>0.050)(表 5)。
2.2.3 Shannon-Wiener指数与气候变量海南岛热带次生草地Shannon-Wiener多样性指数受到最冷月份最低温度(BIO6)和气温年较差(BIO7)的显著影响(图 5), 其中, BIO6与Shannon-Wiener指数显著负相关(P=0.011), BIO7与Shannon-Wiener指数呈显著正相关关系(P=0.043);然而, 其它温度相关因子(BIO1、BIO2、BIO3、BIO4、BIO5、BIO8、BIO9、BIO10、BIO11)对Shannon-Wiener指数均无显著影响(P>0.050)(表 6)。

|
| 图 5 Shannon-Wiener指数与气候变量的线性拟合图 Fig. 5 Linear fitting of Shannon-Wiener index to climate variables |
| 变量 Variables |
Shannon-Wiener指数Shannon-Wiener index | 变量 Variables |
Shannon-Wiener指数Shannon-Wiener index | |||||
| R2 | P | 相关系数 Correlation coefficient |
R2 | P | 相关系数 Correlation coefficient |
|||
| BIO1 | 0.085 | 0.057 | -0.292 | BIO11 | 0.066 | 0.097 | -0.256 | |
| BIO2 | 0.045 | 0.172 | 0.212 | BIO12 | 0.027 | 0.289 | 0.166 | |
| BIO3 | 0.005 | 0.639 | 0.074 | BIO13 | 0.022 | 0.342 | 0.149 | |
| BIO4 | 0.011 | 0.497 | 0.106 | BIO14 | 0.021 | 0.359 | 0.143 | |
| BIO5 | 0.002 | 0.774 | 0.045 | BIO15 | 0.028 | 0.287 | -0.166 | |
| BIO6 | 0.146 | 0.011* | -0.382 | BIO16 | 0.035 | 0.228 | 0.188 | |
| BIO7 | 0.096 | 0.043* | 0.310 | BIO17 | 0.023 | 0.330 | 0.152 | |
| BIO8 | 0.011 | 0.502 | 0.105 | BIO18 | 0.021 | 0.352 | 0.145 | |
| BIO9 | 0.023 | 0.331 | -0.152 | BIO19 | 0.024 | 0.323 | 0.154 | |
| BIO10 | 0.070 | 0.086 | -0.265 | |||||
| *表示显著相关(P < 0.050), * *表示极显著相关(P < 0.010) | ||||||||
如表 6所示, BIO12、BIO13、BIO14、BIO15、BIO16、BIO17、BIO18、BIO19与Shannon-Wiener指数均无显著相关关系(P>0.050)。
3 讨论 3.1 温度因子对物种多样性的影响温差显著影响海南岛热带次生草地丰富度与Shannon-Wiener多样性指数。昼夜温差月均值(BIO2)和气温年较差(BIO7)与海南岛热带次生草地物种多样性显著正相关, 北方部分草地的研究结果也表现出类似特征[33]。可能的原因在于, 温差小的地区, 温度生态位相对较窄[34], 可容纳的植被种类有限;而温差较大的地区需要适应更广泛和多变的环境, 为物种提供更多温度生态位[35], 从而保证丰富的物种多样性[36]。也有研究表明, 环境稳定性可显著促进物种多样性[37], 原因可能是, 长期稳定的气候条件下, 极端事件发生的可能性较低, 从而减少了物种因环境压力而区域灭绝的风险[38]。本研究结果显示(图 3), 等温性(BIO3)显著促进了海南岛热带次生草地物种丰富度。BIO3越大, 温度波动越稳定, 说明在气候稳定的地区物种多样性更高, 反映出热带次生性草地已经对海南长期相对较温暖的气候形成了一定的适应性, 当温度发生较为剧烈的波动时可能对热带草地的生存和恢复产生不利的影响。
年均气温(BIO1)、最冷月份最低温度(BIO6)和最冷季度平均温度(BIO11)与Simpson优势度指数显著负相关, BIO1、BIO6和BIO11的负相关性可能与热带次生草地物种对温度的耐受性有关, 反映了热带次生草地物种对低温的耐受性较差。优越的水热条件为植物生长提供更多能量和资源, 从而显著促进物种多样性[39], 在热带生态系统中, 物种由于长期适应高温环境, 通常对低温具有较强的敏感性, 且极端低温显著降低次生草地的恢复能力[40]。例如, Sandvik等人[41]发现, 高寒草地部分物种因无法适应温度的极端变化, 导致物种多样性降低。因此, 最冷月份和最冷季度较低的温度可能会限制部分物种的生长和分布[42], 进而影响物种多样性。温度季节性变化(BIO4)、最暖月份最高温度(BIO5)、最湿季度平均温度(BIO8)、最干季度平均温度(BIO9)和最暖季度平均温度(BIO10)均与物种多样性没有显著相关性, 原因可能是海南岛气温波动较平稳, 温度季节性变化不明显[43], 且研究区所有样点的BIO5、BIO8和BIO10低于33℃, BIO9高于18℃, 均未达到海南岛的高温和低温下限[44]。
3.2 降水因子对物种多样性的影响本研究发现降水相关因子(BIO12、BIO13、BIO14、BIO15、BIO16、BIO17、BIO18、BIO19)对海南岛热带次生草地物种多样性均无显著影响, 这与北方次生草地物种多样性与气候的研究结果不同, 降水是北方次生草地物种多样性的关键性气候驱动因子。原因可能是:一方面, 南北方次生草地气候与地理条件不同。虽然海南岛热带次生草地主要分布在西北部, 但由于气候和纬度, 年均降水量仍大于1000 mm[28], 降水充沛, 植被对降水因子的敏感度低;而温度因子则会显著影响海南次生草地物种多样性。北方次生草地受温带大陆性气候影响, 年平均降水量在400 mm以下[45], 降水稀缺导致大部分地区常年受干旱困扰, 因此, 降水量的增加能有效减少水分流失[46], 降低蒸散量, 促进植物生长, 产生生态位互补[47], 从而提高物种多样性。另一方面, 南北方次生草地植被的生态习性不同。海南岛热带次生草地主要以喜温暖湿润, 有良好抗涝性的植物为主, 如:三点金(Desmodium triflorum)和铺地黍(Panicum repens)等;但这类植被耐寒性差, 易受低温影响, 相比于降水, 对温度变化较为敏感。北方次生草地长期处于温差大, 降水少的环境中, 其植被类型中耐寒、耐旱、不耐水淹的植物占主导地位[48], 如:羊草(Leymus chinensis)和大针茅(Stipa grandis)等, 对温度变化有较强的适应性, 但是降水能显著缓解干旱, 促进植被生长。
4 结论综上所述, 海南岛热带次生草地物种多样性主要受到温度波动和极端低温的影响, 而降水因子的改变没有使物种多样性发生显著变化。说明海南岛热带次生草地物种多样性已经对热带长期相对较为稳定的高温条件形成了一定的适应性, 但对温度的剧烈波动的适应性较差, 尤其是对低温较为不适应, 低温条件(尤其发生极端低温时)对次生草地的物种多样性将产生明显的负反馈影响。研究丰富了我国南方热带次生草地物种多样性及气候相关研究结果, 为未来全球气候变化背景下热带草地生物多样性保护与管理及恢复提供了科技支撑。
| [1] |
Xu X, Niu S L, Sherry R A, Zhou X H, Zhou J Z, Luo Y Q. Interannual variability in responses of belowground net primary productivity (NPP) and NPP partitioning to long-term warming and clipping in a tallgrass prairie. Global Change Biology, 2012, 18(5): 1648-1656. DOI:10.1111/j.1365-2486.2012.02651.x |
| [2] |
戚超, 刘晓, 闫艺兰, 袁国富. 不同降水年型黄土高原半干旱撂荒草地水分收支特征. 水土保持研究, 2019, 26(1): 106-112. |
| [3] |
姚强, 赵成章, 郝青, 董小刚. 旱泉沟流域次生植被不同恢复阶段的多样性特征. 生态环境学报, 2010, 19(4): 849-852. |
| [4] |
Nerlekar A N, Veldman J W. High plant diversity and slow assembly of old-growth grasslands. Proceedings of the National Academy of Sciences of the United States of America, 2020, 117(31): 18550-18556. |
| [5] |
Klimek S, Richter gen Kemmermann A, Hofmann M, Isselstein J. Plant species richness and composition in managed grasslands: the relative importance of field management and environmental factors. Biological Conservation, 2007, 134(4): 559-570. DOI:10.1016/j.biocon.2006.09.007 |
| [6] |
孙成明, 刘涛, 陈雯, 陈瑛瑛, 田婷, 李建龙. 南方草地NPP的水热估算模型及其应用. 生态环境学报, 2013, 22(9): 1457-1461. |
| [7] |
Yao Z Y, Xin Y, Yang L, Zhao L Q, Ali A. Precipitation and temperature regulate species diversity, plant coverage and aboveground biomass through opposing mechanisms in large-scale grasslands. Frontiers in Plant Science, 2022, 13: 999636. DOI:10.3389/fpls.2022.999636 |
| [8] |
贾鹏. 青藏高原高寒草地群落物种多样性、生产力及其影响因素的研究[D]. 兰州: 兰州大学, 2019.
|
| [9] |
Klopfer P H. Environmental determinants of faunal diversity. The American Naturalist, 1959, 93(873): 337-342. DOI:10.1086/282092 |
| [10] |
Theobald D M, Harrison-Atlas D, Monahan W B, Albano C M. Ecologically-relevant maps of landforms and physiographic diversity for climate adaptation planning. PLoS One, 2015, 10(12): e0143619. DOI:10.1371/journal.pone.0143619 |
| [11] |
Bita C E, Gerats T. Plant tolerance to high temperature in a changing environment: scientific fundamentals and production of heat stress-tolerant crops. Frontiers in Plant Science, 2013, 4: 273. |
| [12] |
Reynaert S, De Boeck H J, Verbruggen E, Verlinden M, Flowers N, Nijs I. Risk of short-term biodiversity loss under more persistent precipitation regimes. Global Change Biology, 2021, 27(8): 1614-1626. DOI:10.1111/gcb.15501 |
| [13] |
Pérez-Navarro M Á, Serra-Diaz J M, Svenning J C, Esteve-Selma M Á, Hernández-Bastida J, Lloret F. Extreme drought reduces climatic disequilibrium in dryland plant communities. Oikos, 2021, 130(5): 680-690. DOI:10.1111/oik.07882 |
| [14] |
Korell L, Auge H, Chase J M, Harpole W S, Knight T M. Responses of plant diversity to precipitation change are strongest at local spatial scales and in drylands. Nature Communications, 2021, 12: 2489. DOI:10.1038/s41467-021-22766-0 |
| [15] |
Yang H J, Wu M Y, Liu W X, Zhang Z, Zhang N L, Wan S Q. Community structure and composition in response to climate change in a temperate steppe. Global Change Biology, 2011, 17(1): 452-465. DOI:10.1111/j.1365-2486.2010.02253.x |
| [16] |
Tabares X, Mapani B, Blaum N, Herzschuh U. Composition and diversity of vegetation and pollen spectra along gradients of grazing intensity and precipitation in southern Africa. Review of Palaeobotany and Palynology, 2018, 253: 88-100. DOI:10.1016/j.revpalbo.2018.04.004 |
| [17] |
Frank D, Reichstein M, Bahn M, Thonicke K, Frank D, Mahecha M D, Smith P, van der Velde M, Vicca S, Babst F, Beer C, Buchmann N, Canadell J G, Ciais P, Cramer W, Ibrom A, Miglietta F, Poulter B, Rammig A, Seneviratne S I, Walz A, Wattenbach M, Zavala M A, Zscheischler J. Effects of climate extremes on the terrestrial carbon cycle: concepts, processes and potential future impacts. Global Change Biology, 2015, 21(8): 2861-2880. DOI:10.1111/gcb.12916 |
| [18] |
Bai X H, Zhao W W, Wang J, Santos Ferreira C S. Precipitation drives the floristic composition and diversity of temperate grasslands in China. Global Ecology and Conservation, 2021, 32: e01933. DOI:10.1016/j.gecco.2021.e01933 |
| [19] |
干珠扎布, 段敏杰, 郭亚奇, 张伟娜, 梁艳, 高清竹, 旦久罗布, 白玛玉珍, 西绕卓玛. 喷灌对藏北高寒草地生产力和物种多样性的影响. 生态学报, 2015, 35(22): 7485-7493. |
| [20] |
胡海英, 李惠霞, 倪彪, 师斌, 许冬梅, 谢应忠. 宁夏荒漠草原典型群落的植被特征及其优势植物的水分利用效率. 浙江大学学报: 农业与生命科学版, 2019, 45(4): 460-471. |
| [21] |
李想, 于红博, 刘月璇, 常帅. 锡林郭勒不同草原类型群落生物量及多样性研究. 草地学报, 2022, 30(1): 196-204. |
| [22] |
费诚, 董乙强, 安沙舟. 新疆北疆荒漠群落植被的物种多样性及影响因素研究. 中国草地学报, 2023, 45(7): 49-59. |
| [23] |
Robertson T R, Zak J C, Tissue D T. Precipitation magnitude and timing differentially affect species richness and plant density in the sotol grassland of the Chihuahuan Desert. Oecologia, 2010, 162(1): 185-197. DOI:10.1007/s00442-009-1449-z |
| [24] |
李敏菲, 马煜成, 刘耘华, 盛建东, 程军回. 新疆草地群落谱系多样性变化特征及影响因素. 生态学报, 2020, 40(7): 2285-2299. |
| [25] |
Tian F P, Zhang Z N, Chang X F, Sun L, Wei X H, Wu G L. Effects of biotic and abiotic factors on soil organic carbon in semi-arid grassland. Journal of Soil Science and Plant Nutrition, 2016, 16(4): 1087-1096. |
| [26] |
谭谌淼. 海南省草地资源调查[D]. 海口: 海南大学, 2020.
|
| [27] |
陈德祥, 李意德, Heping Liu, 许涵, 肖文发, 骆土寿, 周璋, 林明献. 尖峰岭热带山地雨林生物量及碳库动态. 中国科学: 生命科学, 2010, 40(7): 596-609. |
| [28] |
唐少霞, 赵志忠, 毕华, 谢跟踪. 海南岛气候资源特征及其开发利用. 海南师范大学学报: 自然科学版, 2008, 21(3): 343-346. |
| [29] |
周圻. 海南岛热带草地饲料资源浅析. 饲料研究, 2003(6): 24-26. |
| [30] |
杨小波, 陈宗铸, 李东海, 陈玉凯, 陈辉, 林泽钦, 王力军, 丁琼, 李意德, 莫燕妮, 周亚东, 洪小江, 王世力, 邓勤, 卢刚, 龙文兴, 黄瑾, 岑举人, 车秀芬, 欧芷阳, 郭涛, 黄运峰, 罗涛, 党金玲, 叶凡, 杨立荣, 徐中亮, 农寿千, 吕晓波, 张孟文, 张彩凤, 吕洁杰, 罗召美, 汪小平, 龙成, 周威, 卜广发, 万春红, 杨琦, 周文嵩, 陶楚, 冯丹丹, 罗文启, 邢莎莎, 李英英, 张育霞, 张凯, 李苑菱, 周双清, 周婧, 张萱蓉, 李丹, 吴庭天, 刘子金, 熊梦辉, 赵瑞白, 苏凡, 李嘉吴, 黄绪壮. 海南植被志-第一卷. 北京: 科学出版社, 2019.
|
| [31] |
中国南方草地牧草资源调查项目组. 中国南方草地牧草资源调查执行规范: 2017-2022. 北京: 科学出版社, 2019.
|
| [32] |
马克平, 刘玉明. 生物群落多样性的测度方法Ⅰα多样性的测度方法(下). 生物多样性, 1994, 2(4): 231-239. |
| [33] |
Peng Y, Feng J C, Sang W G, Axmacher J C. Geographical divergence of species richness and local homogenization of plant assemblages due to climate change in grasslands. Biodiversity and Conservation, 2022, 31(3): 797-810. DOI:10.1007/s10531-022-02364-2 |
| [34] |
Kolomiytsev N, University C S, Poddubnaya N, University C S. Temporal and spatial variability of environments drive the patterns of species richness along latitudinal, elevational, and depth gradients. Biological Communications, 2018, 63(3): 189-201. DOI:10.21638/spbu03.2018.305 |
| [35] |
Schippers P, Ozinga W A, Pouwels R. Factors affecting functional diversity of grassland vegetations. Ecological Modelling, 2022, 472: 110078. DOI:10.1016/j.ecolmodel.2022.110078 |
| [36] |
张全国, 张大勇. 生物多样性与生态系统功能: 进展与争论. 生物多样性, 2002, 10(1): 49-60. |
| [37] |
Liang M W, Baiser B, Hallett L M, Hautier Y, Jiang L, Loreau M, Record S, Sokol E R, Zarnetske P L, Wang S P. Consistent stabilizing effects of plant diversity across spatial scales and climatic gradients. Nature Ecology & Evolution, 2022, 6: 1669-1675. |
| [38] |
Mathes G H, van Dijk J, Kiessling W, Steinbauer M J. Extinction risk controlled by interaction of long-term and short-term climate change. Nature Ecology & Evolution, 2021, 5: 304-310. |
| [39] |
Schumm M, Edie S M, Collins K S, Gómez-Bahamón V, Supriya K, White A E, Price T D, Jablonski D. Common latitudinal gradients in functional richness and functional evenness across marine and terrestrial systems. Proceedings Biological Sciences, 2019, 286(1908): 20190745. |
| [40] |
Harrison S P, Gornish E S, Copeland S. Climate-driven diversity loss in a grassland community. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(28): 8672-8677. |
| [41] |
Sandvik S M, Heegaard E, Elven R, Vandvik V. Responses of alpine snowbed vegetation to long-term experimental warming. Écoscience, 2004, 11(2): 150-159. DOI:10.1080/11956860.2004.11682819 |
| [42] |
Catford J A, Dwyer J M, Palma E, Cowles J M, Tilman D. Community diversity outweighs effect of warming on plant colonization. Global Change Biology, 2020, 26(5): 3079-3090. DOI:10.1111/gcb.15017 |
| [43] |
Abernethy K, Bush E R, Forget P M, Mendoza I, Morellato L P C. Current issues in tropical phenology: a synthesis. Biotropica, 2018, 50(3): 477-482. DOI:10.1111/btp.12558 |
| [44] |
杨德军, 罗勇, 邱琼, 陈勇, 钟萍, 段宗亮, 张快富. 西双版纳采矿区植被恢复中的树种选择研究. 贵州林业科技, 2015, 43(1): 4-9. |
| [45] |
张仁平, 冯琦胜, 郭靖, 尚振艳, 梁天刚. 2000—2012年中国北方草地NDVI和气候因子时空变化. 中国沙漠, 2015, 35(5): 1403-1412. |
| [46] |
于静, 高亚敏, 付铭. 科尔沁草原NDVI时空变化特征及其对气候的响应. 中国草地学报, 2020, 42(6): 82-90. |
| [47] |
Bunting E L, Munson S M, Villarreal M L. Climate legacy and lag effects on dryland plant communities in the southwestern U.S. Ecological Indicators, 2017, 74: 216-229. DOI:10.1016/j.ecolind.2016.10.024 |
| [48] |
钟悦鸣, 王健铭, 张天汉, 李景文, 冯益明, 卢琦. 中国青藏高原北部戈壁区种子植物物种组成及其区系特征. 植物科学学报, 2017, 35(4): 525-533. |