 2024, Vol. 44
2024, Vol. 44文章信息
- 白昱欣, 刘润洪, 苏洁桦, 申卫军
- BAI Yuxin, LIU Runhong, SU Jiehua, SHEN Weijun
- 树种混交对马尾松和红锥根际与非根际土壤微生物资源限制的影响
- Effects of tree species mixing on bulk and rhizosphere soil microbial resource limitation in stands of Pinus massoniana and Castanopsis hystrix
- 生态学报. 2024, 44(23): 10770-10781
- Acta Ecologica Sinica. 2024, 44(23): 10770-10781
- http://dx.doi.org/10.20103/j.stxb.202310312359
-
文章历史
- 收稿日期: 2023-10-31
- 采用日期: 2024-10-10





2. 亚热带农业生物资源与保护利用国家重点实验室, 南宁 530004;
3. 内蒙古自治区环境监测总站呼和浩特分站, 呼和浩特 010030
2. State Key Laboratory for Conservation and Utilization of Agro-bioresources, Nanning 530004, China;
3. Inner Mongolia Environmental Monitoring Station Hohhot Branch, Hohhot 010030, China
混交林因具有提高土壤养分有效性, 增强人工林生态系统功能与服务等诸多优越于纯林的优势而成为全球广泛推广的造林模式[1—4]。与纯林相比, 混交林可以通过改变凋落物或者根系分泌物的数量和质量等途径而影响微生物群落的组成和活性, 进而影响人工林生态系统碳(C)及氮(N)和磷(P)养分循环过程[2, 5—7]。微生物会根据自身所受到养分资源(如C、N、P)的限制, 通过产生胞外酶来获取受限制的营养元素。因此, 土壤胞外酶的化学计量比可以表征土壤养分的有效性以及土壤微生物在特定的资源投资过程(如C、N、P养分的矿化)中所受资源限制的状况[8—11]。通过测定土壤胞外酶活性、计算其化学计量比来探究树种混交对土壤微生物的资源限制状况的影响, 进而指导人工林的树种选择和养分管理等具有重要理论和实践意义。
目前, 国内外学者已经针对树种混交对土壤胞外酶活性及其化学计量比的影响开展了一些研究。Wu等人[12]通过研究多树种混交对亚热带马尾松人工林土壤胞外酶活性化学计量的影响发现, 在混交林的土壤中, 与C获取相关的水解酶活性降低, 但酚氧化酶活性却增加了, 阔叶树种与马尾松的混交抑制了土壤中不稳定碳的分解, 却促进了土壤中顽固性碳的分解, 微生物需要通过改变C获取酶的活性来应对C限制, 表明树种的混交能够刺激微生物快速调节其分解代谢和合成代谢的能力, 从而影响了地下碳储量[13]。此外, Sheng等人[13]的研究表明, 由于与油松混交增加了刺槐和沙棘根际胁迫, 因此过氧化氢酶的活性增加, 但是却降低了沙棘根际土壤中脱氢酶的活性, 作为参与土壤有机质矿化的酶, 脱氢酶活性的降低表明与油松混交对沙棘根际区域养分的矿化和养分释放有一定的抑制作用, 养分释放受到抑制, 土壤中微生物受到的C源限制也会响应的加剧。研究表明, 混交林中, 由于地下根系生物量和根系分泌的改变可能会通过影响土壤中酶的活性, 从而对土壤中P的矿化过程产生影响, 而当真菌参与土壤有机质的分解过程时, 可增加土壤中P的释放, 增加土壤中P的可利用性[2, 13], 这一系列的过程可能使得使土壤中的P限制得到缓解。
由此可见, 酶种类的差异、混交树种的不同以及林龄和土壤状况等诸多因素会影响最终的研究结论;究其原因, 研究结论的差异可能是土壤养分有效性和pH值等环境因素的变化所导致的[2, 7, 13—14]。构建同龄混交和异龄混交林是林业生产实践中两种主要混交模式, 二者的树龄、林分结构等方面存在明显差异[15]。先前关于树种混交对酶活性及其化学计量比的研究大多集中在异龄混交林, 尚不清楚同龄和异龄混交林对土壤酶活性及其化学计量比的影响之间是否存在差异。因此仍需要针对不同类型的混交林进行深入研究, 以揭示其普适性规律。
根际区域是受植物根系活动影响最为强烈的土壤微环境, 也是物质循环和能量流动最活跃的区域, 在森林生态系统的养分循环过程中发挥着至关重要作用[16—17]。尽管已经有大量研究表明树种混交会改变土壤微生物资源限制状况, 但遗憾的是, 大多数研究都将土壤视为一个整体, 忽略了根际和非根际土壤微生物代谢活性对树种混交的差异化响应。研究表明, 根际土壤的物理、化学以及生物学特性, 如土壤C、N和P含量, 微生物群落以及参与营养物质循环的酶的活性等明显不同于非根际土壤[18—20]。阐明树种混交对根际和非根际土壤酶活性及其化学计量特征的影响, 对于理解树种混交对根际和非根际土壤微生物资源限制状况和养分循环过程具有重要意义。
马尾松(Pinus massoniana)作为我国亚热带地区最主要的用材林树种及荒山造林的先锋树种, 因其具有速生丰产、材质较佳、用途广泛、耐旱耐瘠、适应性强等特性而在我国亚热带地区被广泛种植, 并给人们带来了巨大的经济价值与生态效益。然而, 我国60%的马尾松人工林为纯林, 长期连栽和大面积纯林经营导致生物多样性降低、病虫害频发、养分周转缓慢、地力衰退、生产力下降, 生态系统稳定性差, 生态系统功能和服务退化等一系列生态环境问题, 严重阻碍了马尾松人工林的多目标可持续经营[21]。通过引入阔叶树种, 如:红锥(Castanopsis hystrix)等对马尾松纯林进行近自然化改造, 已成为增强其生态系统功能并实现其可持续发展的有效方法[22—23]。然而, 马尾松人工纯林引入红锥形成针阔混交林后, 其凋落物的数量和质量、根系生物量和根系分泌物等均会产生不同程度的变化, 这些改变可能会对其根际和非根际的土壤胞外酶活性及其化学计量特征产生影响。鉴于此, 本研究提出, 以往的研究表明树种混交通常能够改善土壤养分条件, 那么树种混交将如何影响土壤中微生物的资源限制状况, 其限制程度是加剧还是有所缓解?产生影响的主要驱动因素是什么?除此之外, 对于土壤微生物所受到的C限制和P限制, 在根际区域和非根际区域是否存在差异?针对以上科学问题, 本研究以马尾松-红锥混交林及其对应的纯林作为研究对象, 通过测定5种不同林分类型根际和非根际土壤理化性质、酶活性, 并通过土壤酶活性计算化学计量比, 探究马尾松与红锥混交对土壤微生物资源限制状况的影响在非根际土和根际土之间的差异, 并结合土壤理化因子的变化辨识产生影响的主要驱动因素。基于上述研究背景, 本研究提出以下科学假设:(1)马尾松与红锥混交能够通过改善土壤养分状况来影响土壤酶活性, 从而改变微生物的资源限制状况, 加剧微生物的C限制, 缓解P限制;(2)与非根际土相比, 根际土壤中微生物受到的C限制更强, 但微生物受到混交的影响在同龄混交林和异龄混交林之间存在差异。本研究结果能够从酶活性及其化学计量比的角度揭示不同的混交类型对土壤养分可利用性的影响及其调控机制, 可为马尾松和红锥人工混交林的配置模式以及养分管理等提供重要的科学依据。
1 材料与方法 1.1 研究区概况研究样地位于广西壮族自治区凭祥市中国林业科学研究院热带林业实验中心伏波试验场(106°51′—106°53′ E, 22°02′—22°04′ N, 海拔430—680 m), 地貌类型以低山丘陵为主, 地形自西而东倾斜。该地区属于南亚热带季风型半湿润-湿润气候, 年均气温20.5—21.7℃, 最热月(7月)平均气温约为27.6℃, 最冷月(1月)平均气温约为13.5℃, ≥10℃积温为6000—7600 ℃;年降雨量为1200—1500 mm, 降雨量年际间波动较大, 干季(10月至翌年3月)和湿季(4—9月)明显;年蒸发量1261—1388 mm, 相对湿度为80%—84%。土壤类型主要是由花岗岩发育而形成的砖红壤和红壤, 属酸性土, 土层厚度超过1 m。该地区的地带性植被为热带季雨林和南亚热带常绿阔叶林, 由于人类活动的长期干扰, 原生植被几乎无存, 现已被各种类型的人工林所替代。20世纪50年代在原生植被的皆伐迹地上营造杉木和马尾松等树种的人工纯林。为了改造大面积的杉木和马尾松人工纯林并探索珍贵乡土阔叶树种大径材培育模式, 1983年对部分马尾松纯林进行间伐并引入红锥形成针阔异龄混交林。此外, 1983年在杉木人工林皆伐迹地上分别营造了混交比例为1 ∶ 1的马尾松与红锥混交林、马尾松纯林和红锥纯林。
1.2 样地设置本研究选取1959年营造的马尾松纯林(P59)、1983年营造的马尾松纯林(P83)、1983年种植的红锥纯林(C83)、1959年马尾松-1983红锥异龄混交林(UMF)以及1983年马尾松-红锥同龄混交林(EMF)作为研究对象。于2021年7月, 在上述5个林分类型中各设置3个20 m × 20 m的样方, 各样方之间间隔20 m以上, 共建立15个样方。对样方内所有胸径(DBH)≥ 5 cm的植株进行调查, 记录种名、胸径、高度、枝下高等信息, 并记录每个样方的海拔、坡向、坡位、坡度、郁闭度等生境信息。根据植物群落调查的结果, 按照平均树高(H)和平均胸径(D)且干型中等选择标准木, 用于采集根际土壤, 纯林中选择6棵标准木, 混交林中马尾松和红锥各选择6棵标准木。
1.3 样品采集于2022年1月对非根际土和根际土进行采集。具体而言, 非根际土壤样品采用“五点法”采集, 即在每个样方的4个顶点和中心位置, 去除地表的凋落物和腐殖质后, 利用直径5 cm的土钻钻取0—40 cm土层的土壤样品, 同一样方内5个采样点的土壤样品运用“四分法”混合均匀成为1份土样带回实验室。同时, 在采集土壤样品的附近位置利用环刀(体积100 cm3)采集原状土壤, 用于测定土壤容重和土壤含水量。对于根际土壤, 在清除标准木树干基部的地表凋落物和腐殖质后, 挖掘树干基部的土壤(0—40 cm)以露出侧根, 追踪其至树干基部以识别其是否为标准木的根系;若为标准木的根系则顺着侧根追踪其分支的下一级根系, 直至根系末端, 利用“抖根法”收集距细根(直径<2 mm)表面5 mm以内的土壤作为根际土样品。将采集好的非根际以及根际土壤样品装入做好标记的无菌自封袋中, 放入装有干冰的保温箱中低温保存, 立即带回实验室, 剔除其中的根系、石砾和动植物残体等杂质, 过2 mm土壤筛后分成2份, 其中1份置于4 ℃冰箱保存, 用于土壤硝态氮、铵态氮和土壤胞外酶活性指标的测定, 1份置于室内阴凉处风干, 称取部分样品用于土壤pH值测定, 其他样品研磨后过0.149 mm土壤筛用于土壤有机质、总氮、总磷和有效磷等土壤化学性质的测定。
1.4 土壤样品测定 1.4.1 土壤理化性质测定土壤容重通过环刀法进行测定;土壤含水量采用烘干法测定, 称取10 g新鲜土壤放入铝盒中, 将其置于105 ℃烘箱中烘干至恒重后称量干土质量。将土壤样品与1 g/mol KCl溶液按照土水比为1 ∶ 2.5混合, 搅拌静置30min后。使用pH计(FE28-Standard, 梅特勒)测定pH值[24]。其余土壤化学性质均参考《土壤农化分析》[25]的方法进行测定, 其中, 土壤有机质(SOM)使用重铬酸钾容量法测定;土壤全氮(TN)采用凯氏定氮法测定, 土壤经过浓H2SO4消煮后, 利用凯氏定氮仪(K9860, 海能)进行测定;将新鲜土壤与0.01 mol/L CaCl2溶液混合后震荡2 h, 提取液分装后利用连续流动化学分析仪(Auto Analyzer 3, 德国, SEAL)测定土壤硝态氮(NO3--N)和铵态氮(NH4+-N);土壤总磷(TP)使用用H2SO4-HClO4消煮, 土壤有效磷(AP)采用HCl-H2SO4浸提, 均使用钼锑抗比色法显色, 最后利用多功能酶标仪(Infinite M200 Pro, TECAN), 在波长880 nm(TP)和882 nm(AP)处测定吸光值。
1.4.2 土壤水解酶活性测定参考Bell等[26]的研究方法, 采用荧光标记底物法测定5种参与C、N和P获取的水解酶活性, 即α-1, 4-葡萄糖苷酶(AG)、β-1, 4-葡萄糖苷酶(BG)、β-1, 4-N-乙酰葡糖胺酶(NAG)、亮氨酸氨基肽酶(LAP)和酸性磷酸酶(ACP), 其功能及所用的反应底物等信息如表 1所示。具体方法如下:称取1 g鲜土放入烧杯中, 加入125 mL醋酸钠缓冲溶液(50 mmol/L, pH=5.0), 在涡旋振荡器上震荡1 min, 制成土壤均质悬浮液。利用移液枪向96孔微孔板的每个孔内加入200 μL土壤悬浮液和50 μL的相应底物(200 μmol/L, 超纯水配制)。以同样的方式制备样品对照(200 μL土壤悬浮+50μL乙酸盐缓冲液)、底物对照(200 μL乙酸盐缓冲液+50 μL底物)和荧光标准(200 μL乙酸盐缓冲液+50μL标准溶液)。所有微孔板在25 ℃的黑暗条件下培养2 h, 培养结束后加入10 μL NaOH(1g/mol)终止反应, 并使用多功能酶标仪(Infinite M200 Pro, TECAN), 在365 nm的激发波长和450 nm的发射波长条件下测定用于计算酶活性的荧光值, 单位为nmol g-1 h-1。
| 酶的类型Enzyme type | 酶名称(缩写)Enzyme (Abbreviation) | 功能Function | 底物Substrate |
| C循环水解酶 | α-1, 4-葡萄糖苷酶(AG) | 可溶性糖降解 | 4-MUB-α-D-吡喃葡萄糖苷 |
| C-cycling hydrolytic enzymes | β-1, 4-葡萄糖苷酶(BG) | 纤维素降解 | 4-MUB-β-D-吡喃葡萄糖苷 |
| N循环水解酶 | β-N-乙酰葡糖胺酶(NAG) | 甲壳素和肽聚糖降解 | 4-MUB-N-乙酰基-β-D-氨基葡萄糖 |
| N-cycling hydrolytic enzymes | 亮氨酸氨基肽酶(LAP) | 蛋白质和肽的降解 | L-亮氨酸-7-酰胺基-4-甲基香豆素 |
| P循环水解酶P-cycling hydrolytic enzymes | 酸性磷酸酶(ACP) | 催化磷酸单酯水解 | 4-MUB-磷酸盐 |
| MUB:4-甲基伞形酮4-Methylumbelliferone;MUC:7-氨基-4-甲基香豆素7-Amino-4-methylcoumarin | |||
参考Moorhead等[8, 27]的方法, 利用以下公式计算酶活性向量的长度(Length)和角度(Angle)。
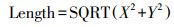
|
(1) |
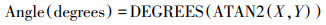
|
(2) |
式中X代表(AG+BG)/(AG+BG+ACP), Y代表(AG+BG)/(AG+BG+NAG+LAP), 向量的长度表示土壤微生物受土壤C限制的程度, 向量长度越长, 表示土壤微生物受C限制的程度越大。向量的角度大小表示土壤微生物受土壤N、P的限制程度, 角度<45°表示土壤微生物受到N限制, 角度越小, N限制越强;角度>45°则表示土壤微生物受P限制, 角度越大, P限制越强。
1.5 数据处理本研究所有数据统计分析与制图均在SPSS 25.0(IBM SPSS Inc, Chicago, USA)和R v4.2.1(https://www.r-project.org/)软件中完成。其中, 利用SPSS软件进行单因素方差分析(One-way ANOVA), 检验不同林分类型和微生境(非根际土和根际土)之间土壤理化性质、土壤酶活性及其化学计量比是否存在显著差异, 随后运用Tukey HSD方法进行多重比较, 确定哪些林分类型之间存在显著差异, 显著性水平设置为P<0.05。此外, 使用R语言“vegan”程序包中的adonis函数进行置换多元方差分析(PERMANOVA), 检验林分类型和生境的变化对于土壤酶活性及其化学计量比差异的贡献;基于R语言“linkET”程序包中的Mantel检验评估土壤理化性质与土壤酶活性及其化学计量比之间的Spearman相关性。数据的可视化均通过R语言“ggplot2”程序包实现。
2 结果与分析 2.1 不同林分类型和微生境土壤理化性质特征单因素方差分析结果表明(表 2), 土壤理化性质在不同林分类之间差异显著(P<0.05)。同龄混交林的非根际土壤土壤含水量显著高于马尾松和红锥纯林, 异龄混交林土壤含水量显著低于红锥纯林, 与马尾松纯林间无显著差异。土壤pH值在针叶林和混交林之间无显著差异, 红锥纯林的土壤pH值显著高于其他林分。除AP以外, 非根际土壤中其余养分含量在同龄混交林中均显著高于异龄混交林。在根际土中, 异龄混交对马尾松根际土养分含量影响不显著, 但同龄混交显著增加了马尾松根际土SOM、TN、TP和NH4+-N的含量。与马尾松纯林相比, 混交能够增加土壤养分含量, 且同龄混交比异龄混交更有利于土壤养分的积累。
| 微生境Microhabitat | 林分类型Stand types | 容重Bulk density/(g/cm3) | 含水量Soil water content/% | pH | 有机质SOM/(g/kg) | 总氮TN/(g/kg) | 总磷TP/(g/kg) | 铵态氮NH4+-N/(mg/kg) | 硝态氮NO3--N/(mg/kg) | 有效磷AP/(mg/kg) |
| 非根际土Bulk soil | UMF | 1.40a(0.06) | 21.95a(1.00) | 3.51Ba(0.04) | 27.62Aab(1.71) | 1.17Aab(0.05) | 0.23Ab(0.02) | 6.85Aab(1.15) | 1.91Ab(0.60) | 2.04Ac(0.10) |
| P59 | 1.45a(0.06) | 22.21a(1.43) | 3.53Ba(0.04) | 25.35Aa(0.67) | 1.04Aa(0.03) | 0.21Aa(0.02) | 6.62Aa(1.16) | 1.64Aab(0.30) | 1.97Abc(0.21) | |
| C83 | 1.41a(0.06) | 23.60b(1.76) | 3.68Bc(0.03) | 29.63Ab(1.79) | 1.29Abc(0.10) | 0.23Ab(0.03) | 8.28Ab(1.36) | 1.15Aa(0.20) | 1.76Aa(0.07) | |
| P83 | 1.42a(0.07) | 21.06a(0.90) | 3.58Bab(0.03) | 32.70Ac(4.74) | 1.29Aab(0.12) | 0.21Aa(0.01) | 7.85Aab(0.73) | 2.10Ab(0.76) | 1.86Aab(0.19) | |
| EMF | 1.41a(0.05) | 26.98c(2.32) | 3.64Cbc(0.02) | 36.61Ad(3.32) | 1.47Ac(0.09) | 0.27Ac(0.01) | 11.30Ac(2.25) | 2.67Bc(0.57) | 1.82Aab(0.11) | |
| 根际土Rhizosphere soil | UMC | - | - | 3.17Aab(0.10) | 63.30Ba(16.25) | 2.23Ba(2.66) | 0.27Aa(0.03) | 18.87Ba(3.23) | 3.73Bc(0.32) | 1.92Aa(0.42) |
| UMP | - | - | 3.16Aab(0.07) | 52.67Ba(9.38) | 1.75ABa(0.28) | 0.26Aa(0.02) | 23.46Cbc(2.29) | 3.15Bbc(0.26) | 1.78Aa(0.08) | |
| P59 | - | - | 3.34Abc(0.05) | 52.55Ba(3.74) | 1.79Ba(0.05) | 0.25Ba(0.03) | 27.18Bb(0.50) | 2.83Abc(0.71) | 2.26Aab(0.10) | |
| C83 | - | - | 3.40Ac(0.05) | 56.52Ba(7.36) | 2.29Ba(0.45) | 0.27Aa(0.05) | 30.31Bb(3.41) | 1.07Aa(0.14) | 2.11Aab(0.37) | |
| P83 | - | - | 3.30Abc(0.13) | 52.61Ba(9.94) | 1.76Aa(0.33) | 0.21Aa(0.02) | 16.92Ba(2.48) | 1.31Aa(0.39) | 2.15Aab(0.44) | |
| EMP | - | - | 3.25Babc(0.09) | 86.20Bb(11.52) | 3.37Bb(0.60) | 0.34Bb(0.03) | 27.84Bb(4.51) | 1.11Aa(0.44) | 2.99Bbc(0.74) | |
| EMC | - | - | 3.07Aa(0.04) | 94.95Bb(1.04) | 3.71Bb(0.10) | 0.35Bb(0.02) | 38.18Cc(5.56) | 2.35Bb(0.39) | 3.44Bc(0.39) | |
| 表中数据为平均值(标准差), n=3;同一列中不同小写字母表示该变量在不同林分类型之间存在显著差异(P<0.05);同一列中不同大写字母表示该变量在不同微生境之间存在显著差异(P<0.05);缩写:UMF表示马尾松-红锥异龄混交林;EMF表示马尾松-红锥同龄混交林;P59和P83分别表示1959年和1983年营造的马尾松纯林;C83表示1983年营造的红锥纯林;MP表示对应混交林中的马尾松;MC表示对应混交林中的红锥; -:无数据 | ||||||||||
对比不同微生境的土壤理化性质(表 2), 与非根际土相比, 根际土的pH值更低且养分含量更高, 但在不同的林分类型之间有一定差异。SOM、TN和NH4+-N在所有林分的根际土中均显著高于非根际土, TP在马尾松纯林和同龄混交林的根际土中显著高于非根际土, NO3--N在异龄混交林的根际土中显著高于非根际土, AP在同龄混交林的根际土中显著高于非根际土。
2.2 不同林分类型和微生境土壤酶活性及其资源限制特征土壤酶活性在林分类型和微生境之间均存在显著差异, 但差异的显著性因土壤水解酶的种类而异(图 1)。具体而言, AG、BG和NAG酶活性在根际土中均显著高于非根际土;LAP酶活性在马尾松纯林(P83)根际土中显著高于非根际土, 但在其他林分类型的根际土中显著低于非根际土;ACP酶活性在同龄混交林和纯林根际土中均显著高于非根际土, 但在异龄混交林根际土中显著低于非根际土(P<0.05)。同龄混交林非根际土中的BG和NAG酶活性显著低于异龄混交林, LAP和ACP酶活性显著高于异龄混交林(P<0.05);除LAP酶以外, 根际土中其余土壤酶的活性受混交的影响均较大, 同龄混交林根际土的酶活性均显著高于异龄混交林(P<0.05)。

|
| 图 1 不同林分类型及生境土壤酶活性 Fig. 1 Soil extracellular enzyme activities in bulk and rhizosphere soils of different forest types 不同小写字母表示林分类型之间有显著性差异, 不同大写字母表示生境之间的酶活性有显著差异, *表示同龄混交和异龄混交之间有显著差异(P<0.05);MF代表混交林, PP代表马尾松纯林, PC代表红锥纯林, MP代表混交林中的马尾松, MC代表混交林中的红锥 |
根据土壤酶活性计算其化学计量比, 结果表明土壤酶活性向量的长度和角度在不同林分类型之间存在显著差异(图 2)。混交类型对土壤中微生物养分限制的影响因树种和土壤生境的不同而产生差异, 相对于马尾松纯林, 无论是同龄混交还是异龄混交, 均显著降低了非根际土壤的P限制(P<0.05;图 2)。同龄混交显著降低了根际土的P限制, 而异龄混交林中, 仅红锥根际土的P限制程度显著降低。非根际土的P限制程度在同龄混交林中显著高于异龄混交林, C限制程度没有显著差异;P限制在同龄混交林的马尾松根际土中显著低于异龄混交林, 但在红锥根际土中显著高于异龄混交林, C限制程度在同龄混交林根际土中显著低于异龄混交林(P<0.05;图 2)。对比不同的土壤生境, 根际土中P限制显著低于非根际土, 但C限制显著增强(P<0.05;图 2)。总体来说, 混交林非根际土的C限制强度显著高对应纯林, 但根际土中C限制程度显著低于纯林, 该规律在同龄及异龄混交林中均有体现。根据向量角度所表征的养分限制情况, 在所有林分类型的土壤中, 微生物都受到强烈的P限制, 且该规律不受土壤生境变化的影响。

|
| 图 2 土壤细胞外酶化学计量分析比较相对氮/磷和碳限制 Fig. 2 Comparison of relative nitrogen/phosphorus and carbon limitations based on the analysis of soil extracellular enzyme stoichiometry 向量角度图中,虚线表示45°阈值,角度<45°表示微生物受到N限制,反之则表示微生物受到P限制 |
置换多元方差分析结果(表 3)显示, AG和BG活性的变异在同龄混交组和异龄混交组中均主要由林分类型的变化来解释(同龄混交:AG:R2=0.568, BG:R2=0.519, 异龄混交:AG:R2=0.712, BG:R2=0.521;P<0.001), NAG和LAP活性的变异在同龄混交林组中主要由林分类型的变化来解释(NAG:R2=0.634, LAP:R2=0.356;P<0.001), 但在异龄混交林组中则主要由林分类型和微生境变化的交互作用解释(NAG:R2=0.496, LAP:R2=0.906;P<0.001), ACP活性的变异在同龄混交林组中主要由微生境的变化来解释(R2=0.201;P<0.001), 但在异龄混交林组中则主要由林分类型的变化解释(R2=0.737;P<0.001)。向量长度(C限制)的变异在同龄混交组主要由林分类型的变化解释(R2=0.749;P<0.001), 在异龄混交组则主要由林分类型和微生境变化的交互作用解释(R2=0.540;P<0.001), 向量角度(P限制)的变异在同龄和异龄混交组均主要由林分类型和微生境变化的交互作用解释(同龄混交:R2=0.160, 异龄混交:R2=0.398;P<0.001)。
| 林分类型Forest type | 微生境Microhabitat | 林分类型×微生境Forest type×Microhabitat | |||||||||
| F | R2 | P | F | R2 | P | F | R2 | P | |||
| 同龄混交Even-aged mixture | |||||||||||
| AG | 84.665 | 0.568* | <0.001 | 12.673 | 0.043 | <0.001 | 8.526 | 0.057 | <0.001 | ||
| BG | 83.034 | 0.519* | <0.001 | 33.741 | 0.105 | <0.001 | 10.749 | 0.067 | <0.001 | ||
| NAG | 272.698 | 0.634* | <0.001 | 136.832 | 0.159 | <0.001 | 39.746 | 0.092 | <0.001 | ||
| LAP | 42.840 | 0.356* | <0.001 | 10.442 | 0.043 | <0.001 | 22.724 | 0.189 | <0.001 | ||
| ACP | 7.714 | 0.081 | <0.001 | 38.095 | 0.201* | <0.002 | 18.420 | 0.195 | <0.001 | ||
| 角度Angle(°) | 469.268 | 0.749* | <0.001 | 172.107 | 0.137 | <0.001 | 21.410 | 0.034 | <0.001 | ||
| 长度Length | 0.815 | 0.014 | 0.448 | 0.469 | 0.004 | 0.504 | 9.599 | 0.160* | <0.001 | ||
| 异龄混交Uneven-aged mixture | |||||||||||
| AG | 757.842 | 0.712* | <0.001 | 210.395 | 0.099 | <0.001 | 151.720 | 0.143 | <0.001 | ||
| BG | 1112.405 | 0.521* | <0.001 | 172.698 | 0.040 | <0.001 | 888.082 | 0.416 | <0.001 | ||
| NAG | 333.244 | 0.434 | <0.001 | 7.961 | 0.005 | 0.005 | 381.044 | 0.496* | <0.001 | ||
| LAP | 101.765 | 0.054 | <0.001 | 53.792 | 0.014 | <0.001 | 1710.791 | 0.906* | <0.001 | ||
| ACP | 445.586 | 0.737* | <0.001 | 122.343 | 0.101 | <0.001 | 48.574 | 0.080 | <0.001 | ||
| 角度Angle(°) | 678.460 | 0.399 | <0.001 | 108.471 | 0.032 | <0.001 | 917.389 | 0.540* | <0.001 | ||
| 长度Length | 137.326 | 0.366 | <0.001 | 78.152 | 0.104 | <0.001 | 149.107 | 0.398* | <0.001 | ||
| 每一行中标注* 的R2为解释该指标最大程度变异的因素 | |||||||||||
Spearman相关性分析结果表明(图 3), 酶活性与土壤养分含量之间的相关性在根际土中低于非根际土。具体而言, AG、BG和NAG酶活性均与pH值呈显著负相关, 并与SOM、TN和AP呈显著正相关, 此外, BG酶活性与TP呈显著正相关(P<0.05)。在同龄混交林组, 与非根际土相比, 根际土中pH值与TN和TP之间的负相关性增强, TP与AP之间的正相关性增强(P<0.05);AG、BG和NAG酶活性与土壤pH值及养分含量之间的相关性减弱, 但ACP酶活性与pH值、NH4+-N和AP之间的相关性增强(P<0.05)。与同龄混交林相比, 异龄混交林非根际土pH值与SOM、TN以及AG、BG、NAG酶活性之间的相关性减弱, 但TP与NH4+-N、AP以及AG、BG和NAG酶活性之间相关性增强;LAP酶活性与pH值之间的相关性由负相关转变为显著正相关, ACP酶活性与SOM、TN和NO3--N的相关性增强(P<0.05);异龄混交林根际土中LAP和ACP酶活性与pH值、NH4+-N和NO3--N之间的相关性增强, ACP与SOM之间的负相关关系也增强。

|
| 图 3 土壤理化指标与酶活性之间的相关性 Fig. 3 Correlations between soil physicochemical characteristics and soil enzyme activities. |
林分内凋落物的数量和质量会显著影响土壤养分含量[28], 混交林中凋落物的种类和输入量增加, 加速了土壤有机质的分解以及其他矿质养分的积累[29], 促进土壤肥力的提高。本研究表明, 无论是同龄混交还是异龄混交, 都能明显改善土壤的养分状况, 大多数养分的含量在同龄混交林中均高于异龄混交林, 尤其是根际土中SOM的含量。由于树种本身特性的差异, 使根际分泌物对周围土壤的理化性质产生影响[30], 植物光合作用所产生的C在传输到根系之后会被细根的特定生理活动(如根系分泌)迅速消耗[31], 大量的光合C通过根系释放并被转化成为土壤有机质[31-32]。在异龄混交林中, 马尾松为1959年种植, 第24年时(1983年)引入红锥。研究表明, 马尾松在快速生长阶段十分消耗土壤养分, 针叶凋落物中较高的木质素/N和C/N比例不利于土壤有机质的积累, 使养分积累效率较低, 导致土壤养分与其他龄组的林分相比更加贫瘠[33]。因此, 虽然同龄与异龄混交林同在1983年营造, 但是林分中土壤初始养分含量的差异可能是导致同龄混交林的土壤养分含量优于异龄混交林的重要原因。
土壤中的有机质能够为酶反应提供底物和能量, 因此土壤中有机质的含量会对酶活性产生影响[34]。本研究的结果表明, 混交(尤其是同龄混交)增加了根际以及非根际土壤中的有机质含量, 这可能是由于混交林中凋落物种类、输入量和质量的改变加速了土壤有机质的积累[29];而在根际区域, 根系分泌物能够为该区域的微生物提供易获得的C源[32]。因此, 充足的有机质使酶促反应中底物的可利用性增强[34], 从而增加了土壤中与C循环相关的酶活性。与非根际土壤相比, 根际区域是酶促反应的热点区域, 植物的根际沉积物为根际区域提供了养分, 增强了营养物质的可利用性, 使根际区域的微生物活性与非根际区域有很大区别, 从而导致养分含量和酶活性在根际土壤与非根际土壤之间的显著差异[35-37]。
3.2 树种混交对根际和非根际土壤微生物资源限制的影响土壤酶活性的化学计量比常被用于表征土壤微生物对C、N、P养分的需求, 是评估微生物资源限制状况十分有效的指标[8, 27]。在全球范围内, C ∶ N ∶ P获取酶活性的比例接近1 ∶ 1 ∶ 1[38], 在本研究中, C ∶ N ∶ P比例介于1 ∶ 0.96 ∶ 0.69—1 ∶ 1.09 ∶ 0.87之间, 一定程度上偏离严格的1 ∶ 1 ∶ 1的规律, 表明所有林分类型根际和非根际土壤微生物均受到强烈的P限制。根据本研究的结果, 混交增强了非根际土中的C限制, 但是减弱了根际土中的C限制, 除此之外, 树种混交能够在一定程度上缓解土壤中微生物所受的P限制, 尤其是在同龄混交林的根际土中。总体来讲, 混交效应在根际区域的响应更直接, 也更强烈。马尾松纯林根际土中的P限制程度低于红锥混林, 在混交林中也是如此;单一树种的特性在混交林中得到了很好的保留, 但同龄混交更有利于降低马尾松根际区域微生物的P限制。除此之外, 混交对于C限制的影响在不同的土壤生境之间有明显差异, 具体表现为混交增强了非根际土中的C限制, 但缓解了根际土中的C限制, 这与本研究提出的假设基本相反。非根际土中的C限制在林分之间的差异可能与凋落物的组成与质量有关, C、N、P养分输入比例的变化可能会影响土壤中C的可利用性[39]。微生物受到某种养分的限制程度越强, 表明对该养分的需求越高[27], 在根际区域, 微生物能够直接利用根际沉积物中的大量C源以缓解C对于自身的限制。P通常是亚热带地区森林生态系统生产力的主要限制因子, 研究表明, 根际土与非根际土之间的P限制程度具有显著差异[7]。本研究中, 土壤微生物所受的P限制得到缓解, 这与本研究所提出的假设基本一致, 其中原因可能是多方面的。首先, ACP酶活性的变化是微生物对P养分需求变化最直接的反应[7];其次, 本研究中的两个树种, 马尾松和红锥均是我国亚热带地区分布广泛的典型外生菌根树种, 真菌对土壤有机质的分解可能会增加P的释放, 使土壤中P的可利用性增加, 从而缓解P限制[13, 40];除此之外, 混交林中根系生物量和根系分泌物的变化也可以影响酶的化学计量比并改变P的矿化过程[2], 使土壤中的P限制得到缓解。
4 结论本研究发现, 相对于针叶纯林, 针阔混交显著改善土壤养分状况, 且同龄混交对土壤养分的改善作用优于异龄混交。与非根际土壤相比, 根际区域土壤养分含量和酶活性受到混交的影响更强烈, 可见树种混交效应在根际区域的响应更为强烈。。本研究中, 不同林分类型的土壤微生物均受到C限制和强烈的P限制, 但与纯林相比, 混交加剧了非根际土壤中微生物所受的的C限制, 缓解了根际土中微生物所受的C限制;对于P限制而言, 混交在很大程度上能够缓解土壤整体的P限制, 但P仍然是土壤中微生物的主要限制因子。土壤pH值、有机质和氮磷养分的含量是驱动酶活性变化和影响微生物资源限制的关键因子。本研究结果进一步完善了土壤酶化学计量比理论体系, 同时也可为马尾松和红锥人工混交林的树种选择和养分管理等提供科学依据。
植被向土壤输入的资源在数量和质量上的差异是造成土壤酶活性变化和资源限制差异的重要因素, 而根际区域与非根际区域最大的区别就在于根系分泌物的影响, 但本研究并未设计进一步的试验内容。如果能够在本研究的基础上进一步结合凋落物与根系分泌相关的试验内容, 相信能够有更深一步的发现。在今后的人工林管理实践和研究中, 应重点关注树种混交缓解土壤微生物P限制的机制和作用过程以及树种混交效应在根际土中的具体表现。
| [1] |
Feng Y H, Schmid B, Loreau M, Forrester D I, Fei S L, Zhu J X, Tang Z Y, Zhu J L, Hong P B, Ji C J, Shi Y, Su H J, Xiong X Y, Xiao J, Wang S P, Fang J Y. Multispecies forest plantations outyield monocultures across a broad range of conditions. Science, 2022, 376(6595): 865-868. DOI:10.1126/science.abm6363 |
| [2] |
Huang Y X, Wu Z J, Zong Y Y, Li W Q, Chen F S, Wang G G, Li J J, Fang X M. Mixing with coniferous tree species alleviates rhizosphere soil phosphorus limitation of broad-leaved trees in subtropical plantations. Soil Biology and Biochemistry, 2022, 175: 108853. DOI:10.1016/j.soilbio.2022.108853 |
| [3] |
Liu C L C, Kuchma O, Krutovsky K V. Mixed-species versus monocultures in plantation forestry: Development, benefits, ecosystem services and perspectives for the future. Global Ecology and Conservation, 2018, 15: e00419. DOI:10.1016/j.gecco.2018.e00419 |
| [4] |
刘世荣, 杨予静, 王晖. 中国人工林经营发展战略与对策: 从追求木材产量的单一目标经营转向提升生态系统服务质量和效益的多目标经营. 生态学报, 2018, 38(01): 1-10. DOI:10.3969/j.issn.1673-1182.2018.01.001 |
| [5] |
Pereira A P A, Durrer A, Gumiere T, Gonçalves J L M, Robin A, Bouillet J P, Wang J T, Verma J P, Singh B K, Cardoso E J B N. Mixed Eucalyptus plantations induce changes in microbial communities and increase biological functions in the soil and litter layers. Forest Ecology and Management, 2019, 433: 332-342. DOI:10.1016/j.foreco.2018.11.018 |
| [6] |
Garau G, Morillas L, Roales J, Castaldi P, Mangia N P, Spano D, Mereu S. Effect of monospecific and mixed Mediterranean tree plantations on soil microbial community and biochemical functioning. Applied Soil Ecology, 2019, 140: 78-88. DOI:10.1016/j.apsoil.2019.04.005 |
| [7] |
Xu C B, Chang Y N, Penuelas J, Sardans J, Cheng D L, Li B Y, Zhong Q L. Mixing Machilus pauhoi with Cerasus campanulata improves soil P availability and changes the soil G+/G- in a mid-subtropical region of China. Plant and Soil, 2023, 486(1): 409-424. |
| [8] |
Moorhead D L, Sinsabaugh R L, Hill B H, Weintraub M N. Vector analysis of ecoenzyme activities reveal constraints on coupled C, N and P dynamics. Soil Biology and Biochemistry, 2016, 93: 1-7. DOI:10.1016/j.soilbio.2015.10.019 |
| [9] |
Cui Y X, Bing H J, Moorhead D L, Delgado-Baquerizo M, Ye L P, Yu J L, Zhang S P, Wang X, Peng S S, Guo X, Zhu B, Chen J, Tan W F, Wang Y Q, Zhang X C, Fang L C. Ecoenzymatic stoichiometry reveals widespread soil phosphorus limitation to microbial metabolism across Chinese forests. Communications Earth & Environment, 2022, 3: 1-8. |
| [10] |
Sinsabaugh R L, Follstad Shah J J. Ecoenzymatic Stoichiometry and Ecological Theory. Annual Review of Ecology, Evolution, and Systematics, 2012, 43: 313-343. DOI:10.1146/annurev-ecolsys-071112-124414 |
| [11] |
Sinsabaugh R L, Hill B H, Follstad Shah J J. Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment. Nature, 2009, 462(7274): 795-798. DOI:10.1038/nature08632 |
| [12] |
Wu L F, Zou B Z, Wang S R, Zhou L H, Zheng Y, Huang Z Q, He J Z. Effects of multispecies restoration on soil extracellular enzyme activity stoichiometry in Pinus massoniana plantations of subtropical China. Soil Biology and Biochemistry, 2023, 178: 108967. DOI:10.1016/j.soilbio.2023.108967 |
| [13] |
Sheng M, Chen X D, Yu X J, Yan J, Zhang X L, Hamel C, Sheng Y Y, Tang M. Neighborhood effects on soil properties, mycorrhizal attributes, tree growth, and nutrient status in afforested zones. Restoration Ecology, 2020, 28(2): 459-467. DOI:10.1111/rec.13099 |
| [14] |
Yang F, Tian J, Fang H J, Gao Y, Zhang X Y, Yu G R, Kuzyakov Y. Spatial heterogeneity of microbial community and enzyme activities in a broad-leaved Korean pine mixed forest. European Journal of Soil Biology, 2018, 88: 65-72. DOI:10.1016/j.ejsobi.2018.07.001 |
| [15] |
You Y M, Huang X M, Zhu H G, Liu S R, Liang H W, Wen Y G, Wang H, Cai D X, Ye D. Positive interactions between Pinus massoniana and Castanopsis hystrix species in the uneven-aged mixed plantations can produce more ecosystem carbon in subtropical China. Forest Ecology and Management, 2018, 410: 193-200. DOI:10.1016/j.foreco.2017.08.025 |
| [16] |
Kuzyakov Y, Razavi B S. Rhizosphere size and shape: Temporal dynamics and spatial stationarity. Soil Biology and Biochemistry, 2019, 135: 343-360. DOI:10.1016/j.soilbio.2019.05.011 |
| [17] |
Smalla K, Sessitsch A, Hartmann A. The Rhizosphere: 'soil compartment influenced by the root'. FEMS Microbiology Ecology, 2006, 56(2): 165. DOI:10.1111/j.1574-6941.2006.00148.x |
| [18] |
Ling N, Wang T T, Kuzyakov Y. Rhizosphere bacteriome structure and functions. Nature Communications, 2022, 13(1): 836. DOI:10.1038/s41467-022-28448-9 |
| [19] |
Ma Y D, Yue K, Heděnec P, Li C H, Li Y, Wu Q Q. Global patterns of rhizosphere effects on soil carbon and nitrogen biogeochemical processes. Catena, 2023, 220: 106661. DOI:10.1016/j.catena.2022.106661 |
| [20] |
Gan D Y, Feng J G, Han M G, Zeng H, Zhu B. Rhizosphere effects of woody plants on soil biogeochemical processes: A meta-analysis. Soil Biology and Biochemistry, 2021, 160: 108310. DOI:10.1016/j.soilbio.2021.108310 |
| [21] |
温远光, 李海燕, 周晓果, 朱宏光, 李运筹, 蔡道雄, 贾宏炎, 黄雪蔓, 尤业明. 马尾松×红锥异龄混交林对土壤微生物群落结构和功能的影响. 广西科学, 2019, 26(02): 188-198. |
| [22] |
Knoke T, Ammer C, Stimm B, Mosandl R. Admixing broadleaved to coniferous tree species: a review on yield, ecological stability and economics. European Journal of Forest Research, 2008, 127(2): 89-101. DOI:10.1007/s10342-007-0186-2 |
| [23] |
Li W Q, Huang Y X, Chen F S, Liu Y Q, Lin X F, Zong Y Y, Wu G Y, Yu Z R, Fang X M. Mixing with broad-leaved trees shapes the rhizosphere soil fungal communities of coniferous tree species in subtropical forests. Forest Ecology and Management, 2021, 480: 118664. DOI:10.1016/j.foreco.2020.118664 |
| [24] |
Lu J Z, Scheu S. Response of soil microbial communities to mixed beech-conifer forests varies with site conditions. Soil Biology and Biochemistry, 2021, 155: 108155. DOI:10.1016/j.soilbio.2021.108155 |
| [25] |
鲍士旦. 土壤农化分析 (3版). 北京: 中国农业出版社, 2000.
|
| [26] |
Bell C W, Fricks B E, Rocca J D, Steinweg J M, McMahon S K, Wallenstein M D. High-throughput fluorometric measurement of potential soil extracellular enzyme activities. Journal of Visualized Experiments, 2013(81): e50961. |
| [27] |
Moorhead D L, Rinkes Z L, Sinsabaugh R L, Weintraub M N. Dynamic relationships between microbial biomass, respiration, inorganic nutrients and enzyme activities: informing enzyme-based decomposition models. Frontiers in Microbiology, 2013, 4: 223. |
| [28] |
Thoms C, Gattinger A, Jacob M, Thomas F M, Gleixner G. Direct and indirect effects of tree diversity drive soil microbial diversity in temperate deciduous forest. Soil Biology and Biochemistry, 2010, 42(9): 1558-1565. DOI:10.1016/j.soilbio.2010.05.030 |
| [29] |
陈璟, 杨宁. 亚热带红壤丘陵区5种人工林对土壤性质的影响. 西北农林科技大学学报(自然科学版), 2013, 41(12): 167-173, 178. |
| [30] |
Likulunga L E, Rivera Pérez C A, Schneider D, Daniel R, Polle A. Tree species composition and soil properties in pure and mixed beech-conifer stands drive soil fungal communities. Forest Ecology and Management, 2021, 502: 119709. DOI:10.1016/j.foreco.2021.119709 |
| [31] |
Chen H Y, Dong Y F, Xu T, Wang Y P, Wang H T, Duan B L. Root order-dependent seasonal dynamics in the carbon and nitrogen chemistry of poplar fine roots. New Forests, 2017, 48(5): 587-607. DOI:10.1007/s11056-017-9587-3 |
| [32] |
Kuzyakov Y, Domanski G. Carbon input by plants into the soil. Review. Journal of Plant Nutrition and Soil Science, 2000, 163(4): 421-431.
|
| [33] |
葛晓改, 肖文发, 曾立雄, 黄志霖, 付甜, 封晓辉. 不同林龄马尾松凋落物基质质量与土壤养分的关系. 生态学报, 2012, 32(3): 852-862. |
| [34] |
Cenini V L, Fornara D A, McMullan G, Ternan N, Carolan R, Crawley M J, Clément J C, Lavorel S. Linkages between extracellular enzyme activities and the carbon and nitrogen content of grassland soils. Soil Biology and Biochemistry, 2016, 96: 198-206. DOI:10.1016/j.soilbio.2016.02.015 |
| [35] |
Fan K K, Weisenhorn P, Gilbert J A, Chu H Y. Wheat rhizosphere harbors a less complex and more stable microbial co-occurrence pattern than bulk soil. Soil Biology and Biochemistry, 2018, 125: 251-260. DOI:10.1016/j.soilbio.2018.07.022 |
| [36] |
Zhang Y L, Sun C X, Chen Z H, Zhang G N, Chen L J, Wu Z J. Stoichiometric analyses of soil nutrients and enzymes in a Cambisol soil treated with inorganic fertilizers or manures for 26 years. Geoderma, 2019, 353: 382-390. DOI:10.1016/j.geoderma.2019.06.026 |
| [37] |
吉艳芝, 冯万忠, 陈立新, 段文标, 张笑归. 落叶松混交林根际与非根际土壤养分、微生物和酶活性特征. 生态环境, 2008(01): 339-343. |
| [38] |
Sinsabaugh R L, Lauber C L, Weintraub M N, Ahmed B, Allison S D, Crenshaw C, Contosta A R, Cusack D, Frey S, Gallo M E, Gartner T B, Hobbie S E, Holland K, Keeler B L, Powers J S, Stursova M, Takacs-Vesbach C, Waldrop M P, Wallenstein M D, Zak D R, Zeglin L H. Stoichiometry of soil enzyme activity at global scale. Ecology Letters, 2008, 11(11): 1252-1264. DOI:10.1111/j.1461-0248.2008.01245.x |
| [39] |
Qi D D, Feng F J, Lu C, Fu Y M. C ∶ N ∶ P stoichiometry of different soil components after the transition of temperate primary coniferous and broad-leaved mixed forests to secondary forests. Soil and Tillage Research, 2022, 216: 105260. DOI:10.1016/j.still.2021.105260 |
| [40] |
Chen X L, Chen H Y H, Chang S X. Meta-analysis shows that plant mixtures increase soil phosphorus availability and plant productivity in diverse ecosystems. Nature Ecology & Evolution, 2022, 6(8): 1112-1121. |