 2024, Vol. 44
2024, Vol. 44文章信息
- 温馨, 王天罡, 程小琴
- WEN Xin, WANG Tiangang, CHENG Xiaoqin
- 山西太岳山不同径级油松种内竞争与叶功能性状
- The intraspecious competition and leaf functional traits of Pinus tabulaeformis with different diameter classes in Taiyue Mountain, Shanxi Province
- 生态学报. 2024, 44(22): 10300-10308
- Acta Ecologica Sinica. 2024, 44(22): 10300-10308
- http://dx.doi.org/10.20103/j.stxb.202404210885
-
文章历史
- 收稿日期: 2024-04-21
- 采用日期: 2024-08-18





2. 北京市园林绿化规划和资源监测中心(北京市林业碳汇与国际合作事务中心), 北京 100029
2. Beijing Forestry and Parks Planning and Resource Monitoring Center(Beijing Forestry Carbon and lnternational Cooperation Center), Beijing 100029, China
竞争是指两个及两个以上的有机体在资源不足时, 为争夺有限资源而产生的相互作用关系[1—3]。在森林群落内部, 竞争关系主要发生在相邻个体之间, 涉及光照、水分和养分的争夺[4—5]。已有研究表明, 竞争主要通过密度制约效应调节树木的数量与种群结构, 随着种群年龄的增长, 植物需要更多的资源来满足自身生长, 这加剧了个体间的相互作用, 导致自疏现象, 进而降低植物的种群密度[6]。竞争分为种间竞争与种内竞争。同一物种由于生态位相同, 种内竞争是竞争的主要形式[7]。从个体角度看, 种内竞争会导致植物个体能量供给不足, 限制其生长发育。从种群角度看, 种内竞争有利于淘汰较弱个体, 促进强健个体的生存, 有助于种群的进化发展[8]。为量化林木间的竞争关系, 竞争指数的研究为分析竞争强度提供了重要途径[9]。竞争指数本质上反映了林分对资源的需求与实际资源占有量之间的关系[4, 10—11]。Hegyi单木竞争指数模型因其简单易算, 能够很好地描述植物间的竞争压力[12], 广泛应用于相关研究中。
植物功能性状是影响个体生长、存活与繁殖的表型特征, 并间接影响个体适合度[13]。作为植物与环境沟通的桥梁, 植物功能性状反映了植物在进化过程中对环境的适应性与竞争能力[14]。在生长过程中, 植物会形成一系列的性状组合以更好地适应环境变化[15]。这些性状组合之间存在内在的权衡与联系[16—17]。通常, 叶片寿命长、光合速率高的植物属于“快速投资收益型”, 而叶片寿命短、叶氮磷含量低的植物属于“缓慢投资收益型”[17—19]。植物功能性状一直是研究热点, 绝大多数研究者关注植物的种间变异, 但种内变异同样不容忽视。例如, Siefert[20]对全球629个群落36个功能性状进行分析, 发现种内变异占群落性状总变异的25%, 占群落间性状总变异的32%。在资源丰富的环境中, 植物主要通过调整叶片性状来缓解种内竞争带来的的生存压力[21]。环境变化会导致植物叶功能性状的改变, 进而加剧种内竞争[22]。研究植物的种内竞争及其叶功能性状的变化, 有助于理解森林植物群落结构的动态变化。
油松(Pinus tabuliformis)具有较长的生活史, 且1年生与多年生针叶共存。尽管目前关于油松种内竞争、功能性状的研究较多, 但对其在生长发育过程中的变化研究较少。因此, 本研究以山西太岳山油松人工林为研究对象, 利用Hegyi单木竞争指数模型, 研究不同径级下油松的种内竞争及其叶功能性状的变化, 从而揭示植物个体间的相互作用与适应机制。
1 研究方法 1.1 研究区概况实验区位于山西省太岳山七里峪三眼窑林场(36°21′N—36°45′N, 111°20′E—111°40′E)。该区域属于暖温带大陆性气候区, 具有良好的水热条件。年平均气温为9.3℃, 昼夜温差较大, 无霜期长达125d。年降水量为500—700mm, 降雨主要集中在7月、8月和9月份。海拔范围为1850—1890m。土壤类型主要为山地褐土, 林下物种丰富, 生物多样性较高。该林场于20世纪70—80年代采用农林混播造林的方法进行种植, 林龄47年。2011年进行过低强度的抚育, 现在林分的平均胸径为21.32cm, 蓄积量为181.5m2, 郁闭度为0.75。乔木层的优势树种为油松, 灌木层的优势种包括茅梅(Rubus parvifolius)、胡枝子(Lespedeza bicolor)、沙棘(Hippophae rhamnoides)、黄刺梅(Rosa davurica)等, 草本层优势种有小红菊(Dendranthema chanetii)、羊胡子草(Incarvillea arguta)、细叶苔草(Carex rigescens)等。
1.2 实验设计2019年8月, 在油松人工林的分布范围内, 选取坡度、海拔和密度较为一致的典型样地3个, 样地大小设置为20m×30m。在每个样地内记录林木的X、Y坐标、冠幅、胸径(DBH)、树高、活枝下高。根据胸径大小, 将油松按5cm间隔进行径级划分, 样地内的油松划分为6个径级:Ⅰ(DBH≤15cm);Ⅱ(15cm<DBH≤20cm);Ⅲ(20cm<DBH≤25cm);Ⅳ(25cm<DBH≤30cm);Ⅴ(30cm<DBH≤35cm);Ⅵ(>35cm)。为消除边缘效应并且保证足够的样本进行拟合。研究对象木范围是以样地中心为圆心, 半径为样地短边的1/2减去样地林木最大冠幅直径。竞争木范围是整个样地, 竞争木是距离对象木最近的4棵邻接木[23]。样地信息基本概况见表 1。
| 样地 Plot |
密度 Density/(株/hm2) |
海拔 Elevation/m |
坡向 Slope aspect |
平均胸径 Average DBH/cm |
平均树高 Average height/m |
| 1 | 1050 | 1880 | 东偏南12° | 21.58±5.7 | 16.53±1.7 |
| 2 | 1100 | 1889 | 东偏南12° | 21.58±6.0 | 16.53±2.0 |
| 3 | 917 | 1853 | 东偏北35° | 20.47±6.1 | 17.39±3.9 |
| DBH:胸径Diameter at breast height | |||||
根据样地调查, 在每个径级中选取3株生长旺盛且具有代表性的油松作为标准木, 共18株标准木。在树冠中部的东、西、南、北四个方向上, 选取生长良好的标准木轮生枝, 再从轮生枝中选取里、中、外三个部位的具有代表性的1年生叶和多年生叶各100束(油松1年生叶为嫩绿色, 质地较脆;多年生叶为深绿色, 韧性较大)。采集时, 用事先准备好的湿润滤纸包裹松针, 随后将其装入自封袋, 并放入保鲜盒中保存。
1.3.2 功能性状相关指标及测定方法从自封袋中随机选取20束松针, 用滤纸吸干表面水分, 使用精度为万分之一的电子天平称量松针的叶鲜重(LFW)。将随机选取的20束针叶整齐平铺, 固定在透明文件夹中, 用精度为0.01mm的数显游标卡尺测量叶长(LL)、叶宽(LD)、叶厚(LT)。在将松针置于85℃烘箱烘干48h, 直至恒重, 并测量叶干重(LDW)。在测量完上述指标后, 再进行其他相关指标的测定。油松的叶面积可以通过计算得到, 计算公式如下:
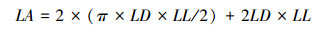
|
(1) |
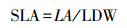
|
(2) |
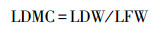
|
(3) |
烘干后的样品利用粉碎机磨粉过细筛, 经硫酸—过氧化氢消煮后, 用于测定叶氮含量(LNC)与叶磷含量(LPC)。叶氮含量(LNC)采用凯氏定氮法测定, 叶磷含量(LPC)通过钼锑抗比色法测定。
1.4 数据处理 1.4.1 竞争强度为表征研究区样地的种内竞争状况, 选用Hegyi的单木竞争指数模型[12], 公式如下:
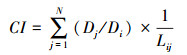
|
(4) |
式中, j、i分别表示竞争木与对象木, CI为对象木竞争指数, Dj为竞争木胸径, Di为对象木胸径, N为竞争木的株数, Lij为对象木和竞争木之间的距离。CI值越大, 表示对象木受到的竞争强度越大。
1.4.2 数据分析利用Excel对油松对象木的胸径、竞争指数以及叶功能性状进行统计计算, 利用SPSS 27.0对不同径级的油松的种内竞争指数和叶功能性状进行单因素方差分析(One-way ANOVA), 通过Pearson相关性分析油松叶功能性状之间的相关性。使用OriginPro 2024b软件对各个性状进行绘图及线性回归拟合。
2 结果与分析 2.1 单木竞争共调查对象木153株, 平均胸径21.67cm, 平均树高16.75m(表 2)。在各径级中油松树木分布不均匀, 其中Ⅱ、Ⅲ、Ⅳ径级的树木分布比较多, 占总数的70%以上。相比之下, Ⅰ径级油松数量比较少, 占总数的13.73%, 而Ⅴ和Ⅵ径级油松数量最少, 占总数的9.74%。竞争木共调查198株, 平均胸径为21.53cm, 平均树高为16.65m。在前四个径级中, 竞争木的株数和平均胸径分布较为均匀。对象木与竞争木的胸径随着树高的增加而增加, 二者呈正相关性关系。
| 径级 Diameter class/cm |
对象木Objective tree | 竞争木Competitor | |||||||
| 株数 Number/株 |
百分比 Number/% |
平均胸径 Average DBH/cm |
平均树高 Average height/m |
株数 Number/株 |
百分比 Percentage/% |
平均胸径 Average DBH/cm |
平均树高 Average height/m |
||
| Ⅰ | 21 | 13.73 | 13.53±1.16 | 14.17±2.60 | 40 | 20.2 | 13.38±1.18 | 13.97±2.83 | |
| Ⅱ | 43 | 28.10 | 17.33±1.34 | 15.97±2.07 | 45 | 22.73 | 17.38±1.23 | 16.18±2.31 | |
| Ⅲ | 42 | 27.45 | 22.46±1.50 | 17.10±2.38 | 49 | 24.75 | 22.54±1.49 | 17.20±2.34 | |
| Ⅳ | 32 | 20.92 | 27.08±1.19 | 18.17±1.90 | 43 | 21.72 | 27.02±1.37 | 18.01±1.96 | |
| Ⅴ | 12 | 7.84 | 31.46±0.75 | 18.54±4.32 | 18 | 9.09 | 31.50±0.99 | 18.51±2.82 | |
| Ⅵ | 3 | 1.9 | 37.57±1.91 | 19.96±2.89 | 3 | 1.52 | 37.57±1.91 | 19.96±2.89 | |
| 合计 | 153 | 100 | 21.67±5.82 | 16.75±2.77 | 198 | 100 | 21.53±6.24 | 16.65±2.86 | |
| Ⅰ:DBH≤15cm;Ⅱ:15cm<DBH≤20cm;Ⅲ:20cm<DBH≤25cm;Ⅳ:25cm<DBH≤30cm;Ⅴ:30cm<DBH≤35cm;Ⅵ:>35cm | |||||||||
油松的种内竞争强度随径级的增大而减小(表 3)。其中径级Ⅵ油松的种内竞争强度最低, 株数最少, 竞争指数在径级Ⅰ、Ⅱ与其他径级之间存在显著差异, 径级Ⅲ、Ⅳ与径级Ⅵ有显著差异。
| 径级 Diameter class/cm |
株数 Number/株 |
竞争指数 Competitive index(CI) |
径级 Diameter class/cm |
株数 Number/株 |
竞争指数 Competitive index(CI) |
|
| Ⅰ | 21 | 4.012a | Ⅴ | 12 | 1.415bc | |
| Ⅱ | 43 | 3.232a | Ⅵ | 3 | 0.889c | |
| Ⅲ | 42 | 2.010b | 合计Total | 153 | 13.408 | |
| Ⅳ | 32 | 1.850b | ||||
| 不同字母代表差异显著(P<0.05) | ||||||
将对象木胸径与竞争指数拟合, 二者符合幂函数关系(图 1), 对象木胸径越大, 受到的种内竞争压力越小, 当对象木胸径增大到20cm之后, 竞争指数逐渐趋于平稳。

|
| 图 1 对象木胸径与竞争指数关系拟合 Fig. 1 Relationship fitting between DBH and competition index of object trees |
根据不同径级油松叶功能性状变化规律结果(图 2), 随径级的增加, 1年生叶与多年生叶的SLA逐渐降低, 且1年生叶的SLA均明显高于多年生叶的SLA。1年生叶与多年生叶的SLA随径级的变化趋势基本相同, 径级Ⅰ的SLA与其他径级的差异显著, 径级Ⅱ、Ⅲ的SLA与径级Ⅳ、Ⅴ、Ⅵ的差异显著。但1年生叶SLA径级Ⅳ、Ⅴ、Ⅵ之间无显著差异, 而多年生叶径级Ⅵ与Ⅳ的差异显著, 与径级Ⅴ的不存在显著差异。1年生叶与多年生叶的LDMC均随径级增加呈上升趋势, 且1年生叶的LDMC显著低于多年生叶LDMC。1年生叶的LDMC径级Ⅰ、Ⅴ与其他径级的呈显著性差异, 径级Ⅱ、Ⅲ、Ⅳ、Ⅵ的1年生叶LDMC之间无显著差异;多年生叶径级Ⅰ、Ⅱ的LDMC与其他径级呈显著性差异, 且径级Ⅰ与径级Ⅱ, 径级Ⅳ与径级Ⅴ多年生叶的LDMC也呈显著差异关系。

|
| 图 2 叶功能性状随径级变化规律 Fig. 2 Changes of leaf functional traits with diameter class of Pinus tabulaeformis 不同小写字母表示差异显著(P<0.05);Ⅰ:DBH≤15cm;Ⅱ:15cm<DBH≤20cm;Ⅲ:20cm<DBH≤25cm;Ⅳ:25cm<DBH≤30cm;Ⅴ:30cm<DBH≤35cm;Ⅵ:>35cm |
1年生叶和多年生叶的LNC整体上随着径级增加逐渐上升, 其中1年叶的LNC要大于多年生叶的LNC。1年生叶与多年生叶的LNC在Ⅰ、Ⅱ径级之间不存在显著差异, 但与径级Ⅵ之间差异显著, 径级Ⅵ 1年生叶与多年生叶的LNC达到最大值。1年生叶的LPC波动不大, 各径级之间无显著性差异;多年生叶的LPC随径级呈现出先下降后上升的趋势, 且在径级Ⅵ多年生叶的LPC最大, 径级Ⅳ多年生叶的LPC最小, 径级Ⅵ与其他径级多年生叶的LPC存在显著性差异。1年生叶的N/P在不同径级之间波动不大, 未达到显著差异, 而多年生叶的N/P则存在显著差异, 径级Ⅲ、Ⅳ多年生叶的N/P值最大值, 且Ⅲ、Ⅳ径级的与其他径级的存在显著差异。
2.2.2 不同径级叶功能性状线性拟合对油松叶片的各个性状进行回归分析结果显示(图 3), 1年生叶的SLA、LDMC、LNC与径级之间拟合效果较好, 呈现良好的线性关系;而1年生叶的LPC、N/P与径级的回归关系模型拟合度较低, 不存在显著的线性关系。多年生叶的SLA、LDMC、LNC、LPC与径级之间的拟合R2较好, 线性关系良好, 多年生叶的N/P与径级的回归关系模型拟合度较低, 不存在显著的线性关系。

|
| 图 3 油松叶功能性状与径级回归分析 Fig. 3 Leaf functional traits and diameter classes regression analysis of Pinus tabulaeformis |
油松的1年生叶与多年生叶功能性状之间具有一定的相关性(图 4)。总体来看, 1年生叶与多年生叶的LDMC和SLA、LPC和N/P之间呈极显著负相关, LDMC和LNC存在显著正相关。1年生叶中SLA和LNC存在显著负相关, LNC和N/P存在显著正相关。多年生叶中, LNC与LPC存在极显著正相关, SLA和LNC存在极显著负相关, SLA和LPC存在显著负相关。

|
| 图 4 1年生叶与多年生叶叶功能性状间相关性 Fig. 4 Correlation of leaf functional traits between annual leaves and perennial leaves LDMC:叶干物质含量Leaf dry matter content;SLA:比叶面积Specific leaf area;LNC:叶氮含量Leaf nitrogen concentration;LPC:叶磷含量Leaf phosphorus concentration;N/P:叶氮磷比Leaf N/P ratio;** P<0.01;* P<0.05 |
在群落内部, 生态习性相似的植物, 生态位重叠越高, 对相同资源的竞争就越为激烈[5, 25]。对于林木生长来说, 光照是竞争的主要资源。只有充足的光照, 林木才能进行正常的光合作用。个体较大的林木会对冠层下的植物产生遮蔽, 使得林下的植物可获得的光较少[26]。这使得个体较小的林木遭受来自个体较大林木的不对称性竞争[27]。在本研究中, 小径级林木株数较多, 树高偏低, 竞争指数偏高, 而大径级林木则相反。这说明, 小径级林木为了保持稳定生长, 在争夺有限环境资源时, 种内竞争尤为激烈。
通过对153株对象木与198株竞争木调查, 发现竞争指数与对象木胸径之间符合幂函数关系, 这与以往的研究成果相一致[1—2, 6]。当胸径大于20cm, 油松的种内竞争强度明显减小。随着径级的增加, 油松的种内竞争强度变化不明显。进一步说明幼龄级油松胸径小, 对于环境资源的竞争能力较弱;当油松的胸径超过20cm, 此时树木居于乔木层, 获取资源的能力增强, 使其比胸径较小的树木具有更大的竞争优势, 受到竞争的影响较少[28]。总体而言, 油松的种内竞争强度取决于自身所处的生长阶段, 随着油松胸径增大, 种内竞争逐渐减弱。
3.2 不同径级油松叶功能性状变化规律叶片是植物对环境最敏感的器官, 叶片性状能够体现植物对环境的适应, 并对环境的变化有着积极响应[29]。研究发现, 1年生叶的SLA、LNC、LPC比多年生叶的高, 而LDMC则反之。油松的1年生叶是光合作用的主要场所, SLA越大, 光合作用越强, 碳同化能力也越强。氮磷元素作为植物中可以相互转移的元素, 当多年生叶的代谢能力下降后, 便将氮磷元素转移到1年生叶中, 来提高1年生叶的营养水平, 增强营养物质的合成[30]。
研究结果还显示, 随着油松径级的增加, SLA逐渐下降, LDMC逐渐上升。小径级的油松的种内竞争明显高于大径级油松, 说明小径级油松在面临更大的竞争压力时, 通过增大叶面积来获得更多的光资源[31];而大径级油松因竞争压力较小, 能够更好地享有充足的光照、土壤水分及营养物质, 因此更注重在叶组织密度和叶片厚度上的投资, 以抵御不良环境因素[32]。通常, 以资源获取型策略为主的植物SLA高、LDMC低, 具有更高的养分获取能力;以资源保守型策略为主的植物SLA低、LDMC高, 具有更高的资源存储能力[33]。这表明, 油松在在生长过程中逐渐从资源获取型向资源保守型转变。
植物叶片中的N、P元素是叶绿素和酶的主要元素, 对植物的光合作用与呼吸作用有着重要影响[34]。研究发现, LNC随径级的增加呈上升趋势, LPC随径级增加变化不明显。这表明, 随着油松径级的增加, 油松生长加快, 树干材积快速增长, 植物的细胞生命活动旺盛, 需合成大量蛋白质来满足需求[35]。N/P是评估生态系统受N或受P限制的重要指标, 当N/P低于14时, 植物受N限制;当N/P高于16时, 植物受P限制;当N/P介于14和16之间时, 植物同时受N和P限制[36]。研究表明, 油松1年生叶主要受到N限制, 多年生叶的生长受限和径级大小有关, 径级Ⅰ、Ⅱ、Ⅴ、Ⅵ的多年生叶受到N限制, Ⅲ、Ⅳ径级的多年生叶受到N、P共同限制。
3.3 油松叶功能性状的相关性植物功能性状之间存在紧密的联系[37]。1年生叶与多年生叶的SLA与LDMC负相关, LDMC和LNC呈显著正相关, 1年生叶的SLA和LNC呈显著负相关;而多年生叶的SLA和LNC呈极显著负相关, SLA和LPC呈显著负相关。这表明, 油松对资源利用能力和保存能力较高, SLA大的叶片叶面积大, 叶片厚度低、质量低, 因而叶片薄, LDMC低[38], 有利于降低叶片建成成本, 减少N消耗, 植物会将更多的N分配至叶绿体中, 提高光合作用速率[39]。N、P元素作为植物生长的重要因素, LNC与LPC存在耦合关系, LPC调控植物生理过程和LNC与生产力的关系, 在N限制的环境下, 林木生长与N/P呈正相关性, 在P限制的环境下, 林木生长与N/P呈负相关[40]。1年生叶与多年生叶LPC与N/P呈极显著负相关, 由此表明, P是限制油松生长的主要元素。
4 结论(1) 小径级油松种内强度最高, 随油松径级的增加, 种内竞争强度逐渐下降。(2)胸径与竞争指数之间存在幂函数关系。当胸径小于20cm, 油松的种内竞争强度较大, 当胸径大于20cm, 油松的种内竞争强度逐渐平缓。(3)不同径级油松叶功能性状采取不同适应策略, 随着树木的生长, 油松逐渐由资源获取型向资源保守型转变。(4)油松叶功能性状之间存在一定的相关性, SLA与LDMC、LPC与N/P、SLA与LNC呈现显著负相关性, 多年生叶中LPC与LNC呈正相关, 油松在不同生活史阶段叶片存在不同制约性。
| [1] |
李颜斐. 中亚热带杉木人工林空间结构及种内竞争关系[D]. 长沙: 中南林业科技大学, 2020.
|
| [2] |
杜秀芳, 汤孟平, 潘建勇, 沈钱勇, 杨帆. 临安区不同森林类型竞争指数比较研究. 生态学报, 2020, 40(12): 4064-4072. |
| [3] |
Trinder J C, Brooker W R, Robinson D. Plant ecology's guilty little secret: understanding the dynamics of plant competition. Functional Ecology, 2013, 27(4): 918-929. DOI:10.1111/1365-2435.12078 |
| [4] |
李际平, 房晓娜, 封尧, 孙华, 曹小玉, 赵春燕, 李建军. 基于加权Voronoi图的林木竞争指数. 北京林业大学学报, 2015, 37(3): 61-68. |
| [5] |
巢林, 刘艳艳, 张伟东, 杨庆朋, 汪思龙. 沼泽交错带白桦个体生长动态及径级结构与种内种间竞争. 生态学杂志, 2017, 36(3): 577-583. |
| [6] |
赵长兴, 赵维俊, 张兴林, 刘思敏, 牟文博, 刘金荣. 祁连山排露沟流域青海云杉种群种内竞争与促进作用分析. 植物生态学报, 2022, 46(9): 1027-1037. |
| [7] |
Yang X Z, Zhang W H, He Q Y. Effects of intraspecific competition on growth, architecture and biomass allocation of Quercus Liaotungensis. Journal of Plant Interactions, 2019, 14(1): 284-294. DOI:10.1080/17429145.2019.1629656 |
| [8] |
周文嵩. 华北落叶松次生林种内、种间关系及影响机制研究[D]. 北京: 北京林业大学, 2018.
|
| [9] |
李超, 裴顺祥, 张连金, 郭嘉, 辛学兵. 北京油松人工林竞争指数的适用性评价. 浙江农林大学学报, 2019, 36(6): 1115-1124. |
| [10] |
廉琪, 张弓乔, 萨日娜, 卢彦磊, 刘文桢, 胡艳波, 赵中华. 基于树冠重叠面积的天然混交林林木竞争指数. 生态学报, 2024, 44(5): 2057-2068. |
| [11] |
郝芳华. 太行山油松人工林结构与竞争特征及经营模式研究[D]. 太谷: 山西农业大学, 2016.
|
| [12] |
周泽宇, 周超凡, 胡兴国, 陈科屹, 杜满义, 张会儒, 符利勇. 基于距离相关Hegyi指数的云冷杉天然林单木胸径生长模型. 北京林业大学学报, 2023, 45(10): 59-69. DOI:10.12171/j.1000-1522.20210510 |
| [13] |
陈世仁, 臧丽鹏, 吴宇航, 杨泽宇, 张广奇, 刘庆福, 陈丹梅, 张树梓, 隋明浈. 植物功能性状种内变异的来源、量化及应用. 生态学杂志, 2024, 43(5): 1442-1452. |
| [14] |
徐睿, 刘静, 王利艳, 颜耀, 马祥庆, 李明. 不同地理种源杉木根叶功能性状与碳氮磷化学计量分析. 生态学报, 2022, 42(15): 6298-6310. |
| [15] |
税伟, 郭平平, 朱粟锋, 冯洁, 孙祥, 李慧. 云南喀斯特退化天坑木本植物功能性状变异特征及适应策略. 地理科学, 2022, 42(7): 1295-1306. |
| [16] |
刘聪聪, 何念鹏, 李颖, 张佳慧, 闫镤, 王若梦, 王瑞丽. 宏观生态学中的植物功能性状研究: 历史与发展趋势. 植物生态学报, 2024, 48(1): 21-40. |
| [17] |
李祖婵, 玄锦, 王秋雪, 彭书婷, 孙耿敏, 黄柳菁. 闽江江心洲不同生境的植物叶功能性状差异及其对土壤特性的响应. 生态学报, 2024, 44(14): 1-16. |
| [18] |
刘文倩, 李家湘, 龚俊伟, 赵丽娟, 项文化, 程幸, 王维嘉, 张意灵. 柯-青冈常绿阔叶林优势树种叶片性状变异及适应策略. 生态学报, 2022, 42(17): 7256-7265. |
| [19] |
Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T L, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets U, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821-827. DOI:10.1038/nature02403 |
| [20] |
Siefert A, Violle C, Chalmandrier L, Albert C H, Taudiere A, Fajardo A, Aarssen L W, Baraloto C, Carlucci M B, Cianciaruso M V, de L Dantas V, de Bello F, Duarte L D S, Fonseca C R, Freschet G T, Gaucherand S, Gross N, Hikosaka K, Jackson B, Jung V, Kamiyama C, Katabuchi M, Kembel S W, Kichenin E, Kraft N J B, Lagerström A, Bagousse-Pinguet Y L, Li Y Z, Mason N, Messier J, Nakashizuka T, Overton J M, Peltzer D A, Pérez-Ramos I M, Pillar V D, Prentice H C, Richardson S, Sasaki T, Schamp B S, Schöb C, Shipley B, Sundqvist M, Sykes M T, Vandewalle M, Wardle D A. A global meta-analysis of the relative extent of intraspecific trait variation in plant communities. Ecology Letters, 2015, 18(12): 1406-1419. DOI:10.1111/ele.12508 |
| [21] |
陈娟, 张小晶, 李巧玉, 陶建平. 不同坡向川西亚高山林木竞争与叶片表型可塑性的关系研究. 生态学报, 2022, 42(5): 1788-1797. |
| [22] |
何雁, 姚玉萍, 姚义鹏, 姜勇, 梁士楚, 李月娟, 梁欢欢, 赵庆年, 黄勇焙, 林才健. 桂林岩溶石山青冈群落植物功能性状的种间和种内变异研究. 生态学报, 2021, 41(20): 8237-8245. |
| [23] |
孜来比·买木提名, 杨华, 赵广亮, 王玲. 单木竞争指标的研究进展. 西北林学院学报, 2012, 27(6): 152-158. |
| [24] |
张凯, 侯继华, 何念鹏. 油松叶功能性状分布特征及其控制因素. 生态学报, 2017, 37(3): 736-749. |
| [25] |
罗梅, 陈绍志. 不同龄组长白落叶松种内及种间竞争研究. 北京林业大学学报, 2018, 40(9): 33-44. |
| [26] |
陈仁飞, 姬明飞, 关佳威, 邓建明. 植物对称性竞争与非对称性竞争研究进展及展望. 植物生态学报, 2015, 39(5): 530-540. |
| [27] |
Zhang X Q, Wang Z, Chhin S, Wang H C, Duan A G, Zhang J G. Relative contributions of competition, stand structure, age, and climate factors to tree mortality of Chinese fir plantations: long-term spacing trials in Southern China. Forest Ecology and Management, 2020, 465: 118103. DOI:10.1016/j.foreco.2020.118103 |
| [28] |
项小燕, 吴甘霖, 段仁燕, 闫玉梅, 张小平. 大别山五针松种内和种间竞争强度. 生态学报, 2015, 35(2): 389-395. |
| [29] |
李蕊希, 吴雪, 贡璐. 塔里木河上游典型荒漠植物叶片性状及其与土壤因子的关系. 生态学报, 2022, 42(13): 5360-5370. |
| [30] |
董玲玲. 辽河源油松次生林林分结构和功能性状对生产力的影响研究[D]. 北京: 北京林业大学, 2020.
|
| [31] |
Sultan S E. Phenotypic plasticity for plant development, function and life history. Trends in Plant Science, 2000, 5(12): 537-542. DOI:10.1016/S1360-1385(00)01797-0 |
| [32] |
Reich P B, Walters M B, Ellsworth D S, Vose J M, Volin J C, Gresham C, Bowman W D. Relationships of leaf dark respiration to leaf nitrogen, specific leaf area and leaf life-span: a test across biomes and functional groups. Oecologia, 1998, 114(4): 471-482. DOI:10.1007/s004420050471 |
| [33] |
黄林娟, 于燕妹, 安小菲, 余林兰, 薛跃规. 天坑森林植物群落叶功能性状、物种多样性和功能多样性特征. 生态学报, 2022, 42(24): 10264-10275. |
| [34] |
Zhu D H, Hui D F, Wang M Q, Yang Q, Yu S X. Light and competition alter leaf stoichiometry of introduced species and native mangrove species. The Science of the Total Environment, 2020, 738: 140301. DOI:10.1016/j.scitotenv.2020.140301 |
| [35] |
王平安. 华北落叶松不同林龄人工林生态化学计量特征分析[D]. 太谷: 山西农业大学, 2020.
|
| [36] |
Koerselman W, Meuleman A F M. The vegetation N: P ratio: a new tool to detect the nature of nutrient limitation. The Journal of Applied Ecology, 1996, 33(6): 1441. DOI:10.2307/2404783 |
| [37] |
艾也博. 北京松山油松叶功能性状对海拔和土壤的响应[D]. 北京: 北京林业大学, 2018.
|
| [38] |
黄海侠, 杨晓东, 孙宝伟, 张志浩, 阎恩荣. 浙江天童常绿植物当年生与往年生叶片性状的变异与关联. 植物生态学报, 2013, 37(10): 912-921. |
| [39] |
史作民, 唐敬超, 程瑞梅, 罗达, 刘世荣. 植物叶片氮分配及其影响因子研究进展. 生态学报, 2015, 35(18): 5909-5919. |
| [40] |
田地, 严正兵, 方精云. 植物生态化学计量特征及其主要假说. 植物生态学报, 2021, 45(7): 682-713. |