 2024, Vol. 44
2024, Vol. 44文章信息
- 宓桂音, 平原, 赵娅君, 刘福君, 郭忠录
- MI Guiyin, PING Yuan, ZHAO Yajun, LIU Fujun, GUO Zhonglu
- 红壤丘陵区苔藓结皮土壤分离能力季节变化特征
- Seasonal variation of soil detachment capacity induced by moss crusts in red soil hilly area
- 生态学报. 2024, 44(22): 10391-10400
- Acta Ecologica Sinica. 2024, 44(22): 10391-10400
- http://dx.doi.org/10.20103/j.stxb.202402200353
-
文章历史
- 收稿日期: 2024-02-20
- 网络出版日期: 2024-08-26





2. 中国电力工程顾问集团中南电力设计院有限公司, 武汉 430071
2. Central Southern China electric Power Design Institute, China Power Engineering Consulting (Group) Corporation, Wuhan 430071, China
土壤侵蚀是侵蚀动力(降雨、径流)和下垫面(土壤侵蚀阻力)的相互作用过程[1], 而近地表特性直接影响侵蚀动力与下垫面特征, 进而作用于侵蚀过程[2—3]。土壤分离是土壤侵蚀的初始阶段, 是土壤颗粒在雨滴溅蚀或径流冲刷作用下从土体中分离的过程[4]。土壤分离是产生细沟最终形成冲沟的关键过程[5]。当作用于土壤的侵蚀动力超过土壤的临界剪切力时, 土壤开始发生分离, 土壤分离能力随着侵蚀动力或能量的增加而增大。土壤可蚀性和临界剪切力是表征土壤抗蚀性的重要参数。许多物理土壤侵蚀模型中的土壤分离过程都是根据土壤分离能力(Dc)、侵蚀力(τ)和抗侵蚀性(Kr和τc)参数之间的定量关系表示[6—7]。
生物土壤结皮(生物结皮)是由藻类、地衣、苔藓以及微生物与下层土壤颗粒胶结形成的复合层次[8]。作为近地表的主要特征之一, 生物结皮发育势必会对土壤表层水文循环、土壤侵蚀产生重要作用[9—10], 能够改善表土理化性质, 提高土壤抗侵蚀能力[11—12]。已有研究表明, 生物结皮土壤分离能力随盖度增加而显著降低, 土壤黏结力和砂粒含量也影响其变化[13—14]。生物结皮能提高土壤侵蚀阻力, 降低土壤分离能力, 但对其发育节律引起的土壤分离能力变化的研究较少。
研究发现, 生物结皮发育表现出宏观地带性特征。目前, 生物结皮的研究主要集中在干旱半干旱区域, 对湿润区生物结皮研究较少。与干旱半干旱区相比, 南方红壤丘陵区具有降雨量大且降雨量集中、多暴雨、地形破碎度大、母质风化程度高、抗蚀性弱、林下植被覆盖度低等特点[15], 土壤侵蚀模数在3000 t km-2 a-1以上[16], 相对土壤流失量较大。南方红壤丘陵区水热条件较好, 加上经果林下遮阳的同时提供了适当散光, 生物结皮可以在经果林下间隙中迅速生长[17]。且由于生物结皮较难与维管植物竞争[18], 该区生物结皮常生长在地表植被稀少、土壤养分有限的侵蚀劣地, 生物结皮特殊的覆盖层对控制土壤侵蚀产沙具有重要作用, 尤其是对土壤侵蚀初始的分离过程。鉴于此, 本研究从时间尺度开展研究, 分析生物结皮表面特征及其对下伏土壤性质影响, 探究季节变化生物结皮对土壤分离过程的影响及其机制, 对解析红壤丘陵区生物结皮发育增强土壤抗侵蚀性能和影响侵蚀水动力的机理具有重要意义, 可为该区域林下水土流失防治及生物结皮的利用与管理提供理论参考。
1 材料与方法 1.1 研究区概况研究区位于湖北省咸宁市贺胜桥镇(114°21′35″E, 30°01′45″N), 该区域平均海拔44.3 m, 总面积88 km2, 地貌以丘陵为主, 地形东南高、西北低。属亚热带季风气候, 年均气温16.5 ℃, 年均降雨量1484 mm, 主要集中在3—7月, 8—10月份降雨不足20%, 季节性干旱频发[19]。选取土壤类型为第四纪黏土发育的红壤, 土层深厚, 土壤质地较为黏重。主要种植经济林和水稻, 林下结皮广泛生长。受水热条件影响, 研究区样地生物结皮主要为苔藓结皮, 优势种有狭叶白发藓(白发藓科, Leucobryum bowringii Mitt.)、暖地大叶藓(真藓科, Rhodobryum giganteum(Hook.) Par.)、小扭口藓(丛藓科, Semibarbula)等。
1.2 样品采集测定在分别种植樟树(Cinnamomum camphora)和油茶(Camellia oleifera)的人工林内建立观测小区, 从2022年5月至11月, 每月测定苔藓结皮盖度、厚度、苔藓株高、土壤黏结力。土壤分离能力试验用取土环刀(直径为10 cm, 高为6.35 cm)采集。在8月末, 茶园为避免杂草与油茶树竞争水分和养分, 进行手动拔草措施, 对生物结皮产生人工干扰。
苔藓结皮盖度采用网格法测定, 每块样地重复测定20次;结皮厚度和苔藓株高使用电子游标卡尺测量, 每种处理重复测量20次;土壤黏结力(不铲除结皮)通过便携式剪切仪(TORVANE®)测量, 每种处理重复测量15次。
1.3 土壤分离能力测定于2022年5—11月进行实验, 每月中旬定期采集冲刷样品。实验前, 先将样品放入水中8 h达到饱和后, 再静止8 h除去土壤重力水, 保证样品初始含水量一致。为减少边际效应带来的误差, 用玻璃胶涂抹环刀与放样室之间的缝隙。
土壤分离能力试验在变坡冲刷水槽内进行测定。冲刷水槽长2.20 m, 宽0.13 m, 坡度可在0—30 °范围内调节。在水槽底部粘一层试验用土, 以模拟原生地表的糙度。水流表面流速通过高锰酸钾染色法测定, 重复测量10次, 测定水温, 查表得水运动黏滞系数ν, 计算出雷诺数Re, 判断水流流态, 通过校正系数与平均表面流速V得到平均流速V。雷诺数、平均流速、水流深度、剪切力通过下式计算:
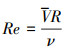
|
(1) |
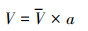
|
(2) |
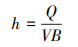
|
(3) |
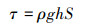
|
(4) |
式中, Re为雷诺数;V为水流表面平均流速(m/s);R为水力半径(m);v为水运动黏滞系数(m2/s);V为水流平均流速(m/s);a为修正系数, 紊流取0.8;h为径流深(m);Q为流量(m3/s);B为水槽宽度(m);τ为水流剪切力(Pa);ρ为水的密度(kg/m3);g为重力加速度(m/s2);S为坡度的正弦值。
通过不同流量和坡度的组合, 本研究选取6组水动力条件进行冲刷试验, 水流剪切力介于7.61—21.08 Pa(表 1), 每组侵蚀动力下重复冲刷5个土样, 每个处理需取30个土样, 共采集并冲刷420个样品。
| 编号 Number |
单宽流量 Flow discharge per unit width/(m2/s) |
水深 Flow depth/mm |
坡度 Slope gradient/% |
平均流速 Mean flow velocity/(m/s) |
流态 Flow regime |
剪切力 Shear stress/Pa |
| 第一组Group 1 | 2.69×10-3 | 4.46 | 10 | 0.56 | 紊流 | 7.61 |
| 第二组Group 2 | 4.08×10-3 | 5.67 | 10 | 0.67 | 紊流 | 9.67 |
| 第三组Group 3 | 4.38×10-3 | 4.87 | 15 | 0.83 | 紊流 | 12.37 |
| 第四组Group 4 | 6.77×10-3 | 6.11 | 15 | 1.03 | 紊流 | 15.52 |
| 第五组Group 5 | 6.00×10-3 | 5.39 | 20 | 1.03 | 紊流 | 18.05 |
| 第六组Group 6 | 6.85×10-3 | 5.62 | 22.5 | 1.13 | 紊流 | 21.08 |
冲刷开始后, 土样被冲刷约2 cm时停止冲刷, 记录冲刷时间(t), 将冲刷后的土样放置于烘箱105 ℃下烘至恒重, 计算可得到土壤分离能力, 根据WEPP模型计算可得细沟可蚀性:
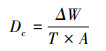
|
(5) |
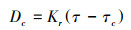
|
(6) |
式中, Dc为土壤分离能力(kg m-2 s-1);ΔW为分离土壤的烘干重(kg);T为冲刷历时(s), 冲刷时间上限5 min;A为冲刷环刀的面积(m2);Kr为细沟可蚀性(s/m);τ和τc分别为水流剪切力(Pa)和临界剪切力(Pa)。以水流剪切力为横坐标、以实测的土壤分离能力为纵坐标, 进行线性拟合, 则拟合直线的斜率为细沟可蚀性(Kr), 而拟合直线在横坐标上的截距为临界剪切力(τc)。
1.4 数据处理使用Excel 2016和SPSS 26对数据进行处理与分析, 应用单因素方差分析不同月度下苔藓结皮表面特征、土壤分离能力差异;简单回归分析拟合苔藓结皮盖度与厚度、土壤黏结力、相对土壤分离能力、细沟可蚀性之间关系;非线性回归拟合土壤分离能力与水动力学特性、苔藓结皮表面特征之间的关系。使用Origin 2023软件绘图。
2 结果与分析 2.1 不同地类苔藓结皮表面特征的季节变化研究区5—11月的气温及降雨量由华中农业大学红壤综合试验站气象站自动监测, 总降雨量为479.6 mm, 最大日降雨量为52.1 mm(图 1)。4—7月降雨较多, 8—11月降雨较少。

|
| 图 1 研究区2022年5—11月降雨量和气温概况 Fig. 1 Study area rainfall and temperature profile for May—November 2022 |
从图 2可得, 两种地类苔藓结皮的结皮盖度、厚度、株高、土壤黏结力均呈先上升后下降再上升趋势, 而相关性分析表明(表 2), 苔藓结皮表面特征的月度变化均与月降雨量显著相关(P<0.01)。其中, 樟树林、油茶园结皮盖度变化范围分别为4%—85%、9%—99%;结皮厚度变化范围分别为2.49—12.03 mm、5.58—9.03 mm;株高变化范围分别为0.54—1.76 mm、0—6.24 mm;土壤黏结力变化范围分别为1.08—20.58 kPa、1.47—26.46 kPa。5—7月份, 水热条件适宜, 苔藓结皮表面特征各指标呈上升趋势。8—10月研究区出现持续高温干旱情况, 各指标呈下降趋势。10月份后研究区出现降雨, 各指标再次回升。

|
| 图 2 樟树林结皮及油茶园结皮盖度、厚度、株高、土壤黏结力季节变化 Fig. 2 Seasonal changes of coverage, thickness, plant height and soil cohesion of moss crusts under camphor forest and moss crusts under oil tea plantation 图中不同小写字母表示不同月份间差异显著(P<0.05) |
| 影响因素 Influencing factors |
结皮盖度 Coverage of crusts |
结皮厚度 Thickness of crusts |
苔藓株高 Plant height of moss |
土壤黏结力 Soil cohesion |
| 降雨量Rainfall | 0.840** | 0.808** | 0.797** | 0.811** |
| * *表示在0.01级别上显著相关 | ||||
受季节变化影响, 两种地类的苔藓结皮土壤分离能力均呈现先增大后减小趋势(P<0.05)(图 3)。分离能力最大值均出现在9月, 樟树林结皮的平均土壤分离能力(0.2107 kg m-2 s-1)大于油茶园结皮(0.1924 kg m-2 s-1), 但两者无显著差异(P≥0.05)。由于8月油茶园出现人工干扰, 土壤分离能力较樟树林结皮大幅度增大。

|
| 图 3 樟树林结皮和油茶园结皮土壤分离能力季节变化 Fig. 3 Seasonal variation in soil detachment capacity of moss crusts under camphor forest and moss crusts under oil tea plantation |
根据WEEP模型, 将土壤分离能力与水流剪切力进行线性拟合, 得到土壤侵蚀阻力(表 3)。樟树林、油茶园结皮覆盖土壤细沟可蚀性分别介于0.0143—0.0659 s/m、0.0015—0.0946 s/m。两地类苔藓结皮细沟可蚀性均表现为先增加后减小, 与季节变化趋势相似, 但9月份樟树林结皮细沟可蚀性较上月份减小39.15%, 与土壤分离能力变化不符, 这可能与数据精度及线性拟合存在误差有关。樟树林结皮的平均细沟可蚀性(0.0374 s/m)小于油茶园结皮(0.0394 s/m), 但两者无显著差异。分析临界剪切力发现, 樟树林结皮和油茶园结皮临界剪切力均呈现无规律变化趋势。
| 不同地类苔藓结皮 Moss crusts of different land types |
月度 Month |
回归方程 Regression equation |
细沟可蚀性 Rill erodibility/(s/m) |
临界剪切力 Critical shear stress/Pa |
R2 |
| 樟树林结皮 | 5 | Dc=0.0216τ-0.2011 | 0.0216 | 9.31 | 0.819 |
| Moss crusts under camphor forest | 6 | Dc=0.0143τ-0.1441 | 0.0143 | 10.08 | 0.607 |
| 7 | Dc=0.0323τ-0.2860 | 0.0323 | 8.85 | 0.882 | |
| 8 | Dc=0.0659τ-0.5806 | 0.0659 | 8.81 | 0.709 | |
| 9 | Dc=0.0401τ-0.1933 | 0.0401 | 4.82 | 0.927 | |
| 10 | Dc=0.0521τ-0.4376 | 0.0521 | 8.40 | 0.808 | |
| 11 | Dc=0.0353τ-0.3212 | 0.0353 | 9.10 | 0.839 | |
| 油茶园结皮 | 5 | Dc=0.0015τ-0.0086 | 0.0015 | 5.73 | 0.841 |
| Moss crusts under oil tea plantation | 6 | Dc=0.0021τ-0.0198 | 0.0021 | 9.43 | 0.703 |
| 7 | Dc=0.0195τ-0.1595 | 0.0195 | 8.18 | 0.905 | |
| 8 | Dc=0.0485τ-0.4689 | 0.0485 | 9.67 | 0.738 | |
| 9 | Dc=0.0946τ-0.8041 | 0.0946 | 8.50 | 0.913 | |
| 10 | Dc=0.0668τ-0.6043 | 0.0668 | 9.05 | 0.872 | |
| 11 | Dc=0.0427τ-0.4101 | 0.0427 | 9.60 | 0.785 |
苔藓结皮发育程度(结皮盖度、厚度、株高)和结皮土壤黏结力影响土壤分离能力, 三者均为苔藓结皮发育程度量化指标。土壤分离能力随结皮盖度、厚度、株高增加呈现指数函数衰减(图 4), 决定系数R2在0.741—0.946之间, 表明随着苔藓结皮发育程度增加, 导致土壤分离能力逐渐下降。对比两地, 樟树林结皮的土壤分离能力与影响因子的拟合效果均较好, 而油茶园结皮的土壤分离能力与结皮厚度和株高的拟合方程中常数量过大, 这导致在厚度和株高较小时, 拟合得到的土壤分离能力过大。

|
| 图 4 土壤分离能力与结皮表面特征之间的关系 Fig. 4 Relationship between soil detachment capacity and crust surface characteristics |
采用非线性回归的方法, 选取决定系数相对较高的土壤黏结力和结皮盖度, 与水动力学参数水流剪切力(τ)共同对土壤分离能力进行拟合(图 5), 纳什效率系数(Nash-Sutcliffe Efficiency, NSE)评价拟合方程。结果表明模拟值与实测值均匀分布于1∶1线两侧, 拟合程度较好。拟合方程能够解释土壤分离能力72.8%的变异, 总体而言, 该模型高估了2.22%的土壤分离能力。

|
| 图 5 土壤分离能力实测值与模拟值之间的比较 Fig. 5 Comparison between measured and modeled values of soil detachment capacity x1:土壤粘结力Soil cohesion; x2:结皮盖度Coverage of crusts |
受气候、降雨、温度等环境因子季节性变化的影响, 苔藓结皮表面特征(盖度、厚度、株高、土壤黏结力)也呈现出季节节律[20—21], 目前关于苔藓结皮季节动态变化的研究多集中于微生物群落及土壤酶活性变化[22—23], 对于苔藓结皮表面特征季节变化的研究相对薄弱。
本研究结果表明, 不同地类苔藓结皮及月度变化显著影响结皮表面特征, 与水热条件变化密切相关(表 2)。季节变化本质上是水热条件的变化。与干旱半干旱区绝对干旱不同, 研究区总体降雨并不少, 春夏之交降雨丰富, 且强度较大, 易发生坡面水蚀, 夏末秋初高温少雨, 降雨分配不均导致季节性干旱严重[24—25]。5—7月份, 水热条件适宜时, 苔藓结皮维持较好发育状态, 生长迅速, 结皮盖度、厚度、株高增加;8—9月份无降雨发生, 出现季节性干旱, 苔藓属于变水植物, 变水特征决定含水量随环境变化而改变, 当环境干旱时苔藓结皮迅速失水呈干燥状态[26], 苔藓结皮表层退化甚至死亡, 代谢活动及微生物活动降低, 苔藓结皮近地表特征随之改变, 结皮厚度、盖度、株高减小;10月份及11月份产生降雨, 解除高温干旱, 苔藓结皮结束休眠状态, 并再次发育生长。土壤黏结力受到季节、降水、日照及结皮表面特征等多种因素影响[27], 苔藓结皮通过菌丝缠绕、多糖分泌物等对土壤颗粒进行胶结、捆绑和吸附, 使土壤黏结力显著增强[28], 因此土壤黏结力季节变化趋势与苔藓结皮发育季节变化趋势相似。
3.2 苔藓结皮发育对土壤分离能力的季节变化特性影响诸多研究表明, 土壤分离能力存在季节性变化规律, 但前人研究重点关注草地、农作物等地表覆被物[29—30], 对于地表生物结皮季节变化的研究较少。南方红壤丘陵区林下植被稀疏, 苔藓结皮广泛存在, 人工干扰较少, 苔藓结皮发育受环境因子影响较大, 月度降雨量等差异导致苔藓结皮发育程度在月度间差异较大(发育或退化), 进而导致土壤分离能力的月度变化响应。
本研究表明, 土壤分离能力随苔藓结皮发育程度增加而减小。苔藓结皮能改变地表微形态, 增加地表粗糙度, 减少地表径流量和径流速度, 从而提升土壤抗蚀性能[9]。苔藓结皮的菌丝和假根在土壤中形成密集的地下网络组织, 将土壤颗粒紧密连接, 能够有效稳定土壤[31]。苔藓结皮表面特征与降雨量显著相关, 水分是制约苔藓结皮发育主要环境限制因子。在水热条件适宜情况下, 苔藓结皮表面特征发育良好, 土壤分离能力减小。在干旱缺水的环境下, 苔藓结皮迅速退化进入休眠状态甚至死亡, 导致土壤分离能力在短时间内提高。但持续的干旱条件不会让土壤分离能力持续增长, 苔藓结皮退化或休眠后, 地表会形成一层硬壳状态, 且下层结皮的菌丝和假根仍然可以固定土壤, 因此土壤分离能力会先迅速增加, 然后逐渐缓慢增长。在本研究的时间尺度上, 由于油茶园的油茶树受到干旱胁迫, 管理人员除草扰动地表, 受到扰动的苔藓结皮进一步退化甚至死亡, 盖度和厚度均有所下降, 土壤分离能力大幅提升, 因此人类活动干扰也能显著影响苔藓结皮生长发育, 进而影响土壤分离能力。
细沟可蚀性和临界剪切力是基于WEPP模型的推导参数, 反映土壤对集中水流侵蚀的抵抗能力[32]。本研究中细沟可蚀性与土壤分离能力变化趋势一致, 但对临界剪切力研究发现, 临界剪切力在月度下呈现无规律变化, 这可能是临界剪切力与土壤性质更为相关, 本研究较小时间尺度上苔藓结皮发育不足以引起临界剪切力的改变[33—34]。
在维管植物存在区域, 可以通过水动力侵蚀、植物根系特征及土壤属性三者对土壤分离能力进行较高精度的拟合[35—36]。对于生物结皮, 有学者通过水流剪切力、结皮厚度、土壤黏结力等指标与土壤分离能力拟合[37—38]。但在本研究中, 分析发现土壤分离能力与苔藓结皮盖度之间的相关系数更大, 结皮盖度更能反映土壤分离能力的变化情况, 因此本研究采用水流剪切力、结皮盖度、土壤黏结力对土壤分离能力进行模拟, 模型精度NSE=0.728, 能够较好解释该区域内经果林下苔藓结皮发育土壤分离能力的季节变化。
4 结论(1) 苔藓结皮表面特征随季节变化显著(P<0.05), 在5—11月, 结皮盖度、厚度、株高、土壤黏结力均呈现先增加后减小再增加的趋势, 苔藓结皮生长发育与月度降雨量显著相关。
(2) 土壤分离能力与苔藓结皮表面特征呈显著负相关(P<0.05), 随苔藓结皮发育程度增加而减小;制约苔藓结皮发育的主要环境限制因子为水分, 因此土壤分离能力的季节性变化与月度降雨量变化有一定相关性。水热适宜情况下苔藓结皮发育迅速, 固结土壤的能力变强, 土壤分离能力下降。
(3) 细沟可蚀性与土壤分离能力变化趋势一致, 临界剪切力在月度下呈现无规律变化。这可能是临界剪切力与土壤性质更为相关, 本研究较小时间尺度上苔藓结皮发育不足以引起临界剪切力的改变。
(4) 多元非线性回归表明, 水流剪切力、苔藓结皮盖度、土壤黏结力能够较好模拟土壤分离能力的季节变化, 模型精度NSE=0.728, 可较好解释该区域经果林下苔藓结皮发育下土壤分离能力的季节变化。
| [1] |
史志华, 宋长青. 土壤水蚀过程研究回顾. 水土保持学报, 2016, 30(5): 1-10. |
| [2] |
Liu Y, Cui Z, Huang Z, Miao H T, Wu G L. The influence of litter crusts on soil properties and hydrological processes in a sandy ecosystem. Hydrology and Earth System Sciences, 2019, 23(5): 2481-2490. DOI:10.5194/hess-23-2481-2019 |
| [3] |
张光辉. 退耕驱动的近地表特性变化对土壤侵蚀的潜在影响. 中国水土保持科学, 2017, 15(4): 143-154. |
| [4] |
Li Z, Zhang G, Geng R, Wang H, Zhang X C. Land use impacts on soil detachment capacity by overland flow in the Loess Plateau, China. Catena, 2015, 124: 9-17. DOI:10.1016/j.catena.2014.08.019 |
| [5] |
Zhang G H, Liu B Y, Liu G B, He X W, Nearing M A. Detachment of undisturbed soil by shallow flow. Soil Science Society of America Journal, 2003, 67(3): 713-719. DOI:10.2136/sssaj2003.7130 |
| [6] |
Liu J X, Chen L D, Wang B, Peng X Y. Effects of physical crust on soil detachment by overland flow in the Loess Plateau region of China. International Soil and Water Conservation Research, 2024, 12(1): 107-120. DOI:10.1016/j.iswcr.2023.05.006 |
| [7] |
Wang D D, Wang Z L, Shen N, Chen H. Modeling soil detachment capacity by rill flow using hydraulic parameters. Journal of Hydrology, 2016, 535: 473-479. DOI:10.1016/j.jhydrol.2016.02.013 |
| [8] |
Kidron G J. The dual effect of sand-covered biocrusts on annual plants: Increasing cover but reducing individual plant biomass and fecundity. Catena, 2019, 182: 104120. DOI:10.1016/j.catena.2019.104120 |
| [9] |
Guida G, Nicosia A, Settanni L, Ferro V. A review on effects of biological soil crusts on hydrological processes. Earth-Science Reviews, 2023, 243: 104516. DOI:10.1016/j.earscirev.2023.104516 |
| [10] |
Gao L Q, Bowker M A, Sun H, Zhao J, Zhao Y G. Linkages between biocrust development and water erosion and implications for erosion model implementation. Geoderma, 2020, 357: 113973. DOI:10.1016/j.geoderma.2019.113973 |
| [11] |
赵允格, 许明祥, 王全九, 邵明安. 黄土丘陵区退耕地生物结皮对土壤理化性状的影响. 自然资源学报, 2006, 21(3): 441-448. |
| [12] |
肖波, 赵允格, 邵明安. 陕北水蚀风蚀交错区两种生物结皮对土壤理化性质的影响. 生态学报, 2007, 27(11): 4662-4670. DOI:10.3321/j.issn:1000-0933.2007.11.034 |
| [13] |
Liu F, Zhang G H, Sun L, Wang H. Effects of biological soil crusts on soil detachment process by overland flow in the Loess Plateau of China. Earth Surface Processes and Landforms, 2016, 41(7): 875-883. DOI:10.1002/esp.3870 |
| [14] |
张冠华, 易亮, 丁文峰, 王一然, 蒲坚, 孙宝洋. 三峡库区生物结皮对土壤分离过程的影响及其机制. 土壤, 2021, 53(3): 610-619. |
| [15] |
梁音, 杨轩, 潘贤章, 张斌, 史德明. 南方红壤丘陵区水土流失特点及防治对策. 中国水土保持, 2008(12): 50-53. |
| [16] |
范亚琳, 刘贤赵, 高磊, 汪亚及, 彭新华. 不同培肥措施对红壤坡耕地土壤有机碳流失的影响. 土壤学报, 2019, 56(3): 638-649. |
| [17] |
Seitz S, Nebel M, Goebes P, Käppeler K, Schmidt K, Shi X Z, Song Z S, Webber C L, Weber B, Scholten T. Bryophyte-dominated biological soil crusts mitigate soil erosion in an early successional Chinese subtropical forest. Biogeosciences, 2017, 14(24): 5775-5788. DOI:10.5194/bg-14-5775-2017 |
| [18] |
Corbin J D, Thiet R K. Temperate biocrusts: mesic counterparts to their better-known dryland cousins. Frontiers in Ecology and the Environment, 2020, 18(8): 456-464. DOI:10.1002/fee.2234 |
| [19] |
林丽蓉, 陈家宙, 王峰, 邓盛华. 几种保护措施对红壤坡地水文过程及干旱的影响. 中国农业科学, 2014, 47(24): 4858-4867. |
| [20] |
Orlovsky L, Dourikov M, Babaev A. Temporal dynamics and productivity of biogenic soil crusts in the central Karakum desert, Turkmenistan. Journal of Arid Environments, 2004, 56(4): 579-601. DOI:10.1016/S0140-1963(03)00075-2 |
| [21] |
Zhang X C, Li J Y, Liu J L, Yuan C X, Li Y N, Liu B R, Yan X F. Temporal shifts in cyanobacterial diversity and their relationships to different types of biological soil crust in the southeastern Tengger Desert. Rhizosphere, 2021, 17: 100322. DOI:10.1016/j.rhisph.2021.100322 |
| [22] |
刘哲, 叶兴旺, 王吉平, 程永韬, 钱隆, 肖敬尚, 吴丽. 生物结皮胞外多糖理化特性及菌群结构的季节动态. 应用生态学报, 2022, 33(7): 1801-1809. |
| [23] |
王凯, 厉萌萌, 刘德权, 张凌基, 刘可欣, 许华, 唐亮, 何明珠, 张珂. 腾格里沙漠不同组成生物结皮特征及其对土壤酶活性的影响. 生态学报, 2022, 42(14): 5859-5868. |
| [24] |
王峰, 李萍, 陈家宙. 亚热带红壤坡地季节性干旱空间特征. 土壤通报, 2016, 47(4): 820-826. |
| [25] |
Chen J Z, Lin L R, Lü G A. An index of soil drought intensity and degree: An application on corn and a comparison with CWSI. Agricultural Water Management, 2010, 97(6): 865-871. DOI:10.1016/j.agwat.2010.01.017 |
| [26] |
张元明, 曹同, 潘伯荣. 干旱与半干旱地区苔藓植物生态学研究综述. 生态学报, 2002, 22(7): 1129-1134. |
| [27] |
唐科明, 张光辉, 孙珍玲. 草地土壤分离能力季节变化特征及其影响因素. 中国水土保持科学, 2016, 14(6): 18-25. |
| [28] |
赵允格, 许明祥, 王全九, 邵明安. 黄土丘陵区退耕地生物结皮理化性状初报. 应用生态学报, 2006, 17(8): 1429-1434. |
| [29] |
Wang B, Zhang G H, Shi Y Y, Zhang X C, Ren Z P, Zhu L J. Effect of natural restoration time of abandoned farmland on soil detachment by overland flow in the Loess Plateau of China. Earth Surface Processes and Landforms, 2013, 38(14): 1725-1734. DOI:10.1002/esp.3459 |
| [30] |
王长燕, 郁耀闯. 黄土丘陵区退耕草地土壤分离能力季节变化研究. 土壤学报, 2016, 53(4): 1047-1055. |
| [31] |
Belnap J. The potential roles of biological soil crusts in dryland hydrologic cycles. Hydrological Processes, 2006, 20(15): 3159-3178. |
| [32] |
Chen N, Wang X P, Zhang Y F, Yu K L, Zhao C M. Ecohydrological effects of biological soil crust on the vegetation dynamics of restoration in a dryland ecosystem. Journal of Hydrology, 2018, 563: 1068-1077. |
| [33] |
Wang H, Zhang G H, Liu F, Geng R, Wang L J. Temporal variations in infiltration properties of biological crusts covered soils on the Loess Plateau of China. Catena, 2017, 159: 115-125. |
| [34] |
许欢欢, 张宝琦, 汪建芳, 王兵. 黄土高原典型生物结皮对坡面产流产沙过程的影响. 水土保持通报, 2020, 40(6): 8-13. |
| [35] |
Zi R Y, Zhao L S, Fang Q, Qian X H, Fang F Y, Fan C H. Path analysis of the effects of hydraulic conditions, soil properties and plant roots on the soil detachment capacity of karst hillslopes. Catena, 2023, 228: 107177. |
| [36] |
Wang B, Zhang G H, Yang Y F, Li P P, Liu J X. Response of soil detachment capacity to plant root and soil properties in typical grasslands on the Loess Plateau. Agriculture, Ecosystems & Environment, 2018, 266: 68-75. |
| [37] |
Zhang G H, Ding W F, Pu J, Li J M, Qian F, Sun B Y. Effects of moss-dominated biocrusts on soil detachment by overland flow in the Three Gorges Reservoir Area of China. Journal of Mountain Science, 2020, 17(10): 2418-2431. |
| [38] |
平原, 澹腾辉, 李雨晨, 曾建辉, 宓桂音, 谭林芳, 郭忠录. 丹江口库区典型小流域生物结皮驱动土壤性质变化及分离响应. 水土保持学报, 2023, 37(3): 87-94. |