 2024, Vol. 44
2024, Vol. 44文章信息
- 汪美宏, 赵慧, 吴晓兰, 陈兆杰, 姜世中
- WANG Meihong, ZHAO Hui, WU Xiaolan, CHEN Zhaojie, JIANG Shizhong
- 考虑气候时间效应的额尔齐斯河流域植被动态变化及其驱动力
- Vegetation dynamics and its driving force in the Irtysh River Basin considering climatic temporal effects
- 生态学报. 2024, 44(22): 10352-10366
- Acta Ecologica Sinica. 2024, 44(22): 10352-10366
- http://dx.doi.org/10.20103/j.stxb.202312262822
-
文章历史
- 收稿日期: 2023-12-26
- 网络出版日期: 2024-08-22





2. 中国科学院、水利部成都山地灾害与环境研究所, 山地表生过程与生态调控重点实验室, 成都 610299;
3. 新疆大学生态与环境学院, 乌鲁木齐 830017;
4. 新疆地矿局第一水文工程地质大队,乌鲁木齐 830091
2. Key Laboratory of Mountain Surface Processes and Ecological Regulation, Institute of Mountain Hazards and Environment, Chinese Academy of Sciences, Chengdu 610299, China;
3. School of Ecology and Environment, Xinjiang University, Urumqi 830017, China;
4. The First Hydrologic Engineering Geology Group, Xinjiang Bureau of Geology and Mineral Resources, Urumqi 830091, China
植被作为陆地生态系统的重要组成部分, 对平衡碳收支、维持气候稳定有重要贡献。近年来研究表明气候和人为因素的耦合效应是区域植被变化的主要驱动因素[1—2], 其中干旱和半干旱地区的植被生长受到的水分压力逐渐增加[3—4], 对气候变化的响应敏感复杂, 农耕、放牧和生态保护工程等人类活动对植被动态变化的速度和程度的影响日益显著[5—6]。植被生长受到年度和年际的气候条件影响, 认识气候对植被生长影响的时间效应是至关重要的[7—8], 忽略时间效应会导致植被动态归因的偏差[9]。气候的时间效应包括滞后和累积效应两个方面, 滞后效应是指植被生长明显受到先前月份气候条件的影响, 累积效应是指植被生长受到先前月份和当月累积气候条件的影响。现有研究表明, 降水、气温的滞后和累积作用能在短时期内对干旱半干旱区植被生长起促进作用, 考虑时间效应的模型比忽略时间效应的模型大幅度提高了气候因素的总体解释力[10—11], 能够更合理明确影响植被动态的气候作用。可见滞后效应和累积效应是影响驱动力量化的重要因素[12—14]。
近年来, 部分干旱半干旱地区研究仅将植被动态与气候的滞后效应联系起来[15], 或主要集中于当前气候因子对植被的影响[16], 未考虑前几个月累积气候条件的作用, 缺乏对滞后和累积耦合效应的研究, 未能全面捕捉植被生长与气候因子的相关特征。目前气候时间效应的相关研究也更多集中在全球、全国或青藏高原等大尺度范围内[7], 难以详细表征干旱半干旱局部重点区域。
额尔齐斯河流域属温带大陆性干旱半干旱气候, 降水分布不均, 植被覆盖率低且具有典型的半干旱区植被特征, 对气候变化响应敏感。以往的研究缺少植被对气候响应时间效应的系统分析[15—17], 植被变化的驱动因素需进一步量化。本文通过探究额尔齐斯河流域植被对降水(月降水量, 下同)和气温(月均温, 下同)响应的时间效应, 进一步量化气候变化对植被动态的贡献率, 明确干旱半干旱区植被动态和气候变化之间的关系, 以期为生态保护提供更准确的数据支撑和管理依据。
1 材料与方法 1.1 研究区概况额尔齐斯河发源于阿尔泰山脉的西南坡, 位于新疆维吾尔自治区北部的阿勒泰地区, 是鄂毕河最大的支流, 地处中国西北内陆(85.53°—91.08°E, 45.00°—49.19°N), 海拔高度为366—4297m, 地形以山地和平原为主, 土壤以棕钙土和栗钙土为主, 年均降水量为107.11—579.32mm,年均温为-12.82—7.22℃, 是干旱半干旱区的相对丰水区。额尔齐斯河流域生态系统脆弱敏感, 是我国的重点生态功能区, 主要生态系统类型为草地、森林、湿地、农田(图 1), 其中草地建群种主要包括沙生针茅、糙隐子草等;森林建群种主要包括西伯利亚落叶松、西伯利亚云杉等;湿地建群种主要包括芦苇、藨草、香蒲等;农田主要种植棉花、油葵等。

|
| 图 1 额尔齐斯河流域地理位置及植被类型分布 Fig. 1 Geographic location and distribution of vegetation types in Irtysh River Basin |
利用Theil-Sen Median趋势分析方法计算NDVI变化趋势, 此方法计算效率高, 对缺失时间序列和异常观测值不太敏感[18]。计算公式如下:
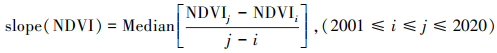
|
(1) |
式中, slope(NDVI)表示生长季归一化植被指数NDVI在时间序列内的变化率;i和j表示年份的序数;NDVIi和NDVIj分别为i和j时刻的NDVI值。
1.3 Mann-Kendall(MK)检验使用Mann-Kendall(MK)趋势检验来确定趋势变化是否显著, 将通过信度为90%、95%、99%显著性检验的NDVI变化趋势分别定义为微显著、显著、极显著变化, 其余为不显著变化[19]。对于NDVI时间序列, 定义检验统计量S:
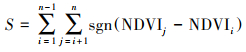
|
(2) |
其中, sgn为符号函数:
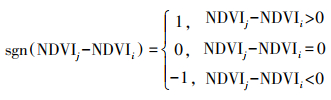
|
(3) |
计算方差:
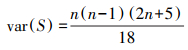
|
(4) |
式中, n为时间序列长度。将S标准化为统计检验值Z, 利用Z进行显著性检验:
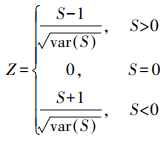
|
(5) |
采用双边趋势检验, 在给定置信水平α下, 当|Z|>Z(1-α)/2时, 表明NDVI在研究时段内发生了显著变化, 反之为无显著变化。本文分别给定置信水平α为0.10、0.05和0.01, 当|Z|大于1.65、1.96和2.58时, 表示NDVI变化趋势分别通过了信度为90%、95%和99%的显著性检验。NDVI变化趋势显著性的判别方法见表 1。
| slope(NDVI) | Z | NDVI变化趋势 Trends in NDVI |
slope(NDVI) | Z | NDVI变化趋势 Trends in NDVI |
|
| slope(NDVI)>0 | 2.58 < Z | 极显著增加 | slope(NDVI) < 0 | Z≤1.65 | 不显著减小 | |
| 1.96 < Z≤2.58 | 显著增加 | 1.65 < Z≤1.96 | 微显著减小 | |||
| 1.65 < Z≤1.96 | 微显著增加 | 1.96 < Z≤2.58 | 显著减小 | |||
| Z≤1.65 | 不显著增加 | 2.58 < Z | 极显著减小 | |||
| NDVI:归一化植被指数Normalized difference vegetation index | ||||||
采用多元线性回归模型分析气候变量影响NDVI的时间效应。植被对气候变化的响应通常发生在一个季度以内, 即时间效应最大为3个月[7], 根据每个像元的多元线性回归模型的最高决定系数R2确定最佳拟合时间效应。计算公式如下:
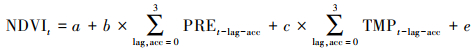
|
(6) |
其中, a为常数项;b、c为回归系数;e为残差;t代表生长季节;PRE为降水;TMP为气温;lag和acc(取值范围为0—3)分别代表气候变量对植被影响的滞后月份和积累月份。lag和acc的各种组合(lag+acc≤3)反映时间效应所有情景, 包括无时间效应、考虑时间滞后效应、考虑时间累积效应以及滞后和累积的耦合效应4种情况, 本文将同时考虑滞后、累积和耦合效应的模型确定为考虑最佳时间效应的模型。
1.5 偏相关分析偏相关分析有助于区分气候变量对植被变化的独立作用。在时间效应的基础上, 采用确定了最佳时间效应的气候数据, 分析气候因子与NDVI的关系, 采用显著性t检验, 并将偏相关系数绝对值较高的对应气候变量认定为影响像元的主导因素[1]。主要计算步骤如下:
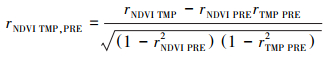
|
(7) |
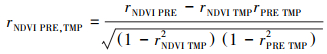
|
(8) |
其中, TMP为温度, PRE为降水, rNDVI TMP, PRE为NDVI与温度的偏相关系数, rNDVI PRE, TMP为NDVI与降水的偏相关系数。
1.6 改进的残差分析以往残差分析中采用多元线性回归计算NDVI的预测值时, 忽略了气候时间效应, 且大多数没有较好地将气候变化和人类活动因素分开[20—21], 导致计算的NDVI预测值并不能精确量化仅受气候变化影响下的植被生长状况。本文为了尽可能多地排除NDVI预测值中人类活动的影响, 使用考虑最佳时间效应的改进残差分析查明额尔齐斯河流域植被变化的驱动力贡献率, 并选取R2>0.5的区域(即气候因子对NDVI的解释度在50%以上)来确定多元线性回归模型的回归系数和常数, 计算研究区的NDVI预测值和残差值, 量化气候变化和人类活动对NDVI变化的贡献。
NDVI预测值的计算见公式(6), NDVI残差值计算公式如下:
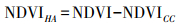
|
(9) |
其中, NDVICC代表NDVI预测值, 即仅受气候变化影响的NDVI;NDVIHA代表NDVI残差值, 即受人类活动影响的NDVI。气候和人类活动对植被NDVI变化的贡献率计算依据见表 2所示。
| NDVI变化趋势 Trends in NDVI |
驱动力 Driving force |
分类Categorization | 贡献率Contribution rate | |||
| slope(NDVICC) | slope(NDVIHA) | 气候变化 >Climate change |
人类活动 >Human activity |
|||
| 增加Increase | 气候变化和人类>活动 | >0 | >0 | 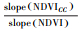 |
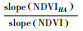 |
|
| 气候变化 | >0 | < 0 | 100 | 0 | ||
| 人类活动 | < 0 | >0 | 0 | 100 | ||
| 减小Reduction | 气候变化和人类活动 | < 0 | < 0 | 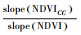 |
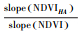 |
|
| 气候变化 | < 0 | >0 | 100 | 0 | ||
| 人类活动 | >0 | < 0 | 0 | 100 | ||
植被NDVI数据从NASA MOD13Q1数据集(https://search.earthdata.nasa.gov/search)获取, 时间分辨率为16天, 空间分辨率为250 m。采用最大值合成法得出NDVI每月最大数据, 然后计算生长季(5—10月)平均NDVI值, 为了减小水体和积雪对分析结果的影响, 排除了多年平均NDVI值小于0.1的像元。气候数据源于国家科技基础条件平台—国家地球系统科学数据中心(http://www.geodata.cn), 包括降水和气温[22], 时间分辨率为逐月, 空间分辨率为1 km。土地利用数据采用武汉大学杨杰、黄昕的土地覆盖数据集(annual China Land Cover Dataset, CLCD)中2020年的数据, 空间分辨率为30 m[23]。对数据进行时间和空间重采样。
本文结合Theil-Sen Median趋势和Mann-Kendall检验方法, 查明额尔齐斯河流域2001—2020年间生长季(5—10月)NDVI变化趋势, 采用多元线性回归模型探究植被对气候变化响应的时间效应, 基于偏相关分析揭示具有最佳时间效应的气候因子对NDVI的影响情况, 并进一步改进残差分析量化气候变化和人类活动对NDVI变化的贡献率。技术路线如图 2所示。

|
| 图 2 研究方法与技术流程 Fig. 2 Research methodology and technical process |
2001—2020年间, 额尔齐斯河流域NDVI整体上以0.0017/a的速率呈显著增加趋势(P < 0.05)(图 3)。从空间变化上看(图 4), NDVI增加和减小区域分别占73.16%和26.84%。其中, 极显著增加(P < 0.01)的区域分布于额尔齐斯河河谷和乌伦古河河谷附近, 显著增加(P < 0.05)的区域分布于北部阿尔泰山脉;极显著减小(P < 0.01)的区域分布于流域西南部古尔班通古特荒漠, 显著减小(P < 0.05)的区域分布于阿尔泰山山麓。四种植被类型NDVI都以增加趋势为主(图 5), 草地、森林、湿地和农田的增长率分别为0.0019/a、0.0014/a、0.0068/a、0.0117/a。其中草地NDVI有76.26%的区域呈增加趋势, 显著增加(P < 0.05)和极显著增加(P < 0.01)的区域分别占草地总面积的9.43%和7.46%;森林NDVI有85.34%的区域呈增加趋势, 显著增加(P < 0.05)和极显著增加(P < 0.01)的区域分别占森林总面积的15.45%和10.71%;湿地NDVI有81.25%的区域呈增加趋势, 显著增加(P < 0.05)和极显著增加(P < 0.01)的区域分别占湿地总面积的6.25%和41.67%;农田NDVI有85.77%的区域呈增加趋势, 显著增加(P < 0.05)和极显著增加(P < 0.01)的区域分别占农田总面积的14.46%和42.49%。

|
| 图 3 额尔齐斯河流域NDVI时间变化趋势 Fig. 3 Temporal trends of NDVI in the Irtysh River Basin |

|
| 图 4 额尔齐斯河流域NDVI空间变化趋势 Fig. 4 Spatial trends in NDVI in the Irtysh River Basin |

|
| 图 5 不同植被NDVI空间变化趋势 Fig. 5 Spatial trends of NDVI for different vegetation |
额尔齐斯河流域植被对降水变化响应的时间效应如图 6所示。降水对植被的影响平均滞后期为(1.92±0.88)月, 平均累积期为(1.90±0.76)月。流域北部阿尔泰山脉植被对降水响应以累积3个月和滞后1月累积2月的耦合效应为主, 南部古尔班通古特荒漠植被对降水响应主要滞后3个月。对于降水滞后效应, 草地平均滞后(1.77±0.86)月, 森林平均滞后(1.89±0.90)月, 湿地平均滞后(1.76±0.79)月, 农田平均滞后(1.82±0.85)月;对于降水累积效应, 草地平均累积(1.91±0.73)月, 森林平均累积(1.84±0.77)月, 湿地平均累积(1.79±0.92)月, 农田平均累积(1.80±0.79)月(图 7)。森林对降水响应的滞后期最长, 对降水变化的反应最不敏感;草地对降水效应的累积期最长, 需要更多时间来累积水分以达到生长条件。

|
| 图 6 额尔齐斯河流域植被对降水、气温响应的时间效应 Fig. 6 Temporal effects of vegetation response to precipitation and temperature in the Irtysh River Basin |

|
| 图 7 不同植被对降水和气温响应的时间效应 Fig. 7 Temporal effects of different vegetation on precipitation and temperature responses |
额尔齐斯河流域植被对气温变化响应的时间效应如图 6所示。气温对植被的影响平均滞后期为(1.70±0.75)月, 平均累积期为(1.57±0.70)月。流域北部阿尔泰山脉植被对气温响应以累积1个月和滞后1月累积1月的耦合效应为主, 南部古尔班通古特荒漠植被对气温响应主要滞后2个月。对于气温滞后效应, 草地平均滞后(1.65±0.76)月, 森林平均滞后(1.72±0.80)月, 湿地平均滞后(1.58±0.68)月, 农田平均滞后(1.61±0.70)月;对于气温累积效应, 草地平均累积(1.57±0.69)月, 森林平均累积(1.56±0.68)月, 湿地平均累积(1.37±0.60)月, 农田平均累积(1.44±0.62)月(图 7)。同样, 与降水的时间效应相似, 森林对气温响应的滞后期最长, 对气温变化的响应最不敏感;草地对气温响应的累积期最长, 植被生长需要更长时间来累积热量。
与不考虑时间效应的模型相比(图 8), 仅考虑气候滞后效应的模型对NDVI变化的平均解释力提高了5.60%, 仅考虑气候累积效应的模型平均解释力提高了4.00%, 仅考虑气候耦合效应的模型平均解释力提高了4.22%。相比之下, 考虑最佳时间效应的模型能对NDVI变化提供更合理的解释, 平均解释力提高了7.84%。

|
| 图 8 考虑不同时间效应与不考虑时间效应的R2差值分布情况 Fig. 8 Distribution of R2 Differences Considering Different Time Effects vs. No Time Effects |
考虑最佳时间效应后, 植被生长被降水和气温主导的区域分别占整个研究区的56.17%和43.83%(图 9), 降水是影响额尔齐斯河流域植被生长的主要气候因子。在降水主导植被生长的区域中, 植被与降水呈正相关和负相关的区域分别占86.65%和13.35%, 其中显著正相关(P < 0.05)区域占32.05%, 分布于流域北部阿尔泰山脉和南部古尔班通古特荒漠, 显著负相关(P < 0.05)区域仅占0.65%。在气温主导植被生长的区域中, 植被与气温呈正相关和负相关的区域分别占51.13%和48.87%, 其中显著正相关关系(P < 0.05)区域占9.50%, 主要集中于北部阿尔泰山脉以及额尔齐斯河和乌伦古河河谷区域, 显著负相关关系(P < 0.05)区域占7.99%, 主要集中在研究区南部古尔班通古特荒漠(图 11)。

|
| 图 9 额尔齐斯河流域降水和气温主要影响的区域 Fig. 9 Areas of main influence of precipitation and temperature in the Irtysh River Basin |

|
| 图 10 考虑最佳时间效应的气候因子变化趋势 Fig. 10 Trends in climate factors considering optimal time effects |

|
| 图 11 植被与降水和气温的偏相关关系 Fig. 11 Partial correlation between vegetation and climate factors |
在降水主导植被生长的区域中, 北部阿尔泰山脉以及额尔齐斯河和乌伦古河河谷降水增加促进植被增加, 流域南部古尔班通古特荒漠降水减少导致植被减少(图 4、图 10、图 11), 草地、森林、湿地、农田与降水呈正相关的区域都占85%以上, 其中有1/3区域呈显著正相关(P < 0.05)(图 12)。在气温主导植被生长的区域中, 北部阿尔泰山脉以及额尔齐斯河和乌伦古河河谷气温升高, 植被增加(图 4、图 10、图 11), 草地、森林、湿地、农田与气温呈显著正相关(P < 0.05)的区域分别占10.41%、11.42%、29.63%、30.45%(图 12);流域南部古尔班通古特荒漠气温升高, 植被减少(图 4、图 10、图 11), 其中草地与气温呈显著负相关(P < 0.05)的区域占11.54%(图 12)。森林和草地受降水主导的区域占比更大, 农田和湿地受气温主导的区域占比更大(图 12)。

|
| 图 12 不同植被与气候因子的关系 Fig. 12 Relationship between different vegetation and climate factors |
考虑最佳时间效应后, 气候变化和人类活动对植被生长的总体贡献率分别为29.27%和70.82%(图 13)。具体而言, 流域内气候变化对NDVI的平均正向和负向贡献率分别为24.19%和34.35%, 受气候变化正向影响贡献率较高的区域主要集中在北部阿尔泰山脉, 而受气候变化负向影响贡献率较高的区域主要分布在流域的南部古尔班通古特荒漠。人类活动对NDVI的平均正向和负向贡献率分别为75.90%和65.74%, 其对整个流域NDVI变化皆有较大影响(图 14)。

|
| 图 13 额尔齐斯河流域气候变化和人类活动的平均贡献率 Fig. 13 The average contribution of climate change and human activities in the Irtysh River Basin |

|
| 图 14 额尔齐斯河流域气候变化和人类活动对NDVI的贡献率 Fig. 14 Contribution of climate change and human activities to NDVI in the Irtysh River Basin |
草地和森林主要受气候变化的正向影响和人类活动的负向影响(图 15)。在草地中, 气候变化的总体贡献率为24.26%, 其中正向贡献率为29.01%, 人类活动的总体贡献率为75.83%, 其中负向贡献率为80.56%;在森林中, 气候变化的总体贡献率为25.82%, 其中正向贡献率为43.51%, 人类活动的总体贡献率为74.34%, 其中负向贡献率为91.91%。农田和湿地主要受气候变化的负向影响和人类活动的正向影响。在湿地中, 气候变化的总体贡献率为5.44%, 其中负向贡献率为9.40%, 人类活动的总体贡献率为94.57%, 其中正向贡献率为94.72%;在农田中, 气候变化的总体贡献率为11.26%, 其中负向贡献率为17.23%, 人类活动的总体贡献率为88.78%, 其中正向贡献率为98.53%。

|
| 图 15 气候变化和人类活动对不同植被类型NDVI的平均贡献率 Fig. 15 The average contribution of climate change and human activities to NDVI for different vegetation types |
额尔齐斯河流域植被对降水变化响应的平均滞后期和平均累积期分别为1.92个月和1.90个月, 与前人研究结果类似, 王会静[24]研究表明新疆有72%植被对降水的响应平均滞后时间达1个多月, Wu[7]研究表明亚洲、美洲中高纬干旱半干旱区植被生长对降水响应滞后和累积1—2个月。干旱半干旱区植被对降水响应的时间效应主要受植被的生理适应机制、土壤等因素的影响。一方面, 植被生长并非直接对降水做出反应, 而是受实际土壤水分的直接影响, 降水发生和植物根系吸水之间存在的时间差, 干旱半干旱区深根植物有效利用水分的适应性策略进一步缓冲了降水波动对NDVI的影响[25];另一方面, 干旱半干旱区降水量总体较少, 粘土含量低导致土壤孔隙大, 水分流失快, 植被生长需要时间累积生长所需水分。森林对降水的滞后期最长, 主要由于森林从土壤深层吸取水分, 且叶片面积大, 水分的传导率低, 深层土壤中的水分上升到地表再被植被吸收的过程需要更多时间[26], 同时林区土壤更厚, 含水量大, 从而能在更长的时间上抵御干旱[27], 森林对降水变化的响应最不敏感。草地土壤较薄, 含水量少, 水分流失快, 对降水变化响应的累积期最长。
额尔齐斯河流域植被对气温响应的平均滞后期和平均累积期分别为1.70个月和1.57个月, 与前人研究结果类似, 黄豪奔[15]研究表明阿勒泰地区植被对气温的响应滞后期主要在2个月内, Wu[7]研究表明北半球中高纬温度较低地区植被对气温响应的累积期可达1个月。植被对气温变化响应的时间效应主要受植被的生理适应机制、土壤、海拔、冰川融化等因素的影响。一方面, 北温带的季节温差大, 植被对温度波动的适应能力较强;另一方面, 植被生长受土壤温度的直接影响, 先前气温升高使得土壤温度升高, 加快植被内部化学反应速率进而促进生长[28];再者, 气温升高使得阿尔泰山脉高海拔地区的冰川融化进而促进植被生长, 这个过程需要足够时间。森林对气温波动的适应力强, 植被根系从深层土壤吸取热量和冰雪融水需要的时间更长, 对气温变化响应的滞后期最长。草地主要分布在北部阿尔泰山脉高海拔区域, 气温低, 土壤保温作用弱, 需要更长时间累积生长所需的热量条件, 对气温变化响应的累积期最长。
另外, 额尔齐斯河流域植被对降水响应的滞后期和累积期都比气温更长, 这是由于在满足植被生长的温度要求之前, 气温对植被生长起主要的限制作用, 当达到植被临界生长温度后, 水分才对植被生长起主导作用[29]。另外, 植被生长具有相应的温度阈值, 在累积温度上升到合适的温度阈值之前, 植被生长良好, 当温度超过阈值时, 气孔关闭和酶活性降低不利于植被光合作用, 温度的持续升高也会加速土壤水分的蒸发, 导致干旱, 进而削弱植被的生长[30—31]。
3.2 影响因素的区域贡献率分析北部阿尔泰山脉植被生长受气候变化的正向影响明显, NDVI呈增加趋势, 与降水和气温变化都呈正相关, 这由于山区迎风坡降水增加有利于植被生长, 以及高海拔缓解了气温升高带来的水分蒸发, 相反气温的升高会促进冰雪融水的增加, 进而促进植被生长。流域南部古尔班通古特荒漠植被生长受气候变化的负向影响明显, NDVI呈减小趋势, 与气温变化呈负相关, 与降水变化呈正相关, 主要由于干旱半干旱区降水量减少, 气温升高促使蒸发增加, 降低土壤含水量, 从而抑制植被的生长[32]。
额尔齐斯河流域植被受人类活动的干扰影响明显, 主要集中在额尔齐斯河和乌伦古河河谷及周边区域, 当地放牧、打草活动频繁, 长期存在强烈的林牧矛盾对部分植被生长产生抑制作用[33]。但2000年以来流域实施了许多重大生态保护工程[34], 以及新疆推广的种植柽柳、铺设草网、防沙治沙等工程, 对部分植被恢复产生了明显的促进作用。人为耕作活动也有利于荒漠区植被覆盖度的增加, 从区域植被指数反应的结果来看, 滴灌喷灌等农业新技术改良了沙化地, 间接增大了植被盖度[35]。
4 结论与展望基于考虑植被对气候响应的最佳时间效应模型基础上, 系统分析了额尔齐斯河流域NDVI的时空变化规律以及驱动因素。近二十年额尔齐斯河流域NDVI总体上呈增加趋势, 年际增长率达0.0017/a, 增加区域主要集中分布于阿尔泰山脉和河谷区域。仅考虑气候影响的条件下, 由于土壤、海拔、冰川融水以及不同植被类型的生理适应机制差异, 降水影响植被生长的滞后期和累积期都长于气温。降水和气温对植被的影响分别平均滞后1.92、1.70个月, 平均累积1.90、1.57个月。考虑最佳时间效应后, 气候因素对植被变化的解释力提高了7.84%, 对植被变化的总体贡献率为29.27%。气候驱动因素存在空间差异, 流域北部阿尔泰山脉降水增加、气温升高带来的冰川融化促进植被生长, 流域南部荒漠区域降水减少、气温升高导致蒸发增加进而抑制植被生长。本研究更多地考虑了植被生长对气候变化响应的时间效应, 有利于提高植被动态变化归因分析的解释力, 但主要分析了气温和降水因子, 存在高估人类活动对NDVI影响力的潜在局限, 今后应考虑更多其他影响植被生长的气候变量。
| [1] |
Yang S K, Liu J, Wang C H, Zhang T, Dong X H, Liu Y L. Vegetation dynamics influenced by climate change and human activities in the Hanjiang River Basin, central China. Ecological Indicators, 2022, 145: 109586. DOI:10.1016/j.ecolind.2022.109586 |
| [2] |
Zhang Y H, Ye A Z. Quantitatively distinguishing the impact of climate change and human activities on vegetation in mainland China with the improved residual method. GIScience & Remote Sensing, 2021, 58(2): 235-260. |
| [3] |
解晗, 同小娟, 李俊, 张静茹, 刘沛荣, 于裴洋. 2000—2018年黄河流域生长季植被指数变化及其对气候因子的响应. 生态学报, 2022, 42(11): 4536-4549. DOI:10.5846/stxb202104271108 |
| [4] |
Zhang Z Z, Wang Z P, Lai H X, Wang F, Li Y B, Feng K, Qi Q Q, Di D Y. Lag Time and Cumulative Effects of Climate Factors on Drought in North China Plain. Water, 2023, 15(19): 3428. DOI:10.3390/w15193428 |
| [5] |
Liu Z J, Liu Y S, Li Y R. Anthropogenic contributions dominate trends of vegetation cover change over the farming-pastoral ecotone of northern China. Ecological Indicators, 2018, 95P1(Dec.): 370-378. |
| [6] |
Du J Q, Quan Z J, Fang S F, Liu C C, Wu J H, Fu Q. Spatiotemporal changes in vegetation coverage and its causes in China since the Chinese economic reform. Environmental Science and Pollution Research, 2020, 27(1): 1144-1159. DOI:10.1007/s11356-019-06609-6 |
| [7] |
Wu D H, Zhao X, Liang S L, Zhou T, Huang K C, Tang B J, Zhao W Q. Time-lag effects of global vegetation responses to climate change. Global Change Biology, 2015, 21(9): 3520-3531. DOI:10.1111/gcb.12945 |
| [8] |
Zhang Z Y, Ju W M, Zhou Y L, Li X Y. Revisiting the cumulative effects of drought on global gross primary productivity based on new long-term series data (1982—2018). Global Change Biology, 2022, 28(11): 3620-3635. DOI:10.1111/gcb.16178 |
| [9] |
Liu N, Ding Y X, Peng S Z. Temporal effects of climate on vegetation trigger the response biases of vegetation to human activities. Global Ecology and Conservation, 2021, 31: e01822. DOI:10.1016/j.gecco.2021.e01822 |
| [10] |
Jin K, Jin Y S, Wang F, Zong Q L. Should time-lag and time-accumulation effects of climate be considered in attribution of vegetation dynamics? Case study of China's temperate grassland region. International Journal of Biometeorology, 2023, 67(7): 1213-1223. DOI:10.1007/s00484-023-02489-1 |
| [11] |
Liu N, Shi Y F, Ding Y X, Liu L, Peng S Z. Temporal effects of climatic factors on vegetation phenology on the Loess Plateau, China. Journal of Plant Ecology, 2023, 16(2): rtac063. DOI:10.1093/jpe/rtac063 |
| [12] |
Ma M Y, Wang Q M, Liu R, Zhao Y, Zhang D Q. Effects of climate change and human activities on vegetation coverage change in northern China considering extreme climate and time-lag and -accumulation effects. Science of the Total Environment, 2023, 860: 160527. DOI:10.1016/j.scitotenv.2022.160527 |
| [13] |
Zhu Y Y, Zhang H M, Ding M J, Li L H, Zhang Y L. The Multiple Perspective Response of Vegetation to Drought on the Qinghai-Tibetan Plateau. Remote Sensing, 2023, 15: 902. DOI:10.3390/rs15040902 |
| [14] |
陆建涛, 彭建, 李刚勇, 关靖云, 韩万强, 刘亮, 王仁军, 郑江华. 干旱对中亚草地总初级生产力时滞和累积效应的影响评估. 生态学报, 2023, 43(23): 9745-9757. DOI:10.20103/j.stxb.202209162644 |
| [15] |
黄豪奔, 徐海量, 林涛, 夏国柱. 2001—2020年新疆阿勒泰地区归一化植被指数时空变化特征及其对气候变化的响应. 生态学报, 2022, 42(7): 2798-2809. DOI:10.5846/stxb202107021768 |
| [16] |
陈春波, 李刚勇, 彭建. 1981—2018年新疆草地归一化植被指数时空特征及其对气候变化的响应. 生态学报, 2023, 43(4): 1537-1552. DOI:10.5846/stxb202201260251 |
| [17] |
Han F F, Yan J J, Ling H B. Variance of vegetation coverage and its sensitivity to climatic factors in the Irtysh River basin. PeerJ, 2021, 9: e11334. |
| [18] |
杨靖, 戴君虎, 姚华荣, 陶泽兴, 朱梦瑶. 1992—2020年横断山区植被分布与植被活动变化. 地理学报, 2022, 77(11): 2787-2802. |
| [19] |
雷茜, 胡忠文, 王敬哲, 张英慧, 邬国锋. 1985—2015年中国不同生态系统NDVI时空变化及其对气候因子的响应. 生态学报, 2023, 43(15): 6378-6391. DOI:10.5846/stxb202208032216 |
| [20] |
金凯, 王飞, 韩剑桥, 史尚渝, 丁文斌. 1982—2015年中国气候变化和人类活动对植被NDVI变化的影响. 地理学报, 2020, 75(5): 961-974. |
| [21] |
耿庆玲, 陈晓青, 赫晓慧, 田智慧. 中国不同植被类型归一化植被指数对气候变化和人类活动的响应. 生态学报, 2022, 42(9): 3557-3568. DOI:10.5846/stxb202012313335 |
| [22] |
Peng S Z, Ding Y X, Liu W Z, Li Z. 1 km monthly temperature and precipitation dataset for China from 1901 to 2017. Earth System Science Data, 2019, 11(4): 1931-1946. |
| [23] |
Yang J, Huang X. The 30 m annual land cover dataset and its dynamics in China from 1990 to 2019. Earth System Science Data, 2021, 13(8): 3907-3925. |
| [24] |
王会静, 郭玉川, 白运保, 王宁, 魏宣, 周明通. 新疆植被动态格局及其对气候的时滞效应. 农业工程学报, 2023, 39(11): 137-145. |
| [25] |
Chen T, de Jeu R A M, Liu Y Y, van der Werf G R, Dolman A J. Using satellite based soil moisture to quantify the water driven variability in NDVI: A case study over mainland Australia. Remote Sensing of Environment, 2014, 140: 330-338. |
| [26] |
Ershadi A, McCabe M, Evans J, Wood E. Impact of model structure and parameterization on Penman-Monteith type evaporation models. Journal of Hydrology, 2015, 525: 521-535. |
| [27] |
Yang C P, Wang Y Q, Wu J J, Shen H Z, Ma X Y. Spatiotemporal evolution and lag effect of drought and vegetation dynamics in southwest China. Applied ecology and environmental research, 2022, 20(4): 3447-3466. |
| [28] |
Zhang H X, Yuan N M, Ma Z G, Huang Y. Understanding the soil temperature variability at different depths: effects of surface air temperature, snow cover, and the soil memory. Advances in Atmospheric Sciences, 2021, 38(3): 493-503. |
| [29] |
Zhou X C, Geng X J, Yin G D, Hänninen H, Hao F H, Zhang X, Fu Y H. Legacy effect of spring phenology on vegetation growth in temperate China. Agricultural and Forest Meteorology, 2020, 281: 107845. |
| [30] |
Tang W X, Liu S G, Kang P, Peng X, Li Y Y, Guo R, Jia J N, Liu M C, Zhu L J. Quantifying the lagged effects of climate factors on vegetation growth in 32 major cities of China. Ecological Indicators, 2021, 132: 108290. |
| [31] |
Jiang H L, Xu X. Impact of extreme climates on vegetation from multiple scales and perspectives in the Agro-pastural Transitional Zone of Northern China in the past three decades. Journal of Cleaner Production, 2022, 372: 133459. |
| [32] |
Jin K, Wang F, Zong Q L, Qin P, Liu C X, Wang S X. Spatiotemporal differences in climate change impacts on vegetation cover in China from 1982 to 2015. Environmental Science and Pollution Research, 2022, 29(7): 10263-10276. |
| [33] |
刘时栋, 刘琳, 张建军, 王柯, 郭义强. 基于生态系统服务能力提升的干旱区生态保护与修复研究——以额尔齐斯河流域生态保护与修复试点工程区为例. 生态学报, 2019, 39(23): 8998-9007. DOI:10.5846/stxb201905281104 |
| [34] |
周立华, 刘洋. 中国生态建设回顾与展望. 生态学报, 2021, 41(8): 3306-3314. DOI:10.5846/stxb202001200154 |
| [35] |
Guan J Y, Yao J Q, Li M Y, Zheng J H. Assessing the spatiotemporal evolution of anthropogenic impacts on remotely sensed vegetation dynamics in Xinjiang, China. Remote Sensing, 2021, 13(22): 4651. |