 2024, Vol. 44
2024, Vol. 44文章信息
- 李旭, 田永莲, 贺禹文, 孙秀锋
- LI Xu, TIAN Yonglian, HE Yuwen, SUN Xiufeng
- 我国城市鸟类多样性特征及鸟类保护导向的城市绿地营建
- Characteristics of urban avian diversity in China and the construction of urban green space guided by bird protection
- 生态学报. 2024, 44(22): 10234-10246
- Acta Ecologica Sinica. 2024, 44(22): 10234-10246
- http://dx.doi.org/10.20103/j.stxb.202312192756
-
文章历史
- 收稿日期: 2023-12-19
- 网络出版日期: 2024-08-22

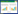




城市生物多样性关乎人类的生存、福祉和可持续发展。当前城市生物多样性的降低已是世界范围广泛关注的问题[1—2]。鸟类作为生态金字塔顶端的上层物种, 直接反映下层生态群的状态[3], 同时作为对环境最为敏感的野生动物类群, 是衡量城市生态质量最直观和最有效的因子之一[4—5], 在城市人居环境高质量发展需求背景下, 认识城市鸟类多样性特征并进行合理保护有重要意义。由于鸟类对栖息地环境变化的敏感性, 高速城市化过程的土地利用、异质性、多边界特征、景观破碎化, 以及热岛效应、人为干扰等诸多因素均对城市鸟类多样性造成巨大冲击[6—8], 深刻改变着城市的生物多样性分布格局。从总体上看, 由于全球城市都是由居住区、商业区及道路构成的整体生境, 具有相似和规则的生境结构[9], 导致城市环境具有筛子效应, 过滤掉不适应种而保留适应种[10], 同时也会有新的鸟类迁入, 使得城市成为引进外来物种和灭绝本地物种的焦点[11], 由此学者将城市鸟类分为了城市适应种、回避种及开拓种三个类型[12]。以上机制使不同城市间鸟类群落逐渐趋向同质化及单一性, 并导致城市鸟类物种组成及β多样性的空间变化[13]。
城市绿地作为鸟类重要的繁殖及觅食空间, 对其多样性有重要影响[14]。首先鸟类多样性和绿地面积成正相关, 鸟类多样性格局符合经典生态学的种-面积关系理论, 且面积较大的绿地斑块的破碎化对多样性降低的影响更大[15—17]。对不同类型鸟类的影响则较为复杂, 如树木营巢、边缘生境鸟类及杂食性留鸟更易在城市繁殖与生存[1, 9, 13], 而虫食性鸟类、筑地面巢和树洞巢鸟类等, 则在城市中数量减少。外来鸟类一般属于城市开拓种, 所以本地鸟类受栖息地破坏及隔离效应的压力更大[1], 甚至以城市为停歇点的迁徙鸟类也会受到城市绿地变化的显著影响[18]。
目前城市鸟类学研究的热点主要集中在三个方面:城市鸟类的基础研究、影响城市鸟类的环境因素研究及城市鸟类的保护研究[6]。基础研究主要关注于城市鸟类的生态习性、分布特点及群落构成等[19], 影响因素主要从城市、绿地和生境斑块三个尺度来探讨[20—23], 保护研究则主要聚焦于栖息地保护规划及鸟类招引技术等方面[24—25]。但目前总体来看研究成果对实践指导作用受限, 城市生物多样性规划和建设实践中对鸟类多样性关注不足[4, 26], 现实中常见违背生态规律的绿地建造, 导致绿林地成为有绿无虫、有绿无鸟的“绿色沙漠”[27], 其原因之一即是研究多是基于城市内部中小尺度的实证研究, 如以群落的多样性、丰富度以及均匀性作为群落物种丰富性表征来反映城市化对鸟类的影响[28], 缺少系统性城市鸟类保护框架, 难以应对复杂多样的城市环境, 而景观尺度上哪些因素对城市鸟类分布有重要影响这一问题仍未达成共识[22], 这也限制了城市鸟类保护和恢复的进一步发展。而实际上城市鸟类多样性受城市区位、研究尺度及栖息地结构等因子的共同影响, 难以将一个尺度或一个城市的研究结果推演到另外的尺度或城市, 因此有必要从多重组织水平和多空间尺度开展鸟类多样性研究[9], 以形成对城市鸟类多样性特征规律的总体认识。
基于此, 本研究拟在跨城市尺度上分析城市鸟类多样性的基本特征, 包括与全国鸟类多样性特征进行对比, 其次对城市鸟类多样性特征进行聚类, 进而对影响城市鸟类多样性的因素进行综合分析, 试图回答以下问题:(1)我国城市鸟类多样性的总体特征及其在地域空间上的分布规律;(2)驱动城市鸟类多样性差异的主导因素;依据分析结果提出鸟类多样性保护导向的城市绿地营建策略, 旨在为城市鸟类多样性保护及生态城市建设提供参考。
1 数据来源与分析方法 1.1 数据来源样本城市的确定是基于鸟类名录的可得性并兼顾鸟类区系的覆盖性, 在中英文文献中检索包含有“鸟类名录”的城市, 名录鸟类均指城市建成区内的鸟类, 剔除单一季节鸟类的城市, 确定了21个城市为样本, 并通过“鸟人课堂”(http://www.birder.cn)和“中国观鸟记录中心”(http://www.birdreport.cn)对数据做了补充, 时间截止于2022年12月, 依据《中国鸟类分类与分布名录》(第4版)[29]对鸟类名称进行了统计并确定其科、属。确定的21个城市分布于我国7个鸟类地理区(图 1), 具有一定的代表性, 虽然不同鸟类地理分区样本城市数量不一, 但距离较近的城市往往具有相似的鸟类多样性构成, 可以交互验证, 并不会影响分析结果。

|
| 图 1 鸟类地理分区及样本城市位置图 Fig. 1 Geographical zoning of birds and location of sample cities |
鸟类多样性影响因子在综合参考相关文献基础上, 从城市空间距离、自然环境及社会经济3个维度选取9个因子, 其中年均温度及年均降雨量来自世界气象网站, 城市绿化覆盖度及人口、面积等来自中国城市统计年鉴[30], 绿视率及植物丰富度等因子通过相关文献获得[31]。
1.2 分析方法 1.2.1 城市鸟类多样性特征及地域型划分为全面准确掌握各城市鸟类基本信息, 通过数据透视表(Pivot Table)统计各城市每种鸟类的科属种、居留类型及食性类型信息, 并形成各城市上述三类特征的占比或结构分析图, 进而应用SPSS从三个维度对城市鸟类特征进行聚类, 聚类方法采用组间联接法, 聚类数范围暂不设置, 并绘制聚类树状图;根据聚类结果统计各类型鸟类多样性特征, 并以桑基图的形式分析三种聚类结果的关联性, 进而根据关联性城市的地域分布并结合鸟类地理区划分出城市鸟类多样性类型。基于此划分出的鸟类多样性类型具有相似的种属(科)比、居留类型及食性类型结构, 在空间上具有聚集性且比鸟类地理区单元更具体, 本研究中将其称之为城市鸟类多样性的“地域型”。
1.2.2 城市鸟类多样影响因素分析为解析影响城市鸟类多样性差异的因素, 应用Mantel检验分析方法进行, 其原理是对一组观察对象来说, 在某个特征属性上差距越大, 则在与之相关的另一方面的特征属性差距也越大, 该方法通过对两个矩阵计算其相关性, 避免了数据类型的限制, 本研究中城市距离矩阵通过样本城市地理坐标计算欧式距离获得。为揭示不同城市鸟类多样性组成差异的影响因素, 应用POD法将鸟类多样性进行分解[32], 则城市鸟类组成差异由物种替换和丰富度差异这两方面反映, 如两个样本城市共有鸟类种数为A, 而各自特有的鸟类种数为B和C, 那么两个城市间鸟类组成的差异为B+C, 丰富度差异为∣B-C∣, 而物种替换为2min(B, C)[33]。
2 结果与分析 2.1 城市鸟类多样性基本特征我国是跨两个动物地理界的仅有两个国家之一[34], 共有鸟类1507种[29], 约占世界鸟类的1/6, 全球8条鸟类重要迁徙线路中有3条和我国鸟类迁徙密切相关[35], 鸟类多样性十分丰富。
分析发现21个样本城市共记录鸟类763种, 占我国鸟类总种数的50.63%, 隶属于27目, 92科, 289属(表 1)。从城市鸟类科、属、种的构成看, 属科比及种科比分别为3.14及8.29, 但属内种的数量频次与科内种的数量频次分布极不均衡(图 2)。城市属内种数量频次分布虽然与全国鸟类频次分布相似, 但相同数量的频次低于全国值;而科内种数量全国来看频次最高的是有2—3个种的科[36], 而城市中频次最高的是只有1个种的科, 且显著高于其它频次, 这说明城市鸟类的科内种更单一, 从而反映了城市鸟类多样性具脆弱性。从食性结构看, 城市鸟类食性以杂食及虫食性为主, 两者占总构成的77%, 其次是鱼食性及肉食性鸟类, 而最少的植食性鸟类仅占总比例的2%;从类型差异看, 杂食性占比在各城市间差异最小, 占比最大的乌鲁木齐为47%, 最小的宜州为32%, 其次虫食性占比在各城市间差2倍左右, 而肉食性占比差异达10倍, 北京为20%, 最低的呼和浩特仅为2%。从居留类型构成来看, 主要受我国所处气候带影响, 北方城市多夏候鸟, 南方城市多留鸟, 但整体看占比最多的是留鸟, 为36%, 夏候鸟、冬候鸟及旅鸟依次为25%、20%及19%(表 1)。
| 城市 City |
居留类型Residence type | 食性类型Feeding habits type | 总计 Total |
||||||||||||||||||||||||||||||||||||
| 旅鸟 Migrant bird |
留鸟 Resident |
夏候鸟 Summer bird |
冬候鸟 Winter bird |
杂食性 Omnivorous |
植食性 Phytophagous |
鱼食性 Fish eating birds |
肉食性 Carnivority |
虫食性 Insectivorous |
|||||||||||||||||||||||||||||||
| 科 | 属 | 种 | 科 | 属 | 种 | 科 | 属 | 种 | 科 | 属 | 种 | 科 | 属 | 种 | 科 | 属 | 种 | 科 | 属 | 种 | 科 | 属 | 种 | 科 | 属 | 种 | 科 | 属 | 种 | ||||||||||
| 北京 | 29 | 65 | 127 | 28 | 51 | 67 | 37 | 69 | 98 | 22 | 30 | 42 | 31 | 64 | 116 | 4 | 6 | 9 | 13 | 32 | 48 | 10 | 33 | 66 | 25 | 44 | 95 | 65 | 166 | 334 | |||||||||
| 成都 | 31 | 58 | 90 | 37 | 63 | 86 | 24 | 41 | 60 | 30 | 56 | 86 | 30 | 64 | 119 | 3 | 4 | 4 | 12 | 27 | 38 | 7 | 16 | 27 | 30 | 66 | 134 | 63 | 165 | 322 | |||||||||
| 福州 | 16 | 29 | 49 | 42 | 78 | 100 | 23 | 40 | 48 | 28 | 48 | 84 | 33 | 58 | 96 | 4 | 6 | 8 | 13 | 27 | 45 | 5 | 13 | 21 | 27 | 64 | 111 | 66 | 160 | 281 | |||||||||
| 广州 | 13 | 20 | 28 | 47 | 100 | 135 | 19 | 29 | 39 | 32 | 56 | 93 | 33 | 63 | 105 | 4 | 6 | 9 | 9 | 23 | 32 | 5 | 15 | 20 | 31 | 73 | 129 | 67 | 173 | 295 | |||||||||
| 哈尔滨 | 9 | 11 | 15 | 16 | 23 | 29 | 33 | 64 | 99 | 4 | 7 | 7 | 17 | 36 | 62 | 2 | 2 | 2 | 8 | 16 | 21 | 4 | 7 | 12 | 20 | 33 | 53 | 42 | 92 | 150 | |||||||||
| 杭州 | 17 | 24 | 41 | 41 | 69 | 90 | 21 | 32 | 43 | 30 | 47 | 71 | 31 | 54 | 96 | 4 | 5 | 6 | 11 | 23 | 29 | 4 | 13 | 17 | 27 | 51 | 97 | 63 | 140 | 245 | |||||||||
| 呼和浩特 | 7 | 11 | 12 | 9 | 12 | 15 | 22 | 26 | 28 | 4 | 4 | 4 | 15 | 19 | 23 | 1 | 1 | 1 | 4 | 10 | 11 | 1 | 1 | 1 | 15 | 20 | 23 | 34 | 51 | 59 | |||||||||
| 昆明 | 20 | 33 | 48 | 48 | 91 | 137 | 15 | 26 | 40 | 16 | 20 | 32 | 29 | 52 | 96 | 2 | 2 | 2 | 7 | 16 | 21 | 5 | 11 | 16 | 29 | 60 | 122 | 57 | 134 | 257 | |||||||||
| 拉萨 | 9 | 12 | 14 | 23 | 44 | 59 | 18 | 30 | 34 | 7 | 8 | 11 | 18 | 33 | 53 | 4 | 5 | 7 | 5 | 11 | 14 | 3 | 10 | 13 | 14 | 22 | 31 | 33 | 77 | 118 | |||||||||
| 兰州 | 18 | 32 | 54 | 26 | 51 | 81 | 29 | 44 | 69 | 8 | 12 | 14 | 22 | 50 | 87 | 2 | 4 | 4 | 10 | 18 | 26 | 3 | 10 | 18 | 20 | 40 | 83 | 48 | 118 | 218 | |||||||||
| 临沂 | 15 | 21 | 24 | 21 | 26 | 29 | 26 | 36 | 43 | 6 | 7 | 10 | 21 | 33 | 48 | 2 | 2 | 2 | 7 | 15 | 17 | 4 | 6 | 6 | 17 | 22 | 33 | 43 | 75 | 106 | |||||||||
| 柳州 | 4 | 4 | 4 | 44 | 67 | 88 | 11 | 12 | 13 | 15 | 23 | 40 | 23 | 34 | 57 | 3 | 4 | 4 | 4 | 7 | 7 | 6 | 9 | 11 | 25 | 42 | 66 | 51 | 93 | 145 | |||||||||
| 牡丹江 | 8 | 8 | 10 | 14 | 22 | 26 | 29 | 38 | 51 | 3 | 3 | 3 | 15 | 24 | 36 | 2 | 2 | 2 | 4 | 5 | 6 | 2 | 4 | 6 | 20 | 27 | 40 | 37 | 62 | 90 | |||||||||
| 南宁 | 6 | 7 | 8 | 31 | 62 | 84 | 13 | 17 | 19 | 19 | 29 | 41 | 18 | 32 | 48 | 2 | 3 | 4 | 7 | 15 | 17 | 5 | 10 | 11 | 23 | 45 | 72 | 46 | 102 | 152 | |||||||||
| 上海 | 25 | 48 | 93 | 37 | 59 | 83 | 31 | 44 | 63 | 29 | 55 | 102 | 30 | 56 | 109 | 4 | 8 | 10 | 15 | 33 | 65 | 5 | 11 | 22 | 28 | 60 | 135 | 64 | 159 | 341 | |||||||||
| 潍坊 | 16 | 26 | 38 | 22 | 28 | 33 | 27 | 40 | 49 | 9 | 13 | 14 | 21 | 34 | 53 | 2 | 3 | 3 | 7 | 18 | 21 | 3 | 5 | 6 | 20 | 31 | 51 | 46 | 89 | 134 | |||||||||
| 乌鲁木齐 | 3 | 7 | 9 | 14 | 26 | 40 | 28 | 42 | 63 | 5 | 6 | 8 | 17 | 35 | 56 | 1 | 2 | 3 | 7 | 10 | 14 | 4 | 8 | 9 | 16 | 23 | 38 | 37 | 75 | 120 | |||||||||
| 武汉 | 24 | 33 | 56 | 35 | 65 | 86 | 27 | 49 | 64 | 26 | 52 | 101 | 30 | 57 | 103 | 3 | 5 | 8 | 13 | 33 | 43 | 5 | 15 | 27 | 30 | 64 | 126 | 62 | 162 | 307 | |||||||||
| 西安 | 22 | 36 | 47 | 29 | 55 | 88 | 29 | 46 | 63 | 14 | 22 | 33 | 25 | 49 | 90 | 2 | 4 | 5 | 11 | 26 | 29 | 3 | 10 | 16 | 27 | 45 | 91 | 55 | 129 | 231 | |||||||||
| 宜州 | 4 | 7 | 8 | 36 | 59 | 71 | 6 | 6 | 8 | 15 | 19 | 24 | 19 | 26 | 33 | 2 | 3 | 4 | 4 | 8 | 8 | 5 | 8 | 9 | 23 | 40 | 57 | 45 | 81 | 111 | |||||||||
| 长沙 | 6 | 7 | 9 | 40 | 71 | 101 | 28 | 47 | 62 | 17 | 29 | 46 | 30 | 55 | 86 | 3 | 3 | 3 | 9 | 20 | 27 | 5 | 8 | 14 | 27 | 50 | 88 | 60 | 128 | 218 | |||||||||

|
| 图 2 城市与全国鸟类科(属) 内种数量频次分布对比 Fig. 2 Comparison of the frequency distribution of bird species in urban and national bird families (genera) |
通过对种属(科)比、食性结构及居留结构进行聚类, 发现当距离系数d=10时, 三种聚类均可将样本城市鸟类多样性分为4个明显的类型(图 3)。从种(属)在高级分类阶元的多样性特征看, 在属/科比、种/属比、种/科比三个方面, 临沂等10个城市为“低低低”型, 杭州等4个城市为均衡型, 北京等4个城市为“高高高”型, 而成都等3个城市为“高低高”型(图 4)。“低低低”型意味着更加单一的种在为高级分类阶元的多样性发挥贡献, 也即单一鸟类种在城市空间迁出或灭绝, 则意味着一个鸟类属或科的迁出或灭绝, 而这一类型占样本城市的近50%, “高高高”型城市仅占样本城市的20%, 这也反映了城市鸟类多样性的脆弱性。从食性结构类型看(图 4), 有杭州等12个城市属于均衡型, 食性结构接近于平均值, 昆明等5个城市属于虫高型, 虫食性鸟类占47%, 明显高于其它类型, 临沂等3个城市属于杂食型, 杂食性鸟类占46%, 而北京独自成为肉高型城市, 肉食性鸟类占比达20%, 比其他城市的平均值7%高出近三倍。从居留结构类型看(图 4), 四种居留类型城市的空间分布具有强地域规律, 符合我国的鸟类地理区分布特征, 杭州等5个城市为均衡型, 结构较为接近平均值, 该类型主要分布于鸟类地理区中的华中区, 西安等6个城市冬候鸟明显偏少, 仅占11%, 属于冬少型, 主要分布于华北区, 哈尔滨等3个城市夏候鸟明显高于其它类型, 占比达60%, 属于夏多型, 而广州等7个城市留鸟占比达52%, 属于典型的留鸟型居留结构。

|
| 图 3 种属(科) 比、食性结构、居留结构聚类图 Fig. 3 Cluster diagram of species (family) ratio, feeding structure, and residence structure |

|
| 图 4 聚类类型构成对比 Fig. 4 Comparison chart of cluster results |
为进一步解析这三种类型结构之间的城市对应关系, 桑基图分析发现其有明显的规律性(图 5)。其中食性结构类型与种(属)科比类型有非常强的对应规律, 比如虫食型及杂食型城市基本属于低低低型, 而这两种食性结构类型也基本对应居留类型中的夏多型和留鸟型, 在地理空间上处于我国的南北方两端, 也即反映出广大中部地区城市的鸟类食性结构更加均衡, 且多数城市的种(属)科比也较高。这其中北京、临沂及牡丹江有其特殊性, 北京单一城市属于肉高型, 但对应的种属比却属于高高高型;临沂属于冬少型, 但却与拉萨及乌鲁木齐相似属于杂食型, 且种科比偏低, 拉萨及乌鲁木齐由于其高原及内陆干旱的区域环境导致城市鸟类以城市适应种为主, 因此杂食性鸟类占比高, 而同属于冬少型的其他城市多数食性结构较均衡;牡丹江市是唯一非留鸟型中的虫高型。由此该三个城市的鸟类食性结构是否由生态金字塔中的中下层生物多样性差异导致, 值得进一步探究。

|
| 图 5 聚类城市对应关系桑基图 Fig. 5 Sankey diagram of city correspondence for three clusters |
依据桑基图, 可将城市鸟类多样性分为以下10类地域型(表 2):低-杂(均)-夏多型, 该类型主要分布于东北及蒙新鸟类亚区, 包括乌鲁木齐、哈尔滨及牡丹江, 主要特征是科(属)内种单一, 以杂食性为主, 夏季鸟类种数突出, 夏候鸟占总鸟类种数的60%;低-均(杂)-冬少型, 主要分布于华北鸟类亚区的东半部, 包括呼和浩特、潍坊及临沂, 主要特征是科(属)内种单一, 鸟类食性均衡, 旅鸟、留鸟及夏候鸟种数相当, 冬候鸟种数较少, 仅占鸟类总种数的11%;均-均-冬少型, 主要分布于华北鸟类亚区的偏西部, 包括西安及兰州, 食性构成及居留构成特征与低-均(杂)-冬少型类似, 但却较其科(属)内种丰富;高-肉-旅多型, 代表城市为北京, 分布于华北地区二三阶梯过度地带, 特点是科(属)内种丰富, 肉食性种类突出, 旅鸟占比高;低-杂-留鸟型, 主要指拉萨等青藏鸟类亚区, 特征是科(属)内种单一, 以杂食性留鸟种类突出, 留鸟占鸟类种数的52%;高-虫-留鸟型, 该类型主要分布于以昆明为代表的西南鸟类亚区, 主要特征是科(属)内种丰富, 以虫食性鸟类占47%, 留鸟占总种数的52%;均-均-均衡型, 该类型主要分布长江中下游区域, 属于华中鸟类亚区, 包括成都、武汉、上海、杭州及福州, 主要特征是科(属)内种数量、食性及居留类型构成较均衡;长沙也属于华中鸟类亚区, 但留鸟种类却偏多, 与华南鸟类亚区类似, 为低-虫-留鸟型, 主要分布于华南鸟类亚区, 包括南宁、柳州、宜州等城市, 主要特征是科(属)内种单一, 虫食性鸟类较多, 占47%, 居留类型以留鸟为主, 占52%;此外广州虽然也处于该亚区且以留鸟为主, 食性构成却较均衡, 且科(属)内种丰富, 这一现象值得研究, 可称之为高-均-留鸟型。
| 地域型 Regional type |
分布区域 Distribution area |
对应鸟类地理区 Corresponding bird geographic areas |
多样性特征 Diversity characteristics |
| 低-杂(均)-夏多型 Type of L-O(Bd)-S |
东北及西北地区 | 东北、蒙新鸟类地理区 | 科(属)内种单一, 杂食性为主, 夏季鸟种数突出 |
| 低-均(杂)-冬少型 Type of L-Bd(O)-Wf |
华北地区中东部 | 华北鸟类地理区东半部 | 科(属)内种单一, 食性均衡, 冬候鸟种数较少 |
| 均-均-冬少型Type of M-Bd-Wf | 华北地区西部 | 华北鸟类地理区西半部 | 科(属)内种较丰富, 食性均衡, 冬候鸟种数少 |
| 高-肉-旅多型Type of H-C-T | 华北地区中北部 | 华北鸟类地理区中北部 | 科(属)内种丰富, 肉食性种类突出, 旅鸟占比高 |
| 低-杂-留鸟型Type of L-0-R | 青藏高原区 | 青藏鸟类地理区 | 科(属)内种单一, 以杂食性及留鸟种类突出 |
| 高-虫-留鸟型Type of L-I-R | 云贵等西南地区 | 西南鸟类地理区 | 科(属)内种丰富, 虫食性及留鸟类型占比高 |
| 均-均-均衡型Type of M-Bd-Br | 长江中下游区域 | 华中鸟类地理区 | 科(属)内种数量、食性类型及居留类型较均衡 |
| 均-均-留鸟型Type of M-Bd-R | 华中地区南部 | 华中鸟类地理区南部 | 科(属)内种数量、食性类型均衡, 留鸟偏多 |
| 低-虫-留鸟型Type of L-I-R | 岭南地区 | 华南鸟地理区 | 科(属)内种单一, 虫食鸟类多, 留鸟占比突出 |
| 高-均-留鸟型Type of H-Bd-R | 岭南地区中南部 | 华南鸟地理区 | 科(属)内种丰富, 食性结构较均衡, 留鸟占比突出 |
| L:低Low;M:均衡居中Medium;H:高High;O:杂食性Omnivorous;Bd:食性均衡Balanced dietary;C:肉食性Carnivorous;I虫食性Insectivorous;S:夏候鸟Summer bird;Wf:少冬候鸟Winter bird fwe;Br:居留型均衡Balanced residency;T:旅鸟Traveling bird;R:留鸟Resident bird | |||
鸟类多样性差异影响因素分析如下表(表 3)。从组成差异(Diff)看, 有显著关系的四个因子中城市距离及海拔因子均由城市区位决定, 且其差异程度最终会通过自然环境因子差异表现, 而温度与降雨量也属于自然环境因子, 因此可以看出造成城市鸟类组成差异的因子还是以自然环境因子为主导, 这符合生物多样性的生产力假说;其中物种替换(Repl), 反映的是多样性相当情景下的种类差异性, 与其有显著关系的因子仍是自然环境因子为主, 但与组成差异(Diff)相比, 温度与降水等直接因子的影响显著降低, 而海拔因子影响则显得相对提高, 即城市相对独特的地理条件对鸟类物种的替换作用明显, 反映了城市周边鸟类自然类群对城市鸟类多样性构成差异的基础性作用, 而植物丰富度和建成区面积对物种替换呈负相关关系, 即城市面积与植物丰富度差异越大鸟类物种替换度相对越小, 这可以结合丰富度差异(Rich)来解释, 如两个面积及植物丰富度差异较大的城市, 其鸟类组成差异(Diff)主要通过丰富度差异(Rich)来体现, 而物种替换(Repl)度反而较小, 也即城市面积及植物丰富度差异越小而自然环境因子差异大的城市物种替换度越大;从丰富度差异(Rich)看, 植物丰富度等城市自身自然环境因子成为重要的影响因素, 也即生态金字塔的下层物种丰富度对鸟类种类丰富度有显著影响, 同时面积效应在此也发挥了重要作用, 这为通过植物丰富度提升城市鸟类多样性提供了证据。
| 多样性差Fiversity differences 影响因子Impact factors |
组成差异 Composition differences(Diff) |
物种替换 Species replacement (Repl) |
丰富度差异 Difference in richness (Rich) |
|||||
| R | P | R | P | R | P | |||
| 城市距离City distance | 0.643 | 0.001 | 0.440 | 0.001 | -0.014 | 0.510 | ||
| 海拔Altitude | 0.382 | 0.014 | 0.306 | 0.006 | -0.049 | 0.613 | ||
| 年均温度Annual temperature | 0.612 | 0.001 | 0.244 | 0.001 | 0.147 | 0.057 | ||
| 年均降雨量Annual rainfall | 0.455 | 0.001 | 0.297 | 0.001 | 0.004 | 0.392 | ||
| 绿视率Green view index | 0.148 | 0.094 | -0.012 | 0.554 | 0.100 | 0.098 | ||
| 绿化覆盖度Greening coverage | 0.117 | 0.243 | 0.016 | 0.462 | 0.056 | 0.240 | ||
| 植物丰富度Plant richness | 0.167 | 0.066 | -0.300 | 0.998 | 0.375 | 0.002 | ||
| 人口密度Population density | 0.194 | 0.064 | 0.022 | 0.413 | 0.097 | 0.134 | ||
| 建成区面积Built-up Area | -0.050 | 0.608 | -0.478 | 1.000 | 0.408 | 0.001 | ||
| R为相关系数;P为相关性显著水平 | ||||||||
分析得知城市鸟类是我国鸟类多样性的重要构成, 但城市鸟类多样性更具脆弱性, 城市鸟类多样性保护更需要科学、精准和对应性策略。10个地域型较鸟类地理区有更细化的空间划分和一致的鸟类多样特征, 鸟类替换(Repl)因素分析说明城市区位决定的其周边鸟类自然类群对城市鸟类多样性差异的基础性作用, 而植物丰富度等城市自身的自然环境因子对城市鸟类丰富度(Rich)影响显著。因此有必要从宏观尺度入手, 构建“地域型-城市-绿地”多尺度多层级的城市鸟类多样性保护体系, 在地域型尺度重点着眼于各类型鸟类多样性特征的差异性, 城市尺度主要考虑城市鸟类类群与周边自然类群的扩散关系, 而绿地尺度则是在上层策略框架下重点关注于植物丰富度等生境的营造。
3.1 地域型尺度城市鸟类多样性保护路径鉴于城市植被对鸟类多样性的支撑作用, ①首先, 应根据城市鸟类多样地域型特征, 加强其与城市植被绿地依存关系的研究, 建议在中国植被分区调查基础上制定城市鸟类地域型绿化植物选择导则, 由此形成基于地域型集体尺度上的城市绿化价值观、规则和行动, 不仅只是减少城市增量扩张时期不当城市绿化对自然的生态足迹和生态债务[37], 更着眼于城市绿地对鸟类多样性提升的正向价值, 以宏观指导各地域型城市的植物多样性规划。②其次, 由于不同地域型鸟类多样性差异, 需加强各地域型不同时令鸟类组成更替的细化研究, 以便针对性营建鸟类更合适的栖息地。如就居留类型差异而言, 由于旅鸟的灭绝率和恢复率与绿地面积负相关, 而留鸟的灭绝率和恢复率与绿地距城市核心区的距离负相关[28, 38], 因此在高-肉-旅多型及高-虫-留鸟型等旅鸟及留鸟分别占比大的地域型进行城市绿地布局时, 前者则要注重绿地面积的保障, 而后者则要关注绿地的位置;再如均-均-均衡型所处的长江下游是全球鸟类最大的越冬地[39], 而越冬鸟种数与冬季挂果树种数呈正相关, 且越冬鸟类物种相似性与树种相似性较为一致[40—41], 由此该地域型应着重考虑越冬鸟类的具体物种特征及其食源及繁殖需求;就食性结构类型而言, 如虫食性鸟类对绿地的稳定性和面积更敏感[42], 且多在灌木及地被层取食[43], 由此高-虫-留鸟型及低-虫-留鸟型虫食性鸟类比重较高的城市不仅要更加重视绿地面积的保障, 也要注重灌木层和地被层的质量;而植食性鸟类主要在乔木层取食, 则相应类型城市则应注重乔木层的营建。
3.2 城市尺度下城市鸟类多样性保护路径由于城市所在区域的自然环境因子对城市间鸟类的物种替换(Repl)有重要影响, 即每个城市在各自地域型的背景下形成各自独特的类群, 而城市中每个子类群也反映了城市鸟类内部资源利用及种间共存关系, 这其中可能会形成数个觅食及繁殖方式相似的“共位群”。因此对具体城市而言, 在其对应地域型导则下, ①首先, 可在每个共位群中筛选确定一个“伞护种”, 标准是其在共位群中的代表性及关键性, 深入研究伞护种分布、扩散及生存能力受到的影响, 制定针对伞护种的保护策略, 则共位群集体受益。②其次, 要考虑具体城市鸟类类群的独特性, 在城市发展建设中考虑独特性类群的生境需求, 特别是关注对该类群有不可替代生境价值的关键性地段或区域, 予以保留与保护, 如北京市肉食性鸟类丰富且旅鸟占比较高, 这就与其处在重要的旅鸟迁徙通道的区位和山区与平原过渡带的独特环境条件有关。③第三, 重点关注城市中现有栖息地特化本地鸟类种的生境需求, 特别是分布区狭窄的特有种[44], 由于其生境需求的特殊性, 一般扩散能力较弱[45], 一旦栖息地丧失将很难再次引入;④第四, 是对已由城市化影响而迁出城市的本地种, 应通过城市鸟类物种历史变化及与相邻区域的对比, 确定关键恢复鸟种, 利用城市种群与自然种群之间通过扩散发生联系的规律[38], 建立“栖息地需求分析—栖息地修复指引—栖息地恢复目标”的逻辑思路, 逐步恢复被城市化过滤掉的本地鸟类。
3.3 绿地尺度下城市鸟类多样性保护策略 3.3.1 城市绿地植物选择与其丰富度植物丰富度是除建成区面积外导致城市鸟类丰富度差异(Rich)的首要原因, 由此在城市绿地营造中植物的选择应把其种类的多样性放在极为重要的位置, 但同时避免以外来植物为主的多样性。城市绿地及景观营建过程中美学标准被作为重要准则, 该准则下的植物选择成为外来植物引入的主要途径, 从而城市成为外来植物主导的生物多样性贫乏区域[46], 而外来植物经常由于无法提供本地鸟类所需的食物源和栖息需求而导致本地鸟类物种多样性下降[47], 相反, 乡土植物可为本地鸟类提供的益处却比外来植物多10—50倍[48], 结果就是城市中本地鸟类物种的丰富度及数量与本地植物的比例呈正相关, 而外来鸟类种数与外来植物的比例正相关[49]。由此在绿地营建中应选择地带性植被建群种与优势种之中能适应城市生境的近缘种类, 尽最大可能的丰富植物种类及避免简单便利化的纯林营造手法, 构建以多样化乡土植物支撑本地鸟类为主体的近自然型生物群落, 同时兼顾美学准则在植物景观塑造上使其成为能够表征城市区域特征的生命地标。
3.3.2 城市绿地群落结构植物群落结构与植物种类丰富度相辅相成, 追求植物种类丰富性的前提下兼顾群落结构也是提升鸟类多样性的重要途径, 相关研究也证实植被结构特征对鸟类多样性的保护甚至比景观格局更具实际意义[9]。目前在城区植物群落的形成、更新及演替, 普遍上是按人的绿化政策发展的“偏途演替”[50], 群落结构简单且同质性特征远远高于郊区, 其生存策略、生活型及系统分类具有高度相似性[51]。已有多个研究表明植物群落特征和鸟类多样性关系密切, 随着植被种类及结构复杂性的增加鸟类多样性也会提高[52], 尤其灌木层的质量提高对鸟类多样性非常关键[3], 同等情况下小乔木越密大乔木越高越有利于提升鸟类丰富度[53], 同时将不同直径的树木互相搭配以及避免群落的叶群集中在特定的层次结构中, 也更有利于鸟类多样性的维持[54—55]。当然由于具有不同营巢、觅食和迁徙偏好的鸟类对城市的耐受力有所差别, 这些一般性准则也要根据地域型鸟类多样性特征有所调整, 如高纬度城市中高位觅食对鸟类城市适应性起决定作用[2], 因此北方地区的低-杂(均)-夏多型城市需要更加注重乔木层的营建。总体来说城市绿地群落结构宜以近自然化为导向, 营造复层—异龄—混交的群落结构, 并优化景观外貌及栽植方式等, 充分发挥物种共存的自维持机制和作用, 以提升鸟类多样性。
4 结语快速城市化背景下, 城市鸟类多样性在多种尺度上面临着前所未有的威胁与压力, 通过恰当的城市绿地营建以保护鸟类的多样性显得尤为重要, 这既是在各种变化与冲击下维持与提升城市鸟类多样性的重要手段, 也是目前促进城市人居环境高质量发展的关键所在。
本文通过对多个城市鸟类多样性特征的对比与分析, 形成对城市鸟类多样性整体性与地域性特征及规律的一些认识。研究发现仅样本城市鸟类种数既占全国鸟类总种数的一半以上, 表明城市空间对我国鸟类多样性的维持有重要作用, 但同时城市鸟类多样性相对于自然地域更具脆弱性, 以上两点说明维护与提升城市鸟类多样性具有实践及研究上的重要意义;同时发现城市鸟类多样性特征具有显著的地域聚集性, 地域型的划分基本符合鸟类地理区分布但比其更加细化;城市区位是影响鸟类种替换的首要因素, 而植物丰富度等城市自身内部环境条件是导致城市鸟类丰富度差异的重要因子, 这证实了城市绿地营造中植物种类选择的丰富性等对鸟类多样性支撑的重要性。基于以上结论本研究认为, 对城市鸟类多样性的维护与提升需要从城市鸟类多样性地域型的层面认识并思考, 建议构建“地域型-城市-绿地”三级尺度及层级的保护框架。目前我国的鸟类地理区是动物地理区的延用, 而鸟类迁徙灵活及活动范围广, 可能存在对鸟类分布的针对性不强的问题, 而在城市尺度上城市数量多, 属于具体策略的落地层次, 所以这其中的“地域型”尤为关键, 可重点通过地域型层面构建鸟类多样性保护在集体尺度上价值观、规则和行动, 共享地域成果, 并充分凸显保护价值, 以促进城市鸟类多样性维持与提升。
城市鸟类多样性受区域环境、城市规模与开发强度、人为干扰特点及强度、鸟类迁徙路线等多重因素叠加影响, 使得城市鸟类多样性及与环境变量之间的关系非常复杂。目前我国仍缺乏长期大尺度的陆地鸟类监测工作, 本研究仅是在对有限样本城市鸟类多样性特征分析基础上进行的初步思考, 并未考虑城市中鸟类种的珍稀度、数量及多度等问题, 这对城市间鸟类多样性特征及城市栖息地的重要性认识造成了一定的限制性。同时, 在对城市鸟类多样性影响因素的分析中, 受数据可获得性限制涵盖的相关因子不够全面, 本文提出的相关策略是基于现有分析结果的应对, 未来对城市鸟类多样性地域型的划分仍需深入研究。
| [1] |
马远, 李锋, 杨锐. 城市化对生物多样性的影响与调控对策. 中国园林, 2021, 37(5): 6-13. |
| [2] |
吴佳雨, 杨梦夏, 周侃. 基于公民科学数据的中国鸟类城市化响应及适应性. 地理学报, 2023, 78(11): 2850-2863. |
| [3] |
福井亘. 城市的生物多样性与环境——以日本城市与近郊的鸟类指标为例. 西部人居环境学刊, 2019, 34(3): 8-18. |
| [4] |
张庆费, 许源. 城市森林生物多样性保育途径探讨. 中国城市林业, 2016, 14(3): 1-5. |
| [5] |
Gregory R D, van Strien A, Vorisek P, Gmelig Meyling A W, Noble D G, Foppen R P B, Gibbons D W. Developing indicators for European birds. Philosophical Transactions of the Royal Society of London Series B, Biological Sciences, 2005, 360(1454): 269-288. DOI:10.1098/rstb.2004.1602 |
| [6] |
张征恺, 黄甘霖. 中国城市鸟类学研究进展. 生态学报, 2018, 38(10): 3357-3367. |
| [7] |
兰思仁, 李霄鹤, 彭东辉, 余韵. 城市建设中的生物多样性及其保育对策. 中国城市林业, 2013, 11(5): 10-13. |
| [8] |
Callaghan C T, Bino G, Major R E, Martin J M, Lyons M B, Kingsford R T. Heterogeneous urban green areas are bird diversity hotspots: insights using continental-scale citizen science data. Landscape Ecology, 2019, 34(6): 1231-1246. DOI:10.1007/s10980-019-00851-6 |
| [9] |
毛齐正, 马克明, 邬建国, 唐荣莉, 张育新, 罗上华, 宝乐, 蔡小虎. 城市生物多样性分布格局研究进展. 生态学报, 2013, 33(4): 1051-1064. |
| [10] |
Bonier F, Martin P R, Wingfield J C. Urban birds have broader environmental tolerance. Biology Letters, 2007, 3(6): 670-673. DOI:10.1098/rsbl.2007.0349 |
| [11] |
Sol D, González-Lagos C, Lapiedra O, Díaz M. Why are exotic birds so successful in urbanized environments? Murgui E, Hedblom M. Ecology and Conservation of Birds in Urban Environments. Cham: Springer, 2017: 75-89.
|
| [12] |
Conole L E, Kirkpatrick J B. Functional and spatial differentiation of urban bird assemblages at the landscape scale. Landscape and Urban Planning, 2011, 100(1/2): 11-23. |
| [13] |
Bellocq M I, Leveau L M, Filloy J. Urbanization and bird communities: spatial and temporal patterns emerging from southern South America. Murgui E, Hedblom M. Ecology and Conservation of Birds in Urban Environments. Cham: Springer, 2017: 35-54.
|
| [14] |
殷鲁秦, 王成, 韩文静. 基于取食行为探究北京居民区鸟类的食源特征及多样性. 生物多样性, 2023, 31(5): 37-50. |
| [15] |
Fujita A, Maeto K, Kagawa Y, Ito N. Effects of forest fragmentation on species richness and composition of ground beetles (Coleoptera: Carabidae And Brachinidae) in urban landscapes. Entomological Science, 2008, 11(1): 39-48. DOI:10.1111/j.1479-8298.2007.00243.x |
| [16] |
Smith R M, Thompson K, Hodgson J G, Warren P H, Gaston K J. Urban domestic gardens (IX): composition and richness of the vascular plant flora, and implications for native biodiversity. Biological Conservation, 2006, 129(3): 312-322. DOI:10.1016/j.biocon.2005.10.045 |
| [17] |
谢世林, 曹垒, 逯非, 欧阳志云. 鸟类对城市化的适应. 生态学报, 2016, 36(21): 6696-6707. |
| [18] |
Biamonte E, Sandoval L, Chacón E, Barrantes G. Effect of urbanization on the avifauna in a tropical metropolitan area. Landscape Ecology, 2011, 26(2): 183-194. DOI:10.1007/s10980-010-9564-0 |
| [19] |
顾灵桢, 闫淑君, 吕蔚, 程敏, 胡文静. 福州城市绿地冬季鸟类多样性分析. 林草资源研究, 2024(1): 125-133. |
| [20] |
王春晓, 何建华, 刘殿锋, 于秀波, 石青青. 土地利用变化对鸟类栖息地连通性的影响——以鄂州市为例. 生态学报, 2022, 42(10): 4197-4208. |
| [21] |
游巍斌, 蔡新瑜, 王英姿, 王瑞, 张锦琳, 陈莹, 邱晓月, 方冰, 汤绍圣, 坚文倩. 基于不同网格尺度福州主城区鸟类多样性与景观特征的关系研究. 生态学报, 2023, 43(18): 7670-7681. |
| [22] |
邓卓迪, 丁志锋, 曾小悦, 张春兰, 李孝恩, 何子晴, 梁健超, 胡慧建. 广州城市绿地景观特征及其对鸟类多样性的影响. 四川动物, 2023, 42(5): 568-578. |
| [23] |
刘巷序. 城市绿地景观结构特征对鸟类群落的影响[D]. 上海: 华东师范大学, 2023.
|
| [24] |
张国月, 雷凯茜, 樊雅东, 安玫, 邓树庆, 杨栋, 陈修家. 城市垃圾填埋场生态修复后鸟类多样性变化研究. 环境生态学, 2024, 6(2): 71-76. |
| [25] |
陈珏洁, 郭雨蕊, 阳佩琳, 刘家琳. 塘类海绵生境鸟类多样性特征及其影响因素研究. 中国园林, 2023, 39(11): 111-117. |
| [26] |
邓一荣, 肖荣波, 黄柳菁, 胡慧建, 李智山, 丁晓龙. 城市生物多样性恢复途径与实例研究. 风景园林, 2015(6): 25-32. |
| [27] |
达良俊. 基于本土生物多样性恢复的近自然城市生命地标构建理念及其在上海的实践. 中国园林, 2021, 37(5): 20-24. |
| [28] |
张淑萍. 城市化对鸟类分布的影响. 生态学杂志, 2008, 27(11): 2018-2023. |
| [29] |
郑光美. 中国鸟类分类与分布名录. 4版. 北京: 科学出版社, 2023.
|
| [30] |
国家统计局城市社会经济调查司. 中国城市统计年鉴—2019. 北京: 中国统计出版社, 2020.
|
| [31] |
肖路. 基于街景大数据的城市绿视率与影响因素多尺度分析研究[D]. 哈尔滨: 中国科学院大学(中国科学院东北地理与农业生态研究所), 2021.
|
| [32] |
Podani J, Schmera D. A new conceptual and methodological framework for exploring and explaining pattern in presence-absence data. Oikos, 2011, 120(11): 1625-1638. |
| [33] |
斯幸峰, 赵郁豪, 陈传武, 任鹏, 曾頔, 吴玲兵, 丁平. Beta多样性分解: 方法、应用与展望. 生物多样性, 2017, 25(5): 464-480. |
| [34] |
韩联宪. 中国鸟类地理分布及多样性. 人与自然, 2002(8): 8-19. |
| [35] |
孔颖. 我国鸟类迁徙路线上的湿地资源现状及保护对策. 环境与发展, 2020, 32(3): 201-202, 207. |
| [36] |
冯永军, 胡慧建, 蒋志刚, 王祖望. 物种与科属的数量关系——以中国鸟类为例. 动物学研究, 2006, 27(6): 581-587. |
| [37] |
Canedoli C, Manenti R, Padoa-Schioppa E. Birds biodiversity in urban and periurban forests: environmental determinants at local and landscape scales. Urban Ecosystems, 2018, 21(4): 779-793. |
| [38] |
Husté A, Boulinier T. Determinants of local extinction and turnover rates in urban bird communities. Ecological Applications: a Publication of the Ecological Society of America, 2007, 17(1): 168-180. |
| [39] |
王鑫. 食物对越冬小白额雁分布、能量与氮平衡以及行为的影响[D]. 合肥: 中国科学技术大学, 2013.
|
| [40] |
曹加杰, 白项鸽, 方静. 南京城市公园树种配置对鸟类越冬的影响. 西部林业科学, 2020, 49(6): 43-53. |
| [41] |
郝日明, 张明娟. 中国城市生物多样性保护规划编制值得关注的问题. 中国园林, 2015, 31(8): 5-9. |
| [42] |
Zhou D Q, Fung T, Chu L M. Avian community structure of urban parks in developed and new growth areas: a landscape-scale study in Southeast Asia. Landscape and Urban Planning, 2012, 108(2/3/4): 91-102. |
| [43] |
杨刚, 许洁, 王勇, 丁由中, 袁晓, 裴恩乐, 马波, 王小明, 王正寰. 城市公园植被特征对陆生鸟类集团的影响. 生态学报, 2015, 35(14): 4824-4835. |
| [44] |
武美香, 吴建国, 况明生, 衡涛. 我国鸟类特有种地理分布与气候要素关系的初步分析. 环境科学研究, 2011, 24(4): 409-420. |
| [45] |
Fernández-Juricic E. Bird community composition patterns in urban parks of Madrid: the role of age, size and isolation. Ecological Research, 2000, 15(4): 373-383. |
| [46] |
Doody B J, Sullivan J J, Meurk C D, Stewart G H, Perkins H C. Urban realities: the contribution of residential gardens to the conservation of urban forest remnants. Biodiversity and Conservation, 2010, 19(5): 1385-1400. |
| [47] |
MacKenzie J A, Hinsley S A, Harrison N M. Parid foraging choices in urban habitat and their consequences for fitness. Ibis, 2014, 156(3): 591-605. |
| [48] |
Zhou D Q, Chu L M. How would size, age, human disturbance, and vegetation structure affect bird communities of urban parks in different seasons?. Journal of Ornithology, 2012, 153(4): 1101-1112. |
| [49] |
赵伊琳, 王成, 白梓彤, 郝泽周. 城市化鸟类群落变化及其与城市植被的关系. 生态学报, 2021, 41(2): 479-489. |
| [50] |
王娟, 达良俊, 李艳艳, 商侃侃. 城市化进程中上海植被的多样性、空间格局和动态响应(Ⅴ): 管护放弃后城市水杉林林下植被自然演替格局的研究. 华东师范大学学报: 自然科学版, 2009(6): 12-22. |
| [51] |
Knapp S, Kühn I, Schweiger O, Klotz S. Challenging urban species diversity: contrasting phylogenetic patterns across plant functional groups in Germany. Ecology Letters, 2008, 11(10): 1054-1064. |
| [52] |
Paker Y, Yom-Tov Y, Alon-Mozes T, Barnea A. The effect of plant richness and urban garden structure on bird species richness, diversity and community structure. Landscape and Urban Planning, 2014, 122: 186-195. |
| [53] |
Tilghman N G. Characteristics of urban woodlands affecting breeding bird diversity and abundance. Landscape and Urban Planning, 1987, 14: 481-495. |
| [54] |
Sasaki T, Imanishi J, Fukui W, Morimoto Y. Fine-scale characterization of bird habitat using airborne LiDAR in an urban park in Japan. Urban Forestry & Urban Greening, 2016, 17: 16-22. |
| [55] |
Palomino D, Carrascal L M. Urban influence on birds at a regional scale: a case study with the avifauna of northern Madrid Province. Landscape and Urban Planning, 2006, 77(3): 276-290. |