 2024, Vol. 44
2024, Vol. 44文章信息
- 李明, 王军邦, 张秀娟, 张溢, 王之嵘, 杨永胜
- LI Ming, WANG Junbang, ZHANG Xiujuan, ZHANG Yi, WANG Zhirong, YANG Yongsheng
- 未来气候情景下青藏高原高寒草原与高寒草甸优势种潜在适宜区分布预测
- Distribution of potential suitable areas of dominant species in alpine grasslands and alpine meadows in the TP under future climate scenarios
- 生态学报. 2024, 44(22): 10162-10177
- Acta Ecologica Sinica. 2024, 44(22): 10162-10177
- http://dx.doi.org/10.20103/j.stxb.202310242314
-
文章历史
- 收稿日期: 2023-10-24
- 网络出版日期: 2024-08-22

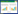




2. 中国科学院地理科学与资源研究所, 生态系统网络观测与模拟重点实验室, 生态系统大数据与模拟中心, 北京 100101;
3. 中国科学院西北高原生物研究所, 中国科学院高原生物适应与进化重点实验室, 西宁 810008
2. National Ecosystem Science Data Center, Key Laboratory of Ecosystem Network Observation and Modeling, Institute of Geographic Sciences and Natural Resources Research, Beijing 100101, China;
3. Key Laboratoryof PlateauAdaptation and Evohution, Northwest stitte of Platea Biology, Chinese Academy of Sciences, Xining 810008, China
全球气候在持续变暖, 并在未来很有可能突破1.5 ℃温升, 使极端气候加剧[1—2]。气候变化可能会改变未来草地空间分布格局, 严重威胁全球陆地生态系统的稳定性[3—4]。青藏高原属大陆性高原气候, 具海拔高, 植被类型多, 地带性差异明显等特点, 是我国乃至亚洲的重要生态屏障, 对气候变化响应更为敏感脆弱[5], 因此评估气候变化对青藏高原草地空间分布及其变迁给草地管理者提出了巨大挑战[6]。
高寒草地是青藏高原最主要的生态系统类型, 是高原畜牧业的基础, 是维持高寒生物多样性和高寒草地生态服务功能的重要条件[7]。近年来青藏高原暖湿化趋势影响高寒草地空间格局, 水热组合格局变化推动草地空间变化[8—9], 未来气候情景下高寒草地分布如何变化, 不仅是全球气候变化研究的主要科学问题, 还是牧业生产和生态保护所面临的重要现实问题[10]。高寒草原与高寒草甸作为高寒草地最主要的两种草地类型, 是研究高寒草地响应未来气候变化的重点[11—13]。白乌云等指出草地优势种分布和数量变化直接影响草地植物群落结构和功能, 是推绎草地生态系统变化趋势的重要组成部分[14];Wang和王譞等发现当前高寒草原与高寒草甸优势种空间分布占高寒草地总面积的92%, 而气候变暖会增加优势种竞争优势[4, 13];Ma等认为优势种耐寒耐旱的生物学特性是高寒草地响应气候变化的关键[15]。高寒草地优势种通常包含早熟禾(Poa annua)类高寒草原, 紫花针茅(Stipa purpurea)类高寒草原, 高山嵩草(Carex parvula)类高寒草甸和矮生嵩草(Carex alatauensis)类高寒草甸[13, 16—19]。目前已有学者对其适宜区变化及响应机制开展研究, 如胡忠俊等利用最大熵模型(MaxEnt)重塑了古气候时期紫花针茅类和嵩草类的空间格局, 发现其起源和冰期避难所是喜马拉雅山与青海湖, 受降水量影响较大, 未来(2080年)其适宜区会出现扩张的情况[18, 20], Zhao等也采用生态位模型研究并发现新疆地区早熟禾多生长于新疆伊犁河流域附近[21]。然而较多研究只是模拟单一物种局部的空间分布, 对于未来青藏高原不同情景下草地变化的预测研究关注较少。
物种分布模型(SDMs)是利用环境变量模拟物种生境适宜性及其适宜区分布变化的主要方法[22]。常用的物种分布模型如规则集的遗传算法模型(Genetic Algorithm for Ruleset Prediction, GARP)、生物种群增长模型(CLIMEX)和最大熵模型(Maximum Entropy Model, MaxEnt)[23—24]等。国内外学者基于受试者工作特征曲线值(Receiver Operating Characteristic, ROC)和Kappa系数对模型模拟的准确性进行评估, 发现MaxEnt评价指标结果表现最好, 且在主导环境因子的筛选、变量的定量描述及生境模拟都表现出极佳的性能[25—26], 此外MaxEnt具有操作简单、运行速度快、预测效果好等优点, 即使在分布样本数量少且不完整的情况下仍表现出较高的预测精度[27], 是谱系地理、种质资源保护以及生境模拟预测等研究领域的首选模型[28—29]。运用ArcGIS和MaxEnt可了解气候变化影响草地优势种空间分布变化, 对草地资源合理利用及畜牧业发展具有重大意义。
本研究基于早熟禾类高寒草原、紫花针茅类高寒草原、高山嵩草类高寒草甸、矮生嵩草类高寒草甸在青藏高原的分布情况, 采用MaxEnt模拟潜在适宜区, 分析影响其空间分布的主导因子, 预测在未来不同气候情景下的空间分布格局变化, 比较高寒草原与高寒草甸两种主要草地类型优势种未来的分布差异, 探究影响其分布差异的原因, 以此推绎未来草地类型空间分布变化。本研究为维持高寒生态系统平衡、保护青藏高原生物多样性、合理配置管理草地提供参考。
1 研究区概况青藏高原(79°19′—104°47′E和26°00′—39°47′N)是中国最大、世界海拔最高的高原, 有“世界屋脊”、“第三极”之称, 总面积约250×104 km2。气候特点是气温低、湿度小, 海拔3000 m以上无夏季, 气温日较差较大, 降水量自高原东南部向西北部递减。区域内主要生态系统类型及面积占比:高寒草原(25.51%)、高寒草甸(38.36%)、高寒荒漠(3.59%)、温性草原(2.97%)、温性荒漠(3.17%)等17种草地类型, 主要草地类型为高寒草甸和高寒草原, 占青藏高原总面积的63.87%(图 1)。

|
| 图 1 优势种青藏高原分布点 Fig. 1 Distribution of dominant species on the Qinghai-Tibetan Plateau |
物种分布数据主要来源于全球生物多样性信息网络(Global Biodiversity Information Facility, GBIF, http://www.gbif.org/)和物种生长季(6—8月)进行的野外调查, 物种标本信息通过在线查阅中国数字植物标本馆(CVH, http://www.cvh.ac.cn)。为使物种分布数据更具时效性, 在GBIF中初步筛选出记录了经纬度的物种样点;参考《中国嵩草草甸》、《中国植物志》[30—31]和相关文献[32—34]中所提及的研究物种分布范围, 并将第二次青藏科考数据作为补充。最终得到早熟禾类高寒草原、紫花针茅类高寒草原、高山嵩草类高寒草甸和矮生嵩草类高寒草甸的地理分布样点数量分别为176、195、178和121(图 1), 保存为CSV格式。
2.2 气候数据来源及预处理本研究共选取19个生物气候因子(bio1—bio19)和1个地形数据用于模型构建, 以上数据均下载自世界气候数据库(http://worldclim.org/), 采用2.1版最新更新的版本。初始模型构建采用1970—2000年时间段气候数据, 空间分辨率为30″(约1 km)。未来气候数据选自WorldClim网站中适合我国地理环境的BCC-CSM2-MR气候模式下的共享社会经济路径(Shared socio-economic path, SSPs), 包括中期(2041—2060, 2050s)和远期(2061—2080, 2070s)这两个时间段下的4种排放路径情景, 即SSP126(可持续发展路径), SSP245(中间路径), SSP370(区域竞争路径), SSP585(以传统化石燃料为主的路径), 空间分辨率为2.5′(约5 km)[35—37], 海拔数据空间分辨率为2.5′(约5 km)。上述获取数据均选用1984世界大地坐标系(World Geodetic System, WGS 1984)进行空间投影[38]。
本研究使用刀切检验法(Jackknife)评估环境变量对模型预测结果的重要性, 通过逐一去除某个环境变量再重新训练模型并比较模型性能变化, 从而确定该变量对模型的重要性。然而WorldClim提供的某些气候变量数据之间可能存在强相关性[27], 为消除可能出现的多重共线性问题影响刀切法的评估结果, 使用皮尔逊分析法测试环境变量之间的相关性(图 2)。然后将所有环境变量数据导入最大熵模型, 根据刀切检验结果剔除贡献率为0的气候因子并结合相关性对剩下的因子做进一步分析, 如果两个因子之间的相关性系数|R|≥0.8, 保留贡献率高的气候因子[39], 最后16个环境变量作为本研究的评估因子, 分别是:年平均气温(bio1)、平均温度日较差(bio2)、等温性(bio3)、气温季节性变动系数(bio4)、最冷月份最低温度(bio6)、气温年较差(bio7)、最湿季度平均温度(bio8)、最干季度平均温度(bio9)、最冷季度平均温度(bio11)、年降水量(bio12)、最干月份降水量(bio14)、降水量季节性变化(bio15)、最干燥季度降水量(bio17)、最暖季度降水量(bio18)、最冷季度降水量(bio19)、海拔(elev)(表 1)。

|
| 图 2 草地优势环境变量相关性图 Fig. 2 Correlation diagram of environmental variables for grassland dominant species Bio1:年平均气温;Bio2:平均温度日较差;Bio3:等温性;Bio4:气温季节性变动系数;Bio5:最热月份最高温度;Bio6:最冷月份最低温度;Bio7:气温年较差;Bio8:最湿季度平均温度;Bio9:最干季度平均温度;Bio10:最热季度平均温度;Bio11:最冷季度平均温度;Bio12:年降水量;Bio13:最湿月份降水量;Bio14:最干月份降水量;Bio15:降水量季节性变化;Bio16:最湿季度降水量;Bio17:最干燥季度降水量;Bio18:最暖季度降水量;Bio19:最冷季度降水量;Elev:海拔 |
| 环境因子代号 Code of environmental factor |
英文名称 English name |
中文名称 Chinese name |
单位 Unit |
| Bio1 | Annual mean temperature | 年平均气温 | ℃ |
| Bio2 | Mean diurnal range (mean of monthly (max temp-min temp)) | 平均温度日较差 | ℃ |
| Bio3 | Isothermality (Bio2/Bio7×100) | 等温性 | — |
| Bio4 | Temperature seasonality (standard deviation×100) | 气温季节性变动系数(标准偏差×100) | — |
| Bio6 | Min Temperature of coldest month | 最冷月份最低温度 | ℃ |
| Bio7 | Temperature annual range | 气温年较差 | ℃ |
| Bio8 | Mean temperature of wettest quarter | 最湿季度平均温度 | ℃ |
| Bio9 | Mean temperature of driest quarter | 最干季度平均温度 | ℃ |
| Bio11 | Mean temperature of coldest quarter | 最冷季度平均温度 | ℃ |
| Bio12 | Annual precipitation | 年降水量 | mm |
| Bio14 | Precipitation of driest month | 最干月份降水量 | mm |
| Bio15 | Precipitation seasonality | 降水量季节性变化 | — |
| Bio17 | Precipitation of driest quarter | 最干燥季度降水量 | mm |
| Bio18 | Precipitation of warmest quarter | 最暖季度降水量 | mm |
| Bio19 | Precipitation of coldest quarter | 最冷季度降水量 | mm |
| Elev | Elevation | 海拔 | m |
将物种分布数据(CSV文件)和环境变量(ASC)导入模型, 选择绘制响应曲线和制作预测图片以及刀切法(Jackknife)计算各因子贡献率和置换重要性。选择逻辑变换(Logistic)为输出方式, 其参数组与MaxEnt相同, 适用性更强[40]。重复迭代方式(Replicated run type)选择自举检验法(Bootstrap), 随机选择25%的分布点作为测试集, 剩下75%的分布点作为训练集, 模型重复运行20次, 以此消除训练数据与测试数据产生的误差[41—42]。此外, 采样点从10000个背景网格单元中随机挑选(Random seed)并记下绘图数据(Write plot data)[43], 其余参数保持默认值。
通常使用受试者工作特征曲线(Receiver Operating Characteristic, ROC曲线)下的面积值(Area Under Curve, AUC)评估模拟结果的准确度, 取值范围介于0—1之间, AUC值越接近1, 模型预测能力越高, 预测结果可划分为以下指标区间:非常好(0.90—1.00), 良好(0.80—0.90), 一般(0.70—0.80), 勉强可接受(0.60—0.70), 失败(0.50—0.60)[27]。
模型运行结束后可获得预测物种的适生指数图, 文件格式为“ASC”。在ArcGIS软件中使用转换工具将模型输出的“ASC”文件转化成值域为[0, 1]的栅格图层, 阈值越大, 表明预测物种在研究区域的适生率越高。基于IPCC第六次评估报告中对“可能”的定义, 确定划分适生区与非适生区的阈值是0.2[44]。使用ArcGIS将各草地优势种叠加并结合SDM Toolbox计算草地空间分布质心, 迁移轨迹及分布面积变化[20], 分析未来气候在不同情境下的潜在适宜区空间变化及不同优势种的迁移能力。
2.4 其他数据与软件说明草地类型数据下载自于《1 ∶ 100万中国草地资源图集》[45], 空间分辨率为1 km。MaxEnt软件下载自网页资源(http://biodiversity informatics.amnh.org/), 版本为3.4.4;SDM toolbox版本为v2.5, 从网页资源(http://www.sdmtoolbox.org/downloads)获取。
3 结果与分析 3.1 模型精度和主导环境变量对当前和未来预测的重要性ROC曲线检验结果显示, 在当前气候下高寒草甸优势种与高寒草原优势种的AUC值都超过0.9, 表明最大熵模型运算效果好, 预测物种适宜分布区可靠性高(图 3)。

|
| 图 3 高寒草原优势种与高寒草甸优势种ROC曲线验证和AUC值 Fig. 3 ROC curve and AUC value for the dominant of alpine grassland and alpine meadows ROC:受试者工作特征曲线Receiver operating characteristic curve;AUC:ROC曲线下的面积Area under the curve |
刀切检验的贡献率结果可以确定影响物种分布的主导环境因子(表 2)。当前和未来贡献率结果显示海拔(Elev)和温度(bio2, bio11)对早熟禾类高寒草原和矮生嵩草类高寒草甸影响最大, 未来海拔与温度影响其分布的权重将增加;紫花针茅类高寒草原与高山嵩草类高寒草甸分布受降水量(bio12, bio17)限制, 且未来情景下降水量对其影响愈加明显。当前贡献率结果表明, 年降水量(bio12)、海拔(Elev)、平均温度日较差(bio2)及降水量季节性变化(Bio15)对4类草地分布具有重要影响, 且各类草地对环境变量的响应强度存在差异, 其中年降水量, 平均温度日较差及降水量季节性变化对早熟禾类高寒草原影响较小;高山嵩草类高寒草甸对海拔响应较弱, 反之对降水量, 平均温度日较差较为强烈。
| 环境变量 Variables |
早熟禾类高寒草原 P. annua alpine grassland |
紫花针茅类高寒草原 S. purpurea alpine grassland |
高山嵩草类高寒草甸 K. humilis alpine meadow |
矮生嵩草类 C. alatauensis alpine meadow |
|||||||
| 当前 Current |
未来 Future |
当前 Current |
未来 Future |
当前 Current |
未来 Future |
当前 Current |
未来 Future |
||||
| Bio1 | 0.3 | 0.9±0.6 | |||||||||
| Bio2 | 2.7 | 3.45±2.05 | 8.5 | 12.3±3.5 | 21.5 | 27.2±2 | 21.4 | 26.2±3 | |||
| Bio3 | 4.2 | 4.25±2.35 | 2.1 | 4.5±2.4 | |||||||
| Bio4 | 8.7 | 5.25±3.55 | 5.8 | 3.85±1.35 | |||||||
| Bio6 | 18.6 | 9.6±1.6 | 12.1 | 7.55±1.05 | |||||||
| Bio7 | 19.4 | 14.7±3.9 | 9.2 | 6.65±3.15 | |||||||
| Bio8 | 0.9 | 3.85±1.65 | |||||||||
| Bio9 | 1.8 | 1.85±0.65 | |||||||||
| Bio11 | 48.3 | 50±9.7 | |||||||||
| Bio12 | 6.6 | 8.85±3.35 | 30.6 | 21.35±2.85 | 42.6 | 43.65±3.95 | 32.7 | 36.8±5.4 | |||
| Bio14 | 7.1 | 6.55±2.15 | |||||||||
| Bio15 | 2.2 | 5±4.5 | 4.2 | 3.2±1.2 | 5.5 | 10.5±3.2 | 6.4 | 5.8±1.25 | |||
| Bio17 | 5.3 | 5.95±2.65 | 14.1 | 17.5±2.7 | |||||||
| Bio18 | 5.3 | 3.3±3.2 | 1.7 | 1.8±0.75 | |||||||
| Bio19 | 2.2 | 1.7±0.35 | |||||||||
| Elev | 16.6 | 15.55±9.75 | 17.1 | 22.1±4.7 | 2.4 | 3.9±2.1 | 10.5 | 12.4±4.7 | |||
环境变量的响应曲线可以判断物种生境的适宜性范围, 存在概率>0.5表明最适宜物种生境区间(图 4)。受最冷季度平均温度(bio11)影响, 早熟禾类高寒草原适宜范围位于青藏高原东南部, 并且在适宜范围偏南部达到最佳生境条件。紫花针茅类高寒草原与高山嵩草类、矮生嵩草类高寒草甸对年降水量适宜范围较为相似, 适宜范围位于青藏高原中部、东北及西南部。受海拔高度(Elev)影响, 紫花针茅类高寒草原和高山嵩草类、矮生嵩草类高寒草甸的适宜范围沿重要山脉分布, 如祁连山, 喜马拉雅山, 横断山等, 且随着海拔的升高, 矮生嵩草类高寒草甸生存能力更强;相比之下, 早熟禾类高寒草原较适应低海拔地区, 能在祁连山周边的低海拔地区生存, 但在喜马拉雅山生长受限。受平均温度日较差(bio2)影响, 紫花针茅类高寒草原及高山嵩草类、矮生嵩草类高寒草甸的适宜范围集中在青藏高原中部;早熟禾类高寒草原适宜范围位于青藏高原东部至南部边界地区。受降水量季节性变化(bio15)影响, 紫花针茅类高寒草原与高山嵩草类、矮生嵩草类高寒草甸的适宜范围位于青藏高原中部, 早熟禾类高寒草原位于青藏高原西南地区。

|
| 图 4 当前情景下重要环境变量响应曲线及适生区间分布 Fig. 4 Response curve of Important environmental variables and interallogenic distribution under current scenario |
利用ArcGIS软件提取青藏高原优势种潜在适宜区(图 5)。当前气候条件下高寒草地优势种在青藏高原的潜在适宜区面积为(113.68—162.14)×104 km2, 集中分布于三江源、祁连山、唐古拉山、喜马拉雅山以及冈底斯山等地区(表 3)。其中紫花针茅类高寒草原与高山嵩草类高寒草甸及矮生嵩草类高寒草甸分布较为相似, 都是集中在青藏高原东部和中部海拔较高的地区, 主要环境因子也都是年降水量。相比于紫花针茅类高寒草原, 早熟禾类高寒草原适应能力更强且能在降水量多、海拔低的地区生存, 主要适宜区分布在青藏高原东部及南部地区, 分布范围包括四川省、云南省、青海省中部、西藏南部以及新疆和甘肃省的部分区域。

|
| 图 5 当前气候下早熟禾类(Ⅰ), 紫花针茅类(Ⅱ) 高寒草原与高山嵩草类(Ⅰ), 矮生嵩草类(Ⅱ) 高寒草甸在青藏高原的潜在适宜区分布 Fig. 5 Distribution of potential suitable areas of the Qinghai-Tibetan Plateau of alpine grassland of P. annua (Ⅰ) and S. purpurea (Ⅱ), alpine meadow of C. parvula (Ⅰ) and C. alatauensis (Ⅱ) under the current climate |
| 名称 Name |
适宜区/×104 km2 Suitable area |
适宜区比例/% Suitable area ratio |
名称 Name |
适宜区/×104 km2 Suitable area |
适宜区比例/% Suitable area ratio |
|
| 早熟禾类(P. annua) | 122.33 | 48.93 | 高山嵩草类(K. humilis) | 113.68 | 45.47 | |
| 紫花针茅类(S. purpurea) | 162.14 | 64.85 | 矮生嵩草类(C. alatauensis) | 138.24 | 55.29 |
在未来气候情景下, 高寒草原与高寒草甸优势种的主要适宜区仍然分布在青藏高原中部、东部及南部地区(图 6)。早熟禾类与紫花针茅类高寒草原的地理分布在未来均出现扩张和丧失的情况, 早熟禾类适宜区丧失远大于扩张, 丧失率为1.27%—13.42%, 丧失区域集中在青藏高原东部地区, 但是在2040—2060年SSP370路径和2060—2080年SSP245路径下青藏高原南部出现明显扩张趋势, 扩张率为6.07%—8.62%;而紫花针茅类适宜区具备明显的扩张现象, 适宜区由青藏高原中部向西和东南方向扩张, 扩张率为8.03%—13.62%;高山嵩草类与矮生嵩草类高寒草甸的适宜区扩张远大于丧失, 高山嵩草类的扩张率为11.56%—29.51%, 在SSP370情景下扩张最大, 由青藏高原中部向西扩张, 但是在2060—2080年SSP585路径下出现丧失的情况, 丧失区域多集中在青藏高原中部, 丧失率为6.86%;矮生嵩草类的扩张率为13.95%—27.53%, 由当前适宜区向四周扩张, 只有在2040—2060年SSP585路径下出现丧失的情况, 丧失区域多集中在青藏高原东部地区, 丧失率为17.67%(图 7、表 4)。总体而言, 未来优势种适宜区会向喜马拉雅山以南迁移, 青海湖以北出现小规模扩张;紫花针茅类高寒草原和高山嵩草类、矮生嵩草类高寒草甸的潜在适宜区在未来气候变化情景下具有较好的前景, 在多数情景下均保持扩张的趋势, 而早熟禾类高寒草原适生区只有在中期的SSP245和远期的SSP370情景下才能扩张, 其他情景皆丧失(图 6, 7)。

|
| 图 6 未来气候情境下早熟禾类(Ⅰ), 紫花针茅类(Ⅱ) 高寒草原与高山嵩草类(Ⅰ), 矮生嵩草类(Ⅱ) 高寒草甸在青藏高原的的潜在适宜区分布 Fig. 6 Distribution of potential suitable areas of the Qinghai-Tibet Plateau of P. annua (Ⅰ) and S. purpurea (Ⅱ), alpine meadow of C. parvula (Ⅰ) and C. alatauensis (Ⅱ) under future climate change scenarios in the future |

|
| 图 7 未来气候情景对比当前气候情景下早熟禾类, 紫花针茅类高寒草原与高山嵩草, 矮生嵩草类高寒草甸在青藏高原的潜在适宜区空间变化 Fig. 7 Spatial variation of potential habitat of P. annua and S. purpurea, alpine meadow of C. parvula and C. alatauensis under the future different climate scenarios compared with modern climate scenarios |
| 物种名称 Species Name |
气候情景 Climate scenario |
时期 Period |
适宜区 Suitable area |
适宜区变化比例 Change rate/% |
| 早熟禾类高寒草原 P. annua alpine grassland |
SSP126 | 2041—2060 | 86.05 | -13.42 |
| 2061—2080 | 103.90 | -6.83 | ||
| SSP245 | 2041—2060 | 113.45 | -3.37 | |
| 2061—2080 | 139.05 | 6.07 | ||
| SSP370 | 2041—2060 | 145.68 | 8.62 | |
| 2061—2080 | 115.34 | -2.6 | ||
| SSP585 | 2041—2060 | 119.15 | -1.27 | |
| 2061—2080 | 112.37 | -3.77 | ||
| 紫花针茅类高寒草原 S. purpurea alpine grassland |
SSP126 | 2041—2060 | 197.10 | 12.90 |
| 2061—2080 | 189.65 | 10.14 | ||
| SSP245 | 2041—2060 | 199.44 | 13.62 | |
| 2061—2080 | 186.92 | 9.00 | ||
| SSP370 | 2041—2060 | 190.30 | 10.38 | |
| 2061—2080 | 187.51 | 9.35 | ||
| SSP585 | 2041—2060 | 198.28 | 13.20 | |
| 2061—2080 | 184.30 | 8.03 | ||
| 高山嵩草类高寒草甸 C. parvula alpine meadow |
SSP126 | 2041—2060 | 151.34 | 13.89 |
| 2061—2080 | 158.93 | 16.70 | ||
| SSP245 | 2041—2060 | 154.22 | 14.85 | |
| 2061—2080 | 159.27 | 16.71 | ||
| SSP370 | 2041—2060 | 193.58 | 29.51 | |
| 2061—2080 | 163.25 | 18.30 | ||
| SSP585 | 2041—2060 | 145.32 | 11.56 | |
| 2061—2080 | 95.34 | -6.86 | ||
| 矮生嵩草类高寒草甸 C. alatauensis alpine meadow |
SSP126 | 2041—2060 | 212.81 | 27.53 |
| 2061—2080 | 176.05 | 13.95 | ||
| SSP245 | 2041—2060 | 206.34 | 25.00 | |
| 2061—2080 | 188.30 | 18.34 | ||
| SSP370 | 2041—2060 | 191.62 | 19.70 | |
| 2061—2080 | 180.45 | 15.57 | ||
| SSP585 | 2041—2060 | 90.67 | -17.67 | |
| 2061—2080 | 186.00 | 10.36 |
当前气候情景下高寒草原优势种的潜在适宜区质心地理坐标为33°08′N、94°58′E, 位于杂多县东部;高寒草甸优势种的潜在适宜区质心地理坐标为33°40′N、97°43′E, 位于石渠县西部, 两质点间的距离为261.95 km(图 8, 表 5)。

|
| 图 8 不同气候情境下高寒草原与高寒草甸优势种潜在适宜区质心转移 Fig. 8 Centroid transfer in potential suitable areas of alpine grassland dominant species and alpine meadow dominant species under different climate |
| 物种名称 Species name |
气候情景 Climate scenario |
时期 Period |
经度 Longitude |
纬度 Latitude |
迁移距离/km Migration distance |
| 高寒草原优势种 Alpine grassland dominan |
1970—2000 | 94°58′E | 33°08′N | — | |
| SSP126 | 2041—2060 | 93°52′E | 32°35′N | 119.55 | |
| 2061—2080 | 93°46′E | 32°47′N | 118.59 | ||
| SSP245 | 2041—2060 | 94°04′E | 32°36′N | 102.55 | |
| 2061—2080 | 94°36′E | 32°28′N | 81.23 | ||
| SSP370 | 2041—2060 | 93°40′E | 32°37′N | 133.88 | |
| 2061—2080 | 94°04′E | 32°32′N | 106.64 | ||
| SSP585 | 2041—2060 | 94°22′E | 32°41′N | 74.19 | |
| 2061—2080 | 94°18′E | 32°27′N | 96.58 | ||
| 1970—2000 | 97°43′E | 33°40′N | — | ||
| 高寒草甸优势种 Alpine meadow dominant species |
SSP126 | 2041—2060 | 97°18′E | 33°02′N | 80.17 |
| 2061—2080 | 97°19′E | 32°20′N | 152.83 | ||
| SSP245 | 2041—2060 | 97°29′E | 33°02′N | 73.92 | |
| 2061—2080 | 96°53′E | 32°49′N | 120.89 | ||
| SSP370 | 2041—2060 | 97°13′E | 33°28′N | 50.52 | |
| 2061—2080 | 97°33′E | 32°52′N | 90.00 | ||
| SSP585 | 2041—2060 | 97°14′E | 33°10′N | 70.19 | |
| 2061—2080 | 97°36′E | 32°46′N | 100.22 |
高寒草原与高寒草甸优势种的潜在适宜区质心都是向西南方向迁移。SSP126气候情景下高寒草原优势种的潜在适宜区质心向西南方向迁移再向西北方向迁移, 偏移距离最长, 高寒草甸优势种的潜在适宜区质心向南迁移, 偏移距离最长, 达152.83 km;SSP245气候情景下高寒草原优势种的潜在适宜区质心向西南方向迁移再向东迁移, 高寒草甸优势种的潜在适宜区质心向南迁移再向西迁移;SSP370气候情景下高寒草原优势种的潜在适宜区质心向西南方向迁移再向东迁移, 向西南方向迁移, 高寒草甸优势种的潜在适宜区质心向西南方向迁移再向东迁移, 较当前向南迁移, 偏移距离最短;SSP585气候情景下高寒草原优势种的潜在适宜区质心向西南方向迁移, 偏移距离最短, 高寒草甸优势种的潜在适宜区质心较当前向南迁移。
4 讨论 4.1 MaxEnt模型评估四类优势种的AUC值均大于0.9表明模型预测效果很好, 可用于高寒草地优势物种适宜区的模拟预测。以往的研究多专注于对单一物种如红松乔木、长袍铁杉等乔木或珍稀物种适宜区的模拟[46—47], 本研究从草地优势种的分布上推到草地类型空间变化, 探究草地类型适生区的演变趋势, 在定量化研究上又向前迈进了一步。通过分布点的选取和无效点的剔除, 保证了建模结果的准确性和可靠性, 利用MaxEnt和ArcGIS工具, 将4类草地优势种适宜区及空间变化清晰地叠加起来, 既便于观察各物种分布变化趋势, 又便于各物种潜在适宜区的对比。建立高寒草原与高寒草甸优势种分布模型使我们能够了解这些变化趋势, 以便当地政府和居民能够采取有效的措施来适应草地资源的分布变化。然而MaxEnt只需基于现有数据就能建模, 部分数据可能跨越较长的时间, 缺乏对生物相互作用、遗传变异和物种扩散能力等实际条件的考虑, 更容易导致物种分布区域的预测范围偏大[48—49]。因此, 后续的研究可以进一步完善模型, 并进行更多的实地验证, 以获得更全面的物种细节, 从而更好地进行保护和评估。
4.2 影响青藏高原优势种的主导因子水热条件是限制陆生植物分布的最主要因素[44]。青藏高原作为“世界屋脊”, 因海拔高、气候寒冷而形成独特的高寒生态系统[33]。高寒草原通常以密丛型多年生禾草建群种, 草层低且覆盖度小;高寒草甸通常以多年生草本植物建群种, 草层低矮却具有较高的覆盖度[50]。两类草地类型均具有耐寒适寒的特性, 而高寒草原适应的气候为寒冷半干旱地带[51], 易受季节性降雨影响, 能适应更高海拔环境[50];高寒草甸生境昼夜温差大, 光照强烈, 适应的气候地带为寒冷半湿润, 主要受气温影响[6], 但其对降水量的需求较高寒草原更高[50]。早熟禾类高寒草原与紫花针茅类高寒草原均具有较强的耐寒性和耐旱性, 生长于寒冷半干旱的大陆性气候地带, 然而早熟禾类高寒草原抗热性较差, 紫花针茅类高寒草原易受降水量影响, 在降水量低于100 mm的区域生长会受到限制[13, 31, 52];高山嵩草类高寒草甸与矮生嵩草类高寒草甸的生物学特性相似, 皆为喜湿冷的气候环境, 嵩草类高寒草甸因具备草毡层保温特性缓解了青藏高原高山低温的胁迫[53—54]。
根据MaxEnt预测结果显示早熟禾类高寒草原的的主要驱动机制是最冷季度平均温度(bio11), 紫花针茅类高寒草原, 高山嵩草类高寒草甸与矮生嵩草类高寒草甸是年降水量(bio12), 这可能是各草地优势种对气候响应受其生物学特性限制。环境变量响应曲线及适生区间显示年降水量在250—550 mm是紫花针茅类高寒草原与高山嵩草类、矮生嵩草类高寒草甸的适宜生长范围, 紫花针茅类最适宜年降水量为400 mm, 这与王艳芬等对高寒草地与环境变化的研究相符[13], 受年降水量影响3类优势种多分布于青藏高原中部、东北部及西南边界地区;而早熟禾类高寒草原的适宜生长范围为550—2500 mm, 年降水量达1000 mm的横断山区域能成为其生长的最佳地[55]。紫花针茅类高寒草原与高山嵩草类、矮生嵩草类高寒草甸海拔高度的适宜区间都>3800 m, 而早熟禾类高寒草原适宜区间是250—3600 m, 表明4类优势种都能适应高海拔环境, 但是早熟禾类高寒草原的适应能力更强, 分布范围更广, 这也与以往对早熟禾产地生境研究相符合[31]。平均温度日较差(bio2)和降水量季节性变化(bio15)贡献率结果显示4类优势种对温度的响应同样敏感, 研究表明平均温度日较差(bio2)在13.5 ℃—15 ℃和降水量季节性变化(bio15)在80至100是紫花针茅类高寒草原与高山嵩草类、矮生嵩草类高寒草甸的适宜生长范围, 高寒草原早熟禾类适宜范围是平均温度日较差为7—12 ℃且降水量季节性变化 < 90, 表明紫花针茅类高寒草原与高山嵩草类、矮生嵩草类高寒草甸更能适应青藏高原地区气温及降雨的变化。此外本研究发现未来情景下各优势种主导环境变量驱动空间分布权重将增加, 这也是气候变化改变高寒草地优势种空间分布的本质因素, 然而本研究仅分析了气候和海拔对高寒草地优势种分布的影响, 未考虑物种分布对种间关系、人类干扰等生物因子和遗传变异、土壤环境等非生物因子[54], 后续应进一步深入研究。
4.3 青藏高原优势种地理分布对未来气候变化的响应高寒草地是青藏高原适应高寒环境最为典型草地植物群落类型。近40年来气候因素主导高寒草甸分布呈比例增加, 受湿热条件增长驱动高寒草原向高寒草甸转变, 反之高寒草甸植物群落结构随温度与降水量降低愈简并逐渐转变为高寒草原[13], 优势种对气候因子响应曲线及适宜区面积变化也很好的解释了高寒草地类型的变化趋势(图 4, 图 8)。本研究预测了高寒草原优势种与高寒草甸优势种在未来不同气候情景下的潜在适宜区变化, 结果显示早熟禾类高寒草原在青藏高原东部地区最高丧失率为13.42%, 在SSP245和SSP370情景下会向南部扩展, 这可能是未来气温升高迫使其适宜区向西部迁移以应对高温胁迫, 同时未来南部降水量增加将为早熟禾类高寒草原提供新的适生地[56]。紫花针茅类高寒草原未来向西及东南部扩张, 且高寒草原优势种质心总体向西南方向偏移, 符合胡忠俊等对针茅属未来格局分布研究[20];未来高山嵩草类、矮生嵩草类高寒草甸的潜在适宜区向西扩张且最高扩张率为29.51%, 同胡忠俊等与张雪梅等对嵩草属未来会形成更大的分布格局且适宜区向西南方向延伸的研究相吻合[18, 57]。本研究发现未来高寒草原优势种与高寒草甸优势种的潜在适宜区质心总体向西南方向偏移, 而早熟禾类高寒草原东部适宜区丧失, 紫花针茅类高寒草原与矮生嵩草类高寒草甸适宜区向东部扩张, 这表明未来早熟禾类高寒草原将可能转化为紫花针茅类高寒草原和矮生嵩草类高寒草甸适宜区(图 1, 图 8, 表 4), 与王艳芬等对高寒草地近40年变化趋势的研究相似[13]。此外近40年高寒草地优势种空间分布占比增加至92%, 未来青藏高原暖湿化趋势增加其竞争优势, 从而影响群落结构并推动草地类型的变化[4, 13—14, 44], 本研究认为未来紫花针茅类高寒草原在青藏高原的优势地位会进一步提高, 部分高寒草原会转化为高寒草甸。气候变化影响高寒草地优势种空间分布, 在很大程度上还影响人类活动和经济发展[57], 因此本研究预测结果草地类型的变化为合理规划管理牧业活动提供了理论指导。
5 结论本研究利用MaxEnt预测了未来不同情景下高寒草原优势种和高寒草甸优势种在青藏高原的潜在适宜区分布。结果表明:(1)最冷季度平均温度(Bio11)是早熟禾类高寒草原的主导环境因子并驱动其空间分布于青藏高原东南部;年降水量(bio12)是紫花针茅类高寒草原, 高山嵩草类高寒草甸及矮生嵩草类高寒草甸的主导环境变量, 影响其适宜区分布于青藏高原东北部、中部及西南部。海拔(elev)、平均温度日较差(bio2)及降水量季节性变化(bio15)是影响4类高寒草地优势种的重要因子。(2)随着气候变化, 早熟禾类、紫花针茅类高寒草原与高山嵩草类、矮生嵩草类高寒草甸均能适应未来低、中排放的情景, 并沿青海湖及喜马拉雅山向南北两侧扩张, 然而在高排放(SSP585)情景下依然会出现生境退缩的情况, 青藏高原东部及中部是适宜区丧失的集中地区, 喜马拉雅山及青海湖周边适宜区将丧失。(3)未来高寒草原优势种适宜区与质心向西南方向迁移, 高山嵩草类与矮生嵩草类高寒草甸向南迁移, 青海省及西藏等地都可能成为高寒草地优势种的潜在适宜区, 且喜马拉雅山南部适宜区显著增加, 早熟禾类高寒草原丧失的部分区域会转化为紫花针茅类高寒草原或矮生嵩草类高寒草甸适宜区。本研究通过探求影响高寒草地分布的主导气候因子, 模拟并预测其潜在适宜区分布及空间变化, 对当地畜牧业的可持续发展、保护青藏高原生物多样性和维持高寒生态系统平衡都具有积极的参考价值。
致谢: 感谢中国科学院地理科学与资源研究所樊江文研究员和中国科学院西北高原生物研究所李英年研究员对研究和写作给予的帮助。| [1] |
甘小玲, 常亚鹏, 江原, 曹丰丰, 赵传燕, 李伟斌. 气候变化对祁连山蒙古扁桃潜在适生区的影响. 生态学报, 2023, 43(2): 768-776. DOI:10.5846/stxb202111103160 |
| [2] |
ipcc Intergovernmental Panel on Climate Change. Climate Change 2022-Impacts, Adaptation and Vulnerability. Cambridge, UK: Cambridge University Press, 2023.
|
| [3] |
羿明璇, 孙政国, 丛鹏飞, 安志远, 刘冲, 翟雪弟, 马焘, 高继超, 张东明. 草地生态系统功能对气候变化的响应研究进展. 中国草地学报, 2023, 45(12): 112-120. |
| [4] |
王譞. 全球气候变化对天然草地植物种群空间分布格局和种间关系的影响[D]. 呼和浩特: 内蒙古大学, 2018.
|
| [5] |
鲍艳, 魏宇晨, 南素兰, 臧文川. 全球2℃温升背景下青藏高原植被对气候变化的响应. 高原气象, 2023, 42(1): 49-59. |
| [6] |
杨元合. 全球变化背景下的高寒生态过程. 植物生态学报, 2018, 42(1): 1-5. |
| [7] |
董世魁, 王学霞, 张勇, 汤琳. 青藏高原高寒草地植物-土壤系统的生物多样性及其对全球变化的响应. 北京: 科学出版社, 2018.
|
| [8] |
Wang Y F, Lv W W, Xue K, Wang S P, Zhang L R, Hu R H, Zeng H, Xu X L, Li Y M, Jiang L L, Hao Y B, Du J Q, Sun J P, Dorji T, Piao S L, Wang C H, Luo C Y, Zhang Z H, Chang X F, Zhang M M, Hu Y G, Wu T H, Wang J Z, Li B W, Liu P P, Zhou Y, A. W, Dong S K, Zhang X Z, Gao Q Z, Zhou H K, Shen M G, Wilkes A, Miehe G, Zhao X Q, Niu H S. Grassland changes and adaptive management on the Qinghai-Tibetan Plateau. Nature Reviews Earth & Environment, 2022, 3(10): 668-683. |
| [9] |
焦珂伟, 高江波, 吴绍洪, 侯文娟. 植被活动对气候变化的响应过程研究进展. 生态学报, 2018, 38(6): 2229-2238. DOI:10.5846/stxb201702240305 |
| [10] |
王根绪, 胡宏昌, 王一博, 陈琳. 青藏高原多年冻土区典型高寒草地生物量对气候变化的响应. 冰川冻土, 2007, 29(5): 671-679. DOI:10.3969/j.issn.1000-0240.2007.05.001 |
| [11] |
马宁. 近40年来青藏高原典型高寒草原和湿地蒸散发变化的对比分析. 地球科学进展, 2021, 36(8): 836-848. |
| [12] |
王婧, 刘雄洲, 金冠芳, 斯确多吉, 次仁曲宗, 尹淑霞, 杨智永. 季节性不对称模拟增温对青藏高原高寒草甸群落特征的影响. 草地学报, 2022, 30(11): 3056-3062. |
| [13] |
Wang Y F, Xue K, Hu R H, Ding B Y, Zeng H, Li R J, Xu B, Pang Z, Song X N, Li C J, Du J Q, Yang X C, Zhang Z L, Hao Y B, Cui X Y, Guo K, Gao Q Z, Zhang Y J, Zhu J T, Sun J, Li Y M, Jiang L L, Zhou H K, Luo C Y, Zhang Z H, Gao Q B, Chen S L, Ji B M, Xu X L, Chen H, Li Q, Zhao L, Xu S X, Liu Y L, Hu L Y, Wu J S, Yang Q E, Dong S K, He J S, Zhao X Q, Wang S P, Piao S L, Yu G R, Fu B J. Vegetation structural shift tells environmental changes on the Tibetan Plateau over 40 years. Science Bulletin, 2023, 68(17): 1928-1937. DOI:10.1016/j.scib.2023.07.035 |
| [14] |
白乌云, 侯向阳. 气候变化对草地植物优势种的影响研究进展. 中国草地学报, 2021, 43(4): 107-114. |
| [15] |
Ma P F, Zhao J X, Zhang H Z, Zhang L, Luo T X. Increased precipitation leads to earlier green-up and later senescence in Tibetan alpine grassland regardless of warming. The Science of the Total Environment, 2023, 871: 162000. DOI:10.1016/j.scitotenv.2023.162000 |
| [16] |
耿远月, 徐田伟, 王循刚, 张骞, 张晓玲, 康生萍, 胡林勇, 刘宏金, 赵娜, 罗崇亮, 李英年, 徐世晓. 黄河源园区典型高寒草原牧草营养特征及家畜承载力研究. 草地学报, 2022, 30(7): 1829-1835. |
| [17] |
王婷, 史倩倩, 祁得胜, 陈克龙, 杜岩功. 青海湖地区紫花针茅型高寒草原群落特征变化. 草地学报, 2024, 32(4): 1204-1209. |
| [18] |
Hu Z J, Guo K, Jin S L, Pan H H. The influence of climatic changes on distribution pattern of six typical Kobresia species in Tibetan Plateau based on MaxEnt model and geographic information system. Theoretical and Applied Climatology, 2019, 135(1): 375-390. |
| [19] |
刘咏梅, 董幸枝, 龙永清, 朱志梅, 王雷, 盖星华, 赵樊, 李京忠. 退化高寒草甸狼毒群落分类特征及其环境影响因子. 草业学报, 2022, 31(4): 1-11. |
| [20] |
胡忠俊, 张镱锂, 于海彬. 基于MaxEnt模型和GIS的青藏高原紫花针茅分布格局模拟. 应用生态学报, 2015, 26(2): 505-511. |
| [21] |
Zhao W X, Liu T, Sun M M, Wang H Y, Liu X L, Su P. Rapid monitoring of Ambrosia artemisiifolia in semi-arid regions based on ecological convergence and phylogenetic relationships. Frontiers in Ecology and Evolution, 2022, 10(10): 3389. |
| [22] |
Huang X T, Ma L, Chen C B, Zhou H K, Yao B Q, Ma Z. Predicting the suitable geographical distribution of Sinadoxa corydalifolia under different climate change scenarios in the three-river region using the MaxEnt model. Plants, 2020, 9(8): 1015. DOI:10.3390/plants9081015 |
| [23] |
侯柏华, 张润杰. 基于CLIMEX的桔小实蝇在中国适生区的预测. 生态学报, 2005, 25(7): 1569-1574. DOI:10.3321/j.issn:1000-0933.2005.07.006 |
| [24] |
刘璐璐, 赵亮, 蔺诗颖, 冯建龙. 基于MaxEnt和GARP的阿蒙森海域南极磷虾(EUPHAUSIA SUPERBA)的分布区预测. 海洋与湖沼, 2023, 54(2): 399-411. |
| [25] |
Morales N S, Fernández I C. Land-cover classification using MaxEnt: can we trust in model quality metrics for estimating classification accuracy?. Entropy, 2020, 22(3): 342. DOI:10.3390/e22030342 |
| [26] |
施晓灯. 红花玉兰适生区域研究[D]. 北京: 北京林业大学, 2021.
|
| [27] |
陈禹光, 乐新贵, 陈宇涵, 程武学, 杜金贵, 钟全林, 程栋梁. 基于MaxEnt模型预测气候变化下杉木在中国的潜在地理分布. 应用生态学报, 2022, 33(5): 1207-1214. |
| [28] |
周礼茂, 吴海洋, 田斌. 基于MaxEnt优化模型对不同时期金银忍冬潜在适生区动态模拟分析. 西南林业大学学报(自然科学), 2024, 44(3): 43-51. |
| [29] |
朱仁斌. 中国特有植物文冠果(Xanthoceras sorbifolium bunge)的谱系地理研究与应用[D]. 杨凌: 西北农林科技大学, 2016.
|
| [30] |
周兴民. 中国嵩草草甸. 北京: 科学出版社, 2001.
|
| [31] |
中国植物志编委. 中国植物志. 科学出版社, 2002.
|
| [32] |
冯月. 增温与施氮对高寒沼泽草甸植物生理生化特性的影响[D]. 兰州: 兰州交通大学, 2021.
|
| [33] |
陶自为. 青藏高原人工草地群落结构对生态系统功能的影响研究[D]. 兰州: 兰州大学, 2022.
|
| [34] |
鲍根生. 甘肃马先蒿寄生对紫花针茅内生真菌共生体生长和光合特性的影响[D]. 兰州: 兰州大学, 2015.
|
| [35] |
Alexander P, Katherine C, Shinichiro F, Petr H, Florian H, Elke S, Leon B B, Philipp D J, C. D J, Mykola G, Tomoko H, Page K, Michael O, Andrzej T, Kiyoshi T, Hugo V, Stephanie W, Isabelle W, Marshall W, Elmar K, Hermann L C, Oliver F, Keywan R, van V D P. Land-use futures in the shared socio-economic pathways. Global Environmental Change, 2016, 42: 331-345. |
| [36] |
Wu T W, Lu Y X, Fang Y J, Xin X G, Li L R, Li W P, Jie W, Zhang J, Liu Y, Zhang L, Zhang F, Zhang Y, Wu F, Li J, Chu M, Wang Z, Shi X, Liu X, Wei M, Huang A, Zhang Y, Liu X. The Beijing Climate Center Climate System Model (BCC-CSM): the main progress from CMIP5 to CMIP6. Geoscientific Model Development, 2019, 12(4): 1573-1600. DOI:10.5194/gmd-12-1573-2019 |
| [37] |
周天军, 邹立维, 陈晓龙. 第六次国际耦合模式比较计划(CMIP6)评述. 气候变化研究进展, 2019, 15(5): 445-456. |
| [38] |
Kottarathu K R, Youngwook K, Taoufik K. Incorporating satellite remote sensing for improving potential habitat simulation of Prosopis cineraria (L.) Druce in United Arab Emirates. Global Ecology and Conservation, 2022, 37: e02167. DOI:10.1016/j.gecco.2022.e02167 |
| [39] |
唐杨欣, 皮杰, 刘新华, 向建国, 曾聪, 李德亮. 基于最大熵模型预测气候变化下河蚬在中国的潜在分布. 生态学报, 2023, 43(10): 4250-4259. DOI:10.5846/stxb202204110952 |
| [40] |
Phillips S J, Dudík M. Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography, 2008, 31(2): 161-175. DOI:10.1111/j.0906-7590.2008.5203.x |
| [41] |
宋慧芳, 党晓宏, 高永, 蒙仲举, 孙艳丽. 基于MaxEnt模型的内蒙古自治区樟子松潜在分布研究. 四川农业大学学报, 2023, 41(2): 203-208, 248. |
| [42] |
沈国强. 适应气候变化的农作物分布格局研究——以东北地区为例[D]. 哈尔滨: 中国科学院大学(中国科学院东北地理与农业生态研究所), 2017.
|
| [43] |
宦智群, 徐小蓉, 朱冬梅, 耿兴敏, 唐明, 圣倩倩, 祝遵凌. 应用最大熵模型预测不同气候变化情景下西康玉兰潜在地理分布. 东北林业大学学报, 2023, 51(5): 98-105. DOI:10.3969/j.issn.1000-5382.2023.05.016 |
| [44] |
Shi X D, Wang J W, Zhang L, Chen S X, Zhao A L, Ning X D, Fan G R, Wu N S, Zhang L, Wang Z D. Prediction of the potentially suitable areas of Litsea cubeba in China based on future climate change using the optimized MaxEnt model. Ecological Indicators, 2023, 148: 110093. DOI:10.1016/j.ecolind.2023.110093 |
| [45] |
1:100万中国草地资源图编制委员会. 1:100万中国草地资源图集. 北京: 中国地图出版社, 1993.
|
| [46] |
邱华, 舒皓, 吴兆飞, 李明鲁, 殷正, 张春雨, 赵秀海. 长白山阔叶红松林乔木幼苗组成及多度格局的影响因素. 生态学报, 2020, 40(6): 2049-2056. DOI:10.5846/stxb201904020642 |
| [47] |
谭雪, 张林, 张爱平, 王毅, 黄丹, 伍小刚, 孙晓铭, 熊勤犁, 潘开文. 孑遗植物长苞铁杉(Tsuga longibracteata)分布格局对未来气候变化的响应. 生态学报, 2018, 38(24): 8934-8945. DOI:10.5846/stxb201806021236 |
| [48] |
Abla B, Susana P, Nard B, Brahim C, David S F. Towards the identification of hotspots of freshwater biodiversity in North-Western Africa: a case study using species distribution models for water beetles in Morocco. Global Ecology and Conservation, 2023, 43: e02441. DOI:10.1016/j.gecco.2023.e02441 |
| [49] |
Barry S, Elith J. Error and uncertainty in habitat models. Journal of Applied Ecology, 2006, 43(3): 413-423. DOI:10.1111/j.1365-2664.2006.01136.x |
| [50] |
王金亭. 青藏高原高山植被的初步研究. 植物生态学与地植物学学报, 1988, 12(2): 3-12. |
| [51] |
梁大林, 唐海萍. 青藏高原两种高寒草地植被变化及其水温驱动因素分析. 生态学报, 2022, 42(1): 287-300. DOI:10.5846/stxb202012033093 |
| [52] |
哈雪, 张金青, 宋于笑, 马旭东, 马晖玲, 张然. 5份草地早熟禾种子产量性状相关指标综合评价与分析. 中国草地学报, 2023, 45(12): 61-71. |
| [53] |
戴黎聪, 郭小伟, 张法伟, 柯浔, 曹莹芳, 李以康, 李茜, 林丽, 朋措吉, 舒锴, 曹广民, 杜岩功. 青藏高原两种植被类型净初级生产力与气候因素的关系及周转值分析. 生态学报, 2019, 39(14): 5241-5251. DOI:10.5846/stxb201808301852 |
| [54] |
李积兰, 李希来, 马福俊. 黄河上游高寒草甸矮生嵩草生理特征对流域不同生境的响应. 草业学报, 2018, 27(1): 62-72. |
| [55] |
周思儒, 信忠保. 基于多源数据的近40年青藏高原降水量时空变化研究. 长江科学院院报, 2023, 40(10): 186-194. |
| [56] |
Ma L, Zhang Z H, Yao B Q, Ma Z, Huang X T, Zhou B R, Xu M H, Guo J, Zhou H K. Effects of drought and heat on the productivity and photosynthetic characteristics of alpine meadow plants on the Qinghai-Tibetan Plateau. Journal of Mountain Science, 2021, 18(8): 2079-2093. |
| [57] |
张雪梅, 刘涛, 任世杰, 刘思齐, 林长存, 纪宝明. 基于MaxEnt模型的3种野生牧草西藏地区潜在适生区分析. 草地学报, 2023, 31(8): 2314-2322. |