 2024, Vol. 44
2024, Vol. 44文章信息
- 韩爽, 张华兵, 刘玉卿, 蔡青, 王娟, 王成
- HAN Shuang, ZHANG Huabing, LIU Yuqing, CAI Qing, WANG Juan, WANG Cheng
- 盐城滨海湿地芦苇群落和互花米草群落对碱蓬群落挤压的时空变化特征
- Temporal and spatial characteristics of extrusion of Phragmites australis community and Spartina alterniflora community on Suaeda salsa community in Yancheng coastal wetland
- 生态学报. 2024, 44(22): 10309-10321
- Acta Ecologica Sinica. 2024, 44(22): 10309-10321
- http://dx.doi.org/10.20103/j.stxb.202302210290
-
文章历史
- 收稿日期: 2023-02-21
- 网络出版日期: 2024-08-22





2. 南京大学电子科学与工程学院, 南京 210023;
3. 安徽农业大学经济管理学院, 合肥 230036
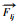 )呈线性增加, 并以1992年和2002年为节点分为3个阶段; 作用点轨迹以向东为主, 移动了约4700 m。互花米草群落对碱蓬群落的挤压力(
)呈线性增加, 并以1992年和2002年为节点分为3个阶段; 作用点轨迹以向东为主, 移动了约4700 m。互花米草群落对碱蓬群落的挤压力( 先增加后降低, 同样表现出3阶段特征; 轨迹点向西移动了约700 m, 向北移动了约270 m。芦苇群落和互花米草群落对碱蓬群落综合挤压力(F合)呈线性增加, 作用力方向2002年之前, 在NNE向的左侧, 2002年之后, 在NE向的右侧; 轨迹点向东北方向移动, 以向东移动为主。(3)RDA排序能够反映碱蓬群落面积与、、F合的关系, 12个时期的碱蓬群落面积变量被F合按照"三二一四"顺时针分布在4个象限; 被变量、按照"三四一二"的顺序分布在4个象限, 阶段性特征明显。(4)多元回归结果显示, 对碱蓬群落面积的影响总体上大于; 景观格局指数ED、LSI、SHDI、AI与、、F合表现出较高的相关性。研究结果可为盐城滨海湿地生态恢复和世界自然遗产地的可持续建设提供参考。
先增加后降低, 同样表现出3阶段特征; 轨迹点向西移动了约700 m, 向北移动了约270 m。芦苇群落和互花米草群落对碱蓬群落综合挤压力(F合)呈线性增加, 作用力方向2002年之前, 在NNE向的左侧, 2002年之后, 在NE向的右侧; 轨迹点向东北方向移动, 以向东移动为主。(3)RDA排序能够反映碱蓬群落面积与、、F合的关系, 12个时期的碱蓬群落面积变量被F合按照"三二一四"顺时针分布在4个象限; 被变量、按照"三四一二"的顺序分布在4个象限, 阶段性特征明显。(4)多元回归结果显示, 对碱蓬群落面积的影响总体上大于; 景观格局指数ED、LSI、SHDI、AI与、、F合表现出较高的相关性。研究结果可为盐城滨海湿地生态恢复和世界自然遗产地的可持续建设提供参考。2. School of Electronic Science and Engineering, Nanjing University, Nanjing 210023, China;
3. School of Economics and Management, Anhui Agricultural University, Hefei 230036, China
) increased linearly and was divided into three stages with 1992 and 2002 as nodes. The track of the action point was mainly eastward, moving about 4700 m. The extrusion force of Spartina alterniflora community on Suaeda salsa community () increased at first and then decreased, which also showed the characteristics of three stages. The track point moved about 700 m to the west and 270 m to the north. The combined extrusion force of Phragmites australis community and Spartina alterniflora community on Suaeda salsa community (F合) showed a linear increase. The force direction was on the left side of NNE direction before 2002, and on the right side of NE direction after 2002. The track point moved to the northeast, mainly to the east. (3) RDA ranking can reflect the relationship among Suaeda salsa community area and , and F合. The area variable of Suaeda salsa in 12 periods was distributed in four quadrants clockwise according to "three- two-one-four" by F合 and distributed in four quadrants in the order of "three-four-one-two" by , and , with obvious stage characteristics. (4) The results of multiple regression showed that the influence of on Suaeda salsa was larger than that of . There was the high correlation among landscape pattern indexes (ED, LSI, SHDI and AI) and , and F合. Research on the interaction relationship of plant communities in Yancheng coastal wetland can provide theoretical reference for the ecological restoration of coastal wetland and the sustainable construction of the world natural heritage site.滨海湿地位于海陆过渡地带, 在自然和人类活动双重作用下, 也是一个脆弱边缘地带和生态敏感区, 景观结构、过程和功能变化显著[1]。滨海湿地生长有多种水生、沼生和耐盐植物群落, 这些植被既是滨海湿地生态系统的重要组成部分, 也是表征生态系统健康状况的重要指标[2], 对滨海湿地生态系统相关功能的发挥起着关键作用[3]。湿地植物在盐土上自然完成其生活史, 与盐土协同进化, 形成了独特的适应盐生环境的生理机制[4]。植物与植物之间的相互作用主导着植物群落的构建, 不同植物也呈现不同的响应形式和机制, 一直以来都是植物生态学研究的核心内容[5]。
盐城滨海湿地是西太平洋海岸最大的淤泥质潮滩湿地, 是黄(渤)海候鸟栖息地(第一期)世界自然遗产地的主体。盐城滨海湿地环境梯度明显、物种组成相对简单, 生长在其中的盐沼植被常表现为带状分布的特征[6], 其中芦苇(Phragmites australis)、碱蓬(Suaeda salsa)和互花米草(Spartina alterniflora)为3种最典型的盐沼植被, 在盐碱地改良、固碳、为鸟类提供栖息地与觅食等方面发挥重要作用[7]。植物种群空间格局变化是其在群落中地位与生存能力的外在表现, 反映了植物对空间资源的利用状况及与其他种群的关系[8—9]。目前关于滨海湿地植被研究聚焦在两个方面:一是植物群落的生理适应及对环境的影响, 不仅阐述了滨海湿地植物性状和功能特征, 还研究了滨海湿地植物地理分布与环境因子的关系[10]以及对变化环境的响应特征[11];二是植被演替特征、机理及生态效应, 研究重点已从景观格局变化向生态环境对景观格局变化的响应方向转变, 既有微观尺度上植被变化对碳、氮组分的影响研究[12], 也有宏观尺度上植被变化对生态环境、生物多样性的影响[13—15];还有研究植被变化对水文地貌过程的影响以及对人类健康的影响[16—18]。目前, 滨海湿地植物群落之间相互作用的研究有运用图像质心分析和叠加分析方法, 分析不同植被相对空间位置的变化[19—20]。客观上, 芦苇群落与碱蓬群落、互花米草群落与碱蓬群落以及芦苇群落与互花米草群落之间都存在不同程度的相互竞争[9];尤其是碱蓬群落在芦苇群落和互花米草群落的双向挤压下, 不断被压缩, 纯碱蓬群落面积更少, 零星分布的碱蓬对鸟类生存优化意义已不明显[21]。鉴于此, 本文以盐城国家级自然保护区核心区为研究对象, 以1983—2021年12期遥感影像为数据源, 引入物理学库仑定律, 通过力学分析, 阐述芦苇群落对碱蓬群落、互花米草群落对碱蓬群落的作用力以及二者合力的时空变化, 研究有助于科学认识滨海湿地植物群落的演变态势, 为盐城国家级自然保护区生态修复提供理论参考。
1 研究区概况江苏盐城国家级珍禽自然保护区介于32°20′N—34°37′N, 119°29′E—121°16′E, 位于江苏省中部沿海地区, 东临黄海, 海岸线长约582 km, 保护区面积为2.47×105 hm2, 是亚洲大陆面积最大的淤泥质滨海湿地。江苏盐城国家级珍禽自然保护区核心区(图 1), 北至新洋港, 南至斗龙港, 为典型的淤长型潮滩湿地, 总面积约1.92×104 hm2。核心区以中路港道路为界, 分为南北两部分, 北部进行了大面积的人工芦苇沼泽恢复, 植被的自然演替被破坏, 为典型的人工管理区;南部主要受潮汐等自然条件影响, 人类活动影响较小, 植被自然演替特征明显, 为典型的自然条件控制区。本研究选择南部自然条件控制区为案例, 地表覆被从陆向海依次为芦苇群落、碱蓬群落、互花米草群落和光滩。芦苇群落主要由芦苇、三菱藨草(Bolboschoenoplectus mariqueter)、白茅(Imperata cylindric)等优势种群组成, 芦苇为建群种。碱蓬群落主要由盐地碱蓬、碱蓬(Suaeda glauca)、碱菀(Tripolium pannonicum)、柽柳(Tamarix chinensis)等种群组成, 在与芦苇群落的交错带有零星三菱藨草、白茅, 与互花米草群落交错带有零星互花米草, 盐地碱蓬、碱蓬为群落的优势种, 又以盐地碱蓬占据主导。互花米草群落几乎由单一种群组成, 即互花米草。

|
| 图 1 研究区位置 Fig. 1 The location of the study area |
数据源为12期遥感影像, 分别为1983年的MSS影像, 1988年、1992年、1997年、2000年、2002年、2006年、2009年、2011年、2014年、2017年和2021年的TM遥感影像。2003年以后由于卫星机载扫描仪, 致使数据条带丢失, 影响图像质量, 图像需要在ENVI中做去条带处理。然后, 对图像依次进行大气校正、几何校正。在ENVI 5.0中, 采用非监督分类和决策树分类的方法进行遥感图像分类, 进一步结合野外调查数据, 在ArcGIS中对遥感图像解译结果进行校正;使得遥感图像整体解译精度大于90%;最后, 在ArcGIS 10.0中, 完成滨海湿地12期植被类型图的制作。
2.2 库仑定律库仑定律是由法国物理学家库仑提出, 是电磁学的第一个基本定律, 用来量度真空中两个静止点电荷之间的相互作用力, 同名电荷相斥、异名电荷相吸, 作用力在两个电荷的连接线上;作用力大小与电荷量与电荷之间距离相关[22]。本研究从生态学的视角, 尝试用库仑定律描述植物群落间相互作用力的大小和方向。在此研究中, 将不同群落在生存空间上存在重叠且相互排斥的现象, 理解为同种电荷的相互作用;如果在空间上能够共存的, 理解为异种电荷相互作用。研究区内3种典型植物群落在空间上存在重叠且相互排斥明显, 可以看作是同种电荷相互作用;作用力方向在两个群落等效作用点的连线上。
2.2.1 等效作用点对任意力场中的刚体, 如果力学系统满足一定条件, 可以把各质点所受场力的作用点移到质心(力的大小和方向不变), 而刚体的运动状态不变, 质心所受场的合力就等于原来各质点所受场力的主矢, 质心也就是刚体所受场力的等效作用点[23]。如图 2所示, 将a、b群落斑块等同于物理学中的刚体, Pa、Pb分别为a、b群落斑块所受作用力的等效作用点。

|
| 图 2 群落相互作用示意图 Fig. 2 Schematic sketch of community interaction a:群落斑块;b:群落斑块;Pa:a群落斑块所受作用力的等效作用点;Pb:b群落斑块所受作用力的等效作用点;rab:群落斑块质心间的距离;ra:a群落至相互作用点的空间位置;rb:b群落至相互作用点的空间位置 |
库仑定律指出真空中两个静止点电荷之间的相互作用力大小与电荷的乘积成正比, 与两电荷之间的距离平方成反比[22]。库仑定律在此研究中应用主要表现为:第一, 盐城滨海湿地植物群落在生存空间上存在重叠, 且在空间上重叠越多, 群落之间竞争作用越明显, 不同群落在同一生态位空间上是相互排斥的;第二, 植物群落规模越大, 不同群落在空间上争夺越明显, 相互之间作用力越强;第三, 滨海湿地不同群落之间的距离越近, 越容易产生竞争。因此, 研究中将群落斑块的质心等同于点电荷的位置, 将群落规模等同于电荷量, 将两个群落斑块质心连接线与群落边界的交点等同于群落相互作用的直接空间位置, 具体公式如下:
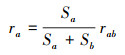
|
(1) |
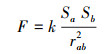
|
(2) |
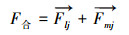
|
(3) |
式中, Sa、Sb为群落a、群落b的面积(hm2), rab为群落斑块质心间的距离(m), ra用来表示群落至相互作用点的空间位置(m);F为不同群落间的作用力, k为不同类型群落相互作用的调节系数, 本文中k取10, 使F计算结果在0—10之间;
以
以碱蓬面积(S)为因变量(y), 以
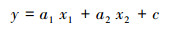
|
(4) |
在景观尺度上选择边缘密度(ED)、景观形状指数(LSI)、景观多样性指数(SHDI)、聚集度指数(AI)等4个景观格局指数, 通过线性回归系数来表征景观格局与芦苇、互花米草对碱蓬挤压力的响应关系。ED表示景观斑块与相邻异质斑块之间的边缘长度, 可以反映景观要素之间相互作用的强度和广泛性。LSI用来表征在挤压力作用下斑块形状的复杂程度。SHDI反映了各类型斑块的非均衡分布情况, 用来表征不同时期景观的异质性特征。AI用来表征在挤压力作用下, 斑块内部的连接程度或者破碎程度。
3 结果分析 3.1 植被结构变化特征盐城国家级珍禽自然保护区核心区植被结构以芦苇群落、碱蓬群落和互花米草群落为主(图 3)。芦苇群落面积从1983年的6.266 hm2扩张至2017年的4178.669 hm2, 至2021年又略有下降, 降至3372.756 hm2, 总体上随着时间呈线性扩张趋势比较明显, 相关系数为0.940(图 4)。互花米草群落面积从1983年的268.761 hm2增加至2021年的3592.081 hm2, 和芦苇群落面积发展的趋势基本一致, 与时间呈线性扩张趋势, 相关系数为0.949(图 4)。

|
| 图 3 1983—2021年研究区植物群落分布图 Fig. 3 Distribution of plant communities in the study area from 1983 to 2021 |

|
| 图 4 研究区植物群落面积变化的线性特征 Fig. 4 Linear characteristics of plant community changes in the study area |
在1983—2002年, 虽然在向陆地一侧, 碱蓬群落受到芦苇群落向海扩张的影响, 但是碱蓬群落在向海一侧还未与互花米草群落相接, 仍能保持一定的速度增长, 碱蓬群落面积从1983年的3301.716 hm2扩张至2002年的4588.773 hm2, 在时间上保持一定的线性增长特征, 相关系数为0.908(图 4)。但是, 2002—2021年, 碱蓬群落受到明显的双向挤压, 向陆一侧受到芦苇群落的挤压, 在向海一侧已与互花米草群落相接, 开始受到互花米草的挤压, 致使碱蓬群落面积从2002年的4588.773 hm2快速降至2017年的578.253 hm2, 后由于保护区开展生境修复等措施, 至2021年碱蓬群落面积又升至1035.099 hm2;总体上2002年以来研究区碱蓬群落面积在时间上呈线性萎缩, 相关系数为0.917(图 4)。
3.2 芦苇群落和互花米草群落对碱蓬群落的挤压作用 3.2.1 芦苇群落对碱蓬群落作用力(

|
|
图 5 研究区植物群落相互作用力变化
Fig. 5 Change of plant community interaction in the study area
:芦苇群落对碱蓬群落作用力;:互花米草群落对碱蓬群落作用力 |
根据等效作用点的坐标位置和公式(1)可以分别计算出不同时间芦苇群落、互花米草群落对碱蓬群落挤压的作用点。从芦苇群落对碱蓬群落挤压的作用点空间轨迹看(图 6), 以1988年和2002年为时间拐点的阶段性特征非常明显, 呈现“东南→西北→东”的空间变化格局。1983—1988年, 作用点向东南方向运动。1988—2002年, 作用点总体上呈现先向东南后向西北方向运动的特征, 其中1997—2000年向东南方向有个折返。2002—2021年, 作用点总体上向东运动, 其中2009—2014年, 作用点出现了波动, 先向东北又折回向西南方向;2014—2017年, 作用点大幅度向东北方向运动, 以向东为主;至2021年, 作用点又折向西南, 以向南方向为主。1983—2021年, 向东累积移动了约4700 m, 在南北方向上累积变化约为0。

|
| 图 6 芦苇群落与碱蓬群落作用点变化 Fig. 6 Changes in action point of Phragmites australis on Suaeda salsa community |
从互花米草群落对碱蓬群落挤压的作用点空间轨迹看(图 7), 空间变化要比芦苇群落对碱蓬群落挤压的作用点轨迹复杂, 以2011年为时间拐点, 在空间上以向“东南—西北”“西南—东北”两组方向变化为主, 经历了“西北→东南→西北→西南→东北→东南”的过程。1983—1988年, 作用点向西北方向运动。1988—1997年, 作用点向东南方向移动。1997—2011年, 作用点总体上向西北方向移动, 其中2000—2002年有一个向东的转向。2002—2011年, 作用点总体上以向西北方向移动, 其中2006—2009年有一个向东南方向的折返。2011—2014年, 作用点开始向西南方向移动;2017年, 转向东北方向;至2021年, 又折向东南方向。空间作用点在东西方向的移动范围大于南北方向的移动距离, 向西累积移动了约700 m, 向北累积移动了约270 m。

|
| 图 7 互花米草群落与碱蓬群落作用点变化 Fig. 7 Changes in action point of Spartina alterniflora on Suaeda salsa community |
进一步比较
从时间上看, 1983—2021年, F合大小呈递增趋势, 与时间的线性相关系数为0.863(图 8)。具体看, 1983年F合最小, 为0.527;2006年F合最大, 为8.185;1983年和1992年, F合小于1;1988年和1997年, F合在1—2之间;2002年, F合为3.959;2006—2021年的F合在5—9之间。总体上可以看出, F合以2002年为时间节点, 2002年以前的F合主要在0—2之间, 2002年以后的F合大部分在5—7之间(2006年除外)。如果不考虑2006年的情况, 其他年份的F合与时间的线性相关系数可达到0.942。

|
| 图 8 芦苇群落和互花米草群落对碱蓬群落综合作用力变化 Fig. 8 Changes of comprehensive force of Phragmites australis and Spartina alterniflora on Suaeda salsa community |
通过碱蓬群落质心变化可以看出综合作用点的轨迹, 1983—2021年, 综合作用点以向北、向东移动为主导方向, 东西移动最大距离为2737.338 m, 南北移动距离最大达到了1424.418 m, 以东西移动占据主导。从时间轨迹看(图 9), 综合作用点移动分为3个阶段, 从轨迹点的分布看也表现出3个集群特征:1983—1992年为第一阶段, 这阶段综合作用比较小, 轨迹变化小;1997—2002年为第二阶段, 轨迹点东移, 综合作用开始增加, 轨迹变化幅度比第一阶段大;2006—2021年为第三阶段, 轨迹点向东北方向移动, 综合作用更加复杂, 出现“西北—东南”往复摆动。进一步结合植物群落分布图, 第一阶段芦苇群落、互花米草群落的规模小, 碱蓬群落处于显著的优势地位;第二阶段, 芦苇群落、互花米草群落的规模扩大, 但是碱蓬群落仍能有空间保持一定速度向海洋方向增长;第三阶段, 碱蓬群落已无扩展空间, 其空间分布主要受芦苇群落、互花米草群落的支配。

|
| 图 9 芦苇群落和互花米草群落对碱蓬群落综合作用点变化 Fig. 9 Change of action point of comprehensive force of Phragmites australis and Spartina alterniflora on Suaeda salsa community |
力是矢量, 既有大小又有方向。1983—2021年, F合的方向主要为北偏西和北偏东, 并且北偏东占据主导(图 10)。F合的方向在时间上具有明显的阶段特征, 2002年之前, F合的方向都在NNE向的左侧, 2002年之后, F合的方向几乎都在NE向的右侧。

|
| 图 10 芦苇群落和互花米草群落对碱蓬群落综合作用力的方向变化 Fig. 10 Direction changes of the comprehensive force of Phragmites australis and Spartina alterniflora on Suaeda salsa community N:北方向;NNE:北北东方向;NE:北东向;ENE:东北东方向;E:东方向;ESE:东南东方向;SE:南东向;SSE:南南东方向;S:南方向;SSW:南南西方向;SW:南西向;WSW:西南西方向;W:西方向;WNW:西北西方向;NW:北西向;NNW为北北西方向 |
运用冗余分析(RDA)方法, 分析碱蓬群落面积对F合的响应关系, 结果显示, 第一主轴和第二主轴的特征值分别为0.433和0.567, 碱蓬群落面积与第一主轴、第二主轴的累积解释率分别为43.3%和100%;碱蓬群落面积与第一主轴的范式典型相关性为0.658, 解释拟合变量率为100%。RDA排序能够反映碱蓬群落面积与F合的响应关系。从图 11中可以看出, 12个时期的碱蓬群落面积变量被分布在4个不同象限, 2006年、2009年分布在第一象限;1997年、2000年、2002年分布在第二象限;1983年、1988年、1992年分布在第三象限;2011年、2014年、2017年和2021年分布在第四象限;1983—2002年分布在纵坐标轴的左侧, 2006—2021年分布在纵坐标轴的右侧。从时空角度看, 坐标系中碱蓬群落呈“三二一四”的顺时针分布;各象限内的要素分布, 既与F合的阶段特征相吻合, 也反映了碱蓬群落面积的时间变化。

|
| 图 11 碱蓬群落面积与挤压力的冗余分析 Fig. 11 Redundancy analysis (RDA) between Suaeda salsa community area and extrusion force F合为芦苇群落和互花米草群落对碱蓬群落综合挤压作用力 |
进一步, 以
运用多元回归方法分析碱蓬群落面积(y)与
| 模型 Model |
相关系数 Multiple regression(R) |
R2 | 修正后R2 Adjust R2 |
显著性检验 Significance test |
F检验 F-test |
| 1 | 0.690a | 0.476 | 0.423 | 0.013a | 9.071 |
| 2 | 0.809b | 0.654 | 0.577 | 0.008b | 8.503 |
| a为预测变量(常量):x1;b为预测变量:x1, x2 | |||||
从表 2中可以看出, 碱蓬群落面积(y)与
| 模型 Model |
非标准化系数 Unstandardized coefficient |
标准系数 Standard coefficient |
T检验 T-test |
显著性检验 Significance test |
||
| 非标准化回归系数 Unstandardized regression coefficient |
标准误差 Standard error |
|||||
| 1 | 常量 | 3840.675 | 467.075 | 8.223 | 0.000 | |
| x1 | -254.273 | 84.427 | -0.690 | -3.012 | 0.013 | |
| 2 | 常量 | 3351.791 | 459.927 | 7.288 | 0.000 | |
| x1 | -384.192 | 94.168 | -1.042 | -4.080 | 0.003 | |
| x2 | 302.209 | 140.351 | 0.550 | 2.153 | 0.060 | |
芦苇群落和互花米草群落对碱蓬群落挤压作用与景观格局变化密切相关, 从表 3中可以看出:
| 相关系数 Correlation coefficient |
ED | LSI | SHDI | AI | |
|
F合 |
|
0.782** | 0.781** | 0.902** | -0.752** | 1.000 | 0.641* | 0.949** |
|
0.435 | 0.433 | 0.833** | -0.386 | 0.641* | 1.000 | 0.670* |
| F合 | 0.720** | 0.720** | 0.882** | -0.687* | 0.949** | 0.670* | 1.000 |
| **在0.01水平上显著相关;*在0.05水平上显著相关;ED:边缘密度Edge density;LSI:景观形状指数Landscape shape index;SHDI:景观多样性指数Shannon′s diversity index;AI:聚集度指数Aggregation index;:芦苇群落对碱蓬群落作用力The force of Phragmites australis on Suaeda salsa community;:互花米草群落对碱蓬群落作用力The force of Spartina alterniflora on Suaeda salsa community;F合:芦苇群落和互花米草群落对碱蓬群落综合挤压作用力The comprehensive force of Phragmites australis and Spartina alterniflora on Suaeda salsa community | |||||||
本研究以盐城国家级珍禽自然保护区为案例区, 运用物理学方法, 研究芦苇群落与互花米草群落对碱蓬群落挤压的时空变化特征, 结果如下:
1983—2021年, 研究区植被变化表现出芦苇群落、互花米草群落面积线性增加, 碱蓬群落面积先增加后减少的特征。
植物群落间的相互作用是滨海湿地生物多样性的重要源泉, 也是维持滨海湿地生态系统多样性的基础。群落间相互作用的时空定量测度有助于科学认识滨海湿地生态系统结构和功能变化。本研究采用的物理学库仑定律测度滨海湿地植物群落种间竞争力, 这一方法简单、直观、可操作性强, 通过植物群落面积和空间分布位置, 用矢量数据描述不同植物群落间的竞争力大小和作用方向, 并判断相互作用的位置, 能够分析不同植物群落相互作用在空间上的变化, 在方向的判断上是基于对区域实际情况有较为统一认知的基础上。相比较其他方法, 库仑定律的应用有其优势也有其局限性:第一, 相比较竞争系数, 库仑定律不需要实测生物量, 但用面积表示植物群落竞争力未能考虑生物量以及处于竞争状态下的不同植物对资源的利用效率;第二, 相比较生态过程模型, 库仑定律是一个静态模型, 是通过竞争的结果来判断竞争力强弱, 不需要对生态过程进行检测, 但是忽略了生境过程要素对植物群落的影响以及植物与生境的相互作用。因此, 今后需要进一步从“格局—过程”的耦合机制来测度群落间相互作用, 从而为滨海湿地生态系统修复和多样性保护提供科学依据。
4.2 讨论 4.2.1 盐城滨海湿地植物群落作用机制生态习性, 强调植物对环境的适应能力。盐城滨海湿地水文地貌过程是海陆两相共同作用的结果, 水文地貌过程是滨海湿地植被演变的基础。盐城滨海湿地不同植物群落的生态习性是生态位产生分异和重叠的基础。芦苇是我国滨海湿地本土的多年生草本植物和优势种, 在维护湿地生态系统的结构、功能和生态系统服务供给方面发挥重要作用[24], 芦苇群落一般位于地下水埋藏较浅的中高潮地带, 距海相对较远, 潮汐入侵频率小于5%, 受淡水影响大, 耐盐能力较强, 耐盐能力在5%—40%, 在长期淹水条件下, 各项生长指标都表现出最高值[25];有研究表明, 芦苇群落面积与土壤含盐量呈负相关关系, 而遗传多样性则与生境中的盐度具有正相关关系[26]。互花米草群落是滨海湿地与海洋最直接作用的植被群落, 生长于平均高潮位与小潮高潮位之间, 潮侵频率在50%—80%, 表现出比芦苇群落更强的耐淹、耐盐能力, 在盐度胁迫下能够保持生理功能稳定。碱蓬是盐城滨海湿地土著物种, 具有较强耐盐能力, 生长于大潮高潮位与平均高潮位之间, 潮侵频率在20%—50%。不同空间地下水位及相应的水盐性状是影响湿地植物群落分布格局、物种多样性和植物生物量的重要因素[27—29]。因此, 盐城滨海湿地3种植物群落从陆向海呈现出“芦苇群落—碱蓬群落—互花米草群落”的带状空间分布格局;植物群落的生态习性的差异, 致使3种群落在滨海湿地环境的适应与竞争能力呈现明显的差异。另一方面, 海岸侵蚀影响了互花米草群落向海洋方向的扩张, 致使2006年以后互花米草群落扩张以向陆地方向为主。
芦苇、盐地碱蓬、互花米草分别是芦苇群落、碱蓬群落和互花米草群落的建群种, 3种植物的生活型、繁殖方式都影响着植物群落之间的相互作用。一般来说, 个体越大、密度越大的植物, 竞争力越强;植物多样性的繁殖方式往往会提升植物对多变环境的适应能力, 从而提升植物种间竞争力。芦苇和互花米草都为多年生草本植物, 植株高大, 高度都可达到3—4 m, 地上部分生物量远高于碱蓬。芦苇和互花米草均为克隆植物, 前者属于游击型结构, 后者倾向于密集型或混合型结构;均具有有性繁殖和无性繁殖的能力;芦苇对碱蓬挤压主要是通过粗大的根茎进行拓展而占据生境;互花米草则通过无性繁殖提高植株数量和植株密度, 通过有性繁殖的方式占据新的生境[30]。碱蓬是1年生草本真盐生植物, 植株低矮、稀疏, 高度在20—80 cm, 以种子繁殖为主, 生物量远低于芦苇和互花米草, 具有独特的耐盐机制。
4.2.2 芦苇群落和互花米草群落对碱蓬群落的挤压过程碱蓬是滨海湿地“先锋植物”, 特别是地上部分聚集土壤盐分, 从土壤中吸收盐分并聚集在体内, 从而转移土壤盐分, 促进土壤脱盐, 而土壤含盐量逐步降低后有利于其他中轻度耐盐植物生长[21, 31—33], 盐分过高或过低都不利于碱蓬生长。因此, 碱蓬群落的上缘, 随着土壤盐分降低, 将更适合芦苇群落生长。互花米草种群对外部环境变化的适应能力较强, 通过促淤功能影响碱蓬的生境使其不适宜继续生长, 进而实现其扩张侵占目的。有研究表明碱蓬群落中互花米草主要通过实生苗入侵新生境, 并向外扩张, 通过互花米草分蘖苗维持种群的稳定, 而且碱蓬滩斑块处的含水量、盐度适合互花米草的生存、扩张[31]。在盐城滨海湿地, 芦苇与互花米草都具有较宽的生态幅与较强潮间带适应能力, 它们的竞争力均强于本地种碱蓬[34, 35], 研究区碱蓬群落中不仅存在大量的芦苇群落与互花米草群落斑块, 甚至出现了芦苇群落与互花米草群落接壤, 也佐证了碱蓬的竞争力较弱[9]。
在互花米草入侵之前, 盐城滨海湿地的潮滩高程剖面是从陆向海逐渐降低的, 潮水可以从海洋向陆推进, 形成稳定的滨海湿地原生生境, 从海洋向陆地形成了“光滩—碱蓬群落—芦苇群落”的分布格局, 形成了“光滩→碱蓬群落→芦苇群落”的单向演替序列。随着互花米草入侵并发育成宽数千米的条带状, 改变了滨海湿地水文地貌格局。由于互花米草群落的超强淤积能力, 互花米花群落的海拔显著增加, 滨海湿地的高程剖面呈“U”形[36]。互花米草群落超强的促淤能力, 使得生境朝着不利于碱蓬群落发育的方向发展[37];同时, 互花米草群落快速扩张并占据了本地物种碱蓬的生存空间, 滨海湿地由碱蓬群落单一格局演变为多层次湿地植物群落分布特征, 由陆向海依次分布芦苇群落、碱蓬群落及互花米草群落;也改变了滨海湿地原有的植被演替序列, 形成了“光滩→互花米草群落”“碱蓬群落→互花米草群落”“碱蓬群落→芦苇群落”的3个演替序列。互花米草群落的扩张改变了滨海湿地的水沙动力环境, 对地貌的重塑产生了重要影响[38—39], 而海拔变化反过来又降低了水沙侵蚀对植被的影响, 有利于植被的更好发展;这种双向动态反馈机制是滨海湿地景观演变的重要环节。
总之, 芦苇群落和互花米草群落的双向挤压, 特别是互花米草入侵改变了本地种间竞争格局, 致使碱蓬群落面临消失的危险[9], 影响湿地功能和生态价值[40]。滨海湿地植被分布格局的变化, 也直接影响滨海湿地生物多样性、能量流动、物质循环等生态特征与过程, 影响动植物种群乃至微生物群落的空间分布[41—42]。因此, 针对滨海湿地复杂的水文地貌过程以及植被快速演替的特征, 需要借助遥感、GIS等空间分析手段, 建立长时序的滨海湿地植被空间格局数据, 定期评估滨海湿地生态系统稳定性和生境适宜性[43], 建立基于自然的生态修复方案(Nbs), 充分发挥滨海湿地栖息地功能, 对盐城世界自然遗产地的可持续建设具有重要意义。
| [1] |
张华兵, 王娟, 刘玉卿, 韩爽, 张亚楠. 人类活动与互花米草扩张对滨海湿地生境质量演变的影响——以盐城国家级珍禽自然保护区核心区为例. 长江流域资源与环境, 2021, 30(5): 1153-1163. |
| [2] |
Fu B L, Wang Y Q, Campbell A, Li Y, Zhang B, Yin S B, Xing Z F, Jin X M. Comparison of object-based and pixel-based Random Forest algorithm for wetland vegetation mapping using high spatial resolution GF-1 and SAR data. Ecological Indicators, 2017, 73: 105-117. DOI:10.1016/j.ecolind.2016.09.029 |
| [3] |
郑云云, 胡泓, 邵志芳. 典型滨海湿地植被演替研究进展. 湿地科学与管理, 2013, 9(4): 56-60. |
| [4] |
郭嘉, 李敬玉, 徐化凌, 许洁, 张明乾, 于一雷, 徐卫刚, 马牧源, 赵娜娜, 李肖夏, 朱耀军. 6种典型滨海湿地植物对土壤盐分的影响. 水生态学杂志, 2021, 42(3): 38-46. |
| [5] |
孔祥龙. 黄河三角洲植物群落种间相互作用研究[D]. 济南: 山东大学, 2016.
|
| [6] |
易雨君, 谢泓毅, 宋劼, 杨志峰. 黄河口盐沼湿地植被群落适宜生境模拟Ⅱ: 应用. 水利学报, 2021, 52(4): 401-408. |
| [7] |
王文硕. 典型盐生植被群落演替退化遥感监测研究[D]. 大连: 大连海洋大学, 2022.
|
| [8] |
赵成章, 姚强, 郝青, 董小刚. 东祁连山地次生林演替过程中种群格局动态. 山地学报, 2010, 28(2): 234-239. |
| [9] |
王娟, 张华兵, 刘玉卿, 韩爽, 徐雅, 张亚楠. 盐城滨海湿地植物群落种间格局与竞争的关系研究. 生态学报, 2020, 40(24): 8966-8973. |
| [10] |
刘利, 季长波, 张梅, 王鹏, 张彦文. 中国不同地区滨海湿地植物区系特点及其影响因素. 东北林业大学学报, 2022, 50(10): 44-48. |
| [11] |
江星浩, 谢柳娟, 叶思源, 周攀, 裴理鑫, 陈皓, 赵俐红. 江苏滨海湿地芦苇和互花米草光合特性对模拟增温的响应. 生态学报, 2022, 42(19): 7760-7772. |
| [12] |
谭立山. 土地利用/覆被变化对滨海湿地碳、氮组分的影响研究[D]. 上海: 华东师范大学, 2022.
|
| [13] |
么秀颖. 大丰麋鹿国家级自然保护区滨海湿地土壤碳储量时空变化[D]. 南京: 南京林业大学, 2022.
|
| [14] |
公慧珍, 李升峰. 江苏东台滩涂垦区植物群落演替及多样性梯度变化研究. 生态科学, 2015, 34(6): 16-21. |
| [15] |
刘庄, 沈永明, 曹学章. 互花米草生态环境影响评价研究——以江苏为例. 生态经济, 2008, 24(3): 149-151, 157. |
| [16] |
王爱军, 高抒, 贾建军. 互花米草对江苏潮滩沉积和地貌演化的影响. 海洋学报, 2006, 28(1): 92-99. |
| [17] |
冯虹毓. 互花米草入侵扰动下滨海湿地地表高程变化的研究[D]. 厦门: 厦门大学, 2020.
|
| [18] |
钦佩. 互花米草与人体健康关系研究进展. 中国野生植物资源, 2019, 38(5): 70-73. |
| [19] |
尹志恒, 甄艳, 杨晓钦, 吴宗攀, 康锦涛, 樊哲. 1997—2017年盐城国家级珍禽自然保护区景观格局时空演变分析. 测绘与空间地理信息, 2021, 44(12): 63-67, 71. |
| [20] |
张华兵. 盐城海滨湿地景观格局变化与生态过程响应. 北京: 科学出版社, 2018.
|
| [21] |
成海, 张亚楠, 陈亚芹, 李征浩, 李洪山. 盐地碱蓬的盐土适应机制及其在滩涂湿地上的群落构建实践. 现代园艺, 2020, 43(22): 179-180. |
| [22] |
张洪欣, 沈远茂, 韩宇南. 电磁场与电磁波 (2版). 北京: 清华大学出版社, 2016.
|
| [23] |
苏双臣, 刘凤祥, 侯石柱, 李文艳. 质心是合力等效作用点的条件研究. 河北工业大学学报, 2003, 32(1): 113-116. |
| [24] |
潘宇, 李德志, 袁月, 徐洁, 高锦瑾, 吕媛媛. 崇明东滩湿地芦苇和互花米草种群的分布格局及其与生境的相关性. 植物资源与环境学报, 2012, 21(4): 1-9. |
| [25] |
Vretare V, Weisner S E B, Strand J A, Granéli W. Phenotypic plasticity in Phragmites australis as a functional response to water depth. Aquatic Botany, 2001, 69(2/3/4): 127-145. |
| [26] |
张俪文, 王安东, 赵亚杰, 陈琳, 宋建彬, 杨长志, 韩广轩. 黄河三角洲滨海湿地芦苇遗传变异及其与生境盐度的关系. 生态学杂志, 2018, 37(8): 2362-2368. |
| [27] |
Fan Y, Li H, Miguez-Macho G. Global patterns of groundwater table depth. Science, 2013, 339(6122): 940-943. |
| [28] |
Xu X L, Zhang Q, Tan Z Q, Li Y L, Wang X L. Effects of water-table depth and soil moisture on plant biomass, diversity, and distribution at a seasonally flooded wetland of Poyang Lake, China. Chinese Geographical Science, 2015, 25(6): 739-756. |
| [29] |
李雅, 于秀波, 刘宇, 张广帅, 张全军, 段后浪. 湿地植物功能性状对水文过程的响应研究进展. 生态学杂志, 2018, 37(3): 952-959. |
| [30] |
阮俊潮, 戴文红, 李文兵, 宋垚彬, 董鸣. 滨海湿地优势植物芦苇和互花米草的生态响应与效应研究进展. 杭州师范大学学报: 自然科学版, 2019, 18(5): 490-498, 509. |
| [31] |
陈柯欣, 丛丕福, 曲丽梅, 韩建波, 韩成伟. 黄河三角洲互花米草、碱蓬种群变化及扩散模拟. 北京师范大学学报: 自然科学版, 2021, 57(1): 128-134. |
| [32] |
陈立华, 张欢, 姚宇阗, 张弛, 郑金海, 张风革. 盐地碱蓬覆被对滨海滩涂土壤理化性质的影响. 植物资源与环境学报, 2021, 30(2): 19-27. |
| [33] |
王文硕, 李微, 孙悦, 孙涛, 刘远. 辽东湾北部碱蓬群落及滩涂土壤盐分遥感反演模型. 华中师范大学学报: 自然科学版, 2022, 56(3): 532-540. |
| [34] |
徐伟伟, 王国祥, 刘金娥, 陈正勇, 杭子清, 王会. 苏北海滨湿地互花米草种群繁殖方式. 生态学报, 2014, 34(14): 3839-3847. |
| [35] |
陈正勇, 王国祥, 刘金娥, 徐伟伟, 王刚. 苏北滨海湿地互花米草两种繁殖体与本地种竞争能力分析. 海洋科学进展, 2012, 30(3): 380-389. |
| [36] |
侯明行, 刘红玉, 张华兵, 王聪, 谭清梅. 地形因子对盐城滨海湿地景观分布与演变的影响. 生态学报, 2013, 33(12): 3765-3773. |
| [37] |
袁红伟, 李守中, 郑怀舟, 方舟易. 外来种互花米草对中国海滨湿地生态系统的影响评价及对策. 海洋通报, 2009, 28(6): 122-128. |
| [38] |
Widdows J, Brinsley M D, Salkeld P N, Elliott M. Use of annular flumes to determine the influence of current velocity and bivalves on material flux at the sediment-water interface. Estuaries, 1998, 21(4): 552-559. |
| [39] |
Cahoon D R, Perez B C, Segura B D, Lynch J C. Elevation trends and shrink-swell response of wetland soils to flooding and drying. Estuarine, Coastal and Shelf Science, 2011, 91(4): 463-474. |
| [40] |
钱凤魁, 周阳, 李婉宁, 郝赫阳, 柳圭泽. 辽河口翅碱蓬湿地退化区土壤理化性质及生态阈值分析. 土壤通报, 2021, 52(5): 1085-1094. |
| [41] |
曹晨晨, 苏芳莉, 李海福, 魏超, 孙迪. 辽河口盐地碱蓬湿地景观破碎化及驱动机制. 生态学报, 2022, 42(2): 581-589. |
| [42] |
王钰祺, 任玉蓉, 廖安邦, 董倩, 刘茂松. 盐城滨海滩涂湿地典型植物群落土壤微生物组成与结构特征. 生态学报, 2023, 43(6): 2336-2347. |
| [43] |
徐力刚, 赖锡军, 万荣荣, 王晓龙, 李相虎. 湿地水文过程与植被响应研究进展与案例分析. 地理科学进展, 2019, 38(8): 1171-1181. |