 2024, Vol. 44
2024, Vol. 44文章信息
- 武晶晶, 王旭峰, 张松林, 李宗省
- WU Jingjing, WANG Xufeng, ZHANG Songlin, LI Zongxing
- 东亚地区植被光合日内变化特征——融合OCO-3和Himawari-8的分析
- Analysis of diurnal variation of vegetation photosynthesis in East Asia by fusing OCO-3 and Himawari-8
- 生态学报. 2024, 44(21): 9884-9895
- Acta Ecologica Sinica. 2024, 44(21): 9884-9895
- http://dx.doi.org/10.20103/j.stxb.202405101051
-
文章历史
- 收稿日期: 2024-05-10
- 网络出版日期: 2024-09-30





2. 中国科学院西北生态环境资源研究院, 黑河遥感试验研究站, 兰州 730000;
3. 中国科学院西北生态环境资源研究院, 祁连山同位素生态水文与国家公园观测站/内陆河流域生态水文重点实验室/甘肃省祁连山生态环境研究中心, 兰州 730000
2. Heihe Remote Sensing Experimental Research Station, Northwest Institute of Eco-Environmental Resources, Chinese Academy of Sciences, Lanzhou 730000, China;
3. Observation and Research Station of Eco-Hydrology and National Park by Stable Isotope Tracing in Qilian Mountains/Key Laboratory of Ecohydrology of Inland River Basin/Ecological Environment Research Center of Qilian Mountain, Gansu Province, Northwest Institute of Eco-environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, China
为了应对气候变化, 我国于2020年9月22日在第75届联合国大会上提出了2030年实现碳达峰、2060年实现碳中和的双碳目标。植被生态系统为巨大、持续的碳汇, 对实现“双碳”目标和减缓全球气候变暖具有重要意义[1—2]。植被光合作用是陆地生态系统中碳循环的关键过程之一[3]。目前, 许多学者已经基于一些植被指数(如NDVI、EVI)对植被光合作用变化进行了广泛的研究, 然而传统的植被指数在进行植被光合动态研究中可能会存在一些局限性[4—5]。随着遥感技术的发展, 日光诱导叶绿素荧光(Sun-Induced Chlorophyll Fluorescence, SIF)遥感成为研究植被光合作用的重要手段[6—8]。SIF是植物在太阳光照条件下, 由叶绿素分子的光合中心以荧光的形式发射出的光谱信号(650—800nm), 具有红光波段(690nm左右)和近红外波段(740nm左右)两个波峰[9—10], 能直接反映植物光合作用的动态变化, 现已成为研究植被光合作用的新工具和重要依据[11—13]。
东亚拥有多样的生态系统, 在全球碳循环中扮演着重要角色。因此, 研究该地区植被的光合作用变化, 不仅能揭示区域生态过程, 还能为全球气候变化模型提供重要数据支持。特别是通过监测日内的太阳诱导荧光(SIF), 能够深入了解植被的光合作用动态变化。首先, 日内SIF变化能够反映植物在一天中不同时间段内光合作用的活跃度和效率, 这对研究植物在不同环境条件下的适应性和生理响应至关重要[14]。其次, 这些数据能够揭示植物对环境胁迫的快速响应能力, 例如在高温、干旱或光照强度变化下, 植物如何调节光合活性以维持生长和生存[15—17]。此外, 日内SIF变化还有助于研究植被生态系统的能量收支平衡, 从而深入理解生态系统碳循环的过程和机制[16]。
近年来, 国内外学者通过卫星遥感技术得到了大量探测全球尺度SIF的产品, 如Zhang等[18]通过训练人工神经网络(ANN)与MODIS表面反射率和来自OCO-2卫星的SIF数据, 生成了2000年以来的全球连续日光诱导叶绿素荧光数据集(CSIF), 时间分辨率为4天, 空间分辨率为0.05°× 0.05°。Li和Xiao[9]利用MODIS EVI(增强型植被指数)和气象再分析MERRA-2(现代研究与应用回顾分析)、PAR(Photosynthetically Active Radiation, 光合有效辐射)、VPD(蒸汽压亏缺)和气温数据, 训练Cubist回归树模型, 生成了全球2000年以来的OCO-2 SIF数据集(GOSIF), 时间分辨率为8天, 空间分辨率为0.05°× 0.05°。佀等人[19]利用GOSIF数据, 发现2001—2021年黄河流域植被光合总体呈上升趋势, 在空间上表现为西部和东南部高、北部低的分布格局。Jeong等[20]利用SIF数据分析了北方高纬度森林植被物候和功能的大尺度变化, 整个生长季节响应整个北方生态系统的温度变化存在季节性滞后, 光照限制了生态系统的生产力, 植被光合作用受到了明显影响, 部分地区的植被光合作用强度呈现下降趋势。然而, 目前利用卫星观测的SIF研究主要集中在植被光合作用的季节变化或年际变化趋势, 对光合作用日内变化的研究仍然局限于小区域或多个站点的观测实验, 难以实现对大区域的观测和分析。传统的太阳同步卫星, 如OCO-2, 虽然提供了高空间分辨率的SIF数据, 但由于其固定的过境时间和轨道布局, 无法有效捕捉一天中不同时间段的植被光合作用变化[21—22]。为解决这一问题, 本研究选择搭载在国际空间站的OCO-3卫星。与OCO-2卫星不同, OCO-3能够提供更灵活的观测时间, 不受固定过境时间的限制, 使得可以更频繁地获取SIF数据。
因此, 本研究的目的是:(1)通过结合OCO-3和Himawari-8以及随机森林回归算法生成东亚地区时空连续的逐小时SIF数据集;(2)分析东亚地区植被SIF的日内变化特征, 探讨其与生长季光照和温度的关系;(3)比较不同植被类型间SIF日内变化的差异, 揭示其在光合作用响应性上的特征。最后为该区域应对气候变化和实现碳中和目标提供理论支撑。
1 研究区概况与数据来源 1.1 研究区概况东亚(10°—60°N, 80°—150°E)位于太平洋西岸, 包括中国、日本、韩国、朝鲜及蒙古五个国家, 占地面积约1250万km2(图 1), 人口数量占全世界人口的22%, 是全球人口最密集的区域之一。东亚地区地势西高东低, 呈阶梯状分布, 西部地形以高原、山地为主, 东部以平原、丘陵为主。该地区横跨热带、亚热带、温带及寒带, 总体上以温带季风气候和亚热带季风气候为主, 夏季炎热多雨、冬季温暖湿润, 且受东亚季风影响, 降雨的季节和年际变化很大[23]。东亚地区的植被生态系统丰富多样, 包括亚热带常绿阔叶林、温带针叶林和寒带草甸等;生长季高峰期在每年的7—9月, 此时的植被覆盖度达到一年内的最大值。

|
| 图 1 研究区地表植被类型 Fig. 1 Land cover vegetation types in the study area 中国藏南地区没有统计数据 |
美国国家航空航天局(National Aeronautics and Space Administration, NASA)的OCO-3(Orbiting Carbon Observatory-3)搭载在国际空间站ISS上, 自2019年5月发射后持续提供每天的日光诱导叶绿素荧光数据。该卫星的Footprint大小为1.3km×2.25km, 空间分辨率在目前为止可用的SIF遥感产品中最高, 观测范围在地球上南北纬约52°之间, 这样的高分辨率更有利于理解景观的格局和异质性对SIF的影响。
本研究中使用的数据集版本为OCO-3 v10r, 选择2019年8月和9月, 2020年至2022年每年的7月、8月和9月的观测数据, 共计覆盖了11个月。采用740nm波段反演的SIF数据(SIF740)作为模型训练的样本数据, OCO-3的数据是瞬时数据, 获取每个OCO-3 SIF Footprint中心位置的经纬度, 并将所有OCO-3 SIF聚合到每小时间隔的0.05°×0.05°网格单元, 单位:W m-2 μm-1 sr-1。OCO-3 SIF数据来源于NASA官网(https://ocov3.jpl.nasa.gov/science/oco-3-data-center/)。
1.2.2 反射率数据向日葵8号卫星(Himawari-8)是日本气象厅于2014年10月发射的静止气象卫星。该卫星搭载的扫描成像仪(Advanced Himawari Imager, AHI)共16个通道, 包含3个可见光通道、3个近红外通道和10个热红外通道, 可以实现区域(80°E—160°W, 60°S—60°N)内1次/10min的高频观测, 空间分辨率最高为500m。本研究选用Himawari-8的1—6波段作为模型的输入要素, 波段1—6的波长分别为470nm、510nm、640nm、860nm、1600nm和2300nm。该数据下载地址为:ftp://ftp.ptree.jaxa.jp/jma/netcdf/。
利用Himawari-8的1、3、4波段的反射率计算植被增强指数(Enhanced Vegetation Index, EVI)。EVI是监测植被状况最广泛采用的植被指数[24], 计算公式为:

|
(1) |
式中, ρNIR、ρRed与ρBlue分别表示近红外波段、红波段和蓝波段的反射率。
1.2.3 气象数据从气候资料库(https://www.ecmwf.int/en/forecasts/datasets/reanalysis-datasets)获取ERA5再分析数据。对ERA5-Land数据进行预处理, 使之符合模型的输入要求。输入模型的气象要素包括逐小时的2m平均空气温度(t2m)、2m露点温度(d2m)、光合有效辐射(PAR)、饱和水汽压差(VPD)以及相对湿度(RH), 其中t2m和d2m从ERA5-Land数据集直接获得, 单位:K。PAR是由ERA5-Land数据集中地面向下短波辐射(SSRD)乘以转化因子0.45得到, 单位:J/m2。SSRD为累计值, 每日0时(UTC)累计前一日的总量。由于ERA5-Land数据集中未提供相对湿度数据, 本文使用联合国粮农组织(FAO)提出的方法, 利用公式(2)计算当前气温下的饱和水汽压差(VPD), 并根据公式(3)计算各像元的相对湿度。
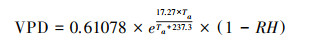
|
(2) |
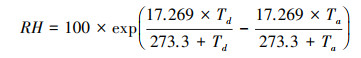
|
(3) |
式中, Td为露点温度, Ta为平均空气温度。
1.2.4 MODIS土地覆盖数据MODIS地表覆盖数据是利用监督决策树分类算法得到的全球土地覆盖类型数据。MODIS官网提供了3种不同空间分辨率(500m、1km和0.05°)的产品。为便于和OCO-3卫星数据匹配, 选择0.05°的产品, 即MCD12C1。以MCD12C1中IGBP的植被类型分类方案为基础, 对植被类型进行重分类, 并归并为7类[25]。其中, 常绿针叶林(ENF)和落叶针叶林(DNF)归并为针叶林(NF), 落叶阔叶林(DBF)和混交林(MF)归并为落叶阔叶林(DBF), 郁闭灌丛(CSH)和开阔灌丛(OSH)归并为灌木(SHR), 有林草地(WSA)和稀树草原(SAV)归并为草原(SAV), 农田(CRO)和自然植被混合地(CNV)归并为农田(CRO), 常绿阔叶林(EBF)和草地(GRA)不变。在NASA的数据中心下载(https://e4ftl01.cr.usgs.gov/)。
1.3 算法使用Python中的scikit-learn库, 实现随机森林回归算法(Random Forest Regression Algorithm, RFRA), 建立Himawari-8波段反射率与OCO-3 SIF之间的回归模型。本研究将东亚地区2019年(8月、9月)、2020年(7月、8月、9月)、2021年(7月、8月、9月)和2022年(7月、8月、9月)共11个月逐小时EVI、PAR、t2m、d2m、VPD和Himawari-8的1—6波段反射率数据进行逐像元对应, 建立空间连续的预测数据集, 剔除误差较大的数据后, 总样本数据为15万条;选取2019—2022年总样本数据的60%作为训练数据, 进行模型的构建;样本剩下的40%数据作为验证数据集, 用于验证模型的精度。模型训练通过粒子群优化算法优化随机森林回归算法的参数组合, 并对模型进行十折交叉验证, 得出模型的最佳预测精度和最小误差。得到随机森林算法预测的SIF后, 需要评估预测结果的准确性;将预测得到的SIF值与OCO-3 SIF观测值进行对比, 差异越小表明预测结果越好。选择决定系数(R2)、平均绝对误差(MAE)以及均方根误差(RMSE)这3个指标来评估模型的准确性。
R2可评估预测结果和观测结果(真实值)之间的符合程度[8], R2值越大表明SIF预测结果和OCO-3 SIF观测值的拟合程度越好[12]。计算公式如下:

|
(4) |
式中, N为数据数量;yi为观测值;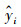
模型拟合的精度通过计算训练样本中预测值和真实值的平均绝对误差(Mean Absolute Error, MAE)来衡量, 计算公式见式(5)。MAE越小, 说明预测值与观测值的偏差越小, 预测精度越高。
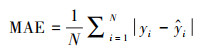
|
(5) |
均方根误差(RMSE)是预测值与真实值偏差的平方和观测次数N比值的平方根, 计算公式见式(6)。RMSE衡量了预测值与真实值的偏差大小, RMSE越小, 说明估计值与真实值的偏差越小, 预测的准确性越高。
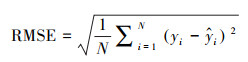
|
(6) |
通过粒子群优化算法确定决策树的个数n、最大深度(max_depth)、分隔节点的随机变量数量m和叶子节点的最小样本数(min_samples_leaf)。当n=500、max_depth=10、m =10和min_samples_leaf=5时, 回归模型的预测精度最高(图 2)。本研究构建RF的R2为0.585, RMSE为0.47W m-2 μm-1 sr-1, RMSE值较大, R2值较小, 可能是由于东亚地区样本的范围和数量较为局限, 导致预测过程累积的误差较大。RF回归模型能够评估每一个特征变量对模型的作用, 图 3统计了训练集中各个参数对SIF观测值的重要性分数, EVI、PAR、VPD、d2m、t2m与Himawari-8的1—6波段反射率分别为0.327、0.051、0.033、0.302、0.035、0.03、0.021、0.032、0.082、0.058、0.029。变量的重要性得分越高, 表明该变量对模型预测的贡献程度越大。可见, EVI是预测SIF的最重要的特征变量, 其次是d2m, 可能原因是d2m数据是逐小时的, 且日变化较大。

|
| 图 2 RF模型SIF预测值与真实值的验证, 图中的虚线表示1 : 1线 Fig. 2 Verification of RF model prediction SIF and observation SIF, the dashed line in the figure indicates a 1 : 1 line SIF:日光诱导叶绿素荧光Solar-induced chlorophyll fluorescence; OCO-3:轨道碳观测者三号Orbiting Carbon Observatory-3; R2:决定系数Coefficient of Determination; RMSE:均方根误差Root Mean Square Error |

|
| 图 3 特征变量重要性 Fig. 3 Importance of characteristic variables EVI:增强植被指数Enhanced Vegetation Index;t2m:2 m高度的平均空气温度2 m Air Temperature;d2m:2 m高度的露点温度2 m Dew Point Temperature;PAR:光合有效辐射Photosynthetically Active Radiation;VPD:饱和水汽压差Vapor Pressure Deficit;Band1—6:波段1至波段6的反射率Reflectance for Bands 1 to 6 |
采用在1.4中训练好的RF回归树模型, 基于Himawari-8静止卫星的1—6波段反射率数据、代表植被生长状况的EVI数据, 代表气候条件的t2m、PAR和VPD数据, 预测东亚地区逐小时0.05°分辨率的SIF。
图 4显示, RF模型预测的2020年至2022年7—9月期间, 东亚地区植被SIF平均值具备较高的空间分辨率和连续性, 适用于进一步的研究。东亚地区的SIF分布整体表现为东部和南部较高, 向西和向北逐渐降低。该地区的SIF分布受多种因素综合影响, 包括气候、地形和植被类型等因素。中国东部沿海和长江流域的SIF值相对较高, 主要受温暖湿润的气候条件和丰富的针叶林、阔叶林以及灌丛等植被的影响;而中国西部地区, 如青藏高原和西南山地, 由于海拔高度较高、气候相对干燥和植被覆盖较少, 其SIF值相对较低。日本、韩国和朝鲜的主要植被类型包括针叶林、阔叶林和草甸, 受温带季风气候和地形多样性影响, 这些地区在7—9月份的SIF值较高。然而, 由于日本、朝鲜和韩国多为多山地形, 它们的SIF分布呈现出较大的空间异质性。相比之下, 蒙古地形相对平坦、气候干冷, 其植被稀疏且主要为草原和荒漠, 因此整体SIF值较低。总体而言, 尽管东亚地区夏季植被的SIF值普遍较高, 但由于该地区地形、气候和植被类型的多样性, 其SIF值空间分布表现出复杂的空间异质性。

|
| 图 4 2020—2022年7—9月东亚地区植被SIF平均值的空间分布特征 Fig. 4 Spatial distribution characteristics of vegetation SIF average in East Asia from July to September, 2020—2022 |
图 5为2020—2022年7—9月东亚地区植被SIF值的空间分布图。由图 5可知, 7月东亚植被SIF高值区大多集中于朝鲜、韩国、日本及中国东北、东南部, 零散分布于中国中部地区及西南部地区, 少数分布于外蒙古国北部地区;SIF低值区分布于外蒙古国西北部及中部地区、中国华北地区及西部的青藏高原地区。8月与7月的空间分布基本一致, 植被SIF高值区集中分布在中国东北部、东部及南部地区、外蒙古国北部、朝鲜、韩国以及日本。到了9月份, 北方气候由热转寒, 植被随寒气增长逐渐萧落, 导致中国东北、东南沿海地区植被SIF值降低。

|
| 图 5 2020—2022年7—9月SIF空间分布特征 Fig. 5 Spatial distribution characteristics of SIF in 7—9, 2020—2022 |
图 6表明, 东亚地区7—9月植被的SIF值存在明显的单峰型日内变化。其中, 2020年7月的SIF值从8时(0.47W m-2 μm-1 sr-1)开始持续上升, 至中午11时达到高峰(0.71W m-2 μm-1 sr-1), 随后逐渐下降, 到16时达到最低值。8月的SIF值日内变化趋势与7月的类似, 峰值出现的时间也一致。这种SIF值日内变化趋势的一致性主要可以归因于2020年7月和8月植被指数(EVI)的高度相似性以及稳定的气象条件。具体来说, EVI作为反映植被绿度和生长活动的指标, 显示出了在这两个月内变化较小的特点, 这直接影响了植被SIF值变化趋势。此外, 光合有效辐射和气温等气象因素在这段时间内变化不大, 进一步促成了SIF日内变化特征的高度一致性。9月的SIF值日内变化的规律类似, 但是峰值较7月和8月减少了0.28W m-2 μm-1 sr-1, 可能是因为9月处于秋初, 日照时间逐渐减少, 因此植物的光合作用活动逐渐减弱。2021年7月SIF值的峰值比2020年7月的更高, 为0.77W m-2 μm-1 sr-1, 比8月的(0.68W m-2 μm-1 sr-1)与9月的峰值(0.49W m-2 μm-1 sr-1)更高, 可能是因为该年的太阳辐射日内变化特征有所差异等原因所致。但是, 该年9月植被的SIF峰值在三个年份中最高。2022年7—9月, 东亚地区植被SIF值的日变化曲线与2021年的类似。图 6表明, 东亚地区植被7—9月的SIF月均值存在年际差异。7月与9月的类似, 但是8月的明显不同, 显示年际间与月份间变化的差异性。

|
| 图 6 2020—2022年7—9月东亚地区植被SIF的日内变化特征与月均值的年变化特征 Fig. 6 Diurnal variation characteristics of vegetation SIF and annual variation characteristics of monthly mean in East Asia from July to September 2020—2022 |
以2020年7月8 : 00—16 : 00为例(图 7), 东亚地区植被的SIF值空间分布具有显著的空间异质性, 整体由西北部向东南部递增;清晨(8:00)很低, 随后逐渐增加, 到中午(11:00)达到峰值, 再逐渐下降, 与太阳辐射的日内变化趋势基本吻合。综上所述, 三年来, 东亚地区植被生长高峰期的SIF值日内变化趋势显示出了明显的单峰型特征、月际差异, 与光合作用活动随着季节变换和光照条件变化有关。

|
| 图 7 2020年7月8 : 00—16 : 00东亚地区植被SIF的空间分布特征 Fig. 7 Spatial distribution characteristics of vegetation SIF in East Asia from 8 : 00 to 16 : 00, July 2020 |
光合作用日变化曲线可直观反映植被的生长特性, 受气候条件、生长期与植被类型的影响较大, 在此仅探讨植被类型对SIF值日内变化的影响。图 8表明, 虽然东亚地区不同类型植被SIF值的日变化曲线均呈现单峰型, 但是不同植被类型之间的差异比较明显, 不同年份与月份之间也存在一定的差异性, 突显了植被类型、季节波动和其它环境变化在光合作用过程中的复杂相互作用。在2020年7月日内变化曲线中, 不同类型植被SIF的峰值从高到低依次为:EBF>CRO>DBF>SAV>NF>SHR>GRA, 其中EBF的峰值最大(1.381W m-2 μm-1 sr-1), 在11 : 00达到峰值;其次是CRO的(1.379W m-2 μm-1 sr-1), 在12 : 00达到峰值;GRA的最小(0.468W m-2 μm-1 sr-1), 在12 : 00达到峰值。8月SIF的峰值从高到低依次为:CRO>EBF>DBF>SAV>NF>SHR>GRA;9月的SIF值排序显著变化, EBF的SIF峰值最高, 峰值从高到低依次为:EBF>CRO>SAV>DBF>NF>SHR>GRA, 7种植被类型的SIF峰值均有所下降。

|
| 图 8 不同植被类型的SIF月均值日内变化 Fig. 8 Intraday variation in monthly mean SIF values for different vegetation types CRO:农田Cropland;EBF:常绿阔叶林Evergreen Broadleaf Forest;GRA:草地Grassland;SHR:灌木Shrubland;NF:针叶林Needleleaf Forest;SAV:草原Savanna;DBF:落叶阔叶林Deciduous Broadleaf Forest |
同样的, 2021—2022年7—9月东亚地区不同类型植被SIF值的日内变化类似:7月份EBF在光照充足时(11点)达到SIF峰值, 反映了其叶面积指数和叶绿素含量高的优势。SAV和CRO也在中午11点时达到SIF峰值, 峰值略低于EBF。与此同时, SHR显示出不同程度的光合作用活性下降, 但相对于DBF, 其下降趋势较为缓和。8月份植被的SIF值可能受到水分限制的影响而降低。相比之下, EBF维持相对稳定的日内变化, 其峰值出现在11点。CRO、SAV、DBF和NF的SIF峰值仍然保持在11点, 略低于7月份, 可能是由于温度升高(不同地区温度的月变化趋势有差异性)导致生长速率减缓。9月份, DBF、NF和CRO的光合作用活性明显下降, 因为叶片开始凋落;其峰值相对于7、8月份显著降低。SHR可能会因为其生长习性和对温度的敏感性不同, 而与DBF相比下降较为缓和。与此同时, EBF、GRA和SAV的光合作用活性继续保持在中午11点左右达到峰值, 但是峰值活性略低于7、8月份, 可能是由于温度下降和日照时间减少所致。
3 讨论 3.1 环境条件对SIF的影响在构建SIF模拟模型时, 本研究全面考虑了多种环境因子对太阳诱导叶绿素荧光(SIF)的影响。选取的关键驱动因素包括植被指数(EVI)、光合有效辐射(PAR)、饱和水汽压差(VPD)、2m露点温度(d2m)、2m平均空气温度(t2m)以及反射率数据。与以往研究相比, 本研究不仅涵盖了更多环境因子, 还对东亚地区的广泛气候、地形和生态系统类型进行了详细分析。前人研究指出, 温度、水分和光照等因素对植物光合作用具有关键性影响[26—27]。本文观察到东亚东部和南部地区的植被生产力较高, 与以往研究中亚热带和温带地区植被生产力较高的观点相符, 这些地区的温暖湿润气候条件有利于植被生长, 进而提高了植被的GPP(总初级生产力)和SIF的值[28—29]。聂冲等人[30]的研究进一步证实温度对长江流域植被生产力的变化起着主导作用, 这与本研究的结果一致。此外, Li和Xiao[9]的全球性研究表明, 不同地区的气候、土壤和植被类型对植被生产力和SIF值有显著影响, 从而解释了中国西部地区相对较低的植被SIF值。因此, 通过综合考虑这些关键环境因子, 能够更准确地分析和预测SIF在不同地理环境下的变化, 为相关研究提供重要的数据支持和科学依据。
3.2 SIF日内变化的控制因素本研究发现东亚地区植被的日内光合作用(SIF)呈现出明显的单峰型变化特征, 即在早上升高至中午达到峰值(11 : 00—12 : 00), 随后逐渐降低。这一日内变化模式与亚马逊雨林的研究结果相似, 表明光照和温度是主要影响因素[31]。尽管地理环境不同, 东亚地区与亚马逊雨林在植被光合作用的日内变化特征上存在一致性。光照条件(太阳辐射)直接影响植被的光合效率。温度对植被的生长和代谢活动具有重要影响, 适宜的温度有助于提高光合作用速率, 但过高的温度可能造成光合作用抑制, 例如光合午休现象[27]。湿度影响植被的水分吸收和蒸腾作用, 进而调节光合速率。不同类型的植被对环境因素的响应可能有所不同, 导致其光合速率及日内变化存在差异[32]。过去的研究已在叶片水平观察到植物光合作用的昼夜模式[33—34];地面SIF监测显示了午后SIF的滞后或抑制现象[7]。例如, Jingyu等[27]在华北平原的农田站点上观测到, 地面温度和光照强度显著影响冬小麦的SIF日变化, 与本研究发现的东亚地区植被“单峰型”日内变化特征相一致。综上所述, 环境因素如光照、温度、湿度以及植被类型在调节东亚地区植被SIF日内变化中起到关键作用。本研究通过深入分析这些因素的影响机制, 为理解和预测不同地理环境下植被光合作用的时空变化提供了重要的科学依据。
3.3 SIF产品的不确定性来源本研究对比了所得到的SIF数据与其它常见SIF产品(如GOSIF、CSIF)之间的差异性及其不确定性来源, 主要归结为以下两个方面:(1)生成方法的差异。本研究采用了随机森林回归算法生成SIF数据, 而其它SIF产品可能使用支持向量机、深度学习或其它算法。不同的算法在模型训练和参数调整上存在差异, 直接影响了SIF数据的生成和结果的不确定性。(2)数据源的差异。本研究基于OCO-3的SIF数据(波长为740nm), 而GOSIF产品基于OCO-2数据生成的SIF使用的是波长757nm。这两种数据在波长、空间分辨率和反演模型等方面存在显著差异。为了进行比较, 本研究使用了波长比例因子(1.69)将波长740nm转换为757nm[35], 并将时间分辨率聚合到月尺度。图 9表明, 2020—2022年7月本研究得到的SIF757nm月均值比GOSIF高出约0.05W m-2 μm-1 sr-1, 而在9月份, 两种产品的SIF月均值几乎一致。此外, 其他SIF遥感产品如CSIF, TROPOMI SIF, 在波段选择、空间分辨率、时间分辨率和反演模型等方面也各有特点[36—37]。不同卫星的观测精度、覆盖范围与传感器性能的差异直接影响了SIF数据的准确性和一致性。此外, 不同卫星数据的配准和校正也可能引入一定的不确定性。综上所述, 为了确保数据分析结果的准确性和可靠性, 需要全面考虑这些不确定性来源, 并进行适当的验证、校正和比较分析, 以便有效地利用不同SIF产品进行科学研究和应用。

|
| 图 9 不同遥感产品在东亚地区植被SIF月均值的比较 Fig. 9 Comparison of monthly vegetation SIF of different remote sensing products in East Asia SIF740:740nm波长处的日光诱导叶绿素荧光(SIF);SIF757:757nm波长处的日光诱导叶绿素荧光(SIF) |
(1) 东亚地区植被SIF值的空间异质性较高。2020—2022年7—9月生长季, 东亚地区植被SIF值呈现出明显的空间分布特征, 东部和南部地区相对较高, 向西和向北逐渐降低。这一差异与地理环境、气候条件及植被类型密切相关。
(2) 东亚地区植被SIF值呈现明显的单峰型特征。2020—2022年7—9月, 东亚地区植被的SIF值从早晨开始逐渐增加, 于11 : 00至12 : 00左右达到高峰, 随后逐渐减小。这与光照条件及植物光合作用活动密切相关。
(3) 植被类型影响东亚地区植被SIF值的日内变化特征。2020—2022年7—9月, 东亚地区7种植被类型的SIF日内变化曲线均为“单峰型”, 但差异明显。2020—2022年的7、8月, CRO和EBF的日均SIF值最高, GRA最低。而9月份EBF的光合作用活性最高, 显示出其光合作用活性存在着季节性变化的趋势。
| [1] |
Piao S L, Liu Q, Chen A P, Janssens I A, Fu Y S, Dai J H, Liu L L, Lian X, Shen M G, Zhu X L. Plant phenology and global climate change : current progresses and challenges. Global Change Biology, 2019, 25(6): 1922-1940. DOI:10.1111/gcb.14619 |
| [2] |
Zheng C, Wang S Q, Chen J M, Chen J H, Chen B, He X L, Li H, Sun L G. Combination of vegetation indices and SIF can better track phenological metrics and gross primary production. Journal of Geophysical Research : Biogeosciences, 2023, 128(7): e2022JG007315. DOI:10.1029/2022JG007315 |
| [3] |
Tang J W, Körner C, Muraoka H, Piao S L, Shen M G, Thackeray S J, Yang X. Emerging opportunities and challenges in phenology : a review. Ecosphere, 2016, 7(8): e01436. DOI:10.1002/ecs2.1436 |
| [4] |
夏传福, 李静, 柳钦火. 植被物候遥感监测研究进展. 遥感学报, 2013, 17(1): 1-16. |
| [5] |
Han J X, Zhang X Y, Wang J H, Zhai J Q. Geographic exploration of the driving forces of the NDVI spatial differentiation in the upper Yellow River Basin from 2000 to 2020. Sustainability, 2023, 15(3): 1922. DOI:10.3390/su15031922 |
| [6] |
Joiner J, Yoshida Y, Vasilkov A P, Schaefer K, Jung M, Guanter L, Zhang Y, Garrity S, Middleton E M, Huemmrich K F, Gu L, Belelli Marchesini L. The seasonal cycle of satellite chlorophyll fluorescence observations and its relationship to vegetation phenology and ecosystem atmosphere carbon exchange. Remote Sensing of Environment, 2014, 152: 375-391. DOI:10.1016/j.rse.2014.06.022 |
| [7] |
Mohammed G H, Colombo R, Middleton E M, Rascher U, van der Tol C, Nedbal L, Goulas Y, Pérez-Priego O, Damm A, Meroni M, Joiner J, Cogliati S, Verhoef W, Malenovský Z, Gastellu-Etchegorry J P, Miller J R, Guanter L, Moreno J, Moya I, Berry J A, Frankenberg C, Zarco-Tejada P J. Remote sensing of solar-induced chlorophyll fluorescence (SIF) in vegetation : 50 years of progress. Remote Sensing of Environment, 2019, 231: 111177. DOI:10.1016/j.rse.2019.04.030 |
| [8] |
Bai Y, Liang S L, Yuan W P. Estimating global gross primary production from Sun-induced chlorophyll fluorescence data and auxiliary information using machine learning methods. Remote Sensing, 2021, 13(5): 963. DOI:10.3390/rs13050963 |
| [9] |
Li X, Xiao J F. A global, 0.05-degree product of solar-induced chlorophyll fluorescence derived from OCO-2, MODIS, and reanalysis data. Remote Sensing, 2019, 11(5): 517. DOI:10.3390/rs11050517 |
| [10] |
Liu L Z, Zhao W H, Wu J J, Liu S S, Teng Y G, Yang J H, Han X Y. The impacts of growth and environmental parameters on solar-induced chlorophyll fluorescence at seasonal and diurnal scales. Remote Sensing, 2019, 11(17): 2002. DOI:10.3390/rs11172002 |
| [11] |
Lu X L, Liu Z Q, Zhou Y Y, Liu Y L, An S Q, Tang J W. Comparison of phenology estimated from reflectance-based indices and solar-induced chlorophyll fluorescence (SIF) observations in a temperate forest using GPP-based phenology as the standard. Remote Sensing, 2018, 10(6): 932. DOI:10.3390/rs10060932 |
| [12] |
Pierrat Z A, Bortnik J, Johnson B, Barr A, Magney T, Bowling D R, Parazoo N, Frankenberg C, Seibt U, Stutz J. Forests for forests : combining vegetation indices with solar-induced chlorophyll fluorescence in random forest models improves gross primary productivity prediction in the boreal forest. Environmental Research Letters, 2022, 17(12): 125006. DOI:10.1088/1748-9326/aca5a0 |
| [13] |
Li C H, Peng L X, Zhou M, Wei Y F, Liu L H, Li L L, Liu Y F, Dou T B, Chen J H, Wu X D. SIF-based GPP is a useful index for assessing impacts of drought on vegetation : an example of a mega-drought in Yunnan Province, China. Remote Sensing, 2022, 14(6): 1509. DOI:10.3390/rs14061509 |
| [14] |
张永江, 刘良云, 侯名语, 刘连涛, 李存东. 植物叶绿素荧光遥感研究进展. 遥感学报, 2009, 13(5): 963-978. |
| [15] |
Wu L S, Zhang X K, Rossini M, Wu Y F, Zhang Z Y, Zhang Y G. Physiological dynamics dominate the response of canopy far-red solar-induced fluorescence to herbicide treatment. Agricultural and Forest Meteorology, 2022, 323: 109063. DOI:10.1016/j.agrformet.2022.109063 |
| [16] |
Rascher U, Gioli B, Miglietta F. FLEX-fluorescence explorer : a remote sensing approach to quantify spatio-temporal variations of photosynthetic efficiency from space. Allen JF, Gantt E, Golbeck JH, Osmond B. Photosynthesis. Energy from the Sun. Dordrecht : Springer, 2008, 1387-1390. |
| [17] |
张永江, 赵春江, 刘良云, 王纪华, 马智宏, 王人潮. 被动荧光探测水分胁迫对玉米叶片影响的初步研究. 农业工程学报, 2006, 22(9): 39-43. DOI:10.3321/j.issn:1002-6819.2006.09.008 |
| [18] |
Zhang Y, Joiner J, Alemohammad S H, Zhou S, Gentine P. A global spatially contiguous solar-induced fluorescence (CSIF) dataset using neural networks. Biogeosciences, 2018, 15(19): 5779-5800. DOI:10.5194/bg-15-5779-2018 |
| [19] |
佀海翔, 王瑞燕, 王秋虹. 基于星载SIF的黄河流域植被光合动态时空演变及归因分析. 生态学报, 2024, 44(13): 1-11. DOI:10.20103/j.stxb.202309212050 |
| [20] |
Jeong S J, Schimel D, Frankenberg C, Drewry D T, Fisher J B, Verma M, Berry J A, Lee J E, Joiner J. Application of satellite solar-induced chlorophyll fluorescence to understanding large-scale variations in vegetation phenology and function over northern high latitude forests. Remote Sensing of Environment, 2017, 190: 178-187. DOI:10.1016/j.rse.2016.11.021 |
| [21] |
Yang S X, Yang J, Shi S, Song S L, Zhang Y Y, Luo Y, Du L. An exploration of solar-induced chlorophyll fluorescence (SIF) factors simulated by SCOPE for capturing GPP across vegetation types. Ecological Modelling, 2022, 472: 110079. DOI:10.1016/j.ecolmodel.2022.110079 |
| [22] |
Buareal K, Kato T, Morozumi T, Ono K, Nakashima N. Red solar-induced chlorophyll fluorescence as a robust proxy for ecosystem-level photosynthesis in a rice field. Agricultural and Forest Meteorology, 2023, 336: 109473. DOI:10.1016/j.agrformet.2023.109473 |
| [23] |
Lamchin M, Lee W K, Jeon S W, Wang S W, Lim C H, Song C, Sung M. Long-term trend and correlation between vegetation greenness and climate variables in Asia based on satellite data. Science of the Total Environment, 2018, 618: 1089-1095. DOI:10.1016/j.scitotenv.2017.09.145 |
| [24] |
Gitelson A A, Viña A, Arkebauer T J, Rundquist D C, Keydan G, Leavitt B. Remote estimation of leaf area index and green leaf biomass in maize canopies. Geophysical Research Letters, 2003, 30(5): 1248. |
| [25] |
Frankenberg C, Fisher J B, Worden J, Badgley G, Saatchi S S, Lee J E, Toon G C, Butz A, Jung M, Kuze A, Yokota T. New global observations of the terrestrial carbon cycle from GOSAT : patterns of plant fluorescence with gross primary productivity. Geophysical Research Letters, 2011, 38(17): L17706. |
| [26] |
Chen N, Zhang Y F, Yuan F H, Song C C, Xu M J, Wang Q W, Hao G Y, Bao T, Zuo Y J, Liu J Z, Zhang T, Song Y Y, Sun L, Guo Y D, Zhang H, Ma G B, Du Y, Xu X F, Wang X W. Warming-induced vapor pressure deficit suppression of vegetation growth diminished in northern peatlands. Nature Communications, 2023, 14(1): 7885. DOI:10.1038/s41467-023-42932-w |
| [27] |
Lin J Y, Zhou L T, Wu J J, Han X Y, Zhao B Y, Chen M, Liu L Z. Water stress significantly affects the diurnal variation of solar-induced chlorophyll fluorescence (SIF) : a case study for winter wheat. Science of the Total Environment, 2024, 908: 168256. DOI:10.1016/j.scitotenv.2023.168256 |
| [28] |
周蕾, 迟永刚, 刘啸添, 戴晓琴, 杨风亭. 日光诱导叶绿素荧光对亚热带常绿针叶林物候的追踪. 生态学报, 2020, 40(12): 4114-4125. DOI:10.5846/stxb201901010004 |
| [29] |
刘啸添, 周蕾, 石浩, 王绍强, 迟永刚. 基于多种遥感植被指数、叶绿素荧光与CO2通量数据的温带针阔混交林物候特征对比分析. 生态学报, 2018, 38(10): 3482-3494. DOI:10.5846/stxb201708211508 |
| [30] |
聂冲, 陈星安, 杨鹊平, 朱延忠, 徐睿, 邓陈宁, 周娟. 基于叶绿素荧光遥感的长江流域植被生产力时空变化及其气候驱动因素. 环境科学研究, 2023, 36: 2200-2209. |
| [31] |
Zhang Z Y, Cescatti A, Wang Y P, Gentine P, Xiao J F, Guanter L, Huete A R, Wu J, Chen J M, Ju W M, Peñuelas J, Zhang Y G. Large diurnal compensatory effects mitigate the response of Amazonian forests to atmospheric warming and drying. Science Advances, 2023, 9(21): eabq4974. DOI:10.1126/sciadv.abq4974 |
| [32] |
Walther S, Voigt M, Thum T, Gonsamo A, Zhang Y G, Köhler P, Jung M, Varlagin A, Guanter L. Satellite chlorophyll fluorescence measurements reveal large-scale decoupling of photosynthesis and greenness dynamics in boreal evergreen forests. Global Change Biology, 2016, 22(9): 2979-2996. DOI:10.1111/gcb.13200 |
| [33] |
Tu Z Q, Sun Y, Wu C Y, Ding Z, Tang X G. Long-term dynamics of peak photosynthesis timing and environmental controls in the Tibetan Plateau monitored by satellite solar-induced chlorophyll fluorescence. International Journal of Digital Earth, 2024, 17(1): 2300311. DOI:10.1080/17538947.2023.2300311 |
| [34] |
Ding Y B, He X F, Zhou Z Q, Hu J, Cai H J, Wang X Y, Li L S, Xu J T, Shi H Y. Response of vegetation to drought and yield monitoring based on NDVI and SIF. CATENA, 2022, 219: 106328. DOI:10.1016/j.catena.2022.106328 |
| [35] |
Yu L, Wen J, Chang C Y, Frankenberg C, Sun Y. High-resolution global contiguous SIF of OCO-2. Geophysical Research Letters, 2019, 46(3): 1449-1458. DOI:10.1029/2018GL081109 |
| [36] |
Chen X G, Huang Y F, Nie C, Zhang S, Wang G Q, Chen S L, Chen Z C. A long-term reconstructed TROPOMI solar-induced fluorescence dataset using machine learning algorithms. Scientific Data, 2022, 9(1): 427. DOI:10.1038/s41597-022-01520-1 |
| [37] |
Ma Y, Liu L Y, Liu X J, Chen J D. An improved downscaled Sun-induced chlorophyll fluorescence (DSIF) product of GOME-2 dataset. European Journal of Remote Sensing, 2022, 55(1): 168-180. DOI:10.1080/22797254.2022.2028579 |