 2024, Vol. 44
2024, Vol. 44文章信息
- 李颖, 何念鹏, 刘聪聪
- LI Ying, HE Nianpeng, LIU Congcong
- 植物功能性状网络: 概念体系发展与应用进展
- Plant functional trait networks: development of conceptual framework and advances in applications
- 生态学报. 2024, 44(18): 7944-7961
- Acta Ecologica Sinica. 2024, 44(18): 7944-7961
- http://dx.doi.org/10.20103/j.stxb.202401090083
-
文章历史
- 收稿日期: 2024-01-09
- 网络出版日期: 2024-07-14






2. 东北林业大学森林生态系统可持续经营教育部重点实验室, 哈尔滨 150040;
3. 中央民族大学生命与环境科学学院, 北京 100081
2. Key Laboratory of Sustainable Forest Ecosystem Management-Ministry of Education, Northeast Forestry University, Harbin 150040, China;
3. School of Life and Environmental Sciences, Minzu University of China, Beijing 100081, China
植物功能性状是植物在长期的进化和演化过程中, 表现出来可度量的、相对稳定的、且与植物各功能密切相关的特征参数[1—5]。在自然界中, 任何植物均具有种类繁多的功能性状;例如, 叶片大小、叶片厚度、叶片各种功能元素含量、植株高度、根系直径、根系长度、种子大小、种子质量等等[6]。这些功能性状既相互独立, 又相互关联;协同、权衡、耦合或解耦等, 总之, 彼此之间的关系非常复杂[7—10]。
20世纪90年代以来, 植物功能性状研究取得了一系列突破性进展, 为其高速发展注入了新的活力, 如化学计量理论、叶经济型谱理论、代谢理论等一系列的理论的提出[11—13]。然而, 纵观前期的研究, 绝大多数还是聚焦于单一功能性状的变异规律或几个功能性状间的协同、权衡关系及其与环境之间的关系。在复杂的自然界中, 理论上任何单一功能性状的改变都不足以实现植物对复杂环境变化的响应与适应。植物为了实现长期生存与繁衍的需求, 必须通过多种功能性状相互作用实现其单一功能优化或者多种功能平衡, 形成多种适应机制以及生长策略, 以应对复杂多变的外界环境。大量研究表明, 多性状之间存在紧密的联系, 然而以往的分析方法大多仅重视单一或两两性状之间相关性的变化, 却忽略了多性状间复杂的关系[5, 7, 14—16]。理论上, 只有综合考虑多性状间这些庞杂的关系, 才有望从性状的角度深入揭示植物对环境变化的适应对策(图 1)。

|
| 图 1 植物功能性状网络将是揭示多种功能性状复杂关系的有效途径 Fig. 1 Plant functional trait networks will be an effective approach to reveal complex relationships among various functional traits |
还原论指向事物的微观性质, 重在从宏观走向微观;而系统论反其道而行, 把目光指向事物的宏观性质和复杂性, 重在从微观走向宏观。复杂科学以复杂系统为研究对象, 尝试采用跨学科的方法来揭示复杂系统背后的统一性规律[17]。当前, 是还原论科学一统天下的时代, 科学理论整体重分析而轻综合。相对而言, 复杂论还很年轻, 甚至还没有一个普遍公认的概念体系和学科框架;然而, 其跨学科范式、多尺度视角以及整体论的世界观, 为揭示各种复杂的自然系统、各类复杂的社会问题等提供了新的科学范式。1977年, 著名科学家伊利亚·普利高津(Ilya Prigogine)就凭借“耗散结构论”这一复杂科学的开创性理论成果获得诺贝尔化学奖。2021年, 复杂科学再次受到了诺贝尔奖的青睐, 由日裔美籍科学家真锅淑郎(Syukuro Manabe)等建立的“地球气候的物理模型、量化其可变性并可靠地预测全球变暖”获奖。在复杂科学研究中, 复杂网络(Complex networks)对于科学研究而言是一种相对较新的思想和理念, 同时对于研究复杂系统而言又是一种新的技术和方法。
目前, 复杂网络已经广泛应用于分子生物学、社会科学、生物科学和交通运输等各个领域[5]。人们通过对复杂网络的研究, 可以对自然界各种复杂关系进行量化和预测。受到复杂科学的启发并结合对植物自身作为一个复杂系统的判断, He等[9]提出, 复杂网络可以提供一种有效的方式探究植物多性状的复杂关系, 并有助于揭示植物对环境变化的多维度适应策略。尽管目前网络分析已广泛应用于多个领域, 但是在植物功能性状研究的应用却非常罕见;除了缺乏完善的植物功能性状网络的概念与方法外, 系统性和配套性的植物功能性状数据缺失也是重要原因。
为突破上述两大难题, 何念鹏团队近期将复杂网络的理念运用到植物功能性状的研究中, 构建了植物功能性状网络(Plant trait networks, PTNs)的理论体系和方法, 为植物功能性状的研究提供了一种新的思路和方法[9]。本文系统梳理了PTNs的理论框架、核心参数和计算方法, 并介绍了近期几个PTNs在自然生态系统的应用案例, 希望能推动该领域的快速发展。
1 植物功能性状网络思维的萌芽众所周知, 植物功能性状是植物在长期的进化和发展过程中, 与环境相互作用而形成的与植物生长、发育、存活和繁殖等过程相关的形态结构和生理特征参数[3—5]。由于它与植物自身的遗传信息密切相关, 对特定植物功能性状而言首先是具有相对稳定性, 并以此为基础应对环境变化时表现出了一定的时空变异性或可塑性[1, 18]。植物功能性状这种相对稳定性和应对外界环境变化的可塑性, 使人们可以通过观测植物功能性状的变来揭示植物在各个层面(从器官、物种、群落到生态系统)适应环境的策略和机制[2, 19]。这也是植物功能性状当前受到越来越多关注的重要原因之一, 人们甚至希望以功能性状为媒介, 打通生态学研究不同时空尺度研究间的壁垒, 推动整个学科的全面、快速发展。
近年来, 国内外学者对植物单一功能性状的时空变异, 功能性状间的关联以及植物功能性状网络进行了大量研究, 并取得了卓有成效的成果[7, 9, 13—14, 20—21], 其发展过程大致如图 2所示。

|
| 图 2 植物功能性状研究进展:从单一功能性状到功能性状网络 Fig. 2 Advances in plant functional trait research-from single functional traits to plant functional trait networks |
事实上, 植物功能性状具有其自身的多功能性(multi-functionality)。一方面, 植物任一种功能性状均具有多种功能和作用机制。以气孔为例, 气孔常被比喻为CO2和水分进出叶片的闸门。作为植物与外界环境进行气体交换的重要通道, 气孔的形态和行为与光能利用效率、水分利用效率和热量平衡等多种功能都密切相关[22]。叶片上的微毛具有多功能性, 它们不仅可以通过反射太阳辐射来减少叶片受损, 同时还有效地防止叶片水分的蒸发散失[23]。另一方面, 植物的各种功能并非仅仅依赖于单一的性状、组织或器官。以光合作用为例, 它并非由栅栏组织或海绵组织单独完成, 而是由二者相互协调、优化的结果。大量研究表明, 植物的多种功能性状之间密切相关, 甚至可以共同调控同一功能。举例来说, 叶脉性状和气孔性状之间存在着密切的相关性, 它们共同影响着植物的水分利用效率[24—25]。由此可见, 在植物适应外部环境变化的过程中, 单一功能性状的改变并不足以满足其所有的适应需求, 而是需要多种功能性状之间的协同作用, 才能共同实现植物功能的平衡与优化[5]。
目前, 植物功能性状的研究对象已经逐渐从单一功能性状转变为多种功能性状[26—28]。最具有代表性的研究成果之一是由Wright等人提出的叶经济型谱理论[13, 16]。此后, 许多学者陆续扩展了该理论, 涵盖了植物其他器官的经济型谱, 如干经济型谱、根经济型谱, 以及一些生理性状谱系(如水力学特征等), 甚至是整体的植物经济型谱[7, 10, 29—32]。然而, 许多研究表明, 这些功能性状间的复杂关系会受到功能群和环境的影响而变化[33], 具有明显的多维度特征[5, 9]。
在还原论指导下, 即使在多种功能性状“集合(constellation)”备受关注的当下, 植物多种功能性状的研究仍然趋向于简单化的趋势, 即“降维处理”。在具体操作过程中, 如果多功能性状紧密相关, 则可将它们共同降为一个简单的维度。诸如主成分分析这类方法, 为降维处理提供了理论和分析的支撑。植物叶经济型谱则是降维处理的典型案例, 叶经济型谱通常被认为是沿单轴的变化, 从快速生长、组织寿命短的一端到缓慢生长、组织寿命长的另一端[5, 13]。
单一维度的简化方法似乎存在一定的局限性, 可能会掩盖自然界植物的整体适应模式。正如前文所述, 植物的功能性状通常与其他性状相互作用, 共同影响多个功能, 从而提高植物的生存、逆境应对和繁殖等能力, 并非仅限于对单一功能的影响[23, 26, 28]。只有解开多种功能性状之间错综复杂的关系, 才能从功能性状整合的角度深入阐释植物整体表型的适应机制(图 1和图 3)。然而, 探究多种功能性状之间的错综复杂关系, 以及这些关系如何随着物种、生境和环境的变化而变化, 依然是一个极具挑战性的科学难题。

|
| 图 3 自然界多种植物功能性状间复杂的关系与植物适合度 Fig. 3 Complex relationships among various plant functional traits impacting plant fitness in nature |
自牛顿力学问世以来, 还原论的研究方法曾在现代科学的多个领域占据主导地位。这种方法将系统分解为许多基本单元, 通过对系统的“分析”和“分解”, 深入探究系统的细节, 揭示系统的组成元素或部件。然而, 这种方法忽视了这些元素是如何相互作用和组合成系统的。因此, 还原论方法在处理“简单系统”时表现出色, 但在面对复杂系统时存在很大的局限性。随着时间的推移, 复杂网络作为一种新兴的研究途径应运而生, 并在21世纪变得越来越重要。复杂网络理论认识到一切复杂系统的结构都可以被描述为复杂网络。自然界中存在着大量的复杂系统, 这些系统可以通过各种形式的网络来加以描述。因此, 复杂网络的出现填补了还原论方法在处理复杂系统时的不足, 为更好地理解和分析复杂系统提供了一种新的方法。
复杂网络的概念起源于图论, 并逐渐演变成解决现实世界中复杂系统问题的重要工具。网络由节点和边组成, 节点代表系统中的个体或基本单元, 而边则表示节点之间的连接或关联关系。随着小世界网络(Small-world network)模型和无标度网络(Scale-free network)模型的提出, 网络研究进入了一个新的时代[34—36]。一切复杂系统的结构都是复杂网络, 自然界中存在的大量复杂系统都可以通过形形色色的网络加以描述。
目前, 复杂网络的思想和理念已经被广泛应用于各个学科领域。在社会科学中, 人与人之间的关系被视为社会网络, 研究人们之间的联系、交流和组织结构[37]。在系统生物学领域, 代谢网络被用于研究生物体内部的化学反应和新陈代谢过程, 从而揭示生物体内部各种分子之间的相互作用[38]。此外, 复杂网络还广泛应用于代谢网络外[39]、交通运输网络[40—41]、基因网络[42]、微生物网络[43—44]等多个领域, 为研究者提供了深入理解系统结构和功能的新途径[5]。为便于理解, 我们在此列举了几种常见的复杂网络在其它领域的应用, 包括节点、边以及相关网络的定义(表 1)。
| 网络Networks | 节点Nodes | 边Edges | 定义Definition | 参考文献References |
| 社会网络Social network | 人、社会岗位 | 亲缘关系、业务往来关系、朋友关系 | 社会网络是指社会个体成员之间由于互动而形成的相对稳定的关系体系, 社会网络关注的是人与人之间的互动和联系, 社会互动会影响人们的社会行为 | [37] |
| 交通网络Transportation network | 交通站点 | 地铁线、飞机航线等 | 交通网络指各种运输网、邮电网构成的整体交通网, 亦称运输网络 | [40—41] |
| 蛋白质互作网络Protein-protein interaction network | 蛋白质分子 | 分子间的相互作用 | 蛋白质互作网络是指由单独蛋白通过彼此之间的相互作用构成的互作网络, 从而参与生物信号传递、基因表达调节、能量和物质代谢及细胞周期调控等环节 | [45] |
| 微生物网络Microbial network | 微生物种类 | 微生物间相互作用 | 微生物网络是指可以探究出不同环境中的不同微生物种间作用关系或者物种的共存模式 | [44] |
| 代谢网络Metabolic network | 分子 | 生化反应 | 代谢网络是指细胞内所有生化反应所构成的网络, 反映了所有参与代谢过程的化合物之间以及所有催化酶之间的相互作用 | [38—39] |
| 信息网络Information network | 网站页面、作者论著 | 网络链接地址、文献引用 | 信息网络专指电子信息传输的通道, 是构成这种通道的线路、设备的总称 | [46] |
| 植物功能性状网络Plant functional trait network | 植物功能性状 | 功能性状之间的协同与权衡关系 | 根据植物多种功能性状间相互关系构成的多维度网络, 其整体特征可反映植物个体和群落对环境的响应与适应机制、植物群落结构维持和生产力优化的机制 | [9] |
在传统的生态学研究领域, Proulx等人[47]率先提出了将复杂网络理论和方法运用于生态和进化研究的设想。该文回顾了网络思想在基因和蛋白质领域的最新应用, 并指出这些研究通过应用现有的网络分析技术, 对生物系统的组织和功能提供了新的见解。此外, 文章还指出, 认识到网络思维和理念在进化和生态应用中的共性以创建生物网络的预测科学是未来的巨大挑战。遗憾的是, 他仅仅建议将复杂网络思想应用到生态学研究中, 但并未给出具体定义、参数和实施方案。
对于植物功能性状而言, 网络分析有助于可视化多个功能性状之间相互依赖的多维度关系, 而非与传统方法一样只关注单一或两两性状之间的二维关系[13, 21]。在前人研究的启发下, 一些近期的研究已经开始勾勒出植物功能性状网络的雏形[48], 并尝试以网络化的形式展示多个性状之间的相互依赖关系[20, 29, 49—53]。此外, 科研人员也在逐步开始尝试应用具体的网络参数来量化这种复杂关系[49, 54]。然而, 植物功能性状网络目前仍处于起步阶段, 其理论框架、构建方法以及生态意义的网络参数都有待进一步研究和完善。特别是对于自然群落, 尤其是在大尺度上, 不同生活型及功能群植物通过植物功能性状间关系的改变适应环境的策略尚不清楚[5, 9]。
生态学致力于研究生物与生物、生物与环境之间的关系。植物是生态系统(包括人类)赖以生存的物质和能量来源, 因此, 植物适应环境及群落生产力形成与稳定机制一直是相关学科的核心科学问题之一。如上所述, 功能性状是植物对环境适应的表现, 功能性状之间存在着紧密的关系, 单一功能性状的改变并不能实现植物对环境的响应与适应。尽管从单个或多个功能性状指标自身变化规律的研究具有其特定的科学价值和进步意义, 但却难以真实地揭示功能性状整体关系与多维响应机制。因此, 如何将复杂网络的研究理念运用到植物功能性状的研究中, 进而构建新型的植物功能性状网络, 是解决植物多种功能性状间复杂关系及其综合适应途径的一条有效途径。
3 植物功能性状网络的定义和内涵何念鹏等人基于前人的研究, 将复杂网络的理念引入了植物功能性状的研究, 并首次提出了植物功能性状网络的概念[9]。植物功能性状网络(Plant trait networks, PTNs)被定义为由多种功能性状之间的相互关系构成的多维度网络, 其整体特征或节点特征能够表征植物性状之间的复杂关系, 揭示植物个体、功能群以及群落对环境变化或干扰等的响应与适应途径(图 4)[5]。在PTNs中, 节点代表着各种功能性状, 而边则代表着两两功能性状之间的关系。

|
| 图 4 多维植物功能性状网络的构建 Fig. 4 Construction of a multidimensional plant functional trait networks LTN, Leaf trait network, 叶片性状网络;r21表示性状1和性状2之间的相关系数 |
通过空间拓扑技术, PTNs具有全面捕获和可视化植物性状之间联系的潜力, 从多维度的角度为揭示植物对环境或资源变化响应与适应策略提供新的视角(见图 4)[5, 9]。在具体操作过程, PTNs可以进一步细分为更具有可操作性的叶片功能性状网络(Leaf trait networks, LTNs)、根系功能性状网络(Root trait networks, RTNs)、花功能性状网络(Flower trait networks, FTNs)、功能元素网络(Multiple element networks, MENs)以及其它性状网络。但无论如何分类, 其核心本质均是利用多维度网络的思维以及网络特征参数来探讨植物对环境适应机制与响应方向。简言之, 植物功能性状网络理论体系的建立, 为植物适应环境研究提供了多维度、定量化、可视化的新途径, 它可以与生态学的经典途径“进化历史、环境过滤、竞争分化”相结合, 去发展、检验相关的机理、假说或探讨植物响应与适应途径(图 5)。

|
| 图 5 植物功能性状网络理论体系为植物适应环境研究提供了多维度、定量化、可视化的新途径 Fig. 5 The theoretical framework of plant functional trait networks provides a multidimensional, quantitative, and visualized approach to studying plant adaptation to the environment |
植物功能性状网络通常应具备以下基本特征[5]:(1)在特定的环境中, 任何一个植物功能性状都有可能成为网络中具有重要意义的中枢性状, 具有不可替代或不可或缺性;(2)功能性状之间的相互作用应具有生态意义, 反映植物对环境的响应和适应策略;(3)基于PTNs的整体参数和关键节点参数可以观测到由单一功能性状或两两功能性状之间相关性无法完全体现的适应机制[9]。参照植物功能性状的分类, PTNs可以按植物不同器官进行分类, 如叶片功能性状网络、根系功能性状网络等;也可以按功能系统划分为经济性状网络或水力性状网络等;以及根据其它分类标准进行多种分类, 功能元素网络等。在具体研究过程中, 可以根据科学目的或数据可获取性等加以灵活应用。
植物功能性状网络理论体系也蕴含或包容了许多生态学的经典理论。以植物叶片功能元素网络为例[55—56], 它可以在一定程度上反映李比希最小因子定律(Liebig′s law of minimum[57])和生态化学计量理论(Theory of ecological stoichiometry), 尤其是与限制元素相关的假设[11, 58]。当外界资源或环境发生较大变化时, 植物会通过调节功能性状网络来适应环境, 因此功能性状网络并非一成不变, 而是随着环境的变化而变化。假设在理想情况下植物具有一个最收敛或相对稳定的球形元素网络, 则植物可以通过调节网络中的复杂关系, 改变整体形状(网络整体参数)或核心功能元素(网络节点参数)来适应环境, 从而提高植物适合度(图 6)[2, 5]。当外界资源、环境发生变化时, 植物功能元素网络可能会沿着改变的资源或环境而发生变形, 凹陷或拉伸等, 但偏移的程度是有限的。在一般环境变化或扰动中, 植物可以通过关键节点的移动和功能性状网络的整体变形来适应环境, 实现生存、生长和长期繁衍的多种功能需求。然而, 当外部资源或环境变化超出了网络的适应能力时, 整个功能元素网络可能会瓦解, 导致植物死亡并从群落中消失[5]。

|
| 图 6 叶片多元素网络对变化的资源和环境的潜在响应 Fig. 6 Leaf multielement networks′ potential responses to changes in resources and environment |
PTNs的应用将提升我们对植物多维适应策略的理解[2, 5, 59]:
(1) 可视化多种功能性状之间的复杂关系。PTNs通过边和节点来描述多种功能性状之间的复杂关系, 将多种功能性状间的复杂关系可视化并使其易于理解[20—21, 48, 52]。
(2) 阐明多种功能性状所构成的“谱”或“维度”。PTNs包含了多个功能模块, 高度相关的功能性状将被汇总到一个“功能模块”中, 而每一个“功能模块”可能对应于一个或多个“功能性状谱”。在PTNs中, “叶经济型谱”显然被组装成单独的模块, 该模块侧重于关键经济功能性状之间的关系[13, 60], 可能对应于与生产力相关的维度(见图 4)。而PTNs可以突出植物多个维度的适应方式, 包括生产力维度、温度维度、水分维度等, 未来可以统一或兼容当前多种不同的理论或观点(图 4)。
(3) 量化多个功能性状之间的整体依赖性[5, 9]。并非所有功能性状都可以直接相连, 两个功能性状之间的间接相连可能需要一个甚至多个中间功能性状作为桥梁。因此, 具有更高的边密度, 更短平均路径长度和更短直径的PTNs, 意味着多个功能性状之间具有更强的协调性[53]。通过分析PTNs的拓扑结构特征, 如边密度、平均路径长度和直径, 可以量化多个功能性状之间的整体依赖关系, 从而揭示出植物功能性状的协同性。
(4) 识别关键功能性状[5]。通过计算PTNs的节点参数(度、紧密度、介数、聚类系数等), 可识别网络中的关键性状。具有较高连接度的功能性状称为“中心性状”(hub traits), 具有较高介数的功能性状则是“中介性状”[49]。Li等[59]发现在中国东部森林叶片功能性状网络中, 与叶片厚度相关的叶经济性状是网络中的中心性状。
(5) 探索功能性状及其关系对不同环境的响应与适应。功能性状间的关系并非一成不变, 而是会随着环境的变化而发生变化, 例如经济功能性状和水力功能性状在比较干燥的地方耦合而在较湿润的地区解耦[28, 31, 61]。Li等[58]发现功能性状网络在温水条件较好的森林叶片功能性状网络中关系更加紧密且复杂。
(6) 量化植株的表型整合。表型整合的研究旨在确定单个功能单元内(或功能模块内)的功能性状是否存在密切相关性, 并辨识不同模块之间的关联性[62]。PTNs的模块性可被用来评估功能性状模块之间的联系, 进而有助于解释表型整合的问题[9, 62]。此外, PTNs还蕴含着更多未来可探索和拓展的应用。总之, 通过揭示功能性状之间的关联, PTNs为我们提供了一种更为有效的途径来理解植物在多个维度上的适应性[5]。
4 植物功能性状网络的构建方法及关键参数 4.1 植物功能性状网络的构建方法网络通常是由一系列节点和边所共同构成。因此, 研究人员将植物功能性状视为节点, 而功能性状-功能性状的关系作为边, 从而构建植物功能性状网络(PTNs)[5]。为了构建PTNs, 首先需要计算功能性状-功能性状关系矩阵。对于功能性状之间的双变量关系, 可以使用Pearson相关系数、Spearman相关系数、系统发育相关系数以及其它参数检验方法对其进行量化[49, 54]。有一点需要特别指出:植物功能性状之间存在紧密关系, 原因主要分为三类[32, 63]。第一, 功能性状之间的关联性是结构优化的结果, 如结构与生理之间的关系, 结构的数量与大小直接决定了植物的生理速率或生长过程。第二, 功能性状之间的关联性是功能平衡的结果, 两个独立的功能性状对更高等级的功能可能具有更大的贡献。例如, 气孔形态性状和叶脉性状都对水分的传导和优化起到至关重要的作用。第三, 功能性状之间的关联性是环境筛选而形成的生态位优化适应选择的结果。例如, 在光受限的情况下, 主叶脉分支的单子叶植物更容易结出肉质多汁的果实。
为了避免功能性状之间的虚假相关性, 研究人员需要设置一个阈值(例如|r|> 0.2, P < 0.05), 以确定功能性状之间是否真的存在相关性[50]。然后, 将超过阈值的关系设为1, 将低于阈值的关系设为0, 从而建立邻接矩阵A=[ai, j]且ai, j∈[0, 1];因此, 邻接矩阵A仅显示了成对植物功能性状之间是否存在联系[5]。最后, 通过借助于R语言软件中的“igraph”软件包, 通过编程完成PTNs可视化并计算PTNs的相关参数, 目前已完成“PTN”R包的主体代码, 预期于近日公开发表(图 7)。

|
| 图 7 植物功能性状网络的关键参数 Fig. 7 Key Parameters of plant functional trait networks 图中每个圆圈代表特定植物功能性状, 边代表功能性状之间的关系; 对于每个参数而言, 从左到右三个网络代表参数值依次增大) |
通常, 科研人员利用网络的节点参数来进一步识别PTNs关键节点, 利用网络的整体参数来表征功能性状间的紧密关系。在此, 本文重点介绍PTNs的4种节点参数和5种整体参数的定义、计算方法和生态意义(见表 2)。其中, PTNs的拓扑角色通常可通过度(Degree)、紧密度(Closeness)、介数(Betweenness)和聚类系数(Clustering coefficient)等参数来识别, 而网络的连通性和复杂性则可通过边密度(Edge density)、平均路径长度(Average path length)、直径(Diameter)、平均聚类系数、(Average clustering coefficient)、模块度(Modularity)来量化[5](见图 7)。
| 分类Categories | 参数Parameters | 定义Definition | 公式Formula | 公式注释Formula notes | 生态意义Ecological significance |
| 网络整体参数Network overall parameters | 边密度 | 性状网络中观测到的边的数量与最大潜在边数量的比值 | 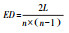 |
式中, L是网络的实际边数, n是节点数。 | 较高的边密度意味着性状间的协同性较好, 此时植物的资源利用效率或生产效率较高 |
| 平均路径长度 | 性状网络中连接两个性状的路径中, 最短一条称为最短路径, 所有最短路径平均值即为平均路径长度 | 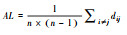 |
式中, dij表示节点vi和vj间的距离, n表示网络中节点的个数。 | 较短的平均路径长度意味着性状间的协同性较好, 此时植物的资源利用效率或生产效率较高 | |
| 直径 | 性状网络中所有最短路径中最长的一条 | 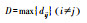 |
式中, dij表示节点vi和vj间的距离。 | 较短的直径意味着性状间的协同性较好, 此时植物的资源利用效率或生产效率较高 | |
| 平均聚类系数 | 网络所有节点聚类系数的均值 |  |
式中, 对于节点vi, li指节点vi的邻居节点间实际相连的边数;ti指节点vi的邻居的数目。 | 较低的平均聚类系数意味着特定性状间的协同性较好, 反之, 更倾向于组成功能小团体实现特定功能 | |
| 模块度 | 性状网络可以划分为子网络或模块, 模块度用来形容子网络(或模块)间分离的程度 | 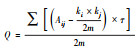 |
式中, m是边的数量, Aij是第i行和第j列的A邻接矩阵的元素, ki是i的阶数, kj是j的阶数;如果j和i相同, 则是1, 否则为0。 | 较高的模块度代表功能模块界限清晰, 功能模块内部连接紧密, 功能模块间的连接很弱, 意味着特定的性状所构成的功能模块执行特定的功能 | |
| 网络节点参数Network node parameters | 度 | 与该节点相连的所有边的数量 |  |
式中, aij表示节点vi、vj间的关系强度。 | 网络中与其它性状连接最多的性状度最高, 是网络的“中心性状”, 可能起着影响整个表型的中心调控作用 |
| 紧密度 | 该性状与其它性状相连的最短路径的平均值的倒数 | 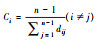 |
式中, ci是指节点i的紧密度, dij是指节点vi和vj之间的最短距离, n是指网络中节点的总集。 | 性状拥有较高的紧密度意味着该性状与网络中较多性状具有紧密的联系, 也是网络中的中心性状 | |
| 介数 | 通过该节点的所有最短路径的数量 | 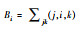 |
式中, (j, i, k)指节点vj和vk之间最短路径经过节点vi的数目。 | 性状拥有较高的介数, 意味着该性状是连接功能模块的“桥梁”或“中介”, 环境对该性状的筛选能够很大程度影响多个功能模块间的协调关系 | |
| 聚类系数 | 与该性状相连的性状为该性状的邻接性状, 聚类系数为邻接性状之间相连的概率 | 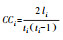 |
式中, 对于节点vi, li指节点vi的邻节点间实际相连的边数;ti指节点vi的邻接性状的数目。 | 性状具有较高的聚类系数, 则意味着该性状是网络功能模块的中心性状, 在实现某一特定功能时起重要作用 |
植物通过多种功能性状的协同/优化来适应环境, 这些功能性状间的复杂关系可以表示为以一个相关关系为依托的多维度网络。目前, 关于不同气候梯度下群落间功能性状网络结构并未开展任何研究, 人们对其空间变异规律、影响因素或调控机制仍然不清楚[9]。基于先前的生态生理学理论, 研究人员提出了几个假设:森林叶片功能性状网络(Leaf trait networks, LTNs)的模块化复杂性将随着物种丰富度、气候温暖度和湿度的增加而增加, 这反映了该气候条件对表型的限制减少和生态位分化的机会增加[5]。此外, 由于叶片厚度及其相关的叶片经济功能性状对植物的多种功能具有不成比例的贡献, 这些功能性状可能是LTNs的中心性状, 它们的变异可能会引起植物表型随之发生明显改变[5, 58]。
为探究以上假设, Li等[59]以叶片性状网络为研究对象, 利用距离长达3700 km的中国东部南北样带(North-South Transect of Eastern China, NSTEC)中九个典型森林394个树种的35种性状数据, 构建每个植物群落的LTNs, 并计算网络的边密度、直径、平均路径长度、平均聚类系数和模块度, 揭示叶片功能性状间关系的紧密性和复杂性, 结合群落物种丰富度和气候数据, 验证上述科学假设(图 8)。同时, 计算网络中每个节点的度、紧密度、介数和聚类系数识别网络中的关键功能性状。结果表明, 由于表型约束的减少和生态位分化机会的增加, LTNs会随着物种丰富度和气候温暖湿度的增加而增加模块化复杂性。此外, 通过节点参数识别出叶片厚度及其相关的叶经济性状与其他性状具有更多的关联, 这些性状是LTNs中的关键性状和骨架, 其稳定性最高, 难以随环境的改变而改变[5, 59]。

|
| 图 8 中国东部9个森林群落的叶片功能性状网络 Fig. 8 Leaf trait networks of nine forest communities in Eastern China |
在自然界中, 植物通过调节多种叶片功能性状及其相互间的关系, 共同优化光合作用以实现维持成本与获取收益之间的平衡。因此, 多种功能性状之间的复杂关系可能形成一个庞大的关系网络, 并通过功能性状网络的整体参数和核心节点参数来适应环境。在该网络中, 节点代表全球尺度的叶片功能性状, 而边代表两两功能性状间的关系;其中, 边的宽度和长度对于该功能性状网络而言也是十分重要的参数。更重要的是, 由于叶片功能性状之间的关系在不同区域中存在着明显的差异(方向或强度), 因此亟需建立基于功能性状间关系强弱进行加权的新型叶片功能性状网络, 以更准确地表达和量化各种叶片功能性状间的相互依赖性(图 9)。

|
| 图 9 用于计算叶片功能性状网络的理论基础和方法 Fig. 9 Theoretical basis and methods for calculating leaf trait networks 植物通过叶片相互作用的多性状协调适应环境或优化功能, 整合的植物性状网络(LTNs)可以帮助捕获不同性状之间的复杂关系, 并探索植物的适应策略; 将植物性状间复杂的关系可视化为植物性状网络; 图中LTN可以表示为由边(线)连接的一组节点(圆圈); 图中不同颜色的圆圈代表不同叶片性状。由于性状之间的距离和强度不同, 准确量化性状间的关系有助于我们更好的了解植物的适应策略; 因此, 我们用边(红线)的宽度代表性状间关系的强弱, 用边的长度代表性状间的距离, 以此对无权重性状网络进行加权, 从而得到加权的性状网络 |
基于上述考虑, Li等[64]利用上述方法和参数, 构建了基于全球尺度2000多种植物六个叶片功能性状的叶片功能性状网络(LTNs);通过LTNs可视化了所有功能性状间的复杂关系, 并通过网络参数量化它们之间的差异[64]。实验结果表明:在全球尺度, 不同生活型和生长型的LTNs存在显著差异, 表明不同类型的植物其生长和生存策略存在明显的差异(图 10)。结果显示阔叶树种的功能性状关系比针叶树更紧密, 这表明阔叶树可能比针叶树在实现吸收和传递物质等功能时更有效率。而灌木的功能性状关系比乔木更紧密, 可能是由于灌木需要多种功能性状的有效合作才能使其在有限的资源中实现多种功能。此外, 叶片氮浓度和叶片寿命在LTNs中具有最高的中心性。因此, 这两个性状的变异可能影响植物的整体表型, 未来应给予更多的关注和重视。

|
| 图 10 不同生长型和生活型植物叶片性状网络 Fig. 10 Leaf trait networks of plants with different growth forms and life forms LL:叶片寿命;LMA:比叶重;Nmass:叶片氮含量;Pmass:叶片磷含量;Amass:光合速率;Rmass:暗呼吸速率 |
目前, 科研人员对藤本植物和乔木之间的功能差异做了大量的探究工作并取得系列进展。然而, 关于藤本植物和乔木的多种功能性状是如何整合在一起的研究却很少。随着功能性状相关网络的发展, 研究人员根据多个功能性状的拓扑关系, 利用网络分析来区分它们的差异。在具体操作中, 本研究使用了公开发表的数据集, 其中包括16种藤本植物和16种乔木的17个植物功能性状, 含12个叶片功能性状, 3个茎干功能性状和2个枝条功能性状[65]。研究结果表明:叶柄长度是乔木性状网络中高度连接的性状, 而磷是藤本植物性状网络中高度连接的性状。与乔木相比, 在两种森林类型中, 藤本植物的特征网络都具有更强的模块化, 表明环境变化对藤本植物的影响小于乔木。如果将植物功能性状重新划分为经济性状、水力性状和机械性状, 则藤本植物的经济性状具有较高的连通性, 而乔木的水力性状则具有较高的连通性。因此, 当资源条件较差时, 增加植物特定关键功能性状的连通性是具有成本效益的[66]。功能性状模块的数量和组成随生活型和环境条件的不同而改变, 植物通常可以选择不同的性状组合来适应当地的环境。该研究表明植物功能性状网络的变异会提高植物的适应性, 也从一个独特的角度为解释全球变化下藤本植物丰富度增加的现象提供了新的见解。
此外, 研究人员借鉴功能性状网络的思维, 提出功能元素网络假设(Multi-element network hypothesis), 即多元素网络的高可塑性可以使植物快速响应环境变化, 从而更具竞争力, 使植物在群落中具有较高优势度, 表现为更高的相对生物量[67]。具体研究中, 研究者基于内蒙生态站的N沉降模拟实验样地采集的5个物种18种元素及长期的生物量测定数据, 采用间隔分析和矩阵对矩阵的方法对多元素网络的特点进行了探究, 并验证了功能元素网络假说。实验数据表明:植物功能元素网络具有独特性, 不同物种在不同氮添加梯度下均可通过多元素网络被识别。不同物种对模拟氮添加的响应不同, 若其多元素网络可以随N添加增加而快速响应(即可塑性较高), 其在群落中的相对生物量呈增加趋势或保持不变, 相反, 若多元素网络可塑性较低, 其相对生物量则呈下降趋势[67]。该研究成功地将功能元素网络应用到全球变化梯度控制研究中, 可能成为未来全球气候变化下预测群落结构和功能变化的新思路。
6 植物功能性状网络的潜在应用领域展望PTNs为植物功能性状的研究提供了综合视角, 实现了单个功能性状到多个功能性状拓展的尝试, 开辟了新的方法阐明植物对环境和资源变化的多维度响应和适应机制。基于多方面考虑, 本研究认为植物功能性状网络可能在诸多领域具有巨大的应用潜力(见图 11), 具体但不限于如下几个方面[5, 9, 59]:

|
| 图 11 植物功能性状网络的潜在应用领域展望 Fig. 11 Prospects for the potential applications of plant functional trait networks |
(1) 探究大尺度下自然群落植物功能性状网络的时空变异特征及其影响因素, 从功能性状网络的角度揭示植物对不同环境的多维度适应机制(见图 8)。
(2) 揭示不同植被类型、功能群、生活型植物功能性状网络的差异, 探讨在区域或全球尺度上植物功能性状网络可能存在的可预测规律性变化, 并基于功能性状网络对复杂自然群落或生态系统进行量化和预测。
(3) 探索叶片功能性状网络和根系功能性状网络的关联, 研究二者在结构上的异同以及其对环境变化的响应是否存在差异, 为深入探究植物地上与地下关系提供新的有效途径。
(4) 在植物由种子萌发到开花结果的不同生长阶段, 研究植物功能性状网络的变化方向以及关键节点是否会发生变化, 以理解植物对不同生长阶段的适应策略。
(5) 利用植物功能性状网络探究植物群落演替或恢复过程中植物对环境的适应策略, 为不同演替阶段植物的响应策略提供科学解释, 同时可能对模型改进具有重要意义。
(6) 在全球变化的大背景下, 通过控制实验探讨植物对气候变化的响应机制, 将植物功能性状网络与模拟全球变化情景相结合, 有助于更好地理解当前状态并预测未来变化趋势, 这可能是蕴含重大原创性突破而特别值得期待的领域。
(7) 植物功能性状网络的研究方法为表型整合(Phylogeny integration)研究提供明确的量化手段, 为花性状及表型之间关系的可视化提供新的思路, 为表型整合研究甚至“物种、群落、生态系统、区域”整合研究提供有效手段和研究方向的指引。
综上所述, 植物功能性状网络是植物性状研究的一个新领域, 目前看还是比较超前的。在此, 本研究大胆预测其方法学上的创新, 未来甚至可能会比肩1990s的通径分析(Path analyses)或2000s的结构方程(Structural equation modeling, SEM)。在应用上, 未来将不仅局限于上述7个领域;其核心思维方式将可能拓展到传统生态系统网络结构的各个领域, 如食物链、食物网、植物不同器官间的关系、不同生物类群间的关系等。无论未来进展如何, 多维度网络分析所提供的新的理念、技术和参数, 将在一定程度上更好地揭示复杂的自然界。在此也必须指出, 开展植物功能性状网络的相关研究需要“系统性和配套性”植物功能性状数据, 很难通过简单的收集数据来获得;诸如世界著名性状数据库TRY(https://www.try-db.org)[68], 目前都难以完美地操作和运行PTNs。因此, 如何在野外调查和控制实验中获取符合要求的数据, 快速拓展数据源, 是当前发展和检验植物功能性状网络最大的难题。未来, 人们可以利用全球广泛分布的长期野外生态站、生物多样性大样地、自然保护区和国家公园等, 快速构建系统性数据库, 突破该数据障碍。
7 结论植物具有多种功能性状, 且这些功能性状具有多适应途径、多功能性的特征, 彼此之间联系密切, 使植物可以维持长期生存以及适应进化过程中所面临的多种胁迫、扰动、甚至突发性灾害。在面对复杂系统时, 传统的相关分析、冗余分析、通径分析、结构方程模型等都难以揭示植物多维度适应机制。此外, 在植物环境适应与功能优化研究中, 人们先后提出了许多科学假设(如最小限制性法测、生态位分化理论、生态位互补理论、经济型谱理论等), 如何在更高维度实现多种理论的统一, 是目前生态学理论研究面临的巨大挑战。在前人研究的基础上发展的植物性状网络理论框架(PTNs), 将利用网络化思维更好地阐明多个植物功能性状之间的复杂关系。本文在先前《Trends in Ecology and Evolution》观点论文的基础上, 系统地梳理了植物功能性状网络的理论基础、构建方法、核心参数及其生态意义;同时, 结合近几年案例研究对PTNs的潜在应用领域进行了展望, 为未来探究植物大尺度的适应、群落演替、对全球变化的响应等问题提供了新的解决方案。目前如何快速建设“系统性和配套性”的新型植物功能性状数据库, 是推动PTNs理论发展和扩大应用范围的关键。
| [1] |
何念鹏, 刘聪聪, 张佳慧, 徐丽, 于贵瑞. 植物性状研究的机遇与挑战: 从器官到群落. 生态学报, 2018, 38(19): 6787-6796. |
| [2] |
He N P, Liu C C, Piao S L, Sack L, Xu L, Luo Y Q, He J S, Han X G, Zhou G S, Zhou X H, Lin Y, Yu Q, Liu S R, Sun W, Niu S L, Li S G, Zhang J H, Yu G R. Ecosystem traits linking functional traits to macroecology. Trends in Ecology & Evolution, 2019, 34(3): 200-210. |
| [3] |
Lavorel S, Garnier E. Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail. Functional Ecology, 2002, 16(5): 545-556. DOI:10.1046/j.1365-2435.2002.00664.x |
| [4] |
Violle C, Navas M L, Vile D, Kazakou E, Fortunel C, Hummel I, Garnier E. Let the concept of trait be functional!. Oikos, 2007, 116(5): 882-892. DOI:10.1111/j.0030-1299.2007.15559.x |
| [5] |
李颖. 叶片性状网络在不同植被类型间的变异规律及其影响因素[D]. 北京: 北京林业大学, 2020.
|
| [6] |
Cornelissen J H C, Lavorel S, Garnier E, Díaz S, Buchmann N, Gurvich D E, Reich P B, ter Steege H, Morgan H D, van der Heijden M G A, Pausas J G, Poorter H. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Australian Journal of Botany, 2003, 51(4): 335. DOI:10.1071/BT02124 |
| [7] |
Chave J, Coomes D, Jansen S, Lewis S L, Swenson N G, Zanne A E. Towards a worldwide wood economics spectrum. Ecology Letters, 2009, 12(4): 351-366. DOI:10.1111/j.1461-0248.2009.01285.x |
| [8] |
Díaz S, Kattge J, Cornelissen J H C, Wright I J, Lavorel S, Dray S, Reu B, Kleyer M, Wirth C, Colin Prentice I, Garnier E, Bönisch G, Westoby M, Poorter H, Reich P B, Moles A T, Dickie J, Gillison A N, Zanne A E, Chave J, Joseph Wright S, Sheremet'ev S N, Jactel H, Baraloto C, Cerabolini B, Pierce S, Shipley B, Kirkup D, Casanoves F, Joswig J S, Günther A, Falczuk V, Rüger N, Mahecha M D, Gorné L D. The global spectrum of plant form and function. Nature, 2016, 529(7585): 167-171. DOI:10.1038/nature16489 |
| [9] |
He N P, Li Y, Liu C C, Xu L, Li M X, Zhang J H, He J S, Tang Z Y, Han X G, Ye Q, Xiao C W, Yu Q, Liu S R, Sun W, Niu S L, Li S G, Sack L, Yu G R. Plant trait networks: improved resolution of the dimensionality of adaptation. Trends in Ecology & Evolution, 2020, 35(10): 908-918. |
| [10] |
Reich P B. The world-wide 'fast-slow' plant economics spectrum: a traits manifesto. Journal of Ecology, 2014, 102(2): 275-301. DOI:10.1111/1365-2745.12211 |
| [11] |
Elser J J, Fagan W F, Denno R F, Dobberfuhl D R, Folarin A, Huberty A, Interlandi S, Kilham S S, McCauley E, Schulz K L, Siemann E H, Sterner R W. Nutritional constraints in terrestrial and freshwater food webs. Nature, 2000, 408: 578-580. DOI:10.1038/35046058 |
| [12] |
Enquist B J, Brown J H, West G B. Allometric scaling of plant energetics and population density. Nature, 1998, 395: 163-165. DOI:10.1038/25977 |
| [13] |
Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavenderbares J, Chapin T, Cornelissen J H C, Diemer M. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821-827. DOI:10.1038/nature02403 |
| [14] |
Moles A T, Ackerly D D, Tweddle J C, Dickie J B, Smith R, Leishman M R, Mayfield M M, Pitman A, Wood J T, Westoby M. Global patterns in seed size. Global Ecology and Biogeography, 2007, 16(1): 109-116. DOI:10.1111/j.1466-8238.2006.00259.x |
| [15] |
Moles A T, Warton D I, Warman L, Swenson N G, Laffan S W, Zanne A E, Pitman A, Hemmings F A, Leishman M R. Global patterns in plant height. Journal of Ecology, 2009, 97(5): 923-932. DOI:10.1111/j.1365-2745.2009.01526.x |
| [16] |
Wright I J, Dong N, Maire V, Prentice I C, Westoby M, Díaz S, Gallagher R V, Jacobs B F, Kooyman R, Law E A, Leishman M R, Niinemets Ü, Reich P B, Sack L, Villar R, Wang H, Wilf P. Global climatic drivers of leaf size. Science, 2017, 357(6354): 917-921. DOI:10.1126/science.aal4760 |
| [17] |
Anderson P. Perspective: complexity theory and organization science. Organization Science, 1999, 10(3): 216-232. DOI:10.1287/orsc.10.3.216 |
| [18] |
何念鹏, 刘聪聪, 徐丽, 于贵瑞. 生态系统性状对宏生态研究的启示与挑战. 生态学报, 2020, 40(8): 2507-2522. |
| [19] |
Reich P B, Walters M B, Ellsworth D S. From tropics to tundra: global convergence in plant functioning. Proceedings of the National Academy of Sciences of the United States of America, 1997, 94(25): 13730-13734. |
| [20] |
Poorter H, Anten N P R, Marcelis L F M. Physiological mechanisms in plant growth models: do we need a supra-cellular systems biology approach?. Plant, Cell & Environment, 2013, 36(9): 1673-1690. |
| [21] |
Reich PB, Walters MB, Ellsworth DS. Leaf life-span in relation to leaf, plant, and stand characteristics among diverse ecosystems. Ecological Monographs, 1992, 62(3): 365-392. DOI:10.2307/2937116 |
| [22] |
Hetherington A M, Woodward F I. The role of stomata in sensing and driving environmental change. Nature, 2003, 424: 901-908. DOI:10.1038/nature01843 |
| [23] |
Sack L, Buckley T N. Trait multi-functionality in plant stress response. Integrative and Comparative Biology, 2020, 60(1): 98-112. DOI:10.1093/icb/icz152 |
| [24] |
毛伟, 李玉霖, 张铜会, 赵学勇, 黄迎新, 宋琳琳. 不同尺度生态学中植物叶性状研究概述. 中国沙漠, 2012, 32(1): 33-41. |
| [25] |
Ackerly D. Functional strategies of chaparral shrubs in relation to seasonal water deficit and disturbance. Ecological Monographs, 2004, 74(1): 25-44. DOI:10.1890/03-4022 |
| [26] |
Bruelheide H, Dengler J, Purschke O, Lenoir J, Jiménez-Alfaro B, Hennekens S M, Botta-Dukát Z, Chytrý M, Field R, Jansen F, Kattge J, Pillar V D, Schrodt F, Mahecha M D, Peet R K, Sandel B, van Bodegom P, Altman J, Alvarez-Dávila E, Arfin Khan M A S, Attorre F, Aubin I, Baraloto C, Barroso J G, Bauters M, Bergmeier E, Biurrun I, Bjorkman A D, Blonder B, Čarni A, Cayuela L, Černý T, Cornelissen J H C, Craven D, Dainese M, Derroire G, De Sanctis M, Díaz S, Doležal J, Farfan-Rios W, Feldpausch T R, Fenton N J, Garnier E, Guerin G R, Gutiérrez A G, Haider S, Hattab T, Henry G, Hérault B, Higuchi P, Hölzel N, Homeier J, Jentsch A, Jürgens N, Kącki Z, Karger D N, Kessler M, Kleyer M, Knollová I, Korolyuk A Y, Kühn I, Laughlin D C, Lens F, Loos J, Louault F, Lyubenova M I, Malhi Y, Marcenò C, Mencuccini M, Müller J V, Munzinger J, Myers-Smith I H, Neill D A, Niinemets Ü, Orwin K H, Ozinga W A, Penuelas J, Pérez-Haase A, Petřík P, Phillips O L, Pärtel M, Reich P B, Römermann C, Rodrigues A V, Sabatini F M, Sardans J, Schmidt M, Seidler G, Silva Espejo J E, Silveira M, Smyth A, Sporbert M, Svenning J C, Tang Z Y, Thomas R, Tsiripidis I, Vassilev K, Violle C, Virtanen R, Weiher E, Welk E, Wesche K, Winter M, Wirth C, Jandt U. Global trait-environment relationships of plant communities. Nature Ecology & Evolution, 2018, 2: 1906-1917. |
| [27] |
de la Riva E G, Olmo M, Poorter H, Ubera J L, Villar R. Leaf mass per area (LMA) and its relationship with leaf structure and anatomy in 34 Mediterranean woody species along a water availability gradient. PLoS One, 2016, 11(2): e0148788. DOI:10.1371/journal.pone.0148788 |
| [28] |
Yin Q L, Wang L, Lei M L, Dang H, Quan J X, Tian T T, Chai Y F, Yue M. The relationships between leaf economics and hydraulic traits of woody plants depend on water availability. The Science of the Total Environment, 2018, 621: 245-252. DOI:10.1016/j.scitotenv.2017.11.171 |
| [29] |
Freschet G T, Cornelissen J H C, Van Logtestijn R S P, Aerts R. Evidence of the 'plant economics spectrum' in a subarctic flora. Journal of Ecology, 2010, 98(2): 362-373. DOI:10.1111/j.1365-2745.2009.01615.x |
| [30] |
Roumet C, Birouste M, Picon-Cochard C, Ghestem M, Osman N, Vrignon-Brenas S, Cao K F, Stokes A. Root structure-function relationships in 74 species: evidence of a root economics spectrum related to carbon economy. New Phytologist, 2016, 210(3): 815-826. DOI:10.1111/nph.13828 |
| [31] |
Sack L, Cowan P D, Jaikumar N, Holbrook N M. The 'hydrology' of leaves: co-ordination of structure and function in temperate woody species. Plant, Cell & Environment, 2003, 26(8): 1343-1356. |
| [32] |
Sack L, Scoffoni C, John G P, Poorter H, Mason C M, Mendez-Alonzo R, Donovan L A. How do leaf veins influence the worldwide leaf economic spectrum? Review and synthesis. Journal of Experimental Botany, 2013, 64(13): 4053-4080. DOI:10.1093/jxb/ert316 |
| [33] |
Wright J P, Sutton-Grier A. Does the leaf economic spectrum hold within local species pools across varying environmental conditions?. Functional Ecology, 2012, 26(6): 1390-1398. DOI:10.1111/1365-2435.12001 |
| [34] |
Barabasi A L, Albert R. Emergence of scaling in random networks. Science, 1999, 286(5439): 509-512. DOI:10.1126/science.286.5439.509 |
| [35] |
Strogatz S H. Exploring complex networks. Nature, 2001, 410: 268-276. DOI:10.1038/35065725 |
| [36] |
Watts D J, Strogatz S H. Collective dynamics of 'small-world' networks. Nature, 1998, 393: 440-442. DOI:10.1038/30918 |
| [37] |
Liljeros F, Edling C R, Amaral L A N, Stanley H E, Åberg Y. The web of human sexual contacts. Nature, 2001, 411: 907-908. DOI:10.1038/35082140 |
| [38] |
Fleischmann R D, Adams M D, White O, Clayton R A, Kirkness E F, Kerlavage A R, Bult C J, Tomb J F, Dougherty B A, Merrick J M. Whole-genome random sequencing and assembly of Haemophilus influenzae Rd. Science, 1995, 269(5223): 496-512. DOI:10.1126/science.7542800 |
| [39] |
Guimerà R, Nunes Amaral L A. Functional cartography of complex metabolic networks. Nature, 2005, 433: 895-900. DOI:10.1038/nature03288 |
| [40] |
Wang C J, Wang J E. Spatial pattern of the global shipping network and its hub-and-spoke system. Research in Transportation Economics, 2011, 32(1): 54-63. DOI:10.1016/j.retrec.2011.06.010 |
| [41] |
Wang J E, Mo H H, Wang F H, Jin F J. Exploring the network structure and nodal centrality of China's air transport network: a complex network approach. Journal of Transport Geography, 2011, 19(4): 712-721. DOI:10.1016/j.jtrangeo.2010.08.012 |
| [42] |
Monaco A, Monda A, Amoroso N, Bertolino A, Blasi G, Di Carlo P, Papalino M, Pergola G, Tangaro S, Bellotti R. A complex network approach reveals a pivotal substructure of genes linked to schizophrenia. PLoS One, 2018, 13(1): e0190110. DOI:10.1371/journal.pone.0190110 |
| [43] |
Wang S, Wang X B, Han X G, Deng Y. Higher precipitation strengthens the microbial interactions in semi-arid grassland soils. Global Ecology and Biogeography, 2018, 27(5): 570-580. DOI:10.1111/geb.12718 |
| [44] |
Feng K, Peng X, Zhang Z, Gu S S, He Q, Shen W L, Wang Z J, Wang D R, Hu Q L, Li Y, Wang S, Deng Y. iNAP: an integrated network analysis pipeline for microbiome studies. iMeta, 2022, 1(2): e13. DOI:10.1002/imt2.13 |
| [45] |
Mongiovì M, Sharan R. Global alignment of protein-protein interaction networks. Methods in Molecular Biology, 2013, 939: 21-34. |
| [46] |
El Gamal A A, Kim Y H. Network information theory. Cambridge, UK: Cambridge University Press, 2011.
|
| [47] |
Proulx S R, Promislow D E L, Phillips P C. Network thinking in ecology and evolution. Trends in Ecology & Evolution, 2005, 20(6): 345-353. |
| [48] |
Osnas J L D, Katabuchi M, Kitajima K, Wright S J, Reich P B, Van Bael S A, Kraft N J B, Samaniego M J, Pacala S W, Lichstein J W. Divergent drivers of leaf trait variation within species, among species, and among functional groups. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(21): 5480-5485. |
| [49] |
Kleyer M, Trinogga J, Cebrián-Piqueras M A, Trenkamp A, Fløjgaard C, Ejrnæs R, Bouma T J, Minden V, Maier M, Mantilla-Contreras J, Albach D C, Blasius B. Trait correlation network analysis identifies biomass allocation traits and stem specific length as hub traits in herbaceous perennial plants. Journal of Ecology, 2019, 107(2): 829-842. DOI:10.1111/1365-2745.13066 |
| [50] |
Mason C M, Donovan L A. Evolution of the leaf economics spectrum in herbs: evidence from environmental divergences in leaf physiology across Helianthus (Asteraceae). Evolution, 2015, 69(10): 2705-2720. DOI:10.1111/evo.12768 |
| [51] |
Niinemets Ü, Sack L. Structural determinants of leaf light-harvesting capacity and photosynthetic potentials. Progress in Botany. Berlin/Heidelberg: Springer-Verlag, 2006: 385-419.
|
| [52] |
Poorter H, Lambers H, Evans J R. Trait correlation networks: a whole-plant perspective on the recently criticized leaf economic spectrum. New Phytologist, 2014, 201(2): 378-382. DOI:10.1111/nph.12547 |
| [53] |
Schneider J V, Habersetzer J, Rabenstein R, Wesenberg J, Wesche K, Zizka G. Water supply and demand remain coordinated during breakdown of the global scaling relationship between leaf size and major vein density. New Phytologist, 2017, 214(1): 473-486. DOI:10.1111/nph.14382 |
| [54] |
Flores-Moreno H, Fazayeli F, Banerjee A, Datta A, Kattge J, Butler E E, Atkin O K, Wythers K, Chen M, Anand M, Bahn M, Byun C, Cornelissen J H C, Craine J, Gonzalez-Melo A, Hattingh W N, Jansen S, Kraft N J B, Kramer K, Laughlin D C, Minden V, Niinemets Ü, Onipchenko V, Peñuelas J, Soudzilovskaia N A, Dalrymple R L, Reich P B. Robustness of trait connections across environmental gradients and growth forms. Global Ecology and Biogeography, 2019, 28(12): 1806-1826. DOI:10.1111/geb.12996 |
| [55] |
Oliveras I, Bentley L, Fyllas N M, Gvozdevaite A, Shenkin A F, Peprah T, Morandi P, Peixoto K S, Boakye M, Adu-Bredu S, Schwantes Marimon B, Marimon B H Jr, Salinas N, Martin R, Asner G, Díaz S, Enquist B J, Malhi Y. The influence of taxonomy and environment on leaf trait variation along tropical abiotic gradients. Frontiers in Forests and Global Change, 2020, 3: 18. DOI:10.3389/ffgc.2020.00018 |
| [56] |
Yan Z B, Hou X H, Han W X, Ma S H, Shen H H, Guo Y L, Fang J Y. Effects of nitrogen and phosphorus supply on stoichiometry of six elements in leaves of Arabidopsis thaliana. Annals of Botany, 2019, 123(3): 441-450. DOI:10.1093/aob/mcy169 |
| [57] |
Liebig J, Playfair L P. Organic chemistry in its applications to agriculture and physiology. London: Printed for Taylor and Walton, 1840.
|
| [58] |
Sterner R W, Elser J J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere. Princeton, NJ: Princeton University Press, 2003.
|
| [59] |
Li Y, Liu C C, Sack L, Xu L, Li M X, Zhang J H, He N P. Leaf trait network architecture shifts with species-richness and climate across forests at continental scale. Ecology Letters, 2022, 25(6): 1442-1457. DOI:10.1111/ele.14009 |
| [60] |
Shipley B, Lechowicz M J, Wright I, Reich P B. Fundamental trade-offs generating the worldwide leaf economics spectrum. Ecology, 2006, 87(3): 535-541. DOI:10.1890/05-1051 |
| [61] |
Li L, McCormack M L, Ma C G, Kong D L, Zhang Q, Chen X Y, Zeng H, Niinemets Ü, Guo D L. Leaf economics and hydraulic traits are decoupled in five species-rich tropical-subtropical forests. Ecology Letters, 2015, 18(9): 899-906. DOI:10.1111/ele.12466 |
| [62] |
Murren C J. Phenotypic integration in plants. Plant Species Biology, 2002, 17(2/3): 89-99. |
| [63] |
Sack L, Scoffoni C. Leaf venation: structure, function, development, evolution, ecology and applications in the past, present and future. New Phytologist, 2013, 198(4): 983-1000. DOI:10.1111/nph.12253 |
| [64] |
Li Y, Liu C C, Xu L, Li M X, Zhang J H, He N P. Leaf trait networks based on global data: representing variation and adaptation in plants. Frontiers in Plant Science, 2021, 12: 710530. DOI:10.3389/fpls.2021.710530 |
| [65] |
Medina-Vega J A, Bongers F, Poorter L, Schnitzer S A, Sterck F J. Lianas have more acquisitive traits than trees in a dry but not in a wet forest. Journal of Ecology, 2021, 109(6): 2367-2384. DOI:10.1111/1365-2745.13644 |
| [66] |
Liu C, Li Y, He N. Differential adaptation of lianas and trees in wet and dry forests revealed by trait correlation networks. Ecological Indicators, 2022, 135: 108564. DOI:10.1016/j.ecolind.2022.108564 |
| [67] |
Zhang J H, Ren T T, Yang J J, Xu L, Li M X, Zhang Y H, Han X G, He N P. Leaf multi-element network reveals the change of species dominance under nitrogen deposition. Frontiers in Plant Science, 2021, 12: 580340. DOI:10.3389/fpls.2021.580340 |
| [68] |
Kattge J, Bönisch G, Díaz S, et al. TRY plant trait database-enhanced coverage and open access. Global Change Biology, 2020, 26(1): 119-188. DOI:10.1111/gcb.14904 |