 2024, Vol. 44
2024, Vol. 44文章信息
- 田新民, 汲旭阳, 王鑫雪, 张子栋, 陈红, 刘小慧, 刘磊, 王启蕃, 马雨含, 钱昕玥, 周绍春, 鞠丹
- TIAN Xinmin, JI Xuyang, WANG Xinxue, ZHANG Zidong, CHEN Hong, LIU Xiaohui, LIU Lei, WANG Qifan, MA Yuhan, QIAN Xinyue, ZHOU Shaochun, JU Dan
- 北极村国家级自然保护区狍栖息地适宜性及活动节律
- Habitat suitability and activity patterns of roe deer in Beijicun National Nature Reserve, Heilongjiang Province, China
- 生态学报. 2024, 44(18): 8175-8184
- Acta Ecologica Sinica. 2024, 44(18): 8175-8184
- http://dx.doi.org/10.20103/j.stxb.202401080057
-
文章历史
- 收稿日期: 2024-01-08
- 网络出版日期: 2024-07-12






2. 黑龙江省野生动物研究所, 哈尔滨 150081;
3. 东北林业大学野生动物与自然保护地学院, 哈尔滨 150040
2. Wildlife Research Institute of Heilongjiang Province, Harbin 150081, China;
3. College of Wildlife and Nature Reserves, Northeast forestry University, Harbin 150040, China
栖息地是野生动物进行生命活动的场所, 对其种群的生存繁衍具有重要意义[1]。随着人类活动范围的不断扩张, 野生动物栖息地呈破碎化, 适宜栖息地面积不断减少[2]。适宜栖息地的缩减已导致许多珍稀野生动物种群数量下降, 甚至在一些区域濒临灭绝[3]。为了保护濒临灭绝的野生动物种群, 野生动物研究人员开展了大量的栖息地适宜性评价工作。至今, 有多种方法被用于栖息地适宜性评价[4—7]。其中, 最大熵模型因其只需物种分布的位置信息和环境数据便可以准确分析物种栖息地分布情况, 具有操作简便、所需样本量少、准确度高等优点被广泛应用于生态学领域研究[8—9]。
探讨动物活动节律是开展野生动物保护管理的主要内容之一。动物的活动节律是动物对昼夜环境变化长期适应的结果[10]。动物在不同时间、季节下的活动强度及变化规律是活动节律研究的核心, 也是行为生态学重要研究内容[11]。动物活动节律除了受到个体差异的直接影响外, 也会受到其他因素的综合影响, 例如:食物[12]、气候条件[13]及人为干扰因素[14]。为此, 研究动物活动节律的变化特征对于了解动物生存状况、种间关系以及针对性制定合理有效的保护措施具有重要意义[15]。
狍(Capreolus pygargus)广泛分布于整个欧亚大陆, 有关狍时空分布方面在国外已有广泛研究, 例如:De[16]等发现分布在欧洲地区的狍对于不同类型栖息地的利用具有明显交替现象;Bonnot[17]等对欧洲北部狍种群的活动节律研究发现, 狍的活动模式具有明显的可塑性, 会根据人类活动和捕食者的密度改变其活动节律。狍作为可塑性较强的物种, 面对地理位置及环境差异, 在栖息地选择及活动模式上表现出不同的对策。近年来, 国内对于狍种群的研究也有许多[18—19], 但北极村国家级自然保护区作为分布在中国最北端的保护区, 有关狍栖息地适宜性及活动节律方面的研究未见报道。因此, 本研究选择保护区内的狍种群作为研究对象, 探究该地区影响狍栖息地选择的环境因子及活动节律的变化特征, 掌握保护区内狍的栖息地适宜性及活动特点, 同时对比纬度更高地区或国家狍种群栖息地选择及活动节律情况, 以期为保护区对该物种的保护管理提供更准确的信息, 为科学、有效、合理制定该物种的保护监测计划服务。
1 研究地区北极村国家级自然保护区位于黑龙江省大兴安岭山地北部(图 1), 地理坐标为53°11′30″—53°33′03″N, 121°40′00″—123°16′00″E, 总面积1375.53 km2, 隶属黑龙江省大兴安岭地区漠河市。保护区属于寒温带大陆性季风气候, 年平均气温为-5℃左右, 与分布在黑龙江省其他地区的保护区相比, 北极村国家级自然保护区冬季持续时间更长, 可达7个月以上, 且日照时间非常短, 夏季较短, 然而从每年的6—8月份, 日照时间长达17 h, 年平均降水量为460.8 mm, 降水主要集中于7、8月份, 占全年降水量的24%左右, 平均无霜期86.2 d, 海拔范围为191—848 m之间。保护区内森林资源丰富, 树种以兴安落叶松(Larix gmelinii)、樟子松(Pinus sylvestris)、白桦(Betula platyphylla)为主, 针叶林占绝对优势, 独特的气候条件和植被资源形成了典型的水域、沼泽、草甸、灌丛和原始森林植被。丰富的植被资源为东北虎(Panthera tigris altaica)、紫貂(Martes zibellina)、猞猁(Lynx lynx)、马鹿(Cervus elaphus)、狍等多种珍稀野生动物提供了栖息条件。

|
| 图 1 北极村国家级自然保护区样线、相机监测布设示意图 Fig. 1 Location of line transect and camera trap in Beijicun National Nature Reserve |
相机布设:于2022年6月—2023年5月累计布设140台红外相机进行监测, 采用随机布设方法, 每个相机位点布设1—2台相机, 相机位点之间至少大于1 km。为了避免相机因晃动产生误拍, 相机均固定在胸径大于16 cm的树干上, 且距离地面约0.5 m。相机设置为拍摄模式(拍照+录像)[20]。相机监测期间换卡3次, 布设相机与换卡时注意清理镜头前方5 m左右30°范围内的杂草与灌木。
样线调查:利用卫星遥感影像图片识别森林覆盖区域, 根据俄罗斯远东地区有蹄类调查中获得的狍平均日行距离的研究结果[21], 在研究区域内随机布设35条样线, 每条长度为3—5 km, 样线间距大于1 km, 并确保样线覆盖保护区所有植被类型。于积雪覆盖期(2022年11月至2023年2月末)开展样线调查工作, 因保护区内除狍外还有马鹿、驼鹿(Alces alces)等有蹄类动物, 根据足迹链特征、粪便形态可对不同有蹄类动物加以区分, 若发现狍的痕迹信息(足迹、卧迹、粪便等)则记录一处活动点并记录活动点坐标。
2.1.2 活动位点数据处理及时对从红外相机获得的数据进行整理和记录。为了尽可能地排除重复记录同一个体, 根据狍的家域范围[22], 在其范围内只保留一台相机数据用于分析, 同时根据成体(雄性有角、雌性无角)、幼体数和体积大小等生物学特征对不同个体(群体)进行判断[23]。将同一相机位点时间间隔30 min以内且被认定为同一个体(群体)的照片判定为一张狍的独立照片[24], 为减少因同一个体重复获得活动位点及模型的过度拟合, 本研究在ArcGIS 10.4中采用系统痕迹点缓冲分析根据狍的平均日行距离剔除距离较近的狍活动位点, 将剩余的232个(相机监测获得79个、样线调查获得153个)狍活动位点转换成MaxEnt软件所需的CSV格式保存。
2.2 环境变量数据的获取及处理环境变量分为4类共计14个, 包括:
(1) 植被:从地理空间数据云(http://www.gscloud.cn)下载分辨率为30 m的Landsat 8 TIRS云覆盖低于5%遥感影像解译获得, 植被类型共分为:林间湿地、常绿针叶林、落叶针叶林、落叶阔叶林、针阔混交林、农田和草地。
(2) 人为干扰因子:居民区通过地理空间数据云(http://www.gscloud.cn)下载分辨率为30 m的Landsat 8 TIRS云覆盖低于5%遥感影像解译获得;公路和林间小路在ArcGIS 10.4中利用林相图矢量化获得。
(3) 地形因子:海拔、坡度和坡向利用ArcGIS 10.4软件的空间分析模块对30 m分辨率的数字高程模型(DEM)进行掩膜提取获得。
(4) 河流:在ArcGIS 10.4中利用林相图矢量化获得。
2.3 狍栖息地适宜性模型构建为了减少环境变量间存在相关性对模型预测精度产生负面影响, 本研究使用SPSS 23.0软件将14个环境变量数据标准化处理后进行Spearman相关性分析, 剔除相关系数|r|≥0.8中贡献值较小的变量。最终保留植被变量(草地、落叶阔叶林、常绿针叶林、针阔混交林)、人为干扰变量(林间小路、公路、居民区)、地形变量(海拔、坡度、坡向)及河流。
模型预测结果的准确性通过受试者工作特征曲线(Relative Operating Characteristic, ROC曲线)判断, 其中AUC值大小代表模型预测结果的精确度[25]。ROC曲线评价标准为:AUC值在0.5—0.6, 不及格;0.6—0.7, 较差;0.7—0.8, 一般;0.8—0.9, 良好;0.9—1.0, 优秀[26]。利用刀切法(Jackknife)检验环境变量对模型预测的重要性和贡献值, 并通过响应曲线分析关键变量对狍产生的影响。此外, 为了确定适宜栖息地斑块面积和分布情况, 对模型预测结果采用自然间断点分级法(Jenks′ natural breaks)进行重分类[27], 将狍适宜栖息地等级划分为不适宜栖息地(0—0.27)、次适宜栖息地(0.27—0.54)和适宜栖息地(0.54—1), 并以此结果为基础将狍适宜栖息地大于10 km2的斑块作为核心栖息地斑块, 进行核心栖息地确定。
2.4 活动节律分析因北极村气候特殊, 冬季寒冷且延续时间较长, 故将一年划分为冷季(10月—次年5月)、暖季(6月—9月)。研究使用拍摄率(capture rate, CR)量化研究区域内红外相机拍摄到狍的相对丰度。
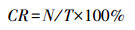
|
式中, N为拍摄到狍的总独立有效照片数, T为相机总有效工作日。拍摄率即为每100个相机工作日所获取某物种的独立有效照片数。
此外, 基于核密度估计法(kernel density estimation)比较冷季和暖季之间狍的日活动节律差异程度。使用R 4.3.1中的overlap包和activity包估算出不同季节下狍活动节律变化特征, 绘制核密度曲线图, 并计算不同季节狍活动节律的重叠指数。在计算重叠指数时, 因本研究样本量≥75, 故选用Δ4值(Δ为0表示完全分离, Δ为1表示完全重叠)进行分析[28]。当Δ>0.75时为“高度重叠”, 0.50<Δ<0.75时为“中度重叠”, Δ<0.5时为“低度重叠”[29]。
将狍的活动时间分为3个不同时间段:白天(日出后1 h至日落前1 h之间)、夜晚(日落后1 h至日出前1 h之间)和晨昏(日出前后1 h之内及日落前后1 h之内), 基于日出日落时间查询平台(https://sunrise.supfree.net/), 通过漠河市每日的日出日落时间计算出冷暖季平均日出日落时间, 统计冷暖季各时间段狍的独立有效照片数并计算选择性指数(Selection ratio index, SRI), 以此分析狍活动时间偏好[30]。
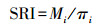
|
式中, Mi为第i个时间段拍摄到狍的独立有效照片数, πi为第i时期长度占所有时期长度的比例。当SRI>1则认为偏好利用该时间段, 反之则表示回避。
3 结果 3.1 栖息地适宜性预测模型的平均训练集AUC值为0.839±0.011, 达到良好水平, 表明模型的预测结果能够真实反映该区域狍栖息地适宜性。环境变量贡献率结果表明:河流是影响狍分布的主要变量, 贡献率为34%;针阔混交林、常绿针叶林、林间小路、海拔、草地和居民是影响狍栖息地质量的次要变量;其他环境变量贡献率均小于5%。
响应曲线分析表明(图 2), 河流作为影响狍栖息地分布最重要的变量, 狍的出现概率与距河流距离表现为负相关, 距河流越近狍出现的概率越高。此外, 分析结果显示, 植被因子在狍适宜栖息地分布中具有十分重要的作用。狍与针阔混交林之间整体呈现下降趋势, 狍距针阔混交林约2 km时出现概率最高;在距常绿针叶林约0—1 km范围内, 随着距离增加狍的出现概率逐渐上升, 大于1 km后随着距离的增加狍出现概率随之降低;狍对草地表现回避, 距草地越远出现概率越高。在林间小路和居民区附近、在海拔700—800 m之间, 狍表现出明显偏好。

|
| 图 2 主要变量响应曲线 Fig. 2 Response curves of main variables |
适宜栖息地分布图表明(图 3), 狍的适宜栖息地、次适宜栖息地和不适宜栖息地面积分别为240.44 km2、490.64 km2和644.42 km2, 分别占研究区域总面积的17.48%、35.67%和46.85%, 利用此结果共确定6块核心栖息地斑块, 总面积为162.18 km2, 占适宜栖息地的67.45%。其中适宜栖息地主要分布于保护区中部的河东林场和东部河湾林场, 西南部的前哨林场和西北部的金沟林场适宜栖息地面积较少且破碎化较为严重。

|
| 图 3 狍适宜栖息地分布图 Fig. 3 Map of the habitat suitability of roe deer |
不同季节日出日落时间存在明显差异, 暖季日出日落时间为:3∶16∶44—5∶16∶44和18∶25∶14—20∶25∶14, 冷季日出日落时间为∶5∶24∶30—7∶24∶30和16∶14∶33—18∶14∶33。研究期间累计监测22155个相机工作日, 获得狍照片1780张, 独立有效照片413张, 拍摄率为1.86%, 其中暖季白天、夜间及晨昏段所拍摄独立有效照片数分别为166张、32张和49张, 冷季白天、夜间及晨昏段所拍摄独立有效照片数分别为115张、27张和24张, 结果表明北极村国家级自然保护区狍在冷暖季活动均表现出昼行性。日活动节律分析结果表明(图 4), 狍的日活动时间集中在6:00—18:00, 其中在9:00附近为一天的最高峰。

|
| 图 4 狍日活动节律 Fig. 4 Daily activity pattern of Roe deer |
冷暖季节狍的日活动节律达到中度重叠(Δ4=0.58)。如图 5所示, 不同季节狍昼夜活动呈现“M”形, 但日活动高峰期有所差异(图 5)。暖季狍的日活动高峰期出现在5:00和18:00左右, 其中5:00附近狍的相对强度指数为当日最高。在冷季狍上午和下午的日活动高峰期表现出延迟和提前, 主要在9:00和14:00左右。

|
| 图 5 不同季节狍日活动节律 Fig. 5 Daily activity pattern of Roe deer in different seasons 阴影部分代表冷暖季节活动时间重叠指数 |
红外相机监测技术是经典的野生动物数据收集方法之一, 因其具有隐蔽性强、造成干扰小、可连续工作和受环境条件限制较小等特征被广泛应用于生态学研究中。过去几十年间关于野生动物调查工作离不开红外相机的布设。但由于研究区域在我国最北端, 冬季温度过低时将导致部分红外相机停止工作。因此, 在冬季进行调查时采用红外相机与样线调查相结合的方式会增加调查结果的准确性。
本研究表明(图 2), 狍出现概率随距河流距离增加而下降, 这是因为北极村国家级自然保护区位于我国最北端, 该地区当年11月至次年4月为河流封冻期, 冰面利于狍行走, 且河流两岸的灌丛能够为狍提供部分食物资源[18], 因此距河流较近狍出现概率较高适宜栖息地分布较多。北极村国家级自然保护区属于典型的寒温带大陆性季风气候和亚寒带针叶林带, 保护区内针叶林所占面积较大。狍偏好在距常绿针叶林、针阔混交林较近且远离草地的区域活动, 这与腾丽薇[31]等研究结果不同, 却与高纬度地区狍栖息地选择研究结果相同[32], 分析其原因为研究区域冬季常伴有大雪, 常绿针叶林郁闭度较高, 林冠截雪作用较强从而引起地面积雪结构发生变化[33], 并且林冠还能调节林下的温度及风速, 对于狍保持体温恒定减少能量消耗具有极其重要的作用[34], 因此狍偏好在靠近常绿针叶林附近活动。此外, 狍偏好在距针阔混交林较近处活动, 是因为冬季杨树、桦树等阔叶树种的枝条能为其提供食物资源[35], 同时针阔混交林产生的微环境对狍的生存具有重要作用[36], 驱使狍对针阔混交林表现出较高的偏好性。对于草地这一植被类型, 不同地区的狍表现出不同的偏好, 高纬度地区的狍种群更倾向在视野开阔的区域活动[37]而在我国其他地区有研究表明狍喜欢在隐蔽性好的生境进行活动[38], 分析其原因可能是由于地理位置不同, 高纬度地区冬季积雪较深, 草地开阔更利于狍行走, 但在本研究区域, 由于保护区内存在人类活动和捕食者存在, 不利于狍的隐蔽, 一定程度上增加其被发现和捕食风险。因此, 在各种因素的综合影响下造成狍对不同植被类型的选择差异。本研究中狍的分布倾向于选择林间小路与居民区附近, 而在挪威南部的狍种群表现出与本研究相反的结果[39], 是由于当地允许居民携带枪支且狩猎属于合法行为, 狍靠近居民区与道路时被捕杀的风险大大增加, 而研究区域内林间小路多为森林采伐期间遗留的集材道, 大兴安岭地区目前处于禁止采伐时期, 通往林间小路的岔口均被挡住, 不允许人类通行, 这为野生动物尤其有蹄类提供了良好的通行条件, 而居民区附近多为农田, 农田中残留的食物资源会吸引狍到附近觅食[18]。狍偏好在海拔700—800米范围内活动, 这与刘艳华等[18]在老爷岭南部的研究结果类似, 而与申立泉[40]在北京周边山区的研究结果有所不同。由于地理位置不同, 环境差异导致有蹄类动物对适宜栖息地的选择也具有明显差异[41], 致使狍在海拔的选择上表现不同。
狍适宜栖息地主要分布于河东林场和河湾林场, 其余区域栖息地破碎化较为严重。研究发现保护区内建设国家级森林公园, 游客和过往车辆较多、人为干扰严重, 大部分区域不适合狍种群生存, 因此相关部门应加大保护区周边人类活动的管控力度, 掌握人类活动时空特征, 分析不同人为干扰强度下狍种群时空分布特征, 从而针对性地进行保护。此外, 由于狍种群对水资源具有强烈偏好, 其适宜栖息地分布呈现条带状, 故可以考虑在保护区内进行栖息地改造, 建设人工水塘、野生动物定点饮水区等方法避免旱期影响狍种群生存。根据核心栖息地斑块情况(图 3), 保护区内存在核心栖息地孤立化, 适宜栖息地破碎化、连通性差等问题。因此, 研究认为在保持现有栖息地同时应对适宜栖息地进行保护, 避免造成不适宜栖息地的持续扩张, 通过加强栖息地连通性管理、构建栖息地廊道等方法使破碎的栖息地连接成整体, 促进保护区狍个体间交流[42]。提高狍种群遗传多样性, 从而维持北极村国家级自然保护区的生物多样性和生态平衡。
4.2 活动节律在长期的自然选择中, 不同的地理环境会形成野生动物独特的活动节律, 形成过程往往受多种因素的共同作用。研究发现, 狍在夜晚的活动频率相对较低, 这与保护区内狍的捕食者密度有关, 在捕食者少的地区, 狍在夜晚的活动强度相对较高, 当保护区内存在一定数量捕食者时, 狍的活动模式产生变化以降低被捕食的风险[43]。调查发现研究区域内存在猞猁、狼等多种狍的捕食者, 这些捕食者大多数在夜晚进行活动(表 1), 迫使狍改变其活动模式。
| 捕食者 Predator |
独立有效照片数 Number of unique valid photos |
夜晚独立有效照片数 Number of unique valid photos at night |
百分比/% Percent |
| 猞猁(Lynx lynx) | 351 | 251 | 71.51 |
| 狼 | 73 | 40 | 54.79 |
面对环境温度的变化, 动物会选择较为适宜的时间进行活动[44], 最终导致日活动节律在不同季节间出现差异。本研究中狍在不同季节活动时间段具有灵活的选择性。在暖季, 狍的两个活动高峰期出现在5:00和18:00附近, 这与江辉龙[45]和陈梦霄等[46]的研究结果相似, 狍的活动均具有一定的晨昏性。这种现象是由多种因素综合所致:(1)夜晚的气温相对较低, 早晨相对凌晨气温有所回升, 并且晨昏这两个时段恰好避免了高温和强阳光, 减少水分的散失有利于狍的活动[47];(2)晨昏时间段树的枝芽多挂有露水, 狍在取食过程中能够一定程度上补充水分;(3)北极村国家级自然保护区对人类活动具有一定的管控, 但仍存在人类、家畜、家犬等干扰, 据调查发现在白天人为干扰均保持较高强度, 而狍在晨昏活动一定程度上避开了人为干扰的影响(图 6)。而在冷季, 狍的活动高峰期表现为上午的延迟和下午的提前, 产生这种现象的原因为:(1)在冷季夜晚和晨昏温度低、食物资源质量较差, 而中午相较其他时间段温度高, 为了减少维持生命活动所消耗的能量, 狍会选择较为温暖的时间活动;(2)冷季因气候影响, 具有经济价值的山货极少, 因此没有人进山采山货, 人为干扰产生的影响较小, 也为狍集中在中午附近活动提供了条件。而不同地区或国家狍的日活动节律表现出相似特征, 在高纬度地区在较为温暖的季节, 狍表现出明显的晨昏性[48], 温度的下降同样导致狍活动高峰期向一天中温度较高的时间段靠近[49], 此外, 分布在欧洲北部地区的狍种群, 当人类活动密度高时狍在白天活动频率也存在明显下降[17]。

|
| 图 6 暖季狍和人为干扰相对强度指数 Fig. 6 Relative intensity index of roe deer and human disturbance in warm season |
通过以上研究发现, 狍在纬度更高的地区通常会选择具有茂密树冠的针叶林作为其栖息地, 而在气候较为温暖的地区, 阔叶林中的丰富的食物是其首要选择, 同时发现无论在任何林型狍都会根据当地捕食者密度、人为干扰类型及强度等适当调整其适宜栖息地选择。此外, 狍的活动节律会根据地理位置及环境发生改变后进行针对性调整, 高纬度地区的狍, 由于冷暖季节温度差异较大, 其活动节律的高峰期变化较为明显, 而在低纬度地区狍的活动节律主要表现出晨昏性, 并且无论纬度高低, 狍在白天活动强度均高于夜晚。正是这种高度的适应能力为狍种群的繁衍生存提供重要保障。
| [1] |
Grinnell J. Field tests of theories concerning distributional control. The American Naturalist, 1917, 51(602): 115-128. DOI:10.1086/279591 |
| [2] |
Ramírez-Delgado J P, Di Marco M, Watson J E M, Johnson C J, Rondinini C, Corredor Llano X, Arias M, Venter O. Matrix condition mediates the effects of habitat fragmentation on species extinction risk. Nature Communications, 2022, 13(1): 595. DOI:10.1038/s41467-022-28270-3 |
| [3] |
姜广顺, 李京芝. 中国濒危兽类栖息地评估与保护研究进展与展望. 兽类学报, 2021, 41(5): 604-613. |
| [4] |
刘玮, 辛美丽, 周健, 詹冬梅, 丁刚, 吴海一. 基于生境适宜性指数模型的俚岛海黍子生境层级分布. 应用生态学报, 2021, 32(3): 1061-1068. |
| [5] |
刘鹏, 代娟, 曹大藩, 李志宏, 张立. 普洱市亚洲象栖息地适宜度评价. 生态学报, 2016, 36(13): 4163-4170. DOI:10.5846/stxb201411142260 |
| [6] |
张学儒, 周杰, 李梦梅. 基于土地利用格局重建的区域生境质量时空变化分析. 地理学报, 2020, 75(1): 160-178. |
| [7] |
王启蕃, 陈红, 牛莹莹, 张子栋, 梁卓, 田新民, 张明海, 周绍春. 黑龙江省老爷岭南部东北梅花鹿栖息地适宜性评价及廊道构建. 兽类学报, 2023, 43(5): 489-500. |
| [8] |
Mahmoodi S, Ahmadi K, Shabani A A, Zeinalabedini M, Javanmard A, Khalilipour O, Banabazi M H. Anthropogenic and natural fragmentations shape the spatial distribution and genetic diversity of roe Deer in the marginal area of its geographic range. Ecological Indicators, 2023, 154: 110835. DOI:10.1016/j.ecolind.2023.110835 |
| [9] |
滕扬, 张沼, 张书理, 杨永昕, 贺伟, 王娜, 张正一, 鲍伟东. 大兴安岭南段马鹿生境适宜性分析与生态廊道构建. 生态学报, 2022, 42(14): 5990-6000. DOI:10.5846/stxb202109062517 |
| [10] |
Seri H, Chammem M, Ferreira L M, Kechnebou M, Khorchani T, Silva S R. Effects of seasonal variation, group size and sex on the activity budget and diet composition of the addax antelope. African Journal of Range & Forage Science, 2018, 35(2): 89-100. |
| [11] |
唐卓. 利用红外相机研究卧龙雪豹和绿尾虹雉及其同域野生动物[D]. 北京: 清华大学, 2016.
|
| [12] |
Gaudiano L, Pucciarelli L, Mori E. Livestock grazing affects movements and activity pattern of Italian roe Deer in Southern Italy. European Journal of Wildlife Research, 2021, 67(4): 66. DOI:10.1007/s10344-021-01506-1 |
| [13] |
Liu S Y, Qi J, Gu J, Long Z, Ma J Z, Jiang G S. What factors relate with the activity synchronization intensity among big cats and their ungulate prey in Northeast China?. Global Ecology and Conservation, 2021, 32: e01899. DOI:10.1016/j.gecco.2021.e01899 |
| [14] |
Petridou M, Benson J F, Gimenez O, Kati V. Spatiotemporal patterns of wolves, and sympatric predators and prey relative to human disturbance in northwestern Greece. Diversity, 2023, 15(2): 184. DOI:10.3390/d15020184 |
| [15] |
Bu H L, Wang F, McShea W J, Lu Z, Wang D J, Li S. Spatial co-occurrence and activity patterns of mesocarnivores in the temperate forests of southwest China. PLoS One, 2016, 11(10): e0164271. DOI:10.1371/journal.pone.0164271 |
| [16] |
De Groeve J, Van de Weghe N, Ranc N, Morellet N, Bonnot N C, Gehr B, Heurich M, Mark Hewison A J, Kröschel M, Linnell J D C, Mysterud A, Sandfort R, Cagnacci F. Back and forth: day-night alternation between cover types reveals complementary use of habitats in a large herbivore. Landscape Ecology, 2023, 38(4): 1033-1049. DOI:10.1007/s10980-023-01594-1 |
| [17] |
Bonnot N C, Couriot O, Berger A, Cagnacci F, Ciuti S, De Groeve J E, Gehr B, Heurich M, Kjellander P, Kröschel M, Morellet N, Sönnichsen L, Hewison A J M. Fear of the dark? Contrasting impacts of humans versus lynx on diel activity of roe Deer across Europe. The Journal of Animal Ecology, 2020, 89(1): 132-145. DOI:10.1111/1365-2656.13161 |
| [18] |
刘艳华, 牛莹莹, 周绍春, 张子栋, 梁卓, 杨娇, 鞠丹. 老爷岭南部狍冬季移动、卧息生境选择及其适宜性评价. 生态学报, 2021, 41(17): 6913-6923. DOI:10.5846/stxb202007131827 |
| [19] |
Bao H, Zhai P H, Wen D, Zhang W H, Li Y, Yang F F, Liang X, Yang F, Roberts N, Xu Y C, Jiang G S. Effects of inter- and intra-specific interactions on moose habitat selection limited by temperature. Remote Sens, 2022, 14: 6401. DOI:10.3390/rs14246401 |
| [20] |
田新民, 刘小慧, 张超宇, 张宇, 张子栋, 梁卓, 陈红, 宋雅祺, 王鑫雪, 周绍春. 黑龙江省老爷岭南部黄喉貂栖息地适宜性评价与廊道构建. 生态学报, 2023, 43(23): 9621-9629. DOI:10.20103/j.stxb.202211013120 |
| [21] |
Stephens P A, Zaumyslova O Y, Miquelle D G, Myslenkov A I, Hayward G D. Estimating population density from indirect sign: track counts and the Formozov-Malyshev-Pereleshin formula. Animal Conservation, 2006, 9(3): 339-348. |
| [22] |
张帅. 存在猞猁捕食风险下西伯利亚狍的家域与生境利用研究[D]. 北京: 北京林业大学, 2014.
|
| [23] |
马逸清. 黑龙江省兽类志. 哈尔滨: 黑龙江科学技术出版社, 1986: 407-412.
|
| [24] |
刘周, 周虎, 郭瑞, 章书声, 许丽娟, 罗远, 徐爱春. 浙江清凉峰国家级自然保护区华南梅花鹿栖息地内人为干扰类型及时空分布格局. 兽类学报, 2020, 40(4): 355-363. |
| [25] |
许仲林, 彭焕华, 彭守璋. 物种分布模型的发展及评价方法. 生态学报, 2015, 35(2): 557-567. DOI:10.5846/stxb201304030600 |
| [26] |
Swets J A. Measuring the accuracy of diagnostic systems. Science, 1988, 240(4857): 1285-1293. |
| [27] |
刘轲, 刘炎林, 李晟. 欧亚猞猁在中国的分布现状与适宜栖息地预测. 兽类学报, 2023, 43(6): 652-663. |
| [28] |
Ridout M S, Linkie M. Estimating overlap of daily activity patterns from camera trap data. Journal of Agricultural, Biological, and Environmental Statistics, 2009, 14(3): 322-337. |
| [29] |
江辉龙, 周绍春, 张明海, 徐万鹏, 李薇, 刘静岩. 同域分布的野猪与狍活动节律及与月光周期的关系. 生态学报, 2023, 43(8): 3128-3136. DOI:10.5846/stxb202203100582 |
| [30] |
Li J, Xue Y D, Liao M F, Dong W, Wu B, Li D Q. Temporal and spatial activity patterns of sympatric wild ungulates in Qinling Mountains, China. Animals: an Open Access Journal from MDPI, 2022, 12(13): 1666. |
| [31] |
滕丽微, 刘振生, 张恩迪, 马建章. 黑龙江省三江自然保护区狍对冬季卧息地的选择. 动物学研究, 2006, 27(4): 403-410. |
| [32] |
李奕, 蔡体久, 盛后财, 俞正祥. 大兴安岭地区天然樟子松林降雪截留及积雪特征. 水土保持学报, 2014, 28(5): 124-128. |
| [33] |
Borkowski J, Ukalska J. Winter habitat use by red and roe Deer in pine-dominated forest. Forest Ecology and Management, 2007, 255(3): 468-475. |
| [34] |
Ossi F, Gaillard J M, Hebblewhite M, Cagnacci F. Snow sinking depth and forest canopy drive winter resource selection more than supplemental feeding in an alpine population of roe Deer. European Journal of Wildlife Research, 2015, 61(1): 111-124. |
| [35] |
周绍春, 张明海, 尹远新, 任梦非. 黑龙江完达山地区狍子冬季生境选择. 北京林业大学学报, 2010, 32(3): 122-127. |
| [36] |
Jean H. Winter habitat of white-tailed Deer at Thirty-one Mile Lake, Quebec. The Canadian Field-Naturalist, 1974, 88(3): 293-301. |
| [37] |
Mysterud A, Bjørnsen B H, Østbye E. Effects of snow depth on food and habitat selection by roe Deer Capreolus capreolus along an altitudinal gradient in south-central Norway. Wildlife Biology, 1997, 3(1): 27-33. |
| [38] |
陈龙, 李月辉, 胡远满, 熊在平, 吴文, 李悦, 问青春. 小兴安岭铁力林业局冬季西伯利亚狍(Capreolus pygargus)的生境选择. 生物多样性, 2017, 25(4): 401-408. |
| [39] |
Mysterud A, Lian L B, Hjermann D Ø. Scale-dependent trade-offs in foraging by European roe Deer (Capreolus capreolus) during winter. Canadian Journal of Zoology, 1999, 77(9): 1486-1493. |
| [40] |
申立泉, 吴佳忆, 周鑫, 吕青昕, 王燕群, 袁乃秀, 孟秀祥. 基于MaxEnt模型的北京周边山区野生狍(Capreolus pygargus)的生境适宜性评价. 生态学杂志, 2023, 42(10): 2555-2560. |
| [41] |
顾佳音, 王巍, 郭晓艺, 姜广顺. 小兴安岭南麓野猪春冬季行为状态的生境选择. 生态学报, 2024, 44(3): 1067-1078. DOI:10.20103/j.stxb.202303180510 |
| [42] |
刘艳华, 刘磊, 陈红, 刘小慧, 王启蕃, 刘鑫鑫, 张子栋, 张宇, 周绍春. 基于MaxEnt模型的北极村国家级自然保护区紫貂栖息地适宜性评价. 生态学报, 2024, 44(2): 559-569. DOI:10.20103/j.stxb.202301010005 |
| [43] |
Mori E, Bagnato S, Serroni P, Sangiuliano A, Rotondaro F, Marchianò V, Cascini V, Poerio L, Ferretti F. Spatiotemporal mechanisms of coexistence in an European mammal community in a protected area of southern Italy. Journal of Zoology, 2020, 310(3): 232-245. |
| [44] |
Wolfson D W, Schlichting P E, Boughton R K, Miller R S, VerCauteren K C, Lewis J S. Comparison of daily activity patterns across seasons using GPS telemetry and camera trap data for a widespread mammal. Ecosphere, 2023, 14(12): e4728. |
| [45] |
江辉龙, 周绍春, 张明海, 徐万鹏, 李薇, 刘静岩. 同域分布的野猪与狍活动节律及与月光周期的关系. 生态学报, 2023, 43(8): 3128-3136. DOI:10.5846/stxb202203100582 |
| [46] |
陈梦霄, 王晓玲, 穆丽光, 姜秀丽, 胡磊, 乌力吉, 王翠, 孟和达来, 鲍伟东. 内蒙古赛罕乌拉国家级自然保护区狍的日活动节律及其季节变化研究. 四川动物, 2019, 38(2): 214-219. |
| [47] |
李耀宇, 陈昕旸, 赵闪闪, 宋虓, 郭瑞, 徐爱春. 野外放归华南梅花鹿活动节律与家域特征. 兽类学报, 2023, 43(5): 501-512. |
| [48] |
Jasińska K D, Jackowiak M, Gryz J, Bijak S, Szyc K, Krauze-Gryz D. Habitat-related differences in winter presence and spring-summer activity of roe Deer in warsaw. Forests, 2021, 12(8): 970. |
| [49] |
Pagon N, Grignolio S, Pipia A, Bongi P, Bertolucci C, Apollonio M. Seasonal variation of activity patterns in roe Deer in a temperate forested area. Chronobiology International, 2013, 30(6): 772-785. |