 2024, Vol. 44
2024, Vol. 44文章信息
- 吴小健, 李江飞, 江宇, 孙麟钧, 吴鹏飞, 马祥庆
- WU Xiaojian, LI Jiangfei, JIANG Yu, SUN Linjun, WU Pengfei, MA Xiangqing
- 增温对杉木凋落叶和宿存叶分解过程养分释放的影响——基于移位试验模拟
- Effect of simulated warming on nutrient release during the decomposition of leaf litter and canopy litter of Chinese fir based on displacement test
- 生态学报. 2024, 44(17): 7713-7724
- Acta Ecologica Sinica. 2024, 44(17): 7713-7724
- http://dx.doi.org/10.20103/j.stxb.202311122451
-
文章历史
- 收稿日期: 2023-11-12
- 网络出版日期: 2024-06-27





2. 国家林业和草原局杉木工程技术研究中心, 福州 350002
2. Chinese Fir Engineering Technology Research Center of National Forestry and Grassland Administration, Fuzhou 350002, China
全球气温变化日趋严重, 对人类生存环境造成极大的威胁[1]。IPCC AR6报告指出, 2021—2040年全球平均气温相较于2001—2020年将升高0.3—0.7 ℃[2]。全球尺度上的气温升高不仅造成陆地生态系统中植被生长策略及分布格局的改变, 还会引起地上与地下各碳库间的失衡, 对陆地生态系统的生产力和生态功能产生深远影响[3]。凋落物作为陆地生态系统中能量流动和养分循环的重要载体[4], 其分解过程中的养分释放不仅决定着土壤养分状况和林木生长策略, 在调节森林生态系统养分平衡以及维持林分长期生产力等方面也扮演着重要角色[5]。因此, 开展模拟增温对森林凋落物分解的影响研究, 对掌握及预测未来森林生态系统的养分循环过程具有重要意义。
杉木(Cunninghamia lanceolata (Lamb.) Hook.)是我国重要的速生用材树种, 具有生长快、病害少和材质好等优点, 广泛分布于我国秦岭淮河以南的17个省区[6]。近年来杉木人工林多代连栽导致林地生产力下降问题日益严重, 极大地限制了杉木人工林的可持续经营发展。研究发现, 杉木人工林凋落物产量低、分解速率慢的特性可能是导致连栽生产力下降的原因之一[7], 且杉木枝叶枯死过程中存在的宿存特性, 致使其枯死后不会立刻掉落地面, 依然会附着在主干上存留四年之久[8]。杉木枝叶的大量宿存极大地延缓了人工林生态系统的养分归还, 且不同发育阶段杉木林枯死枝叶宿存量存在明显差异。郑临训[9]研究发现, 在15年生杉木人工林中, 宿存枝叶占林分枯死枝叶总量的50%以上, 且所占比重随地位指数的增加呈上升趋势。因此, 宿存枝叶是杉木人工林生态系统中不可忽视的重要组成部分。但目前有关杉木人工林枯死叶片的研究多集中于地面凋落叶[10], 忽略了林冠下层的宿存叶, 而针对杉木凋落叶和宿存叶分解对增温的响应研究更鲜有报道。因此, 开展增温对杉木人工林凋落叶和宿存叶分解过程中养分释放的影响研究显得极为必要, 有助于全面地深入认识杉木人工林的养分循环过程。
近年来, 针对增温对森林凋落物分解的影响研究多以室内微缩模拟(培养箱、温室)和原位模拟(埋设电缆、开顶式增温箱)等人为控制试验为主[11—12], 其存在试验条件与野外自然条件差别较大和代表性不足等缺陷, 仅能反映某特定环境因子对森林生态系统的影响规律[13]。而利用不同海拔梯度间气温的自然差异, 通过异地移位试验开展野外增温模拟研究, 能够真实地反映自然环境中多因子共同作用下的凋落物分解进程, 很好地克服人工模拟试验的不足。因此, 移位试验被认为是开展模拟增温对森林生态系统结构和功能影响研究的新尝试。武夷山作为我国中亚热带拥有全球同纬度现存面积最大、保存最完整的常绿阔叶林生态系统, 最高海拔达2160.8 m, 被誉为华东第一峰, 具有明显的垂直气候带分布特征[14], 是开展增温对杉木凋落叶和宿存叶分解过程养分释放影响研究的理想平台。鉴于此, 本研究选取杉木人工林凋落叶和宿存叶作为研究对象, 利用武夷山不同海拔间的气温差异, 采用异地移位试验开展了为期360 d的野外定位模拟研究, 比较不同气温条件下杉木凋落叶和宿存叶分解速率及分解过程中N、P、Mg、Ca养分元素释放规律的差异, 为深入揭示全球气温升高对杉木人工林生态系统养分循环的影响机制提供科学依据。
1 材料与方法 1.1 研究区概况研究区位于福建省南平市武夷山国家公园自然保护区内, 地理位置介于117°27′—117°51′E、27°33′—27°54′N之间。气候属中亚热带季风性气候, 年平均气温为8.5—18.0 ℃, 年均降雨量1486—2150 mm, 主要降雨集中在3—6月, 约占年降雨量的60%, 年均相对湿度83.5%, 年均无霜期253—270 d。从黄岗山顶(2160.8 m) 起, 气温随海拔高度的下降逐渐升高, 土壤和植被呈明显地垂直地带性分布。植被类型沿海拔从高到低分别为高山草甸、亚高山矮曲林、针叶林、针阔混交林、亚热带常绿阔叶林。主要优势植被有毛竹(Phyllostachys edulis)、杉木(Cunninghamia lanceolata)、米槠(Castanopisi carlesii)、马尾松(Pinus massoniana)、黄山松(Pinus taiwanensis)、青冈(Cyclobalanopsis glauca)等。
1.2 试验材料的制备2021年8月在福建省三明市福建农林大学莘口教学林场15年生杉木人工林内设置3个20 m×20 m的标准地, 并在每个标准地内随机布设10个1 m×1 m凋落物收集框。9月对收集框中的杉木凋落物进行收集;同时于每个标准地内选取5棵标准木, 在每棵标准木下铺设尼龙纱网, 立标尺作为高度标记, 利用人工架梯并结合高枝剪的方式将杉木最高枯枝以下2 m范围内的宿存枝叶全部剪下, 对剪落到尼龙纱网上的宿存枝叶进行收集;随后把收集到的杉木凋落枝叶和宿存枝叶带回实验室, 将叶片人工剥离, 并去除杂质, 然后取少许凋落叶和宿存叶于65 ℃下烘干至恒重, 用于测定其含水率和初始养分含量(表 1);其余的凋落叶和宿存叶待其自然风干后分别装入20 cm×20 cm、孔径0.5 mm的尼龙分解袋中, 每袋装10 g±0.005g, 供不同海拔移位分解试验使用。凋落叶和宿存叶分别制备144袋分解样品, 共计288袋。
| 枯死叶类型 Type of dead leaves |
全碳 Total C |
全氮 Total N |
全磷 Total P |
镁 Mg |
钙 Ca |
碳氮比 C/N |
| 凋落叶Leaf litter | 496.42±12.38 | 15.22±0.20 | 0.34±0.02 | 0.64±0.03 | 5.05±0.58 | 40.62±0.83 |
| 宿存叶Leaf canopy litter | 506.03±16.94 | 17.68±0.31 | 0.35±0.11 | 0.68±0.04 | 6.56±0.57 | 28.62±0.76 |
2021年9月在武夷山国家公园自然保护区内选取1948 m、1410 m、1003 m、620 m四个海拔梯度作为本研究的分解试验地, 利用海拔梯度改变而导致增温的方式开展野外模拟试验。分别在不同海拔梯度林分中设置3个20 m×20 m的固定标准地, 将地表凋落物层清除干净, 将制备好的凋落叶和宿存叶分解袋与土壤充分接触, 同组间纵向排列, 不同组间设50 cm间隔带。并在不同海拔试验地中安装大气土壤温湿度自动监测仪(Soil temp & moisture datalogger, TMS series, Microclimate station, TMS-4), 每隔15 min自动监测记录大气的温度, 不同海拔试验地年平均气温增量及每月的月均温度见图 1, 土壤理化性质见表 2。每隔60 d进行一次凋落物分解袋取样, 每个海拔各标准地内每组试验处理每次取2个分解袋, 每次收取凋落叶和宿存叶各24袋(4个海拔×3个标准地×2袋/标准地), 从2021年9月到2022年9月进行为期360 d的定位观测。将取回的分解袋样品清理干净, 在65 ℃烘至恒重后称重, 粉碎过0.149 mm筛后, 进行N、P、Mg、Ca含量测定。采用Vario元素分析仪(Elementar vario EL III, Germany) 测定凋落叶和宿存叶的N含量, 采用硝酸-高氯酸消煮、电感耦合等离子体发射光谱仪(PE OPTIMA 8000) 测定P、Mg和Ca含量。

|
| 图 1 不同海拔试验地年平均气温增量及月均气温变化 Fig. 1 Annual mean temperature increment and monthly mean temperature change at different altitudes test sites |
| 海拔/m Altitude |
土壤容重/(g/cm3) Soil bulk density |
全碳/(g/kg) Total carbon |
全氮/(g/kg) Total nitrogen |
全磷/(g/kg) Total phosphorus |
土壤pH Soil pH |
| 620 | 0.87±0.11 | 4.24±0.69 | 0.43±0.07 | 0.32±0.01 | 4.76 |
| 1003 | 0.86±0.08 | 8.04±0.59 | 0.77±0.12 | 0.30±0.01 | 4.41 |
| 1410 | 0.94±0.10 | 5.24±0.68 | 0.71±0.13 | 0.46±0.06 | 4.38 |
| 1948 | 0.97±0.12 | 6.92±0.65 | 0.54±0.04 | 0.30±0.03 | 4.59 |
根据每次取样的干重, 计算干重残留率(D, %):
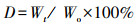
|
(1) |
式中, Wo为样本分解初始干重(g), Wt为分解t时间后的剩余干重(g)。
Olson指数衰减模型[15]:
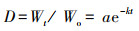
|
(2) |
式中, a为拟合参数, k为分解常数, t为分解时间。
养分残留率(Et, %):
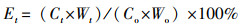
|
(3) |
式中, Co为样本初始养分含量(g/kg), Ct为分解t时间后的养分含量(g/kg);当Et > 100%, 养分累积;Et < 100%, 养分释放。
运用Excel 2010进行数据计算和处理, 使用Origin 2021软件进行相关图表绘制, 采用SPSS 19.0统计软件对不同海拔气温、不同分解时间下干重残留率及N、P、Mg、Ca残留率进行单因素方差分析(one-way ANOVA), 并结合多重比较(Duncan法) 对各处理组进行显著性差异分析(显著水平α=0.05)。以海拔年均气温和分解时间为固定影响因子, 干重残留率与N、P、Mg、Ca残留率为因变量进行双因素方差分析。采用一元线性回归分析检验分解过程中N、P、Mg、Ca残留率与干重残留率之间的相关性。本文图表中的统计数据均为平均值±标准差。
2 结果与分析 2.1 杉木凋落叶和宿存叶干重残留率变化由图 2可知, 随着分解时间的增加, 凋落叶和宿存叶在不同海拔气温条件下的干重残留率(D)均呈下降趋势。干重残留率在分解0—120 d时段内的损失速率要高于120—240 d时段。分解360 d后, 不同海拔气温下的干重残留率差异显著, 杉木凋落叶的干重残留率在各海拔梯度的大小排序表现为:D1948m(82.66%)> D1410m(80.63%)> D1003m(77.99%)> D620m(77.07%);杉木宿存叶的干重残留率表现为D1948m(85.16%)> D1410m (82.56%)> D620m (78.85%)> D1003m(77.84%)。

|
| 图 2 不同海拔气温条件下杉木凋落叶和宿存叶干重残留率的变化 Fig. 2 Changes of dry weight remaining of leaf litter and canopy litter of Chinese fir under different temperature conditions 不同大写字母表示同一时期不同海拔气温间差异显著, 不同小写字母表示同一海拔气温下不同时期间差异显著(P<0.05) |
将杉木凋落叶和宿存叶的干重残留率与分解时间进行指数回归方程拟合, 并估算半衰期(残留率为50%, t0.5) 和周转期(残留率为5%, t0.05)。表 3可知, 所得拟合方程R2均大于0.8, 表明拟合效果较好。凋落叶分解速率在不同海拔气温下表现为K620 m> K1003 m> K1410 m> K1948 m, 宿存叶表现为K1003 m> K620 m> K1410 m> K1948 m。凋落叶在620 m、1003 m、1410 m和1948 m气温下的半衰期分别为3.239 a、3.277 a、3.663 a和4.204 a, 周转期为14.582 a、14.790 a、16.042 a和18.505 a;宿存叶的半衰期分别为3.120 a、3.063 a、3.886 a和4.466 a, 周转期为13.539 a、13.388 a、16.969 a和19.615 a。
| 枯死叶类型 Type of dead leaves |
海拔 Altitude/m |
回归方程 Regression equation |
相关系数(R2) Correlation coefficient |
分解速率(K) Decomposing rate |
t0.5/a | t0.05/a |
| 凋落叶 | 620 | D = 96.499e-0.203t | 0.839 | 0.203 | 3.239 | 14.582 |
| Leaf litter | 1003 | D = 96.305e-0.200t | 0.883 | 0.200 | 3.277 | 14.790 |
| 1410 | D = 98.824e-0.186t | 0.856 | 0.186 | 3.663 | 16.042 | |
| 1948 | D = 98.376e-0.161t | 0.882 | 0.161 | 4.204 | 18.505 | |
| 宿存叶 | 620 | D = 99.636e-0.221t | 0.912 | 0.221 | 3.120 | 13.539 |
| Leaf canopy litter | 1003 | D = 98.995e-0.223t | 0.879 | 0.223 | 3.063 | 13.388 |
| 1410 | D = 99.083e-0.176t | 0.880 | 0.176 | 3.886 | 16.969 | |
| 1948 | D = 98.584e-0.152t | 0.872 | 0.152 | 4.466 | 19.615 | |
| t0.5:半衰期Half-life period;t0.05:周转期Turnover period | ||||||
方差结果表明(表 4), 气温、分解时间对杉木凋落叶和宿存叶干重残留率均呈极显著影响(P<0.001), 气温与时间的交互作用对宿存叶的干重残留率有极显著影响(P<0.01), 但对凋落叶干重残留率无显著影响(P>0.05)。
| 枯死叶类型 Type of dead leaves |
变异来源 Source of variation |
平方和(SS) Sum of squares |
df | F统计值 F-statistic |
P |
| 凋落叶 | 气温 | 248.23 | 3 | 62.15 | < 0.001 |
| Leaf litter | 时间 | 1557.72 | 5 | 233.99 | < 0.001 |
| 气温×时间 | 18.48 | 15 | 0.93 | 0.54 | |
| 宿存叶 | 气温 | 146.89 | 3 | 68.52 | < 0.001 |
| Leaf canopy litter | 时间 | 1583.14 | 5 | 443.20 | < 0.001 |
| 气温×时间 | 76.77 | 15 | 7.16 | < 0.001 |
由图 3可知, 在整个分解观测期内, 凋落叶和宿存叶的N残留率随分解时间的延长呈下降趋势, 同一分解时间下不同海拔气温间的N残留率差异显著(P<0.05)。在分解0—60 d时段内, 凋落叶N残留率在各海拔气温下表现为大于100%, 出现明显N素富集现象;而宿存叶N素仅在1003 m和1948 m气温下出现富集现象。在分解120—360 d时段内, 各海拔气温下凋落叶和宿存叶中的N素均呈释放状态。分解360 d后凋落叶N残留率为70.47%—83.57%, 宿存叶为66.90%—77.70%。P残留率在分解观测期间呈先降低后增加的变化趋势。在分解120—240 d时段内, P素处于释放状态, 且不同海拔气温间的P残留率差异不显著。在分解300—360 d时段内, 凋落叶和宿存叶P残留率在各海拔气温下表现为大于100%, 出现明显累积现象, 且不同海拔气温间的P残留率差异显著。分解360 d后凋落叶P残留率为104.00%—122.03%, 宿存叶为102.27%—111.10%。

|
| 图 3 不同海拔气温条件下杉木凋落叶和宿存叶分解过程中氮、磷残留率 Fig. 3 N, P remaining during the decomposition of leaf litter and canopy litter of Chinese fir under different temperature conditions 不同大写字母表示同一时期不同海拔气温间差异显著, 不同小写字母表示同一海拔气温下不同时期间差异显著(P<0.05) |
由图 4可知, 杉木凋落叶的Mg、Ca残留率低于宿存叶。凋落叶的Mg残留率在分解过程中呈先降后增的变化趋势, 一直处于释放状态。宿存叶的Mg残留率则表现为先增后降再增的变化趋势。在分解0—60 d时段内, 宿存叶在不同海拔气温间的Mg残留率差异不显著(P>0.05), 并出现轻微累积现象, 在分解120—360 d时段内, Mg处于释放状态。分解360 d后凋落叶的Mg残留率为55.87%—64.18%, 宿存叶则为62.43%—92.39%。凋落叶和宿存叶的Ca残留率在分解过程中整体呈先降后增的变化趋势, 在分解0—360 d时段内, 在不同海拔气温下Ca素均处于不同程度的释放状态。分解360 d后凋落叶的Ca残留率为59.12%—83.09%, 宿存叶则为57.75%—89.68%。

|
| 图 4 不同海拔气温条件下杉木凋落叶和宿存叶分解过程中镁、钙残留率 Fig. 4 Mg, Ca remaining during the decomposition of leaf litter and canopy litter of Chinese fir under different temperature conditions 不同大写字母表示同一时期不同海拔气温间差异显著, 不同小写字母表示同一海拔气温下不同时期间差异显著(P<0.05) |
由表 5可知, 气温、分解时间及其二者交互作用均会对凋落叶和宿存叶N、Ca残留率产生极显著影响(P<0.01)。气温对凋落叶Mg残留率产生显著影响(P<0.05), 气温与时间的交互作用凋落叶P残留率产生显著影响(P<0.05)。
| 枯死叶类型 Type of dead leaves |
变异来源 Source of variation |
氮残留率 N remaining |
磷残留率 P remaining |
镁残留率 Mg remaining |
钙残留率 Ca remaining |
| 凋落叶 | 气温 | 109.59*** | 9.79*** | 3.15* | 7.71*** |
| Leaf litter | 时间 | 857.62*** | 274.43*** | 55.96*** | 10.90*** |
| 气温×时间 | 46.95*** | 2.05* | 3.65*** | 2.58** | |
| 宿存叶 | 气温 | 484.23*** | 21.78*** | 19.73*** | 14.19*** |
| Leaf canopy litter | 时间 | 1597.96*** | 593.24*** | 110.12*** | 8.14*** |
| 气温×时间 | 88.55*** | 7.67*** | 8.46*** | 2.90** | |
| ns: P>0.05; *: P<0.05; **: P<0.01; ***: P<0.001 | |||||
将凋落叶和宿存叶的干重残留率与N、P、Mg、Ca残留率进行相关性分析发现(图 5), 凋落叶和宿存叶在各海拔气温下的干重残留率与N残留率呈极显著正相关(P<0.001), 与P呈显著负相关(P<0.05)。在1948 m气温下Mg残留率与干重残留率呈显著正相关, 而在620 m、1003 m和1410 m气温下二者相关性未达显著性水平。凋落叶在各海拔气温下的Ca残留率与干重残留率相关性不显著, 宿存叶在620 m气温下Ca残留率与干重残留率呈显著正相关(P<0.001)。表明增温会改变养分残留率与干重残留率间的相关关系。

|
| 图 5 杉木凋落叶和宿存叶干重残留率与养分残留率的关系 Fig. 5 Relationship between dry weight remaining and nutrient remaining in leaf litter and canopy litter decomposition of Chinese fir ns: P>0.05; *: P<0.05; **: P<0.01; ***: P<0.001 |
凋落物分解是森林土壤养分的重要来源, 森林凋落物经过降水淋溶、动物和微生物的啃食粉碎及一系列的物理化学反应从而造成质量的损失[16]。温度是影响凋落物分解的主要驱动因素之一, 研究表明在外界环境因子相同的情况下, 气温升高将显著提高森林凋落物的分解速率[17—18]。Liu等[17]利用纬度梯度模拟增温对红松(Pinus Koraiensis) 凋落物分解的影响研究中发现, 气温升高显著提高了凋落物分解速率。在本研究中, 杉木凋落叶分解速率随海拔气温的升高呈上升趋势, 表现为K620 m> K1003 m> K1410 m> K1948 m, 与Zhou等[18]、郭忠玲等[19]研究结果相似, 宿存叶则表现为K1003 m> K620 m> K1410 m> K1948 m, 并未表现出随海拔气温升高而增加的变化规律, 可能与分解袋所处海拔环境的异质性密切相关。研究发现, 海拔环境中的温湿度、郁闭度、林下光环境等均对分解有一定影响, 不同海拔气温条件下宿存叶的分解过程同时受到多种因素的综合影响[20], 同时短期的网袋分解法也只能反映局部范围内分解速率的变化规律。在凋落物分解过程中, 微生物群落和相关酶活在质量损失过程中均起着决定性作用[21—22], 气温的升高必将引起分解环境及相关土壤酶活性的变化, 改变微生物菌群的相对丰度及物种多样性, 进而影响凋落物的分解进程。林惠瑛等[23]对武夷山土壤酶活性测定发现, β-N\乙酰氨基葡糖苷酶、酸性磷酸酶活性随气温的升高呈上升的趋势, 并表明在不同温度区间内限制微生物活动的主控因子有所不同。此外, 凋落叶和宿存叶在不同海拔气温下分解速率的差异可能是由二者初始基质质量的不同和微生物菌群的偏好性所导致的。相较于宿存叶而言, 凋落叶经历了更长时间的淋溶和光降解过程, 导致二者初始养分浓度的截然不同。研究发现, 林木宿存枝叶比凋落枝叶具有更高的养分含量, 被视为一种自我养分保护措施, 在土壤养分供应不足时, 林木可以对宿存枝叶中的累积养分进行重吸收, 最大限度地实现自身养分的内循环, 从而减少对土壤养分的依赖[24]。
本研究中, 杉木凋落叶和宿存叶的干重残留率随分解时间的延长呈下降趋势, 在分解0—60 d时段内具有较高的分解速率, 整体呈先降后增的变化趋势, 与李娜等[20]研究结果相似。在凋落物分解初期, 易分解组分和水溶性养分的快速淋失导致前期分解速率较高, 随着分解的进行, 木质素等大分子聚合物浓度相对增加, 延缓了分解速率[25]。此外, 温度的季节性变化也是造成分解速率变化的可能原因之一, 研究表明在一定温度区间内, 微生物活性和酶活性与温度呈正相关关系[26]。在分解120—180 d时段(1月—3月) 正处冬季, 气温较低, 微生物和土壤相关酶活性较低, 进而导致凋落物分解速率降低。
3.2 增温对林木凋落物分解过程中养分释放的影响在凋落物分解过程中, 降水淋溶和微生物降解是影响养分释放的主控因子[27]。温度变化不仅影响微生物群落分布及其活性, 还可能造成降雨格局的改变, 进而影响凋落物养分释放过程。Liu等[28]利用移位试验模拟气温升高对木荷(Schima superba) 凋落叶分解过程中养分释放的影响研究中发现, 增温显著促进K、N、P、Mg和Ca的释放。而有学者采用原位埋设电阻加热电缆模拟增温对亚热带天然林凋落物分解试验中发现, 增温对凋落叶N、P释放无显著影响[29]。本研究发现, 杉木凋落叶和宿存叶中养分残留率受海拔年均气温的显著影响(表 5), 增温显著促进N、P、Mg、Ca的释放, 与Liu等研究结果一致。研究表明, 增温显著提升了土壤微生物C、N、P含量, 加快了微生物矿化速率, 进而加速凋落物内矿质元素的释放[30]。相较于高海拔低温环境而言, 低海拔温润环境为土壤动物和微生物菌群提供了一个更舒适的生存空间, 微生物活性高、周转速率快, 对营养元素的消耗量较大[31]。另外, 增温可能会刺激微生物合成更多的降解酶, 养分释放速率的加快是为相关酶的合成提供足够的养分供应。养分释放速率的加快会促进凋落物的质量损失, 反之抑制[32], 在本研究中, 干重残留率与N残留率呈极显著正相关(图 5), 也证实了这一观点。
在凋落物分解过程中, 养分释放模式主要有:释放、富集-释放、淋溶-富集-释放[33], 养分初始含量和微生物菌群需求量是决定养分释放模式的关键因子, 当养分释放量能够满足微生物活动所需时, 呈释放现象, 反之则呈累积现象[34]。在本研究中, 杉木凋落叶N素在各海拔气温下均呈富集-释放模式, 而宿存叶N素在620 m和1410 m气温下呈净释放模式, 在1003 m和1948 m气温下呈富集-释放模式, 表明增温改变了杉木宿存叶N素的释放模式。不同温度环境下菌群数量及相关酶活性的异质性可能是造成该变化的可能原因之一, 在高温环境下, 菌群对宿存叶中N素具有较高的吸收利用率是为合成更多的凋落物分解酶。研究表明, 凋落物N素的释放对腐殖质层中微生物群落的繁殖和周转至关重要, 在凋落物分解初期, 释放N素并不足以满足异养生物活动所需, 出现N素富集[31]。增温并未引起凋落叶和宿存叶P素释放模式的改变, 在各海拔气温下均呈现释放-富集模式。P作为凋落物内的易分解可溶性养分, 在分解初期易被大量淋溶[35], 而此时由土壤进入分解袋内的微生物菌群数量较少, 淋溶所释放出的P素供给远超微生物活动所需, 呈释放模式。在分解300—360 d, 正处高温多雨季节(7月—9月), 微生物菌群丰度和多样性增加, 活性较高, 分解所释放出的P素无法满足微生物群落的周转, 出现累积现象[24]。此外, 有报道称N、P元素的释放模式与土壤微生物C、N、P间的化学计量密切相关[36]。Pei等[37]研究发现, 当C∶N>25时, N成为限制凋落物分解的主控因子, N就会出现固持累积现象;当C∶N < 25时, 限制元素为P。在本研究中, 凋落叶和宿存叶的初始C∶N均大于25, 分解过程出现N素累积, 与Pei等研究结果一致。
凋落物分解所释放的Mg、Ca是林木能量代谢与光合作用所必需营养元素的重要来源, 在土壤有机质形成和调控林木生长策略方面均发挥重要作用[38]。本研究中, 凋落叶Mg、Ca在各海拔气温下均呈净释放状态, 宿存叶Mg呈富集-释放模式, Ca呈净释放模式, 表明模拟增温并未引起Mg、Ca释放模式的改变。对于Mg、Ca而言, 其释放格局并不像N、P一样易受雨水淋溶的影响, 而是更多地取决于微生物降解[39]。一般而言, Mg主要以游离形态存在于植物细胞溶液中极易从植物凋落物中淋失, 宿存叶Mg在分解初期的富集可能是由于侵入分解袋内微生物数量激增所导致的。Ca主要以螯合态存在于植株体内, 与有机碳的耦合关系较弱[39]。凋落叶和宿存叶初始Ca含量较高, 微生物菌群对Ca的需求量相对较低, 因此在分解过程中Ca释放量远超需求量, 从而呈净释放模式[40]。
4 结论研究发现增温对杉木凋落叶和宿存叶的分解具有显著促进作用, 随气温的升高, 凋落叶和宿存叶分解速率整体呈增大趋势。增温显著促进了凋落叶和宿存叶N、P、Mg、Ca的释放, 有利于林地内的养分循环和土壤培肥, 在今后的杉木人工林管理中适时地将宿存枝叶敲落, 堆积于地表, 将有利于长期维持林地生产力。增温并未改变凋落叶和宿存叶分解过程中P、Mg、Ca素的释放模式, 但改变宿存叶N素的释放模式。杉木凋落叶和宿存叶的N、P残留率与干重残留率密切相关。本研究结果可为预测全球气温变化背景下杉木人工林枯死叶分解过程中的养分循环过程提供一定数据参考。
| [1] |
王亚萍, 王帅, 丁婧祎, 赵文武, 尹嘉敏. 气候变化背景下全球陆地干湿变化研究综述. 生态学报, 2023, 43(2): 475-486. DOI:10.5846/stxb202109072527 |
| [2] |
周天军, 陈梓明, 陈晓龙, 左萌, 江洁, 胡帅. IPCC AR6报告解读: 未来的全球气候——基于情景的预估和近期信息. 气候变化研究进展, 2021, 17(6): 652-663. |
| [3] |
Peng X B, Yu M, Chen H S. Projected changes in terrestrial vegetation and carbon fluxes under 1.5 ℃ and 2.0 ℃ global warming. Atmosphere, 2021, 13(1): 42. DOI:10.3390/atmos13010042 |
| [4] |
马转转, 张全智, 王传宽. 六种温带森林类型凋落物量长期动态及其环境驱动. 生态学报, 2023, 43(17): 7307-7316. DOI:10.20103/j.stxb.202208032214 |
| [5] |
Wu P P, Jiang L X, Zhang Y, Tu Q H, Mao R. Manganese addition accelerates litter decomposition and alters litter mixing effects in the late stage in subtropical plantations of Southern China. Plant and Soil, 2022, 481(1): 501-510. |
| [6] |
余李, 廖杨文科, 自海云, 潘畅, 周梦田, 曹超阳, 李孝刚. 杉木连栽对土壤细菌群落及其抑病能力的影响. 生态学报, 2023, 43(12): 5038-5049. DOI:10.5846/stxb202204281185 |
| [7] |
夏丽丹, 于姣妲, 邓玲玲, 李小艳, 周垂帆, 徐永兴. 杉木人工林地力衰退研究进展. 世界林业研究, 2018, 31(2): 37-42. |
| [8] |
盛炜彤, 范少辉. 杉木及其人工林自身特性对长期立地生产力的影响. 林业科学研究, 2002, 15(6): 629-636. |
| [9] |
郑临训. 杉木人工林枯损枝叶营养特点的研究. 林业科学研究, 1997, 10(6): 48-52. |
| [10] |
Tong R, Zhou B Z, Jiang L N, Ge X G, Cao Y H, Shi J X. Leaf litter carbon, nitrogen and phosphorus stoichiometry of Chinese fir (Cunninghamia lanceolata) across China. Global Ecology and Conservation, 2021, 27: e01542. DOI:10.1016/j.gecco.2021.e01542 |
| [11] |
吴小健, 石焱, 朱嘉宁, 孙麟钧, 马祥庆. 全球变暖对森林凋落物的影响. 世界林业研究, 2023, 36(1): 26-32. |
| [12] |
徐振锋, 尹华军, 赵春章, 曹刚, 万名利, 刘庆. 陆地生态系统凋落物分解对全球气候变暖的响应. 植物生态学报, 2009, 33(6): 1208-1219. |
| [13] |
平云梅, 崔丽娟, 李伟, 潘旭, 雷茵茹, 魏佳明. 湿地挺水植物凋落物分解研究概述. 世界林业研究, 2018, 31(3): 15-20. |
| [14] |
Li X, Wu P F, Guo F T, Hu X S. A geographically weighted regression approach to detect divergent changes in the vegetation activity along the elevation gradients over the last 20years. Forest Ecology and Management, 2021, 490: 119089. DOI:10.1016/j.foreco.2021.119089 |
| [15] |
刘增文, 潘开文. Olson枯落物分解模型存在的问题与修正. 西北农林科技大学学报: 自然科学版, 2005, 33(1): 69-70. |
| [16] |
Mehta N, Dinakaran J, Patel S, Laskar A H, Yadava M G, Ramesh R, Krishnayya N S R. Changes in litter decomposition and soil organic carbon in a reforested tropical deciduous cover (India). Ecological Research, 2013, 28(2): 239-248. DOI:10.1007/s11284-012-1011-z |
| [17] |
Liu L, Yang F, Wang Y J, Shen X, Janssens I A, Guenet B, Xiao C W. Fine-root turnover, litterfall, and soil microbial community of three mixed coniferous-deciduous forests dominated by Korean pine (Pinus koraiensis) along a latitudinal gradient. Frontiers in Plant Science, 2019, 10: 1298. DOI:10.3389/fpls.2019.01298 |
| [18] |
Zhou Y, Clark M, Su J Q, Xiao C W. Litter decomposition and soil microbial community composition in three Korean pine (Pinus koraiensis) forests along an altitudinal gradient. Plant and Soil, 2015, 386(1-2): 171-183. DOI:10.1007/s11104-014-2254-y |
| [19] |
郭忠玲, 郑金萍, 马元丹, 李庆康, 于贵瑞, 韩士杰, 范春楠, 刘万德. 长白山各植被带主要树种凋落物分解速率及模型模拟的试验研究. 生态学报, 2006, 26(4): 1037-1046. |
| [20] |
李娜, 赵传燕, 郝虎, 臧飞, 常亚鹏, 汪红, 杨建红. 海拔和郁闭度对祁连山青海云杉林叶凋落物分解的影响. 生态学报, 2021, 41(11): 4493-4502. DOI:10.5846/stxb201906171275 |
| [21] |
王小平, 杨雪, 杨楠, 辛晓静, 曲耀冰, 赵念席, 高玉葆. 凋落物多样性及组成对凋落物分解和土壤微生物群落的影响. 生态学报, 2019, 39(17): 6264-6272. DOI:10.5846/stxb201805251153 |
| [22] |
葛晓敏, 吴麟, 唐罗忠. 森林凋落物分解与酶的相互关系研究进展. 世界林业研究, 2013, 26(1): 43-47. |
| [23] |
林惠瑛, 周嘉聪, 曾泉鑫, 孙俊, 谢欢, 刘苑苑, 梅孔灿, 吴玥, 元晓春, 吴君梅, 苏先楚, 程栋梁, 陈岳民. 土壤酶计量揭示了武夷山黄山松林土壤微生物沿海拔梯度的碳磷限制变化. 应用生态学报, 2022, 33(1): 33-41. |
| [24] |
Zhou L L, Li S B, Jia Y Y, Heal K V, He Z M, Wu P F, Ma X Q. Spatiotemporal distribution of canopy litter and nutrient resorption in a chronosequence of different development stages of Cunninghamia lanceolata in southeast China. The Science of the Total Environment, 2021, 762: 143153. DOI:10.1016/j.scitotenv.2020.143153 |
| [25] |
Zhang P H, Lin J, Hao J T, Li C C, Quan W X. Decomposition characteristics of lignocellulosic biomass in subtropical Rhododendron litters under artificial regulation. Metabolites, 2023, 13(2): 279. DOI:10.3390/metabo13020279 |
| [26] |
程严, 列志旸, 刘旭军, 李旭, 吴婷, 褚国伟, 孟泽, 刘菊秀. 增温对南亚热带针阔叶混交林凋落物分解酶活性的影响. 应用与环境生物学报, 2021, 27(4): 923-929. |
| [27] |
张晓曦, 胡嘉伟, 胡漫, 黄琰莹, 王羿人, 张蔓. 黄土丘陵区代表性乔灌草植物凋落物关键代谢产物释放的室内模拟研究. 西北植物学报, 2023, 43(4): 688-697. |
| [28] |
Liu J X, Liu S G, Li Y Y, Liu S Z, Yin G C, Huang J, Xu Y. Warming effects on the decomposition of two litter species in model subtropical forests. Plant and Soil, 2017, 420(1/2): 277-287. |
| [29] |
Liu X F, Chen S D, Li X J, Yang Z J, Xiong D C, Xu C, Wanek W, Yang Y S. Soil warming delays leaf litter decomposition but exerts no effect on litter nutrient release in a subtropical natural forest over 450days. Geoderma, 2022, 427: 116139. DOI:10.1016/j.geoderma.2022.116139 |
| [30] |
van Meeteren M J M, Tietema A, van Loon E E, Verstraten J M. Microbial dynamics and litter decomposition under a changed climate in a Dutch heathland. Applied Soil Ecology, 2008, 38(2): 119-127. DOI:10.1016/j.apsoil.2007.09.006 |
| [31] |
赵盼盼, 周嘉聪, 林开淼, 张秋芳, 袁萍, 曾晓敏, 苏莹, 徐建国, 陈岳民, 杨玉盛. 海拔梯度变化对中亚热带黄山松土壤微生物生物量和群落结构的影响. 生态学报, 2019, 39(6): 2215-2225. DOI:10.5846/stxb201804170877 |
| [32] |
Fang S Z, Li H Y, Xie B D. Decomposition and nutrient release of four potential mulching materials for poplar plantations on upland sites. Agroforestry Systems, 2008, 74(1): 27-35. DOI:10.1007/s10457-008-9155-0 |
| [33] |
陈栎霖, 刘语欣, 范川, 李贤伟, 刘运科, 杨正菊, 张军. 台湾桤木-扁穗牛鞭草复合模式下凋落物分解及其养分释放动态. 南京林业大学学报: 自然科学版, 2015, 39(1): 49-54. |
| [34] |
Tu L H, Hu H L, Hu T X, Zhang J, Li X W, Liu L, Xiao Y L, Chen G, Li R H. Litterfall, litter decomposition, and nutrient dynamics in two subtropical bamboo plantations of China. Pedosphere, 2014, 24(1): 84-97. DOI:10.1016/S1002-0160(13)60083-1 |
| [35] |
温达志, 魏平, 张佑昌, 孔国辉. 鼎湖山南亚热带森林细根分解干物质损失和元素动态. 生态学杂志, 1998, 17(2): 1-6. |
| [36] |
Xu X F, Thornton P E, Post W M. A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems. Global Ecology and Biogeography, 2013, 22(6): 737-749. DOI:10.1111/geb.12029 |
| [37] |
Pei G T, Liu J, Peng B, Gao D C, Wang C, Dai W W, Jiang P, Bai E. Nitrogen, lignin, C/N as important regulators of gross nitrogen release and immobilization during litter decomposition in a temperate forest ecosystem. Forest Ecology and Management, 2019, 440: 61-69. DOI:10.1016/j.foreco.2019.03.001 |
| [38] |
Lovett G M, Arthur M A, Crowley K F. Effects of calcium on the rate and extent of litter decomposition in a northern hardwood forest. Ecosystems, 2016, 19(1): 87-97. DOI:10.1007/s10021-015-9919-0 |
| [39] |
Yue K, Ni X Y, Fornara D A, Peng Y, Liao S, Tan S Y, Wang D Y, Wu F Z, Yang Y S. Dynamics of calcium, magnesium, and manganese during litter decomposition in alpine forest aquatic and terrestrial ecosystems. Ecosystems, 2021, 24(3): 516-529. DOI:10.1007/s10021-020-00532-5 |
| [40] |
涂利华, 胡庭兴, 张健, 戴洪忠, 李仁洪, 向元彬, 雒守华. 模拟氮沉降对两种竹林不同凋落物组分分解过程养分释放的影响. 生态学报, 2011, 31(6): 1547-1557. |