 2024, Vol. 44
2024, Vol. 44文章信息
- 赵金发, 刘永, 李纯厚, 石娟, 王腾, 谢宏宇, 康志鹏, 宋晓宇
- ZHAO Jinfa, LIU Yong, LI Chunhou, SHI Juan, WANG Teng, XIE Hongyu, KANG Zhipeng, SONG Xiaoyu
- 西沙群岛北礁珊瑚礁鱼类种类组成及变化特征
- Composition and variation characteristics of coral reef fish species in Beijiao Atoll of the Xisha Islands
- 生态学报. 2024, 44(16): 7384-7400
- Acta Ecologica Sinica. 2024, 44(16): 7384-7400
- http://dx.doi.org/10.20103/j.stxb.202312142726
-
文章历史
- 收稿日期: 2023-12-14
- 网络出版日期: 2024-06-18






2. 西沙岛礁渔业生态系统海南省野外科学观测研究站/海南省深远海渔业资源高效利用与加工重点实验室/三亚热带水产研究院, 三亚 572018
2. Scientific Observation and Research Station of Xisha Island Reef Fishery Ecosystem of Hainan Province/Key Laboratory of Efficient Utilization and Processing of Marine Fishery Resources of Hainan Province/Sanya Tropical Fisheries Research Institute, Sanya 572018, China
珊瑚礁生态系统是地球上生物多样性最高的生态系统之一, 被誉为“海洋中的热带雨林”, 滋养了世界上三分之一的海洋鱼类[1], 并且对人类社会产生了巨大的经济价值, 保障了沿海区域生态安全[2—3]。珊瑚礁生态系统不仅可以为许多海洋鱼类提供栖息地和食物源, 而且还是海洋物质循环和能量流动中关键的一环, 为提升海洋环境的自我调节能力做出了巨大贡献[4]。但是近几十年来, 珊瑚礁生态系统的健康与稳定面临着巨大的压力。人类活动对珊瑚礁生态系统造成了巨大的威胁, 主要体现在海洋污染和不可持续的捕捞方式两方面。例如, 海洋污染由于塑料废物的堆积, 导致珊瑚礁疾病的增加;而过度捕捞则引发了珊瑚礁生态系统中生物群落结构的明显改变[5—6]。全球气候变化也是对珊瑚礁的主要威胁之一, 逐渐升高的海洋温度导致海洋酸化, 这些均会加剧珊瑚白化和死亡[7]。另外, 海洋变暖也会通过增加全球气旋的强度间接威胁珊瑚礁, 强烈的气旋会导致珊瑚礁遭到破坏及相关生物丰度和多样性的损失[8]。
珊瑚礁鱼类是维系珊瑚礁生态系统健康的关键类群之一, 例如蝴蝶鱼(Chaetodontidae)由于它们的取食习性和与珊瑚礁栖息地的密切关系而被认为是珊瑚礁生态系统健康状况的生物指示器[9]。波纹唇鱼(Cheilinus undulatus)等多种鱼类可以通过捕食作用控制长棘海星(Acanthaster planci)的数量, 对保护珊瑚礁有重要的作用[10]。草食性鱼类可以摄食珊瑚礁上的藻类, 控制藻类的生长, 为珊瑚清理出生长空间, 维持珊瑚的活力与繁荣[11]。但是, 人类活动造成了珊瑚礁鱼类群落结构发生变化, 研究发现过度捕捞使大型肉食性鱼类种群数量锐减, 多地珊瑚礁鱼类小型化明显, 这可能会造成珊瑚和藻类的平衡被打破, 大量藻类占据珊瑚的生存空间, 珊瑚礁生境进一步退化[6, 12]。因此, 监测和掌握鱼类群落的长期变化对于珊瑚礁生态系统健康和鱼类资源保护至关重要。国内外学者对于珊瑚礁鱼类群落组成和长期变化对极为重视, 如澳大利亚大堡礁[13]、马来西亚半岛[14]、以及我国西沙的东岛[15]、七连屿[16]、永兴岛[17]以及永乐环礁[18]。
北礁是西沙群岛中距离大陆最近的一座暗礁, 拥有非常丰富的鱼类资源。然而该礁受到人类扰动和气候变化的影响, 导致鱼类群落结构发生变化。为更好地了解北礁珊瑚礁鱼类资源的本底情况, 本研究对1998—2023年的珊瑚礁鱼类调查数据进行了回顾分析。主要研究内容为:(1)汇编北礁至今(1998—2023)的珊瑚礁鱼类名录;(2)北礁珊瑚礁鱼类现状;(3)在多重影响下珊瑚礁鱼类群落结构如何变化?本研究将全面解析北礁珊瑚礁鱼类资源现状, 综合鱼类群落结构的演替特征预测未来鱼类群落演变趋势, 并分析了这一演变原因。本研究将为北礁珊瑚礁鱼类资源研究提供数据资料, 对于西沙珊瑚礁生态系统的保护具有重要科学和实践意义。
1 材料与方法 1.1 研究区域及调查时间北礁(17°03′N——17°06′N, 111°26′E——111°33′E)位于中国西沙群岛最北边, 是永乐环礁北端的椭圆形环礁, 全长约12 km, 宽约4.8 km, 面积约为50 km2。低潮时部分露出海面, 高潮时也有个别点露出, 其环礁口多开于西面, 珊瑚丛生, 海洋生物极为丰富。本研究使用中国水产科学研究院南海水产研究所在1998—1999年春季(4、5月)和2003年春季(5月)在西沙群岛北礁采集的鱼类资源历史档案数据, 并结合2023年春季(3、4月)的实地调查航次数据, 对其珊瑚礁鱼类种类组成及群落结构的变化特征进行了统计分析。
1.2 样品采集1998—1999年的调查方法为手钓、延绳钓和流刺网;2003年的调查方法是延绳钓和流刺网;2023年的调查方法为潜水捕捞、水下视觉普查技术、手钓和环境DNA相结合的方式。流刺网一般在傍晚(17:00)下网, 早上(06:00)收网。放网长度根据地形而定, 在400—640 m之间, 捕捞水深为5—25 m(网具规格:1.0 m×40 m、1.6 m×40 m、1.8 m×40 m, 网目大小:5—12 cm)。延绳钓是指由带有浮子和沉子的干线, 并结有若干带有钓钩的支线而组成的钓具。一般于傍晚(17:00)开始, 次日凌晨(06:00)结束, 放置时间约12小时。捕捞水深为40—110 m(鱼钩:15或16号, 数量:80钩、120钩、160钩、180钩、200钩、300钩)。潜水捕捞时间一般为20:00—24:00, 由渔民在珊瑚礁区使用鱼枪进行无选择性的潜水捕捞, 采样深度为5—30 m。水下视觉普查由受雇专业潜水员在白天使用GoPro(HERO 11)进行水肺潜水, 由两种拍摄方式组成, 一种为在不同水深(5 m、10 m和15 m)铺设样带(长50 m), 由潜水员进行样带拍摄;另一种则由潜水员在水下随机拍摄。手钓时间为上午(7:00—10:00)和晚上(19:00—23:00), 由4—6名钓手在采样船两侧布设鱼钩, 使用2—7号钩, 采样深度为5—40 m。环境DNA的样品采集分为两步:(1)采水, 使用5 L的采水器在每个采样点采集表、中、底层水共15 L, 保存至已消毒的一次性采样袋中, 再置于保温箱中-4℃保存并与1小时内运回大船, 采样深度为0—60 m。每个采样点采集3个重复样品, 采水器在使用前后均用纯水冲洗三次, 采样人员不同采样点更换手套。(2)抽滤, 样品运回大船后立即使用真空抽滤设备将水样过滤至直径47 mm、孔径0.2 μm的聚碳酸酯滤膜上, 将抽滤后的滤膜放入2 mL冻存管并置于液氮中-80℃保存。另外, 每组样品使用相同的设备和程序, 在同一天抽滤相同体积的纯水作为阴性对照。获取的鱼类样本均冰鲜保存带回实验室进行解剖实验, 水下视频影像信息专人进行分析鉴定和整理(图 1)。分类系统参考世界鱼类数据库Fishbase database (https://www.fishbase.se/search.php)、中国台湾鱼类资料库(https://fishdb.sinica.edu.tw/)和中国生物物种名录2023。

|
| 图 1 北礁珊瑚礁鱼类的水下摄影图片 Fig. 1 Underwater photography images of coral reef fish in Beijiao Atoll |
使用Qiagen公司的PowerWater DNA Isolation Kits试剂盒提取滤膜DNA。每个样品进行独立提取, 并使用空白滤膜作为阴性对照。并用1%琼脂糖凝胶电泳检测DNA的完整性, 同时利用NanoDrop和Qubit检测DNA的浓度和纯度。将提取到的DNA溶液保存至-20 ℃冰箱备用。
1.3.2 12S rRNA基因扩增子测序PCR扩增选用MiFish-U引物(F:5′-GTYGGTAAAWCTCGTGCCAGC-3′, R:5′-CATAGTGGGGTATCTAAT CCYAGTTTG-3′)针对线粒体12S rRNA基因的目标区域进行扩增[19]。PCR扩增反应体系(20 μL)包括:2—5 μL模板DNA(10 ng/μL)、各0.8 μL正反向引物(10 μmol/L)、2 μL dNTPs、4 μL 5 ×FastPfu缓冲液、0.4 μL FastPfu聚合酶和7—10 μL ddH2O。PCR反应程序为:95 ℃预变性5 min;27个循环包括:95 ℃变性30 s, 55 ℃退火30 s, 72 ℃延伸45 s;72 ℃最后延伸10 min。每次PCR反应使用ddH2O作为模板进行PCR阴性对照, 每份样品重复扩增三次, 将同一样品的PCR产物混合后, 使用2%琼脂糖凝胶电泳检测。所有阴性对照均未观察到目的条带, 表明从采样、过滤、DNA提取至PCR扩增均未受到外源鱼类DNA污染的影响。扩增产物通过1 ∶ 1磁珠回收后, 按照Qubit浓度等质量原则计算各个样品所需体积, 将各个样本PCR产物进行混合汇集等量, 后续在Illumina NovaSeq 6000平台上进行测序。
1.3.3 序列处理测序得到的reads首先通过Fastp软件(v0.12.4)对原始reads进行质控, 去除接头, 过滤掉低质量的reads, 过滤后将所有样本的拼接序列进行合并, 并将拼接序列按照相似度98%进行聚类。得到的OTU序列与目标基因宏条码数据库的参考序列进行BLAST(v2.2.31)比对, 根据相似度98%, 95%, 90%, 85%的阈值依次赋予种, 属, 科, 目, 纲五个分类水平的界限。
1.4 食性鱼类的食性组成信息根据解剖分析并参考世界鱼类数据库Fishbase database (https://www.fishbase.se/search.php)和中国台湾鱼类资料库(https://fishdb.sinica.edu.tw/)分为杂食性、肉食性和植食性[16]。
1.5 体型鱼类最大全长从世界鱼类数据库Fishbase database (https://www.fishbase.se/search.php)中获取, 极少数未获取到最大全长的, 参考同属鱼类值。鱼类体型以最大全长的35 cm和65 cm为分界线分为大型鱼、中型鱼和小型鱼[16]。
1.6 数据处理利用SPSS 24.0软件分析珊瑚礁鱼类最大全长与种类数的相关性关系, 并采用线性函数、指数函数、幂函数、逻辑函数方程拟合, 以相关系数R2最大者为拟合方程, 分析其相关性特征。同时利用独立样本T检验分析不同食性类型珊瑚礁鱼类最大全长之间的差异显著性, 统计显著性标准设为0.05。采用分类学多样性指数分析珊瑚礁鱼类群落多样性[20]。相比于传统多样性计算方法, 分类学多样性指数不受采样方法、栖息地生态类型自然变化等影响, 可解释群落的种间关系[21]。利用Primer5.2软件的Taxdtest包分析珊瑚礁鱼类群落的分类学多样性指数。
平均分类差异指数(Δ+)和分类差异变异指数(Λ+)计算公式如下[21]:

|
(1) |
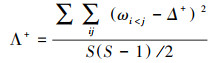
|
(2) |
式中, wij为第i和j个种类在分类系统树中的路径长度;S为种类数。将门、纲、目、科、属、种6个分类等级间的加权路径长度的权重设置为100.000、83.333、66.667、50.000、33.333、16.667。
Jaccard相似性系数(Js)[22]:
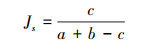
|
(3) |
式中, a为年份a的鱼类种类数或者目、科、属的数量, b为年份b的鱼类种类数或者目、科、属的数量, c为两个年份调查的鱼类共有种类数或者目、科、属的数量。相似级别分别是: 0 < Js < 0.25为极不相似; 0.25≤Js < 0.50为不相似; 0.50≤Js < 0.75为中等相似; 0.75≤Js < 1.00为极相似。
珊瑚礁鱼类多样性指数Coral Fish Diversity Index(CFDI)[23]:

|
(4) |
式中, N为预测总物种数, CFDI为隆头鱼科、蝴蝶鱼科、鹦嘴鱼科、刺盖鱼科、刺尾鱼科和雀鲷科的总物种数。
2 结果 2.1 鱼类种类组成特征综合历史数据, 在北礁珊瑚礁海域共发现鱼类270种(图 2, 附表 1)。其中, 水下潜捕共发现46种, 手钓共发现37种, 水下摄影共发现125种, 流刺网共发现78种, 延绳钓共发现12种, eDNA共发现90种。辐鳍鱼纲(Actinopterygii)共记录10目鱼类, 鲈形目(Perciformes)鱼类种类数最多为212种, 占总种类数的78.52%。软骨鱼纲(Chondrichthyes)共记录2目鱼类, 六鳃鲨目(Hexanchiformes)鱼类种类数最多为2种, 占总种类数的0.74%。

|
| 图 2 北礁珊瑚礁鱼类种类组成 Fig. 2 Composition of coral reef fish species in Beijiao Atoll |
| 序号 No. |
科 Family |
种 Species |
IUCN濒危状态 Conservation Status |
食性 Feeding habits |
1998—2003 | 2023 | 捕捞方式 Fishing methods |
| 1 | 海鳝科Muraenidae | 多带海鳝Echidna polyzona | LC | 肉食性 | 1 | f | |
| 2 | 条纹裸海鳝Gymnomuraena zebra | LC | 肉食性 | 1 | f | ||
| 3 | 伯恩斯裸胸鳝Gymnothorax buroensis | LC | 肉食性 | 1 | f | ||
| 4 | 豆点裸胸鳝Gymnothorax favagineus | LC | 肉食性 | 1 | d | ||
| 5 | 斑点裸胸鳝Gymnothorax meleagris | LC | 肉食性 | 1 | a | ||
| 6 | 白斑裸胸鳝Gymnothorax prismodon | LC | 肉食性 | 1 | d, e | ||
| 7 | 异纹裸胸鳝Gymnothorax richardsonii | LC | 肉食性 | 1 | d, e | ||
| 8 | 鞍斑裸胸鳝Gymnothorax rueppellii | LC | 肉食性 | 1 | a | ||
| 9 | 邵氏裸胸鯙Gymnothorax shaoi | DD | 肉食性 | 1 | a | ||
| 10 | 密点裸胸鳝Gymnothorax thyrsoideus | LC | 肉食性 | 1 | d, e | ||
| 11 | 波纹裸胸鳝Gymnothorax undulatus | LC | 肉食性 | 1 | f | ||
| 12 | 颌针鱼科Belonidae | 黑背圆颌针鱼Tylosurus acus | LC | 肉食性 | 1 | f | |
| 13 | 鳄形圆颌针鱼Tylosurus crocodilus | LC | 肉食性 | 1 | b, f | ||
| 14 | 飞鱼科Exocoetidae | 翼髭须唇飞鱼Cheilopogon pinnatibarbatus | LC | 肉食性 | 1 | f | |
| 15 | 细头燕鳐鱼Cypselurus angusticeps | NE | 杂食性 | 1 | f | ||
| 16 | 鳂科Holocentridae | 红双棘鳂Dispinus ruber | LC | 肉食性 | 1 | d | |
| 17 | 黄鳍锯鳞鱼Myripristis chryseres | LC | 肉食性 | 1 | 1 | b, f | |
| 18 | 格氏锯鳞鱼Myripristis greenfieldi | NE | 杂食性 | 1 | f | ||
| 19 | 康德锯鳞鱼Myripristis kuntee | LC | 肉食性 | 1 | d | ||
| 20 | 白边锯鳞鱼Myripristis murdjan | LC | 肉食性 | 1 | 1 | a, d | |
| 21 | 红锯鳞鱼Myripristis pralinia | LC | 肉食性 | 1 | f | ||
| 22 | 紫红锯鳞鱼Myripristis violacea | LC | 肉食性 | 1 | a | ||
| 23 | 黑鳍新东洋鳂Neoniphon opercularis | LC | 肉食性 | 1 | a, f | ||
| 24 | 莎姆新东洋鳂Neoniphon sammara | LC | 肉食性 | 1 | 1 | a, d, f | |
| 25 | 尾斑棘鳞鱼Sargocentron caudimaculatum | LC | 肉食性 | 1 | 1 | a, b, c, f | |
| 26 | 黑鳍棘鳞鱼Sargocentron diadema | LC | 肉食性 | 1 | d | ||
| 27 | 点带棘鳞鱼Sargocentron rubrum | LC | 肉食性 | 1 | f | ||
| 28 | 尖吻棘鳞鱼Sargocentron spiniferum | LC | 肉食性 | 1 | 1 | b, c, d | |
| 29 | 鲱科Clupeidae | 锈眼小体鲱Spratelloides delicatulus | LC | 肉食性 | 1 | f | |
| 30 | 银带小体鲱Spratelloides gracilis | LC | 肉食性 | 1 | f | ||
| 31 | 管口鱼科Aulostomidae | 中华管口鱼Aulostomus chinensis | LC | 肉食性 | 1 | c | |
| 32 | 烟管鱼科Fistulariidae | 鳞烟管鱼Fistularia petimba | LC | 肉食性 | 1 | d | |
| 33 | 灯笼鱼科Myctophidae | 长鳍虹灯鱼Bolinichthys longipes | LC | 肉食性 | 1 | f | |
| 34 | 刺尾鱼科Acanthuridae | 日本刺尾鱼Acanthurus japonicus | LC | 植食性 | 1 | a, c | |
| 35 | 纵带刺尾鱼Acanthurus lineatus | LC | 植食性 | 1 | a, f | ||
| 36 | 白面刺尾鱼Acanthurus nigricans | LC | 植食性 | 1 | d | ||
| 37 | 黑尾刺尾鱼Acanthurus nigricauda | LC | 植食性 | 1 | f | ||
| 38 | 褐斑刺尾鱼Acanthurus nigrofuscus | LC | 植食性 | 1 | 1 | b, d, f | |
| 39 | 橙斑刺尾鱼Acanthurus olivaceus | LC | 植食性 | 1 | 1 | d, f | |
| 40 | 黑鳃刺尾鱼Acanthurus pyroferus | LC | 植食性 | 1 | c, f | ||
| 41 | 横带刺尾鱼Acanthurus triostegus | LC | 植食性 | 1 | 1 | a, b, d, f | |
| 42 | 双斑栉齿刺尾鱼Ctenochaetus binotatus | LC | 植食性 | 1 | f | ||
| 43 | 栉齿刺尾鱼Ctenochaetus striatus | LC | 植食性 | 1 | a, c, f | ||
| 44 | 短吻鼻鱼Naso brevirostris | LC | 植食性 | 1 | c | ||
| 45 | 六棘鼻鱼Naso hexacanthus | LC | 植食性 | 1 | c, f | ||
| 46 | 颊吻鼻鱼Naso lituratus | LC | 植食性 | 1 | 1 | a, c, d, f | |
| 47 | 方吻鼻鱼Naso mcdadei | LC | 植食性 | 1 | f | ||
| 48 | 拟鲔鼻鱼Naso thynnoides | LC | 植食性 | 1 | d | ||
| 49 | 单角鼻鱼Naso unicornis | LC | 植食性 | 1 | a, c, f | ||
| 50 | 丝尾鼻鱼Naso vlamingii | LC | 植食性 | 1 | c | ||
| 51 | 小高鳍刺尾鱼Zebrasoma scopas | LC | 植食性 | 1 | c | ||
| 52 | 天竺鲷科Apogonidae | 巨牙天竺鲷Cheilodipterus macrodon | NE | 肉食性 | 1 | c | |
| 53 | 五带巨牙天竺鲷Cheilodipterus quinquelineatus | NE | 肉食性 | 1 | f | ||
| 54 | 史密斯氏天竺鲷Jaydia smithi | NE | 肉食性 | 1 | f | ||
| 55 | 丽鳍棘眼天竺鲷Pristiapogon kallopterus | NE | 肉食性 | 1 | f | ||
| 56 | 鳚科Blenniidae | 颊纹穗肩鳚Cirripectes castaneus | LC | 杂食性 | 1 | f | |
| 57 | 紫黑穗肩鳚Cirripectes imitator | LC | 杂食性 | 1 | f | ||
| 58 | 黑带稀棘鳚Meiacanthus grammistes | LC | 杂食性 | 1 | c | ||
| 59 | 细纹凤鳚Salarias fasciatus | LC | 杂食性 | 1 | f | ||
| 60 | 梅鲷科Caesionidae | 双带鳞鳍梅鲷Pterocaesio digramma | LC | 肉食性 | 1 | d | |
| 61 | 马氏鳞鳍梅鲷Pterocaesio marri | LC | 肉食性 | 1 | f | ||
| 62 | 黑带鳞鳍梅鲷Pterocaesio tile | LC | 肉食性 | 1 | c, f | ||
| 63 | 三带鳞鳍梅鲷Pterocaesio trilineata | LC | 肉食性 | 1 | d | ||
| 64 | 鲹科Carangidae | 黑尻鲹Caranx melampygus | LC | 肉食性 | 1 | d | |
| 65 | 颌圆鲹Decapterus macarellus | LC | 肉食性 | 1 | c, f | ||
| 66 | 长身圆鲹Decapterus macrosoma | LC | 肉食性 | 1 | d | ||
| 67 | 高体鰤Seriola dumerili | LC | 肉食性 | 1 | d, e | ||
| 68 | 蝴蝶鱼科Chaetodontidae | 项斑蝴蝶鱼Chaetodon adiergastos | LC | 杂食性 | 1 | d | |
| 69 | 丝蝴蝶鱼Chaetodon auriga | LC | 杂食性 | 1 | a, c, f | ||
| 70 | 叉纹蝴蝶鱼Chaetodon auripes | LC | 杂食性 | 1 | f | ||
| 71 | 双丝蝴蝶鱼Chaetodon bennetti | DD | 杂食性 | 1 | c | ||
| 72 | 鞭蝴蝶鱼Chaetodon ephippium | LC | 杂食性 | 1 | c, f | ||
| 73 | 珠蝴蝶鱼Chaetodon kleinii | LC | 杂食性 | 1 | c | ||
| 74 | 新月蝴蝶鱼Chaetodon lunula | LC | 杂食性 | 1 | c, f | ||
| 75 | 弓月蝴蝶鱼Chaetodon lunulatus | LC | 杂食性 | 1 | a, c | ||
| 76 | 黑背蝴蝶鱼Chaetodon melannotus | LC | 杂食性 | 1 | c | ||
| 77 | 华丽蝴蝶鱼Chaetodon ornatissimus | LC | 杂食性 | 1 | f | ||
| 78 | 斑带蝴蝶鱼Chaetodon punctatofasciatus | LC | 杂食性 | 1 | c | ||
| 79 | 格纹蝴蝶鱼Chaetodon rafflesii | LC | 杂食性 | 1 | f | ||
| 80 | 镜斑蝴蝶鱼Chaetodon speculum | LC | 杂食性 | 1 | c | ||
| 81 | 三纹蝴蝶鱼Chaetodon trifascialis | NT | 杂食性 | 1 | a | ||
| 82 | 三带蝴蝶鱼Chaetodon trifasciatus | LC | 杂食性 | 1 | b, d | ||
| 83 | 乌利蝴蝶鱼Chaetodon ulietensis | LC | 杂食性 | 1 | c | ||
| 84 | 斜纹蝴蝶鱼Chaetodon vagabundus | LC | 杂食性 | 1 | a | ||
| 85 | 黄蝴蝶鱼Chaetodon xanthurus | LC | 杂食性 | 1 | a, c | ||
| 86 | 黄镊口鱼Forcipiger flavissimus | LC | 杂食性 | 1 | c | ||
| 87 | 多鳞霞蝶鱼Hemitaurichthys polylepis | LC | 杂食性 | 1 | c | ||
| 88 | 马夫鱼Heniochus acuminatus | LC | 杂食性 | 1 | d | ||
| 89 | 金口马夫鱼Heniochus chrysostomus | LC | 杂食性 | 1 | c | ||
| 90 | 鱼翁科Cirrhitidae | 翼鱼翁Cirrhitus pinnulatus | LC | 肉食性 | 1 | a | |
| 91 | 副鱼翁Paracirrhites arcatus | LC | 肉食性 | 1 | c | ||
| 92 | 蛇鲭科Gempylidae | 蛇鲭Gempylus serpens | LC | 肉食性 | 1 | f | |
| 93 | 鰕虎科Gobiidae | 丝条凡塘鳢Valenciennea strigata | LC | 肉食性 | 1 | c | |
| 94 | 仿石鲈科Haemulidae | 斑胡椒鲷Plectorhinchus chaetodonoides | NE | 肉食性 | 1 | c, f | |
| 95 | 鮀科Kyphosidae | 低鳍[鱼舵]Kyphosus vaigiensis | LC | 植食性 | 1 | f | |
| 96 | 隆头鱼科Labridae | 乌尾阿南鱼Anampses melanurus | LC | 肉食性 | 1 | c | |
| 97 | 黄尾阿南鱼Anampses meleagrides | LC | 肉食性 | 1 | 1 | c, d | |
| 98 | 星阿南鱼Anampses twistii | LC | 肉食性 | 1 | c | ||
| 99 | 腋斑普提鱼Bodianus axillaris | LC | 肉食性 | 1 | c | ||
| 100 | 双带普提鱼Bodianus bilunulatus | LC | 肉食性 | 1 | c | ||
| 101 | 绿尾唇鱼Cheilinus chlorourus | LC | 肉食性 | 1 | a | ||
| 102 | 横带唇鱼Cheilinus fasciatus | LC | 肉食性 | 1 | 1 | b, c, d | |
| 103 | 尖头唇鱼Cheilinus oxycephalus | LC | 肉食性 | 1 | c | ||
| 104 | 三叶唇鱼Cheilinus trilobatus | LC | 肉食性 | 1 | a | ||
| 105 | 黑缘丝隆头鱼Cirrhilabrus melanomarginatus | LC | 肉食性 | 1 | c | ||
| 106 | 鳃斑盔鱼Coris aygula | LC | 肉食性 | 1 | c | ||
| 107 | 露珠盔鱼Coris gaimard | LC | 肉食性 | 1 | c, f | ||
| 108 | 伸口鱼Epibulus insidiator | LC | 肉食性 | 1 | c | ||
| 109 | 杂色尖嘴鱼Gomphosus varius | LC | 肉食性 | 1 | c, f | ||
| 110 | 双眼斑海猪鱼Halichoeres biocellatus | LC | 肉食性 | 1 | c | ||
| 111 | 金色海猪鱼Halichoeres chrysus | LC | 肉食性 | 1 | c | ||
| 112 | 格纹海猪鱼Halichoeres hortulanus | LC | 肉食性 | 1 | 1 | c, d | |
| 113 | 星云海猪鱼Halichoeres nebulosus | LC | 肉食性 | 1 | f | ||
| 114 | 横带厚唇鱼Hemigymnus fasciatus | LC | 肉食性 | 1 | c | ||
| 115 | 黑鳍厚唇鱼Hemigymnus melapterus | LC | 肉食性 | 1 | c, f | ||
| 116 | 环纹细鳞盔鱼Hologymnosus annulatus | LC | 肉食性 | 1 | c | ||
| 117 | 双色裂唇鱼Labroides bicolor | LC | 肉食性 | 1 | c | ||
| 118 | 裂唇鱼Labroides dimidiatus | LC | 肉食性 | 1 | c | ||
| 119 | 多纹褶唇鱼Labropsis xanthonota | LC | 肉食性 | 1 | c | ||
| 120 | 珠斑大咽齿鱼Macropharyngodon meleagris | LC | 肉食性 | 1 | c | ||
| 121 | 带尾美鳍鱼Novaculichthys taeniourus | LC | 肉食性 | 1 | 1 | d, f | |
| 122 | 西里伯斯唇鱼Oxycheilinus celebicus | LC | 肉食性 | 1 | f | ||
| 123 | 双线尖唇鱼Oxycheilinus digramma | LC | 肉食性 | 1 | c | ||
| 124 | 单带尖唇鱼Oxycheilinus unifasciatus | LC | 肉食性 | 1 | c | ||
| 125 | 姬拟唇鱼Pseudocheilinus evanidus | LC | 肉食性 | 1 | c | ||
| 126 | 八带拟唇鱼Pseudocheilinus octotaenia | LC | 肉食性 | 1 | c | ||
| 127 | 黑星紫胸鱼Stethojulis bandanensis | LC | 肉食性 | 1 | f | ||
| 128 | 钝头锦鱼Thalassoma amblycephalum | LC | 肉食性 | 1 | c, f | ||
| 129 | 鞍斑锦鱼Thalassoma hardwicke | LC | 肉食性 | 1 | c | ||
| 130 | 新月锦鱼Thalassoma lunare | LC | 肉食性 | 1 | c | ||
| 131 | 纵纹锦鱼Thalassoma quinquevittatum | LC | 肉食性 | 1 | c | ||
| 132 | 尖吻鲈科Latidae | 尖吻鲈Lates calcarifer | LC | 肉食性 | 1 | b | |
| 133 | 裸颊鲷科Lethrinidae | 金带齿颌鲷Gnathodentex aureolineatus | LC | 肉食性 | 1 | 1 | a, b, c, d, f |
| 134 | 灰裸顶鲷Gymnocranius griseus | LC | 肉食性 | 1 | b, d, e | ||
| 135 | 扁裸颊鲷Lethrinus lentjan | LC | 肉食性 | 1 | d | ||
| 136 | 长吻裸颊鲷Lethrinus miniatus | LC | 肉食性 | 1 | b, d, e | ||
| 137 | 星斑裸颊鲷Lethrinus nebulosus | LC | 肉食性 | 1 | d | ||
| 138 | 桔带裸颊鲷Lethrinus obsoletus | LC | 肉食性 | 1 | f | ||
| 139 | 短吻裸颊鲷Lethrinus ornatus | LC | 肉食性 | 1 | f | ||
| 140 | 红裸颊鲷Lethrinus rubrioperculatus | LC | 肉食性 | 1 | f | ||
| 141 | 杂色裸颊鲷Lethrinus variegatus | LC | 肉食性 | 1 | d | ||
| 142 | 黄唇裸颊鲷Lethrinus xanthochilus | LC | 肉食性 | 1 | f | ||
| 143 | 单列齿鲷Monotaxis grandoculis | LC | 肉食性 | 1 | c | ||
| 144 | 异牙单列齿鲷Monotaxis heterodon | LC | 肉食性 | 1 | c | ||
| 145 | 笛鲷科Lutjanidae | 叉尾鲷Aphareus furca | LC | 肉食性 | 1 | d | |
| 146 | 红叉尾鲷Aphareus rutilans | LC | 肉食性 | 1 | d, e | ||
| 147 | 蓝短鳍笛鲷Aprion virescens | LC | 肉食性 | 1 | b, c | ||
| 148 | 孟加拉国湾笛鲷Lutjanus bengalensis | LC | 肉食性 | 1 | f | ||
| 149 | 白斑笛鲷Lutjanus bohar | LC | 肉食性 | 1 | b, f | ||
| 150 | 隆背笛鲷Lutjanus gibbus | LC | 肉食性 | 1 | a | ||
| 151 | 四线笛鲷Lutjanus kasmira | LC | 肉食性 | 1 | b, d | ||
| 152 | 黄背若梅鲷Paracaesio xanthura | LC | 肉食性 | 1 | f | ||
| 153 | 黄鳍紫鱼Pristipomoides flavipinnis | LC | 肉食性 | 1 | d | ||
| 154 | 弱棘鱼科Malacanthidae | 短吻弱棘鱼Malacanthus brevirostris | NE | 肉食性 | 1 | b | |
| 155 | 侧条弱棘鱼Malacanthus latovittatus | NE | 肉食性 | 1 | d | ||
| 156 | 羊鱼科Mullidae | 黄带拟羊鱼Mulloidichthys flavolineatus | LC | 肉食性 | 1 | c | |
| 157 | 无斑拟羊鱼Mulloidichthys vanicolensis | LC | 肉食性 | 1 | c, f | ||
| 158 | 纵带羊鱼Mullus surmuletus | LC | 肉食性 | 1 | b, d | ||
| 159 | 条斑副绯鲤Parupeneus barberinus | LC | 肉食性 | 1 | 1 | c, d | |
| 160 | 圆口副绯鲤Parupeneus cyclostomus | LC | 肉食性 | 1 | b, d | ||
| 161 | 七棘副绯鲤Parupeneus heptacanthus | LC | 肉食性 | 1 | d | ||
| 162 | 多带副绯鲤Parupeneus multifasciatus | LC | 肉食性 | 1 | a, c | ||
| 163 | 黑斑副绯鲤Parupeneus pleurostigma | LC | 肉食性 | 1 | 1 | b, c | |
| 164 | 三带副绯鲤Parupeneus trifasciatus | LC | 肉食性 | 1 | b, d | ||
| 165 | 金线鱼科Nemipteridae | 双带眶棘鲈Scolopsis bilineata | LC | 肉食性 | 1 | c | |
| 166 | 线纹眶棘鲈Scolopsis lineata | LC | 肉食性 | 1 | a, f | ||
| 167 | 三带眶棘鲈Scolopsis trilineata | LC | 肉食性 | 1 | c | ||
| 168 | 单鳍鱼科Pempheridae | 黑稍单鳍鱼Pempheris oualensis | NE | 肉食性 | 1 | d | |
| 169 | 黑缘单鳍鱼Pempheris vanicolensis | NE | 肉食性 | 1 | c | ||
| 170 | 肥足鰧科Pinguipedidae | 六睛拟鲈Parapercis hexophtalma | NE | 肉食性 | 1 | c | |
| 171 | 刺盖鱼科Pomacanthidae | 三点阿波鱼Apolemichthys trimaculatus | LC | 杂食性 | 1 | d | |
| 172 | 双棘刺尻鱼Centropyge bispinosa | LC | 杂食性 | 1 | f | ||
| 173 | 条尾刺尻鱼Centropyge fisheri | LC | 杂食性 | 1 | f | ||
| 174 | 海氏刺尻鱼Centropyge heraldi | LC | 杂食性 | 1 | c | ||
| 175 | 福氏刺尻鱼Centropyge vrolikii | LC | 杂食性 | 1 | c | ||
| 176 | 主刺盖鱼Pomacanthus imperator | LC | 杂食性 | 1 | c | ||
| 177 | 双棘甲尻鱼Pygoplites diacanthus | LC | 杂食性 | 1 | a, c | ||
| 178 | 雀鲷科Pomacentridae | 五带豆娘鱼Abudefduf vaigiensis | LC | 杂食性 | 1 | a, c, f | |
| 179 | 金凹牙豆娘鱼Amblyglyphidodon aureus | LC | 杂食性 | 1 | c | ||
| 180 | 腋斑光鳃鱼Pycnochromis atripes | LC | 杂食性 | 1 | c | ||
| 181 | 线纹光鳃鱼Pycnochromis lineatus | LC | 杂食性 | 1 | c | ||
| 182 | 双斑光鳃鱼Pycnochromis margaritifer | LC | 杂食性 | 1 | c | ||
| 183 | 卵形光鳃鱼Pycnochromis ovatiformis | LC | 杂食性 | 1 | c | ||
| 184 | 凡氏光鳃鱼Pycnochromis vanderbilti | LC | 杂食性 | 1 | c | ||
| 185 | 蓝绿光鳃鱼Chromis viridis | NE | 杂食性 | 1 | c | ||
| 186 | 黄尾光鳃鱼Chromis xanthura | NE | 杂食性 | 1 | c | ||
| 187 | 三斑宅泥鱼Dascyllus trimaculatus | NE | 杂食性 | 1 | 1 | a, c, d | |
| 188 | 显盘雀鲷Dischistodus perspicillatus | NE | 杂食性 | 1 | f | ||
| 189 | 眼斑椒雀鲷Stegastes lacrymatus | NE | 杂食性 | 1 | f | ||
| 190 | 安汶雀鲷Pomacentrus ambionensis | NE | 杂食性 | 1 | c | ||
| 191 | 霓虹雀鲷Pomacentrus coelestis | NE | 杂食性 | 1 | c, f | ||
| 192 | 黑缘雀鲷Pomacentrus nigromarginatus | NE | 杂食性 | 1 | c | ||
| 193 | 菲律宾雀鲷Pomacentrus philippinus | NE | 杂食性 | 1 | c | ||
| 194 | 王子雀鲷Pomacentrus vaiuli | NE | 杂食性 | 1 | c | ||
| 195 | 长吻眶锯雀鲷Stegastes lividus | NE | 杂食性 | 1 | f | ||
| 196 | 大眼鲷科Priacanthidae | 金目大眼鲷Priacanthus hamrur | LC | 肉食性 | 1 | d | |
| 197 | 拟雀鲷科Pseudochromidae | 蓝带拟雀鲷Pseudochromis cyanotaenia | LC | 肉食性 | 1 | f | |
| 198 | 小带鳚科Microdesmidae | 大口线塘鳢Nemateleotris magnifica | LC | 肉食性 | 1 | c | |
| 199 | 黑尾鳍塘鳢Ptereleotris evides | LC | 肉食性 | 1 | c | ||
| 200 | 尾斑鳍塘鳢Ptereleotris heteroptera | LC | 肉食性 | 1 | c | ||
| 201 | 鹦嘴鱼科Scaridae | 星眼绚鹦嘴鱼Calotomus carolinus | LC | 植食性 | 1 | f | |
| 202 | 日本绚鹦嘴鱼Calotomus japonicus | LC | 植食性 | 1 | b, d | ||
| 203 | 凹尾绚鹦嘴鱼Calotomus spinidens | LC | 植食性 | 1 | b, d | ||
| 204 | 双色鲸鹦嘴鱼Cetoscarus bicolor | LC | 植食性 | 1 | d | ||
| 205 | 蓝头绿鹦嘴鱼Chlorurus sordidus | LC | 植食性 | 1 | a, c | ||
| 206 | 尾斑绿鹦嘴鱼Chlorurus spilurus | LC | 植食性 | 1 | f | ||
| 207 | 长头马鹦嘴鱼Hipposcarus longiceps | LC | 植食性 | 1 | a, c | ||
| 208 | 弧带鹦嘴鱼Scarus dimidiatus | LC | 植食性 | 1 | d | ||
| 209 | 锈色鹦嘴鱼Scarus ferrugineus | LC | 植食性 | 1 | d | ||
| 210 | 绿唇鹦嘴鱼Scarus forsteni | LC | 植食性 | 1 | 1 | a, b, c, d, f | |
| 211 | 网纹鹦嘴鱼Scarus frenatus | LC | 植食性 | 1 | c | ||
| 212 | 青点鹦嘴鱼Scarus ghobban | LC | 植食性 | 1 | d | ||
| 213 | 黑斑鹦嘴鱼Scarus globiceps | LC | 植食性 | 1 | 1 | a, b, c, d | |
| 214 | 黑鹦嘴鱼Scarus niger | LC | 植食性 | 1 | 1 | c, d | |
| 215 | 黄鞍鹦嘴鱼Scarus oviceps | LC | 植食性 | 1 | 1 | a, b, d | |
| 216 | 突额鹦嘴鱼Scarus ovifrons | DD | 植食性 | 1 | f | ||
| 217 | 绿颌鹦嘴鱼Scarus prasiognathos | LC | 植食性 | 1 | d | ||
| 218 | 棕吻鹦嘴鱼Scarus psittacus | LC | 植食性 | 1 | 1 | a, d, f | |
| 219 | 截尾鹦嘴鱼Scarus rivulatus | LC | 植食性 | 1 | c | ||
| 220 | 许氏鹦嘴鱼Scarus schlegeli | LC | 植食性 | 1 | a, c | ||
| 221 | 刺鹦嘴鱼Scarus spinus | LC | 植食性 | 1 | f | ||
| 222 | 带尾鹦嘴鱼Scarus taeniopterus | LC | 植食性 | 1 | d | ||
| 223 | 鲭科Scombridae | 裸狐鲣Gymnosarda unicolor | LC | 肉食性 | 1 | b, c | |
| 224 | 鮨科Serranidae | 斑点九棘鲈Cephalopholis argus | LC | 肉食性 | 1 | 1 | a, b, c, d |
| 225 | 尾纹九棘鲈Cephalopholis urodeta | LC | 肉食性 | 1 | 1 | a, b, c, d | |
| 226 | 珊瑚石斑鱼Epinephelus corallicola | LC | 肉食性 | 1 | d | ||
| 227 | 横条石斑鱼Epinephelus fasciatus | LC | 肉食性 | 1 | b, d, e | ||
| 228 | 棕点石斑鱼Epinephelus fuscoguttatus | VU | 肉食性 | 1 | f | ||
| 229 | 六角石斑鱼Epinephelus hexagonatus | LC | 肉食性 | 1 | 1 | a, b, d | |
| 230 | 蜂巢石斑鱼Epinephelus merra | LC | 肉食性 | 1 | 1 | a, b, c, d | |
| 231 | 吻斑石斑鱼Epinephelus spilotoceps | LC | 肉食性 | 1 | a | ||
| 232 | 七带下美鮨Hyporthodus septemfasciatus | DD | 肉食性 | 1 | d | ||
| 233 | 点线鳃棘鲈Plectropomus oligacanthus | LC | 肉食性 | 1 | d | ||
| 234 | 刺盖拟花鮨Nemanthias dispar | LC | 杂食性 | 1 | c | ||
| 235 | 高体拟花鮨Pseudanthias hypselosoma | LC | 杂食性 | 1 | c | ||
| 236 | 紫红拟花鮨Mirolabrichthys pascalus | LC | 肉食性 | 1 | c | ||
| 237 | 篮子鱼科Siganidae | 星斑篮子鱼Siganus guttatus | LC | 植食性 | 1 | d | |
| 238 | 眼带篮子鱼Siganus puellus | LC | 植食性 | 1 | c | ||
| 239 | 斑篮子鱼Siganus punctatus | LC | 植食性 | 1 | f | ||
| 240 | 蠕纹篮子鱼Siganus vermiculatus | LC | 植食性 | 1 | a | ||
| 241 | 狐篮子鱼Siganus vulpinus | LC | 植食性 | 1 | 1 | c, b, d | |
| 242 | 鲷科Sparidae | 黄犁齿鲷Evynnis tumifrons | LC | 肉食性 | 1 | d, e | |
| 243 | 魣科Sphyraenidae | 大眼魣Sphyraena forsteri | NE | 肉食性 | 1 | d | |
| 244 | 鰧科Uranoscopidae | 土佐鰧Uranoscopus tosae | NE | 肉食性 | 1 | f | |
| 245 | 镰鱼科Zanclidae | 角镰鱼Zanclus cornutus | LC | 杂食性 | 1 | c, f | |
| 246 | 鲆科Bothidae | 豹纹鲆Bothus pantherinus | LC | 肉食性 | 1 | f | |
| 247 | 鲉科Scorpaenidae | 花彩圆鳞鲉Parascorpaena picta | LC | 肉食性 | 1 | b | |
| 248 | 须拟鲉Scorpaenopsis cirrosa | NE | 肉食性 | 1 | d | ||
| 249 | 玫瑰毒鲉Synanceia verrucosa | LC | 肉食性 | 1 | f | ||
| 250 | 鳞鲀科Balistidae | 褐拟鳞鲀Balistoides viridescens | NE | 杂食性 | 1 | b | |
| 251 | 黑边角鳞鲀Melichthys vidua | NE | 杂食性 | 1 | a, c, f | ||
| 252 | 红牙鳞鲀Odonus niger | NE | 杂食性 | 1 | a, b | ||
| 253 | 叉斑锉鳞鲀Rhinecanthus aculeatus | NE | 杂食性 | 1 | f | ||
| 254 | 黑带锉鳞鲀Rhinecanthus rectangulus | NE | 杂食性 | 1 | d | ||
| 255 | 黄鳍多棘鳞鲀Sufflamen chrysopterum | NE | 杂食性 | 1 | c | ||
| 256 | 缰纹多棘鳞鲀Sufflamen fraenatum | LC | 杂食性 | 1 | b | ||
| 257 | 金边黄鳞鲀Xanthichthys auromarginatus | NE | 杂食性 | 1 | c | ||
| 258 | 线斑黄鳞鲀Xanthichthys lineopunctatus | NE | 杂食性 | 1 | b | ||
| 259 | 单角鲀科Monacanthidae | 单角革鲀Aluterus monoceros | LC | 肉食性 | 1 | f | |
| 260 | 棘尾前孔鲀Cantherhines dumerilii | LC | 杂食性 | 1 | a, c, f | ||
| 261 | 细斑前孔鲀Cantherhines pardalis | LC | 杂食性 | 1 | c | ||
| 262 | 箱鲀科Ostraciidae | 粒突箱鲀Ostracion cubicus | NE | 肉食性 | 1 | c | |
| 263 | 白点箱鲀Ostracion meleagris | NE | 肉食性 | 1 | c | ||
| 264 | 鲀科Tetraodontidae | 纹腹叉鼻鲀Arothron hispidus | LC | 杂食性 | 1 | a | |
| 265 | 黑斑叉鼻鲀Arothron nigropunctatus | LC | 杂食性 | 1 | c | ||
| 266 | 星斑叉鼻鲀Arothron stellatus | LC | 杂食性 | 1 | a | ||
| 267 | 横带扁背鲀Canthigaster valentini | LC | 杂食性 | 1 | c | ||
| 268 | 六鳃鲨科Hexanchidae | 灰六鳃鲨Hexanchus griseus | NT | 肉食性 | 1 | d, e | |
| 269 | 中村氏六鳃鲨Hexanchus nakamurai | NT | 肉食性 | 1 | d, e | ||
| 270 | 魟科Dasyatidae | 迈氏条尾魟Taeniura meyeni | VU | 肉食性 | 1 | d, e | |
| EX:灭绝Extinct;EW:野生灭绝Extinction In The Wild;CR:极危Critically Endangered;VU:易危Vulnerable;NT:近危Near Threatened;LC:最少关注Least Concern;DD:数据不足Data deficient;NE:未评估Not Evaluated;a:水下潜捕;b:手钓;c:水下摄影;d:流刺网;e:延绳钓;f:eDNA | |||||||
不同科之间的鱼类种类数存在较大差异。其中, 有超过20种的科包括隆头鱼科(36种)、蝴蝶鱼科(22种)和鹦嘴鱼科(22种)。而有超过10种的科包括刺尾鱼科(18种)、雀鲷科(18种)、鮨科Serranidae (13种)等。此外, 有超过5种的科包括笛鲷科Lutjanidae (9种)、鳞鲀科Balistidae (9种)、羊鱼科Mullidae (9种)、刺盖鱼科(7种)和篮子鱼科Siganidae (5种)。根据CFDI计算, 预测北礁珊瑚礁鱼类的总物种数为396种。
2.2 体型结构北礁珊瑚礁鱼类的最大全长范围从6 cm到485 cm不等。将这些数据以10 cm为一个区间划分, 并统计不同区间物种的分布情况。在大于350 cm的范围中, 鱼类均不超过1种, 且仅有1组区间有物种分布。因此, 在体型结构分析中, 我们将这些异常值舍弃。总体结果显示, 物种数与最大全长呈指数相关。当最大全长在0—100 cm范围时, 随着最大全长的增加, 物种数下降最明显;而当最大全长超过100 cm时, 物种数的减少趋势相对缓慢。这一结果表明, 北礁鱼类物种主要分布在在最大全长为0—100 cm的范围内, 该范围内的鱼类物种数高达249种, 占总物种数的92.22%。
北礁珊瑚礁鱼类以小型鱼为主, 占总数的52.96%, 其次是中型鱼(30.74%)和大型鱼(16.30%)。肉食性鱼类占绝对优势, 占总数的56.67%, 其次是杂食性鱼类(26.30%)和植食性鱼类(17.04%)。大中型鱼类以肉食性为主, 分别占79.54%和59.04%, 小型鱼类则以杂食性和肉食性为主, 分别占48.25%和44.06%。植食性鱼类的比例最低, 仅占7.69%(图 4)。

|
| 图 3 北礁鱼类最大全长的物种数分布特征 Fig. 3 The distribution characteristics of species number at the maximum total length of coral reef fishes in Beijiao Atoll |

|
| 图 4 北礁不同大小和食性类型鱼类的分布特征 Fig. 4 Distribution characteristics of fishes with different sizes and feeding habits in Beijiao Atoll |
肉食性鱼类中小型鱼类最多, 占45.10%(69种), 其次为中型鱼类32.03%(49种), 大型鱼类最少22.88%(35种);植食性鱼类中中型鱼类60.87%(28种)占据优势, 其次为小型鱼类23.91%(11种)和大型鱼类15.22%(7种);杂食性鱼类中小型鱼类占绝对优势, 比例高达88.73%(63种), 中型鱼类8.45%(6种)和大型鱼类2.82%(2种)均较少(图 4)。三种食性鱼类的最大全长分布范围差异显著, 肉食性鱼类个体最大, 最大全长平均为(56.1±63.28) cm, 分布范围最大为6.00—482.00 cm;杂食性鱼类个体最小, 最大全长平均为(22.0±16.83) cm, 分布范围为6.00—120.00 cm;植食性鱼类最大全长平均为(45.4±16.61) cm, 分布范围最小为21.00—85.00 cm(图 5)。

|
| 图 5 北礁不同食性类型的鱼类大小特征 Fig. 5 Body size characteristics of fishes with different feeding habits in Beijiao Atoll |
随时间变化北礁珊瑚礁鱼类的平均分类差异指数和变异分类差异指数均呈现递减的现象, 其中Delta+(Δ+)为1998—2003(56.72)> 2023(54.57)(图 6), Lambda+(Λ+)为1998—2003(163.8)> 2023(107.6)(图 6)。并且与历史数据相比较, 北礁珊瑚礁鱼类目、科和属的数量与种类数的比例均明显下降, 目的比例减少了48.21%, 科的比例减少了34.67%, 属的比例减少了15.45%。而2023年与1998—2003年的目、科、属、种的相似性指数也均较低分别是0.50、0.37、0.23和0.10。不同功能群的相似性均表现为极不相似, 肉食性、植食性和杂食性分别为0.10、0.22和0.01, 大型鱼、中型鱼和小型鱼分别为0.00、0.13和0.11。不同时间阶段恋礁型鱼类组成的相似性表现为极不相似, 相似度为0.22。

|
| 图 6 北礁不同时间段分类多样性指数的95%置信区间漏斗图 Fig. 6 Funnel plots of 95% confidence limits for taxonomic diversity indices at different time periods in Beijiao Atoll |
与历史数据比相比, 在本研究中共有60种鱼类未采集到, 肉食性鱼类占据优势, 种类数为41种, 占未采集到鱼类的68.33%。在未采集到鱼类中, 不同体型的鱼类均以肉食性鱼类占比最多, 大型鱼中为16种(84.21%), 中型鱼中为15种(65.22%), 小型鱼中为10种(55.56%)。另外, 大型鱼类和肉食性鱼类在其功能群中的未采集到比例也是最高, 分别占其类群的43%和27%(图 7)。这些未采集到的鱼类有86.67%属于鲈形目、鳗鲡目、和鲀形目, 并且软骨鱼类均未再次采集到。另外, 北礁珊瑚礁鱼类中有5种鱼类列于IUCN红色名录, 其中保护状态为易危的有2种, 分别是棕点石斑鱼(Epinephelus fuscoguttatus)和迈氏条尾魟(Taeniura meyeni);保护状态为近危的有3种, 分别是三纹蝴蝶鱼(Chaetodon trifascialis)、中村氏六鳃鲨(Hexanchus nakamurai)和灰六鳃鲨(Hexanchus griseus), 其中位于IUCN红色名录中的鱼类有80%均为大型肉食性鱼类。

|
| 图 7 北礁珊瑚礁鱼类各功能群未采集到的比例 Fig. 7 Proportion of fish functional groups not collected in Beijiao Atoll |
北礁与中国南海西沙群岛其他岛屿相比, 物种数虽然不及永乐环礁(336种)[18]、七连屿(315种)[16]和永兴岛(341种)[17], 但却明显高于单个岛屿的东岛(235种)[15], 可能与北礁具有潟湖有关, 具有更复杂的珊瑚礁结构、微生境和食物获得性, 更适宜作为鱼类的产卵场、育幼场和摄食场[24]。同时, 根据Coral Fish Diversity Index(CFDI)预测北礁珊瑚礁鱼类总物种数为396种[23], 有126个物种尚未被发现, 说明北礁可能拥有非常丰富的珊瑚礁鱼类资源。另外, 西沙群岛位于世界生物多样性最高的“珊瑚礁三角区”北缘, 二者珊瑚礁鱼类物种组成非常相似, 研究表明印度太平洋珊瑚礁鱼类主要由蝴蝶鱼科、隆头鱼科及鹦嘴鱼科等29科鱼类组成[25]。北礁珊瑚礁鱼类中除海龙科, 其余均有分布, 且占总物种数的89.63%, 这一比例高于西沙群岛其他岛屿, 如东岛除拟雀鲷科、金线鱼科、小带鳚科和海龙科未发现, 其余科占总物种数的82.13%[15], 七连屿除小带鳚科未发现, 其余科占总物种数的89.52%[16]。说明北礁珊瑚礁鱼类资源非常丰富, 更加贴近印度太平洋珊瑚礁鱼类的特征。
尽管两个时期在北礁的鱼类资源调查中采用了不同的采样方式, 但在1998—2003年间主要采用的延绳钓仅获得了12种鱼类样本。相比之下, 流刺网(采样深度:5—25 m)和手钓(采样深度:5—40 m)成为了这一时期的主要采样方式, 其采集到的鱼类样本涵盖了延绳钓所采集的种类。至于2023年的四种采样方式则覆盖了水深0—60 m的范围, 其中潜水捕捞和水下视觉普查技术可以更加精细和准确地调查北礁珊瑚礁鱼类的组成情况, 尤其是一些数量较少且隐蔽性强的物种, 并且使用了eDNA方法进行补充[26]。因此, 第二个时期采用的方法不仅可以达到第一个时期的采集效果, 而且在精度和准确性上可能更胜一筹。
3.2 不同体型和食性鱼类的变化北礁珊瑚礁鱼类以肉食性鱼类为主, 符合早期珊瑚礁鱼类大多以肉食性物种为主的研究[27]。但在2023年调查与历史数据的比较中, 肉食性鱼类在未采集到鱼类中比例(27%)明显高于其他食性, 说明肉食性鱼类受到的扰动最大。并且, 在北礁位于IUCN红色名录中的鱼类有大部分均为顶级肉食性鱼类, 这也进一步证明了肉食性鱼类更容易消亡, 符合当前渔业活动导致生态系统营养级下降的规律[15]。另外, 与历史数据相比软骨鱼类在本次调查中均未采集到, 这也反映了软骨鱼类难以抵挡气候变化和人类扰动影响等多重压力。有研究表明, 全球有近三分之二的珊瑚礁鲨和鳐面临灭绝的威胁[28]。肉食性鱼类的减少似乎已经成为全世界珊瑚礁鱼类所经历的演变, 但值得注意的是在肉食性鱼类明显下降的同时, 植食性鱼类的影响却不是那么明显, 在西沙群岛的永乐环礁和东岛均出现了肉食性鱼类下降而植食性鱼类上升的现象[15, 18]。有国外学者对澳大利亚大堡礁的22个独立珊瑚礁上不同程度珊瑚衰退时鱼类群落的反映进行了调查, 发现鱼类多样性很少因珊瑚减少而减少, 原因是珊瑚礁衰退后草食性鱼类受益于藻类生长而丰度增加, 从而鱼类多样性的维持掩盖了鱼类群落的重大变化[29—30]。因此, 要重视鱼类群落结构的变化, 短期来看渔业捕捞向植食性鱼类转变对渔业发现价值显著, 但长期来看这也会破坏生态系统的平衡与功能[31—32]。
在与历史调查数据的对比分析中发现大型鱼的未采集到比例(43%)最高, 说明北礁珊瑚礁鱼类出现小型化的现象。这与当前世界珊瑚礁鱼类体型的变化趋势相符, 在全世界范围内均出现了由于气候变化及人类扰动造成的鱼类小型化现象[33—34]。有学者研究了38个太平洋岛屿的珊瑚礁鱼类群落, 发现均出现了大型鱼类生物量减少的现象, 并且有人居住的岛屿这种现象更加明显[33]。并且有研究预测在21世纪北太平洋中部大型鱼类数量的下降幅度可能将高达75.8%[35]。北礁出现这种现象的原因可能有以下几种:第一, 大型鱼类具有较高的经济价值, 是主要的目标捕捞物种。并且大型鱼类的生长缓慢、性成熟晚等特点决定了在种群受到冲击时其难以快速补充[36]。北礁处于西沙群岛最北缘, 距离大陆最近, 是多数大陆渔民首先到达的地方, 因此北礁可能承担了较大的捕捞压力。第二, 小型鱼更能适应环境。小型鱼游泳能力较弱, 但对环境的需求更低, 相同的生境范围和食物资源可能满足更多的小型鱼类[37]。北礁的小型珊瑚礁鱼类种类非常丰富, 占到总种类数的一半以上。第三, 环境因素的变化也是鱼类小型化的一个因素。目前全球气候变化对海洋鱼类群落造成了影响, 一些研究表明高温与鱼类体型变小有密切关系[38]。有研究发现西沙群岛在近56年气温呈现上升趋势, 平均增暖速率为0.19℃/10a, 并且在近10年差异显著, 平均增暖速率为0.38℃/10a[39]。第四, 栖息地的丧失。近几十年来由于气候变化和人类活动导致珊瑚大量死亡, 失去了三维立体结构和作为鱼类栖息地的功能, 导致鱼类群落结构发生改变[40]。珊瑚礁的健康程度与鱼类群落息息相关, 有学者研究了澳大利亚大堡礁在生境退化和恢复之后的15年内鱼类群落的变化, 发现鱼类群落丰富度和珊瑚物种恢复趋势一致[41]。同时, 本次调查的大型鱼类组成与历史调查数据的相似度为极不相似, 这也进一步证明了大型鱼类种群受到了干扰。值得注意的是恋礁型鱼类与历史调查的相似度也很低, 这些鱼类种群的变化可能反应了北礁珊瑚礁生境受到了影响, 同时, 我们在水下影像中发现了长棘海星(Acanthaster planci)的踪迹, 这也验证了北礁珊瑚礁生境正在出现衰退, 因此需要引起重视并采取必要手段对珊瑚礁生境进行保护。
3.3 鱼类分类多样性降低随着时间的推移北礁珊瑚礁鱼类的平均分类差异指数和变异分类差异指数均表现出了递减的趋势, 平均分类差异指数的降低说明北礁珊瑚礁鱼类的分类学距离越来越近, 即亲缘关系更近[42]。这与南海珊瑚礁鱼类分类多样性研究结果相符, 有研究发现南海珊瑚礁鱼类比东海陆架鱼类的平均分类差异指数更低, 反映了珊瑚礁鱼类在亲缘关系上更为接近[43]。2023年的平均分类差异指数显著低于95%置信区间的下边界, 这意味着环境受到了扰动。另外, 变异分类差异指数的降幅更加明显, 表明北礁珊瑚礁鱼类群落间物种分类关系的均匀度升高。可能是由于北礁生态环境异质性的降低减少了不同物种的共存机会[44]。很多研究均发现了这一现象, 墨西哥湾盆地南部的Terminos Lagoon的鱼类分类多样性在几十年来一直呈现单调下降趋势, 表明其鱼类群落结构在长期尺度上表现出分类树中分支的减少和物种的同质化[45]。另外, 北礁珊瑚礁鱼类的目、科和属的数量与种类数的比例均明显下降, 这也表明北礁目前珊瑚礁鱼类亲缘关系更近, 分类同质化明显。同时, 在本研究中不同时段鱼类物种组成相似度均为不相似和极不相似, 表明北礁珊瑚礁鱼类演替和变迁非常显著。这些变化可能与气候异常和人类活动导致的栖息地丧失有关, 但这些影响不是单向的, 鱼类明显的更替和亲缘距离的减少也会反过来影响环境的稳定性, 降低生态系统的抗压能力[14]。
4 结论本研究整理北礁珊瑚礁鱼类物种名录, 分析鱼类物种组成的变化。西沙群岛北礁的珊瑚礁鱼类资源已经发生了明显的变化, 其中包括物种组成的改变以及群落结构的转变。与历史数据相比, 分类差异指数和变异分类差异指数显著下降, 大型和肉食性鱼类的数量也明显减少。此外, 综合考虑不同时期的相似性分析, 北礁珊瑚礁鱼类物种组成已经发生了重要变化。这些发现表明了北礁珊瑚礁鱼类的一种由肉食性向植食性、大型鱼向小型鱼、以及复杂性向简单性的演变趋势。因此, 对北礁珊瑚礁鱼类的研究不仅有助于更好地了解其物种组成和演变趋势, 而且对于维护和恢复珊瑚礁生态系统的健康具有重要意义。
| [1] |
Fisher R, O'Leary R A, Low-Choy S, Mengersen K, Knowlton N, Brainard R E, Caley M J. Species richness on coral reefs and the pursuit of convergent global estimates. Current Biology: CB, 2015, 25(4): 500-505. DOI:10.1016/j.cub.2014.12.022 |
| [2] |
Robinson J P W, Wilson S K, Robinson J, Gerry C, Lucas J, Assan C, Govinden R, Jennings S, Graham N A J. Productive instability of coral reef fisheries after climate-driven regime shifts. Nature Ecology & Evolution, 2019, 3: 183-190. |
| [3] |
Shi J, Li C H, Wang T, Zhao J F, Liu Y, Xiao Y Y. Distribution pattern of coral reef fishes in China. Sustainability, 2022, 14(22): 15107. DOI:10.3390/su142215107 |
| [4] |
Coker D J, Wilson S K, Pratchett M S. Importance of live coral habitat for reef fishes. Reviews in Fish Biology and Fisheries, 2014, 24(1): 89-126. DOI:10.1007/s11160-013-9319-5 |
| [5] |
Lamb J B, Willis B L, Fiorenza E A, Couch C S, Howard R, Rader D N, True J D, Kelly L A, Ahmad A, Jompa J, Harvell C D. Plastic waste associated with disease on coral reefs. Science, 2018, 359(6374): 460-462. DOI:10.1126/science.aar3320 |
| [6] |
McCallen E, Knott J, Nunez-Mir G, Taylor B, Jo I, Fei S L. Trends in ecology: shifts in ecological research themes over the past four decades. Frontiers in Ecology and the Environment, 2019, 17(2): 109-116. DOI:10.1002/fee.1993 |
| [7] |
Camp E, Schoepf V, Mumby P, Hardtke L, Rodolfo-Metalpa R, Smith D J, Suggett D. The future of coral reefs subject to rapid climate change: lessons from natural extreme environments. Frontiers in Marine Science, 2018, 5: 4. DOI:10.3389/fmars.2018.00004 |
| [8] |
Cheal A J, MacNeil M A, Emslie M J, Sweatman H. The threat to coral reefs from more intense cyclones under climate change. Global Change Biology, 2017, 23(4): 1511-1524. DOI:10.1111/gcb.13593 |
| [9] |
Khalaf M, Crosby M P. Assemblage structure of butterflyfishes and their use as indicators of Gulf of Aqaba benthic habitat in Jordan. Aquatic Conservation: Marine and Freshwater Ecosystems, 2005, 15(S1): S27-S43. DOI:10.1002/aqc.698 |
| [10] |
Kroon F J, Lefèvre C D, Doyle J R, Patel F, Milton G, Severati A, Kenway M, Johansson C L, Schnebert S, Thomas-Hall P, Bonin M C, Cameron D S, Westcott D A. DNA-based identification of predators of the corallivorous Crown-of-Thorns Starfish (Acanthaster cf. solaris) from fish faeces and gut contents. Scientific Reports, 2020, 10: 8184. DOI:10.1038/s41598-020-65136-4 |
| [11] |
Edwards C B, Friedlander A M, Green A G, Hardt M J, Sala E, Sweatman H P, Williams I D, Zgliczynski B, Sandin S A, Smith J E. Global assessment of the status of coral reef herbivorous fishes: evidence for fishing effects. Proceedings Biological Sciences, 2013, 281(1774): 20131835. |
| [12] |
Pratchett M S, Cumming G S. Managing cross-scale dynamics in marine conservation: pest irruptions and lessons from culling of crown-of-thorns starfish (Acanthaster spp.). Biological Conservation, 2019, 238: 108211. DOI:10.1016/j.biocon.2019.108211 |
| [13] |
Syms C, Jones G P. Disturbance, habitat structure, and the dynamics of a coral-reef fish community. Ecology, 2000, 81(10): 2714. DOI:10.1890/0012-9658(2000)081[2714:DHSATD]2.0.CO;2 |
| [14] |
Arai T. Diversity and conservation of coral reef fishes in the Malaysian South China Sea. Reviews in Fish Biology and Fisheries, 2015, 25(1): 85-101. DOI:10.1007/s11160-014-9371-9 |
| [15] |
王腾, 李纯厚, 赵金发, 石娟, 于洋飞, 肖雅元, 刘永. 西沙东岛珊瑚礁鱼类种类组成特征与演变. 水生生物学报, 2023, 47(9): 1456-1463. |
| [16] |
王腾, 刘永, 全秋梅, 肖雅元, 吴鹏, 李纯厚. 七连屿珊瑚礁鱼类种类组成特征分析. 中国水产科学, 2022, 29(1): 102-117. |
| [17] |
王腾, 刘永, 李纯厚, 肖雅元, 林琳, 李纯然, 谢雨芳, 吴鹏. 永兴岛附近海域珊瑚礁鱼类群落结构特征. 水生生物学报, 2023, 47(4): 674-683. |
| [18] |
Zhao J F, Li C H, Wang T, Shi J, Song X Y, Liu Y. Composition and long-term variation characteristics of coral reef fish species in Yongle atoll, Xisha Islands, China. Biology, 2023, 12(8): 1062. DOI:10.3390/biology12081062 |
| [19] |
Liao Y Z, Miao X, Wang R, Zhang R, Li H, Lin L S. First pelagic fish biodiversity assessment of Cosmonaut Sea based on environmental DNA. Marine Environmental Research, 2023, 192: 106225. DOI:10.1016/j.marenvres.2023.106225 |
| [20] |
Mouillot D, Laune J, Tomasini J A, Aliaume C, Brehmer P, Dutrieux E, Do Chi T. Assessment of coastal lagoon quality with taxonomic diversity indices of fish, Zoobenthos and macrophyte communities. Hydrobiologia, 2005, 550(1): 121-130. DOI:10.1007/s10750-005-4368-y |
| [21] |
Clarke K R, Warwick R M. A further biodiversity index applicable to species lists: variation in taxonomic distinctness. Marine Ecology Progress Series, 2001, 216: 265-278. DOI:10.3354/meps216265 |
| [22] |
Zintzen V, Anderson M J, Roberts C D, Diebel C E. Increasing variation in taxonomic distinctness reveals clusters of specialists in the deep sea. Ecography, 2011, 34(2): 306-317. DOI:10.1111/j.1600-0587.2010.06546.x |
| [23] |
Allen G R. Reef and shore fishes of Milne Bay Province, Papua New Guinea[C]//Werner T B, Allen G R (Eds. ), A rapid biodiversity assessment of the coral reefs of Milne Bay Province, Papua New Guinea, 1988.
|
| [24] |
Gladstone W. Selection of a spawning aggregation site by Chromis hypsilepis (Pisces: Pomacentridae): habitat structure, transport potential, and food availability. Marine Ecology Progress Series, 2007, 351: 235-247. DOI:10.3354/meps07108 |
| [25] |
Allen G R. Indo-Pacific coral-reef fishes as indicators of conservation hotspots[C]//Moosa MK, Soemodihardjo S, Soegiarto A, (Eds. ), Proceedings of the Ninth International Coral Reef Symposium, 23-27 October 2000, Bali, Indonesia. 2002(2): 921-926.
|
| [26] |
Rey A, Viard F, Lizé A, Corre E, Valentini A, Thiriet P. Coastal rocky reef fish monitoring in the context of the Marine Strategy Framework Directive: environmental DNA metabarcoding complements underwater visual census. Ocean & Coastal Management, 2023, 241: 106625. |
| [27] |
Valentine J F, Heck K L. Perspective review of the impacts of overfishing on coral reef food web linkages. Coral Reefs, 2005, 24(2): 209-213. DOI:10.1007/s00338-004-0468-9 |
| [28] |
Sherman C S, Simpfendorfer C A, Pacoureau N, Matsushiba J H, Yan H F, Walls R H L, Rigby C L, VanderWright W J, Jabado R W, Pollom R A, Carlson J K, Charvet P, Bin Ali A, Fahmi, Cheok J, Derrick D H, Herman K B, Finucci B, Eddy T D, Palomares M L D, Avalos-Castillo C G, Kinattumkara B, Blanco-Parra M D P, Dharmadi, Espinoza M, Fernando D, Haque A B, Mejía-Falla P A, Navia A F, Pérez-Jiménez J C, Utzurrum J, Yuneni R R, Dulvy N K. Half a century of rising extinction risk of coral reef sharks and rays. Nature Communications, 2023, 14(1): 15. DOI:10.1038/s41467-022-35091-x |
| [29] |
Cheal A J, Wilson S K, Emslie M J, Dolman A M, Sweatman H. Responses of reef fish communities to coral declines on the Great Barrier Reef. Marine Ecology Progress Series, 2008, 372: 211-223. DOI:10.3354/meps07708 |
| [30] |
Lindahl U, Ohman M C, Schelten C K. The 1997/1998 mass mortality of corals: effects on fish communities on a Tanzanian coral reef. Marine Pollution Bulletin, 2001, 42(2): 127-131. DOI:10.1016/S0025-326X(00)00167-3 |
| [31] |
Milne R, Bauch C T, Anand M. Local overfishing patterns have regional effects on health of coral, and economic transitions can promote its recovery. Bulletin of Mathematical Biology, 2022, 84(4): 46. DOI:10.1007/s11538-022-01000-y |
| [32] |
王腾, 石娟, 于洋飞, 赵金发, 肖雅元, 刘永, 李纯厚. 西沙珊瑚礁鱼类研究进展和保护建议. 生态学杂志, 2023, 42(7): 1755-1763. |
| [33] |
Robinson J P W, Williams I D, Edwards A M, McPherson J, Yeager L, Vigliola L, Brainard R E, Baum J K. Fishing degrades size structure of coral reef fish communities. Global Change Biology, 2017, 23(3): 1009-1022. DOI:10.1111/gcb.13482 |
| [34] |
Zgliczynski B J, Sandin S A. Size-structural shifts reveal intensity of exploitation in coral reef fisheries. Ecological Indicators, 2017, 73: 411-421. DOI:10.1016/j.ecolind.2016.09.045 |
| [35] |
Woodworth-Jefcoats P A, Polovina J J, Dunne J P, Blanchard J L. Ecosystem size structure response to 21st century climate projection: large fish abundance decreases in the central North Pacific and increases in the California Current. Global Change Biology, 2013, 19(3): 724-733. DOI:10.1111/gcb.12076 |
| [36] |
Shin Y J, Rochet M J, Jennings S, Field J G, Gislason H. Using size-based indicators to evaluate the ecosystem effects of fishing. ICES Journal of Marine Science, 2005, 62(3): 384-396. DOI:10.1016/j.icesjms.2005.01.004 |
| [37] |
Castro R M C, Polaz C N M. Small-sized fish: the largest and most threatened portion of the megadiverse neotropical freshwater fish fauna. Biota Neotropica, 2020, 20(1): e20180683. DOI:10.1590/1676-0611-bn-2018-0683 |
| [38] |
Li Z L, Shan X J, Jin X S, Dai F Q. Long-term variations in body length and age at maturity of the small yellow croaker (Larimichthys polyactis Bleeker, 1877) in the Bohai Sea and the Yellow Sea, China. Fisheries Research, 2011, 110(1): 67-74. DOI:10.1016/j.fishres.2011.03.013 |
| [39] |
李嘉琪, 白爱娟, 蔡亲波. 西沙群岛和涠洲岛气候变化特征及其与近岸陆地的对比. 热带地理, 2018, 38(1): 72-81. |
| [40] |
Eddy T D, Lam V W Y, Reygondeau G, Cisneros-Montemayor A M, Greer K, Palomares M L D, Bruno J F, Ota Y, Cheung W W L. Global decline in capacity of coral reefs to provide ecosystem services. One Earth, 2021, 4(9): 1278-1285. DOI:10.1016/j.oneear.2021.08.016 |
| [41] |
Emslie M J, Cheal A J, Sweatman H, Delean S. Recovery from disturbance of coral and reef fish communities on the Great Barrier Reef, Australia. Marine Ecology Progress Series, 2008, 371: 177-190. DOI:10.3354/meps07657 |
| [42] |
Barjau-González E, Rodríguez-Romero J, Galván-Magaña F, López-Martínez J. Changes in the taxonomic diversity of the reef fish community of San José Island, Gulf of California, Mexico. Biodiversity and Conservation, 2012, 21(14): 3543-3554. DOI:10.1007/s10531-012-0378-z |
| [43] |
李永振, 史赟荣, 艾红, 董丽娜, 李娜娜, 李夏, 高天翔. 南海珊瑚礁海域鱼类分类多样性大尺度分布格局. 中国水产科学, 2011, 18(3): 619-628. |
| [44] |
Shan X J, Jin X S, Yuan W. Taxonomic diversity of fish assemblages in the Changjiang Estuary and its adjacent waters. Acta Oceanologica Sinica, 2010, 29(2): 70-80. |
| [45] |
Paz-Ríos C E, Sosa-López A, Torres-Rojas Y E, del Río-Rodríguez R E. Long-term multiscale analysis of temporal variability in the fish community in Terminos Lagoon. Estuarine, Coastal and Shelf Science, 2022, 277: 108066. |