 2024, Vol. 44
2024, Vol. 44文章信息
- 吴霞, 王晓丽, 乌音嘎, 阿如汗, 虹霞
- WU Xia, WANG Xiaoli, WU Yinga, A Ruhan, HONG Xia
- 黄河内蒙古段甲烷通量变化特征及甲烷功能菌群落对通量的影响
- Variation characteristics of methane fluxes in the Inner Mongolia section of the Yellow River and the effect of methane-functional bacterial communities on fluxes
- 生态学报. 2024, 44(16): 7105-7118
- Acta Ecologica Sinica. 2024, 44(16): 7105-7118
- http://dx.doi.org/10.20103/j.stxb.202311092433
-
文章历史
- 收稿日期: 2023-11-09
- 网络出版日期: 2024-06-18






2. 内蒙古师范大学化学与环境科学学院, 呼和浩特 010022
2. College of Chemistry and Environmental Sciences, Inner Mongolia Normal University, Hohhot 010022, China
随着温室气体浓度的增加, 引发了全球变暖等一系列环境问题。CO2与CH4作为最主要的两种温室气体, 是导致地表温度上升的主要因素[1]。CH4是仅次于CO2之后的第二大温室气体, 在百年尺度内的增暖潜势是CO2的28倍, 对温室效应的贡献率约为22%[2]。自工业革命以来, 大气中的CH4浓度增加了2.7倍, 虽然大气增长率表现出明显的年际变化, 但由于各种来源, 这些浓度将持续增加[3]。了解温室气体排放源, 分析和统计温室气体排放数据, 准确估算温室气体排放量从而采取合理的应对政策和减排方法对我国非常重要。
内陆水域(湖泊、河流和水库)是大气中CH4的重要来源, 主要从缺氧的陆地土壤和水生沉积物中释放出来[4], 全球从河流中释放的甲烷约为1.5Tg CH4 a-1[5]。产甲烷菌和甲烷氧化菌是影响甲烷排放的主要微生物[6], 甲烷菌的数量、种类组成等与CH4通量紧密相关, 是参与碳循环的重要功能菌群[7]。目前关于河流CH4排放的研究局限于CH4排放通量的测算与影响因素[8―10], 将产甲烷菌和甲烷氧化菌与CH4排放通量结合起来的研究较少。
黄河内蒙古段位于黄河中上游, 流经黄土高原地区, 使黄河具有较大的碳排放潜力, 因此研究该区域温室气体通量特征及影响因素有重要意义。为了观测研究区水-气界面总通量[11], 考虑到研究区域在监测时间段内风速变化较小, 水流较缓, 水面未形成较大风浪, 从估算结果准确性来看, 宜使用静态箱法进行估算。基于静态箱-气相色谱法, 探讨了黄河内蒙古段水-气界面CH4气体通量的时空变化特征, 并对影响气体通量的主要影响因子进行了测定, 最后通过高通量测序技术从微观角度探讨了甲烷功能菌群落对黄河内蒙古段甲烷排放的影响, 可为我国河流温室气体排放量的估算提供数据支持, 有利于准确计算我国碳排放量从而采取更合理的减排方法。
1 区域概况黄河是位于中国北方地区的大河, 属世界长河之一, 中国第二长河, 全长5464km, 支流众多, 流域面积为752443km2, 上中游面积占总面积的97%。自西向东流经青海、甘肃、宁夏、内蒙古、陕西、山西、河南及山东等9个省, 最后流入渤海, 总体呈“几”字型。黄河内蒙古段西起石嘴山市与乌海市交界处的苦水沟, 东至托克托县的头道拐, 位于黄河流域的“几”字头部。黄河上中游流经黄土高原, 黄土高原土质疏松, 易蚀易散, 每逢暴雨冲刷, 则流失大量水土, 奔入黄河, 增加黄河外源碳含量, 对黄河温室气体排放造成影响, 所以黄河内蒙古段在黄河流域中具有代表性和特殊性。本研究以黄河内蒙古段为研究对象, 选取乌海(WH)、临河(LH)、乌拉特前旗(QQ)、包头(BT)、托克托县(TX)、老牛湾(LNW)六个采样点(图 1和表 1)。乌海市境内资源富集, 是内蒙古西部地区的新型工业城市;临河和乌拉特前旗地处巴彦淖尔市, 土地肥沃, 农业和畜牧业基础雄厚;包头是内蒙古最大的工业城市, 促进大量外源性碳输入河流, 影响温室气体排放;托克托县地处黄土高原, 有较好的农业发展, 且受人为影响较大;老牛湾是旅游区, 经济发展主要以农业为主。

|
| 图 1 黄河内蒙古段采样点分布图 Fig. 1 Distribution of sampling points in the Inner Mongolia section of the Yellow River WH 乌海;LH 临河;QQ 乌拉特前旗;BT 包头;TX 托克托县;LNW 老牛湾 |
| 位置名称 Location name |
经度 Longitude |
纬度 Latitude |
位置名称 Location name |
经度 Longitude |
纬度 Latitude |
|
| WH | 106°46′35.4″E | 39°39′20.78″N | BT | 109°56′48.48″E | 40°31′47.71′N | |
| LH | 107°25′7.2″E | 40°18′51″N | TX | 111°8′52.30″E | 40°14′23.89″N | |
| 108°56′55″E | 40°31′5″N | LNW | 111°26′57.12″E | 39°39′0.39″N | ||
| WH 乌海;LH 临河;QQ 乌拉特前旗;BT 包头;TX 托克托县;LNW 老牛湾 | ||||||
本研究使用静态箱-气相色谱法测定黄河内蒙古段水-气界面CH4通量, 采样箱主体由透明的有机玻璃材质构成(与水面的交界面积为0.075m2), 为保持箱内和箱外气压平衡采用导气管连接, 箱室覆盖锡箔纸, 以减少阳光注入。在采样箱顶部连接导气管, 长约1.50m, 外端用三通阀连接100.00mL注射器, 通过注射器将气体储存在采气袋中。在采样前需将采样箱放置在通风处使箱内气体混合均匀, 在采集气体时, 短时间内的气体交换浓度通常较低, 所以当采样箱在水面静置20min后开始使用注射器抽取箱内气体。采样过程中每5min抽取一次, 连续抽取5次, 编号为1—5, 于2023年3月―11月按月份在乌海(WH)、临河(LH)、乌拉特前旗(QQ)、包头(BT)、托克托县(TX)、老牛湾(LNW)采样。野外使用黄河上覆水冲洗聚乙烯瓶3—5次, 再将聚乙烯瓶置于水面以下采集表层水样, 当水完全充满聚乙烯瓶为止, 平行采集三份水样, 将其置于暗箱中, 在4h内带回实验室, 冷藏保存至4℃的冷藏箱中, 用于后续理化性质的测定。并采集表层沉积物样本, 装入聚乙烯密封袋中, 每次采样在4―5d内完成。采集的沉积物一部分低温保存主要用于理化指标的测定及向生物公司送检用于微生物分析, 其中微生物种类为产甲烷菌和甲烷氧化菌, 一部分带回实验室, 去除草木根系及石子等杂质, 经过自然风干后研磨过筛。水体的pH、溶解氧(DO)、电导率、电位等使用便携式水质监测仪(DZB-712型)实时测得, 平均风速由内蒙古气象局提供。
2.2 样品分析本研究测定的水质参数包括总氮(TN)、硝态氮(NO3--N)、氨氮(NH4+-N)、总磷(TP)、溶解有机碳(DOC)、溶解无机碳(DIC)、叶绿素a(Chl-a)。TN采用碱性过硫酸钾消解紫外分光光度法测定;NO3--N采用紫外分光光度比色法测定;NH4+-N用纳氏试剂分光光度法测定;TP采用钼酸铵分光光度法测定;DOC和DIC用总有机碳(TOC)分析仪测定;用热乙醇法测定水体中叶绿素a(Chl-a)的含量。
沉积物参数包括总氮(TN)、总碳(TC)、硝态氮(NO3--N)、氨氮(NH4+-N)、总磷(TP)。TC和TN用元素分析仪(Costech ECS4010/4024)测定; TP使用碱熔-钼锑抗分光光度法测定;NH4+-N用氯化钾溶液提取, 在碱性条件下与纳氏试剂络合生成黄色络合物进行比色测定;NO3--N用紫外分光光度比色法测定。
2.3 通量计算静态箱-气相色谱法, 是通过测定静态箱内CH4气体浓度变化率来计算水-气界面的CH4交换通量, 计算公式:
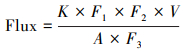
|
式中, Flux为水-气界面CH4气体通量(mg m-2 d-1), K为时间-浓度关系图里的斜率, F1为μL/L与μg/m3的单位转换系数(CH4为655.47μg/m3), F2为min和day的转换系数(1440), V为浮游箱内部的空气体积(m3), F3为μg与mg的单位转换系数(1000), A为通量箱箱底的面积(m2)。
2.4 DNA提取、PCR扩增及高通量测序沉积物样品在生物公司采用DNA试剂盒按照提供的步骤进行DNA提取, 利用1%琼脂糖凝胶电泳检测提取的基因组DNA。利用产甲烷菌的特异性引物MLfF(GGTGGTGTMGGATTCACACARTAYGCWACAGC)和MLrR(TTCATTGCRTAGTTWGGRTAGTT)进行mcrA扩增, 利用甲烷氧化菌的特异性引物A189F(GGNGACTGGGACTTCTGG)和mb661r(CCGGMGCAACGTCYTTACC)进行pmoA扩增, 反应参数如下:95℃预变性3min;95℃变性30s, 60℃退火30s, 72℃延伸45s, 共40个循环;72℃延伸10min。PCR扩增产物用2%琼脂糖凝胶电泳检测, 运用Illumina Miseq测序平台进行PCR产物的高通量测序。
2.5 数据分析甲烷数据使用Excel 2016进行处理, OriginPro进行绘图。使用IBM SPSS Statistics软件对CH4通量与影响因素进行皮尔逊相关性分析。经过生物公司高通量测序后的细菌数据使用Uparse软件平台根据相似度水平OTU划分, 进行生物信息统计分析;运用R软件对CH4通量与产甲烷菌群落和甲烷氧化菌群落属水平前4的菌群进行冗余分析(Redundancy Analysis, RDA)。
3 结果与分析 3.1 黄河内蒙古段水-气界面CH4气体通量时空变化特征本研究中黄河内蒙古段水-气界面CH4气体通量的观测时间为2023年的3―11月, 如图 2所示, 近一年的观测结果发现黄河内蒙古段水-气界面CH4气体通量的范围在-11.93―58.01mg m-2 d-1, 均值为(7.41±10.78)mg m-2 d-1, 中值为5.96mg m-2 d-1。大部分采样点的CH4排放通量集中在-2.42―27.55mg m-2 d-1之间, 少量采样点出现CH4气体通量极高值, 范围在38.24―58.01mg m-2 d-1, 整条流域总体表现为大气CH4的排放源。

|
| 图 2 黄河内蒙古段水-气界面CH4通量逐月变化 Fig. 2 Monthly variation of CH4 fluxes at the water-gas interface in the Inner Mongolia section of the Yellow River |
黄河内蒙古段水-气界面CH4通量的时间变化特征如图 3。不同采样季节黄河内蒙古段流域CH4排放通量差异显著, 春季、夏季、秋季其CH4排放通量分别为(5.92±4.54)mg m-2 d-1, (13.42±18.83)mg m-2 d-1, (1.40±3.81)mg m-2 d-1。同一采样点CH4排放通量总体表现为夏季高于春季, 秋季较低, 各采样点CH4排放通量在不同季节也表现出同样的变化规律。这可能是因为CH4的产生主要受温度的影响, 夏季温度较高会加速微生物对有机质的降解, 而春季正是大量浮游生物繁殖的时期, 对CH4的积累也有一定的促进作用。

|
| 图 3 黄河内蒙古段水-气界面CH4通量的时间变化 Fig. 3 Temporal variation of CH4 fluxes at the water-gas interface in the Inner Mongolia section of the Yellow River |
黄河内蒙古段水-气界面CH4通量的空间变化特征如图 4。黄河内蒙古段水-气界面CH4排放通量空间变化特征明显。春季CH4排放通量最高值出现在BT, 为10.02mg m-2 d-1, 最低值出现在WH, 为3.51mg m-2 d-1, 夏季CH4排放通量最高值出现在QQ, 为29.45mg m-2 d-1, 最低值出现在BT, 为1.48mg m-2 d-1, 秋季CH4排放通量最高值出现在TX, 为3.66mg m-2 d-1, 其次是LH, 最低值出现在BT, 为-0.73mg m-2 d-1。从变化幅度来看, 春季的甲烷通量变化幅度较小, 从上游到中游逐渐增大, 夏季和秋季的变化幅度较大, 可能是降雨的影响对水体造成了扰动, 对甲烷排放影响较大。

|
| 图 4 黄河内蒙古段水-气界面CH4通量的空间变化 Fig. 4 Spatial variation of CH4 fluxes at the water-gas interface in the Inner Mongolia section of the Yellow River |
黄河内蒙古段上覆水体理化性质(表 2)可以看出, 水温在整个研究期间变化范围为10.4—26.9℃, 各采样点之间水温平均值没有明显差异。pH变化范围为7.83—8.25, 水体呈碱性, 这主要受自然因素和人为因素的影响。叶绿素a是估算浮游植物生物量和水体初级生产的重要指标, 在WH和TX采样点Chl-a含量超过20μg/L, 说明这两个采样点呈现藻类较多现象, 这一现象在春季较为明显。DOC和DIC的含量范围分别为3.99—18.93mg/L, 3.33—4.13mg/L, TX采样点的DOC含量最高, 这与其含有大量浮游植物有关, 经过代谢降解产生有机碳。氮素是水体最主要的营养物质, TN的变化范围为6.86—7.65mg/L, 平均值为(7.24±0.30)mg/L, 空间变化不显著, 最低值出现在WH, 最高值出现在LH, NO3--N和NH4+-N含量最高值均出现在TX, 说明该采样点受生活污水和工业废水影响较严重。TP含量范围为0.01—0.43mg/L, 平均值为(0.22±0.19)mg/L, 最高值出现在受生活污水较严重的TX, 其次出现在受农业废水影响的LH。
| 理化参数 Physicochemical properties |
WH | LH | BT | TX | LNW | |
| Tw/℃ | 21.68 | 20.00 | 18.12 | 19.60 | 21.20 | 20.68 |
| PH | 7.83 | 8.03 | 7.95 | 8.12 | 7.99 | 8.25 |
| DO/(mg/L) | 17.68 | 8.56 | 10.94 | 13.77 | 7.22 | 14.18 |
| Chl-a/(μg/L) | 24.18 | 16.78 | 16.97 | 18.88 | 30.77 | 10.60 |
| DOC/(mg/L) | 12.75 | 5.25 | 3.99 | 11.50 | 18.93 | 7.26 |
| DIC/(mg/L) | 3.33 | 3.64 | 4.13 | 4.04 | 4.00 | 3.86 |
| TN/(mg/L) | 6.86 | 7.15 | 7.65 | 6.99 | 7.35 | 7.46 |
| NO3--N/(mg/L) | 1.28 | 1.16 | 1.34 | 0.99 | 1.70 | 1.44 |
| NH4+-N/(mg/L) | 0.25 | 0.68 | 0.52 | 0.69 | 1.70 | 0.46 |
| TP/(mg/L) | 0.33 | 0.12 | 0.38 | 0.03 | 0.43 | 0.01 |
| 数据为3个季节的平均值;Tw:水温Water temperature;DO:溶解氧Dissolved oxygen;Chl-a:叶绿素a Chlorophyll a;DOC:溶解有机碳Dissolved organic carbon;DIC:溶解无机Dissolved inorganic carbon;TN:总氮Total nitrogen;NO3--N:硝态氮Nitrate nitrogen;NH4+-N:氨氮Ammonia nitrogen;TP:总磷Total phosphorus | ||||||
黄河内蒙古段水-气界面CH4通量与水体理化性质的皮尔逊相关性分析见表 3。表中水-气界面CH4通量是在空间采样中分析计算得到的, 与之对应的水质数据是在空间采样时用水质参数仪以及实验室分析得到的。表 3结果分析表明黄河内蒙古段水-气界面CH4通量与DO呈显著负相关, 皮尔逊相关性系数为-0.898(P=0.015, P < 0.05), 与DOC和NH4+-N呈显著正相关, 皮尔逊相关性系数分别为0.927(P=0.008, P < 0.01)和0.812(P=0.05), 与Chl-a、DIC和TN呈弱相关关系, 与其他水质参数不相关。
| FCH4 | Tw | pH | DO | Chl-a | DOC | DIC | TN | NO3--N | NH4+-N | TP | |
| FCH4 | 1 | ||||||||||
| TW | -0.016 | 1 | |||||||||
| pH | 0.193 | -0.128 | 1 | ||||||||
| DO | -0.898* | 0.159 | -0.154 | 1 | |||||||
| Chl-a | 0.376 | 0.452 | -0.636 | -0.167 | 1 | ||||||
| DOC | 0.927** | 0.327 | 0.150 | -0.738 | 0.565 | 1 | |||||
| DIC | 0.376 | -0.662 | 0.437 | -0.311 | -0.102 | 0.215 | 1 | ||||
| TN | 0.340 | 0.681 | -0.222 | -0.011 | 0.851* | 0.639 | -0.063 | 1 | |||
| NO3--N | 0.152 | 0.366 | -0.069 | -0.228 | 0.428 | 0.324 | 0.152 | 0.457 | 1 | ||
| NH4+-N | 0.812* | 0.123 | 0.008 | -0.679 | 0.660 | 0.867* | 0.484 | 0.637 | 0.614 | 1 | |
| TP | 0.085 | 0.258 | -0.795 | -0.093 | 0.838* | 0.209 | -0.109 | 0.537 | 0.622 | 0.496 | 1 |
| ** 表示在0.01水平上显著相关;* 表示在0.05水平上显著相关; FCH4:甲烷通量CH4 Flux | |||||||||||
黄河内蒙古段沉积物中产甲烷菌群落丰富度和多样性(表 4)可以看出, WH、LH、QQ、BT、TX和LNW沉积物产中甲烷菌OTUs数量分别为481、149、121、483、435和48。样品测序文库覆盖率均为99.7%以上, 说明文库达到饱和, 所获得的OTUs能够代表样品的所有产甲烷菌总量。Chao指数与ACE指数表示群落丰富度, 从表 4可以看出, WH、BT采样点产甲烷菌丰富度高, 而LNW采样点产甲烷菌丰富度低。Shannon指数和Simpson指数表示群落多样性, 通过比较各采样点沉积物产甲烷菌的Shannon指数和Simpson指数发现, BT和TX的产甲烷菌多样性高, LNW的多样性低。
| 样点 Sampling site |
分类操作单元 Operational taxonomic unit |
Chao指数 Chao index |
ACE指数 ACE index |
Shannon指数 Shannon index |
Simpson指数 Simpson index |
覆盖率/% Coverage rate |
| WH | 481 | 535 | 527 | 4.64 | 0.0286 | 99.7 |
| LH | 149 | 155 | 156 | 3.99 | 0.0303 | 99.9 |
| 121 | 122 | 123 | 3.06 | 0.1040 | 99.9 | |
| BT | 483 | 518 | 527 | 4.85 | 0.0207 | 99.7 |
| TX | 435 | 449 | 445 | 5.15 | 0.0124 | 99.8 |
| LNW | 48 | 48 | 48 | 2.83 | 0.1060 | 99.9 |
沉积物中产甲烷菌群落组成在属水平的相对丰度(图 5)可以看出, 在属水平上, 样品中的主要产甲烷菌种有Methanosarcina(甲烷八叠球菌属)、Methanobacterium(甲烷杆菌属)和Archaea的未分类属unclassified_k_norank_d_Archaea。BT、LNW、TX、WH样点以Methanosarcina为主要优势菌种, 在WH样点占比最大, 在TX样点占比最小。LH样点以unclassified_k_norank_d_Archaea为主要优势菌种, QQ样点以Methanobacterium为主要优势菌种。

|
| 图 5 产甲烷菌群落组成 Fig. 5 The composition of methanogenic bacteria community |
黄河内蒙古段沉积物中甲烷氧化菌群落丰富度和多样性(表 5)可以看出, WH、LH、QQ、BT、TX和LNW沉积物中甲烷氧化菌OTUs数量分别为208、42、104、240、481和26。样品测序文库覆盖率均为99.8%以上, 说明文库达到饱和, 所获得的OTUs能够代表样品的所有产甲烷菌总量。Chao指数与ACE指数表示群落丰富度, 从表 5可以看出, BT、TX采样点甲烷氧化菌丰富度高, 而LNW采样点甲烷氧化菌丰富度低。Shannon指数和Simpson指数表示群落多样性, 通过比较各采样点沉积物甲烷氧化菌的Shannon指数和Simpson指数发现, BT和TX的甲烷氧化菌多样性高, LNW的多样性低。
| 样点 Sampling site |
分类操作单元 Operational taxonomic unit |
Chao指数 Chao index |
ACE指数 ACE index |
Shannon指数 Shannon index |
Simpson指数 Simpson index |
覆盖率/% Coverage rate |
| WH | 208 | 225 | 223 | 3.64 | 0.0694 | 99.8 |
| LH | 42 | 42 | 42 | 2.77 | 0.0793 | 100.0 |
| 104 | 107 | 110 | 3.75 | 0.0356 | 99.9 | |
| BT | 240 | 257 | 255 | 4.08 | 0.0414 | 99.8 |
| TX | 481 | 482 | 484 | 4.94 | 0.0189 | 99.9 |
| LNW | 26 | 29 | 37 | 2.19 | 0.1670 | 99.9 |
沉积物中甲烷氧化菌群落组成在属水平的相对丰度(图 6)可以看出, 在属水平上, 样品中的主要产甲烷菌种有未识别菌属norank_d_Bacteria、Methylocystis(甲基孢囊菌属)和Methylobacter(甲烷杆菌属)。LNW样点的最大优势菌种为norank_d_Bacteria, 占比达到88.48%, BT、LH、TX、WH样点的最大优势菌种为Methylocystis, LH样点中Methylobacter也是重要的菌种, BT样点中norank_f__Methylococcaceae也是重要的菌种, QQ样点的最大优势菌种为Methylobacter, 其次norank_f__Methylococcaceae也是其重要菌属。

|
| 图 6 甲烷氧化菌群落组成 Fig. 6 The composition of the methane-oxidizing bacteria community |
为研究沉积物中产甲烷菌菌群样本与环境因子的关系, 将样本与环境因子做RDA分析(图 7)。本次研究所选择的沉积物理化性质有:总碳(TC)、总氮(TN)、氨氮(NH4+-N)、硝态氮(NO3--N)和总磷(TP)。由图 7可知, 第一、第二排序轴的特征值分别为44.96%、30.99%, 结果证明TC、TP与TN整体对沉积物中产甲烷菌群落影响较大, 其中对沉积物中产甲烷菌群落影响最大的是TC(r=0.8792), 其次是TP(r=0.4064)和TN(r=0.3160), 且TC、TP与TN呈正相关关系。河流以溶解无机碳和溶解有机碳的形式转移碳, 连接陆地和海洋系统[12]。水体系统中的生物残留物和外源性输入会使得河流中含有丰富的有机质, 有机质沉淀到沉积物中会被降解[13], 当沉积物和水中有机质含量较高时, 产甲烷菌数量增加, 甲烷产量的增加导致排放通量的增加[14]。

|
| 图 7 产甲烷菌群落与环境因子的冗余分析(RDA) Fig. 7 Redundancy analysis(RDA) of methanogenic bacteria communities with environmental factors TC:总碳Total carbon;TP:总磷Total phosphorus;TN:总氮Total nitrogen;NO3--N:硝态氮Nitrate nitrogen;NH4+-N:氨氮Ammonia nitrogen |
为研究沉积物中甲烷氧化菌菌群样本与环境因子的关系, 将样本与环境因子做RDA分析(图 8)。由图 8可知, 第一、第二排序轴的特征值分别为72.38%、20.31%, 结果证明NH4+-N、TC与TP整体对沉积物中甲烷氧化菌群落影响较大, 这与鄱阳湖湿地细菌群落相关的土壤化学指标是一致的[15], 其中对沉积物中甲烷氧化菌群落影响最大的是NH4+-N(r=0.7190), 其次是TC(r=0.6821)和TP(r=0.5907), 且TC与TP呈正相关关系。

|
| 图 8 甲烷氧化菌群落与环境因子的冗余分析(RDA) Fig. 8 Redundancy analysis (RDA) of methane-oxidizing bacteria communities with environmental factors |
为探究甲烷功能菌群落组成与CH4通量的相关性, 将样本中甲烷功能菌群落分别与采样当天的CH4排放通量进行RDA分析(图 9和图 10)。由两图可以得到, 沉积物中产甲烷菌群落与甲烷氧化菌群落对CH4通量变化的解释度分别为76%、92%。样本点QQ中产甲烷菌群落组成与CH4排放通量呈显著正相关, 样本点WH、BT中产甲烷菌群落组成与CH4排放通量呈显著负相关(图 9)。Methanobacterium(甲烷杆菌属)与CH4排放通量呈显著正相关, 未分类属unclassified_k_norank_d_Archaea与CH4排放通量呈正相关。样本点LH、QQ、BT、TX甲烷氧化菌群落组成与CH4排放通量呈显著正相关, 样本点LNW和WH甲烷氧化菌群落组成与CH4排放通量呈显著负相关(图 10)。细菌未分类属norank_d_Bacteria与CH4排放通量呈显著负相关。Ⅰ型甲烷氧化菌Methylobacter与norank_d_Bacteria与CH4排放通量呈显著负相关。

|
| 图 9 产甲烷菌群落与CH4排放通量的冗余分析(RDA) Fig. 9 Redundancy analysis(RDA)of methanogenic bacteria communities with methane emission fluxes |

|
| 图 10 甲烷氧化菌群落与CH4排放通量的冗余分析(RDA) Fig. 10 Redundancy analysis (RDA) of methane-oxidizing bacteria communities with methane emission fluxes |
季节变化是自然界的发展规律, 也是影响河流温室气体排放的关键。黄河内蒙古段水-气界面CH4通量随季节变化特征明显(图 3), 夏季CH4排放通量最高, 这与大多数河流研究结果相似, 且随时间变化幅度大, 这可能是由于CH4是在严格厌氧条件下由产甲烷菌作用于产甲烷底物的结果, 环境中温度的改变会对产甲烷菌活性产生影响, 从而影响水-气界面CH4的通量[16], 且在夏季藻类大量生长繁殖, 为水体CH4的产生提供了必要的有机质, 而且在夏季等高温季节由于水体垂直化作用使得DO含量较低, 为CH4的产生提供厌氧条件[17], 导致水体产生的CH4易在底部累积, 因此温度较高的夏季是CH4产生的源。春季的CH4通量仅次于夏季, 秋季最低(图 3), 这与Sivakiruthika等人对瑞典西部的一条流域研究结果是相似的[18], 可能是由于春季降雨较少, 流量较低, 加之气温升高, 水体微生物代谢开始活跃, 同时, 春季也是亚热带区域富营养水体中浮游生物繁殖的时期, 对水体CH4的积累具有一定的促进作用[13]。
本研究基于多个采样点探究黄河内蒙古段水-气界面CH4气体通量, 各采样点水环境参数大不相同, 特别是碳、氮、磷等外源性要素的输入将影响水体内部的生物地化过程[19], 进而改变流域水体温室气体排放的空间格局。黄河内蒙古段总体表现为大气CH4的排放源, 春季甲烷通量变化幅度较小, 从上游到中游总体呈逐渐增大的趋势, 最高值出现在受工业化影响最严重的BT采样点(图 4), 由于工业废水的注入造成河流营养盐等不断增多[20], 河流呈现高氮负荷及缺氧特征, 直接刺激产甲烷菌的活性[21], 从而促进甲烷排放。而夏季CH4通量的最高值出现在QQ采样点(图 4), QQ采样点附近有部分农户居住并且岸边空地被周边居民开垦成田, 农业活动中氮肥的施用导致大量N源随地表水径流排入水体, 在营养物质充足的情况下, 导致该地区细菌群落丰度高[22], 促进CH4排放。TX采样点CH4排放通量仅次于QQ, TX采样点地处黄河中上游分界点, 流经黄土高原, 黄土本身结构松散, 很容易受侵蚀和崩塌, 助长了水土流失, 大量泥沙涌入黄河, 使得TX采样点受大量外源性碳的输入, 从而含有丰富的有机质, 有机质沉淀到沉积物中被降解[13]。当沉积物和水中有机质含量较高时, 产甲烷菌数量增加, 甲烷产量的增加导致排放通量的增加[14]。所以, 六个采样点受污染状况不同和所处地形的差异从而导致了不同的排放趋势。
4.2 影响因素分析黄河内蒙古段水-气界面CH4气体通量呈现较大的空间差异性, 与复杂的环境因子密切相关。本研究中, CH4通量与DO呈显著负相关(r=-0.898, P=0.015)(表 3), 这与Furlanetto等人对巴西Rio Grande流域的研究结果是相似的[23]。DO是影响CH4产生和氧化的重要因素[24], CH4是厌氧条件下有机碳矿化的主要产物[25], 通过孔隙水进入沉积物和水界面, 然后进入水体向上扩散, 甲烷扩散的过程中被氧化为CO2, 在水扩散期间, 甲烷进一步氧化, 只有少量的甲烷排放到大气中, 所以缺氧条件有利于甲烷的生产和排放。溶解性有机碳(DOC) 丰富程度对水体中CO2和CH4的产生与排放也具有重要影响[26], CH4通量与DOC呈显著正相关关系(r=0.927, P=0.008)(表 3), TX采样点淹没大量植被和沉积物, 水体中易分解的DOC浓度高, 致使春季和夏季的CH4通量较高[27], 这与其他研究结果相似, 而DOC影响水体CO2和CH4排放通量的主要作用机制是DOC为水体中CO2和CH4产生的直接碳源, 其含量高低直接关系到相关微生物活性。人为活动对河流CH4通量有一定影响, 直接表现为河流水体中营养物质的输入, 特别是与碳、氮、磷等生源要素关系密切。在本研究中, CH4通量与NH4+-N呈显著正相关关系(r=0.812, P=0.015)(表 3), 高的NH4+-N含量主要分布在BT和TX采样点, TX采样点作为景观用水, 受污来源较多, 而BT采样点附近有较多大型工厂, 不但受生活污水的影响, 还有工业废水的注入, 使其NH4+-N含量较高, 营养盐影响河流CH4产生与排放的作用机制为: (1)为产甲烷菌产CH4提供底物;(2)营养物质分解会导致DO含量降低, 造成厌氧条件[28];(3)水中的NH4+会对甲烷氧化菌产生抑制作用[29], 所以营养盐会促进CH4排放。本研究中, 黄河内蒙古段流域水体磷含量较低(表 2), 对CH4排放产生影响较小, DO、DOC和NH4+-N是CH4排放的主要影响因子。
4.3 甲烷功能菌群落对甲烷排放的影响产甲烷菌和甲烷氧化菌是介导CH4循环的重要功能菌群, 产甲烷菌是一种广泛分布于厌氧环境的古菌, 在有机物厌氧降解的最后一步将其转化为CH4[30]。产甲烷菌按照底物利用类型差异可分为氢营养型产甲烷菌、乙酸营养型产甲烷菌和甲基营养型产甲烷菌, 不同区域对底物利用的类型不同, 导致产甲烷途径有很大差别[31]。本研究发现, Methanosarcina(甲烷八叠球菌属)、Methanobacterium(甲烷杆菌属)为研究区的优势菌种(图 5), Methanosarcina(甲烷八叠球菌属)为乙酸营养型产甲烷菌, 通过裂解乙酸, 将乙酸的羧基氧化为CO2, 甲基还原为甲烷。Methanobacterium(甲烷杆菌属)为氢营养型产甲烷菌[32], 主要利用H2和甲酸作为电子供体还原CO2产甲烷[33]。有研究表明, CH4的产生和排放与产甲烷菌群落紧密相关[34]。Adam等人对捷克Morava流域的甲烷动态研究发现, 堰坝上游沉积物的甲烷产量和氧化潜力均高于堰坝的下游, 这是因为氢营养型甲烷菌对总甲烷沉积物的贡献为36.7%—89.4%[35]。Franziska等人[36]发现大部分甲烷产生于河流沉积物中的缺氧区, 这是厌氧产甲烷古菌还原二氧化碳、醋酸盐或甲醇的结果。Yu等人[37]也研究发现甲烷通量取决于产甲烷菌或产甲烷营养菌在环境中是否更活跃。Chen等人[38]对珠江口甲烷代谢研究发现沉积物中的产甲烷菌是甲烷的重要来源, 而且微生物的产甲烷作用主要发生在缺氧环境中。
先前大量研究表明, 甲烷氧化菌是淡水系统中温室气体甲烷主要的微生物汇[39], 对大气中甲烷浓度保持平衡起至关重要的作用。好氧甲烷氧化菌主要存在于甲烷与氧气共存的微小界面空间, 包括土壤-空气、水-空气界面、植物根际以及植物内部[40]。本研究的结果表明, LNW样点的最大优势菌种为norank_d_Bacteria, 其他样点的最大优势菌种为Methylocystis(甲基孢囊菌属)和Methylobacter(甲烷杆菌属)(图 6), 其中Methylocystis属于Type Ⅱ型甲烷氧化菌, 归属于Methylocystaceae科, Methylobacter属于Type Ⅰ型甲烷氧化菌, 在王晓琳[41]的研究中, Type Ⅱ型甲烷氧化菌Methylocystis以及Type Ⅰ型甲烷氧化菌Methylobacter为研究区的优势菌属, 这与本研究的结果较为一致。但总体按占比来看, 黄河内蒙古段沉积物中Type Ⅱ型甲烷氧化菌为最大优势菌属。而有大量研究表明, 淡水沉积物中甲烷氧化菌以Type Ⅰ型的Methylomonas、Methylobacter和Methylosarcina等属为主[42]。在张洪勋[43]的研究中, 我国青藏高原若尔盖永冻土湿地中好氧甲烷氧化菌仅有Methylobacter属和Methylocystis两个属, 且以Type Ⅰ型甲烷氧化菌为优势菌群。刘洋[44]研究发现北运河沉积物中甲烷氧化菌以Type Ⅰ型为主, 这与本实验研究结果有所差别, 需要做进一步研究。
5 结论(1) 研究期间, 黄河内蒙古段总体表现为大气CH4的排放源, 春季、夏季、秋季CH4排放通量分别为(5.92±4.54)mg m-2 d-1, (13.42±18.83)mg m-2 d-1, (1.40±3.81)mg m-2 d-1, 存在明显的季节变化, 且不同采样点排放通量存在显著空间差异性。
(2) 黄河内蒙古段水-气界面CH4通量与DO呈显著负相关, 皮尔逊相关性系数为-0.898(P=0.015, P < 0.05), 与DOC和NH4+-N呈显著正相关, 皮尔逊相关性系数分别为0.927(P=0.008, P < 0.01)和0.812(P=0.05)。
(3) 不同样本中甲烷功能菌群落多样性存在显著差异, 在产甲烷菌群落中, Methanobacterium(甲烷杆菌属)和未分类古菌unclassified_k_norank_d_Archaea对CH4排放影响最大, 这类细菌会促进甲烷排放。在甲烷氧化菌群落中, 未识别的甲烷氧化菌norank_d_Bacteria对CH4氧化影响最大, 这类细菌会促进甲烷氧化, 从而减少水-气界面甲烷排放。
| [1] |
曲建升, 孙成权, 张志强, 高峰. 全球变化科学中的碳循环研究进展与趋向. 地球科学进展, 2003, 18(6): 980-987. |
| [2] |
Xue H, Yu R H, Zhang Z Z, Qi Z, Lu X X, Liu T X, Gao R Z. Greenhouse gas emissions from the water-air interface of a grassland river: a case study of the Xilin River. Scientific Reports, 2021, 11: 2659. DOI:10.1038/s41598-021-81658-x |
| [3] |
Li Y H, Zhan L Y, Chen L Q, Zhang J X, Wu M, Liu J. Spatial and temporal patterns of methane and its influencing factors in the Jiulong River Estuary, southeastern China. Marine Chemistry, 2021, 228: 103909. DOI:10.1016/j.marchem.2020.103909 |
| [4] |
Shi W Q, Chen Q W, Zhang J Y, Lu J, Chen Y C, Pang B H, Yu J H, Van Dam B R. Spatial patterns of diffusive methane emissions across sediment deposited riparian zones inhydropower reservoirs. Journal of Geophysical Research: Biogeosciences, 2021, 126(3): e2020JG005945. DOI:10.1029/2020JG005945 |
| [5] |
Bastviken D, Tranvik L J, Downing J A, Crill P M, Enrich-Prast A. Freshwater methane emissions offset the continental carbon sink. Science, 2011, 331(6013): 50. DOI:10.1126/science.1196808 |
| [6] |
张曦. 三峡库区万州段沉积物细菌群落对CH4排放的影响[D]. 重庆: 重庆交通大学, 2020.
|
| [7] |
刘垚垚, 葛继稳, 斯南雍茜, 蒋浩, 袁梦媛, 冯亮, 张志麒. 神农架大九湖亚高山泥炭湿地甲烷通量变化特征及其与产甲烷菌群落组成的关系. 长江流域资源与环境, 2021, 30(6): 1418-1427. |
| [8] |
陈玉鹏, 毛旭锋, 魏晓燕, 苏晓虾, 张帅, 刘小君. 连续筑坝河流水气界面温室气体排放通量及其影响因素——以青海省湟水支流火烧沟为例. 水生态学杂志, 2020, 41(3): 17-23. |
| [9] |
常思琦, 王东启, 俞琳, 任名成, 胡蓓蓓, 陈振楼. 上海城市河流温室气体排放特征及其影响因素. 环境科学研究, 2015, 28(9): 1375-1381. |
| [10] |
吴瑶洁, 李海英, 陈文重, 卢佳, 赵荧, 刘定富. 夏季温榆河温室气体释放特征与影响因素研究. 环境科学与技术, 2016, 39(5): 8-16. |
| [11] |
李航, 王晓锋, 袁兴中, 王继龙, 李贤祥, 周婷. 淡水生态系统温室气体通量监测方法综述. 湖泊科学, 2023, 35(4): 1153-1175. |
| [12] |
Gu C, Waldron S, Bass A M. Carbon dioxide, methane, and dissolved carbon dynamics in an urbanized river system. Hydrological Processes, 2021, 35(9): e14360. DOI:10.1002/hyp.14360 |
| [13] |
Yang H, Andersen T, Dörsch P, Tominaga K, Thrane J E, Hessen D O. Greenhouse gas metabolism in Nordic boreal lakes. Biogeochemistry, 2015, 126(1): 211-225. |
| [14] |
Martinez-Cruz K, Gonzalez-Valencia R, Sepulveda-Jauregui A, Plascencia-Hernandez F, Belmonte-Izquierdo Y, Thalasso F. Methane emission from aquatic ecosystems of Mexico City. Aquatic Sciences, 2017, 79(1): 159-169. DOI:10.1007/s00027-016-0487-y |
| [15] |
王鹏, 陈波, 张华. 基于高通量测序的鄱阳湖典型湿地土壤细菌群落特征分析. 生态学报, 2017, 37(5): 1650-1658. DOI:10.5846/stxb201510052000 |
| [16] |
宋红丽, 刘兴土, 文波龙. 黄河三角洲养殖塘水-气界面CO2、CH4和N2O通量特征. 生态环境学报, 2017, 26(9): 1554-1561. |
| [17] |
杨丽娜, 李正炎, 张学庆. 大辽河近入海河段水体溶解氧分布特征及低氧成因的初步分析. 环境科学, 2011, 32(1): 51-57. |
| [18] |
Natchimuthu S, Sundgren I, Gålfalk M, Klemedtsson L, Crill P, Danielsson Å, Bastviken D. Spatio-temporal variability of lake CH4 fluxes and its influence on annual whole lake emission estimates. Limnology and Oceanography, 2016, 61(S1): S13-S26. |
| [19] |
Marescaux A, Thieu V, Garnier J. Carbon dioxide, methane and nitrous oxide emissions from the human-impacted Seine watershed in France. The Science of the Total Environment, 2018, 643: 247-259. DOI:10.1016/j.scitotenv.2018.06.151 |
| [20] |
韩洋, 郑有飞, 吴荣军, 尹继福, 孙霞. 南京河流夏季水-气界面N2O排放通量. 环境科学, 2014, 35(1): 348-355. |
| [21] |
Liu L L, Greaver T L. A review of nitrogen enrichment effects on three biogenic GHGs: the CO2 sink may be largely offset by stimulated N2O and CH4 emission. Ecology Letters, 2009, 12(10): 1103-1117. DOI:10.1111/j.1461-0248.2009.01351.x |
| [22] |
王晓丽, 其勒格尔. 黄河内蒙古段表层沉积物细菌多样性及群落结构类型. 生态学报, 2020, 40(2): 578-589. DOI:10.5846/stxb201806261405 |
| [23] |
Furlanetto L M, Marinho C C, Palma-Silva C, Albertoni E F, Figueiredo-Barros M P, de Assis Esteves F. Methane levels in shallow subtropical lake sediments: dependence on the trophic status of the lake and allochthonous input. Limnologica, 2012, 42(2): 151-155. DOI:10.1016/j.limno.2011.09.009 |
| [24] |
Schrier-Uijl A P, Veraart A J, Leffelaar P A, Berendse F, Veenendaal E M. Release of CO2 and CH4 from lakes and drainage ditches in temperate wetlands. Biogeochemistry, 2011, 102(1): 265-279. |
| [25] |
Li Y, Shang J H, Zhang C, Zhang W L, Niu L H, Wang L F, Zhang H J. The role of freshwater eutrophication in greenhouse gas emissions: a review. The Science of the Total Environment, 2021, 768: 144582. DOI:10.1016/j.scitotenv.2020.144582 |
| [26] |
Jacinthe P A, Filippelli G M, Tedesco L P, Raftis R. Carbon storage and greenhouse gases emission from a fluvial reservoir in an agricultural landscape. CATENA, 2012, 94: 53-63. DOI:10.1016/j.catena.2011.03.012 |
| [27] |
Wang D Q, Chen Z L, Sun W W, Hu B B, Xu S Y. Methane and nitrous oxide concentration and emission flux of Yangtze Delta Plain river net. Science in China Series B: Chemistry, 2009, 52(5): 652-661. |
| [28] |
Hu B B, Wang D Q, Zhou J, Meng W Q, Li C W, Sun Z B, Guo X, Wang Z L. Greenhouse gases emission from the sewage draining rivers. The Science of the Total Environment, 2018, 612: 1454-1462. |
| [29] |
Conrad R, Rothfuss F. Methane oxidation in the soil surface layer of a flooded rice field and the effect of ammonium. Biology and Fertility of Soils, 1991, 12(1): 28-32. |
| [30] |
彭兴意, 秦宇, 舒钰清, 李银波, 张曦. 三峡库区夏季万州段底泥甲烷功能菌群落对甲烷排放的影响. 环境工程学报, 2022, 16(3): 1028-1038. |
| [31] |
许晓晴, 陈烨, 甄毓, 米铁柱, 李晶, 刘昌岭. 渤海沉积物中产甲烷途径及产甲烷菌群落特征. 海洋地质与第四纪地质, 2022, 42(3): 50-61. |
| [32] |
任秘坊, 李凌燕, 陈琳, 邢婷婷, 刘勇勤, 东秀珠. 青藏高原三个盐碱湖的产甲烷菌群和产甲烷代谢途径分析. 微生物学报, 2020, 60(1): 161-171. |
| [33] |
王洁, 袁俊吉, 刘德燕, 项剑, 丁维新, 蒋先军. 滨海湿地甲烷产生途径和产甲烷菌研究进展. 应用生态学报, 2016, 27(3): 993-1001. |
| [34] |
顾航, 肖凡书, 贺志理, 颜庆云. 湿地微生物介导的甲烷排放机制. 微生物学报, 2018, 58(4): 618-632. |
| [35] |
Bednarˇík A, Blaser M, Matouš A, Hekera P, Rulík M. Effect of weir impoundments on methane dynamics in a river. The Science of the Total Environment, 2017, 584/585: 164-174. |
| [36] |
Broman E, Sun X L, Stranne C, Salgado M G, Bonaglia S, Geibel M, Jakobsson M, Norkko A, Humborg C, Nascimento F J A. Low abundance of methanotrophs in sediments of shallow boreal coastal zones with high water methane concentrations. Frontiers in Microbiology, 2020, 11: 1536. |
| [37] |
Yu X L, Yang X Q, Wu Y J, Peng Y S, Yang T, Xiao F S, Zhong Q P, Xu K, Shu L F, He Q, Tian Y, Yan Q Y, Wang C, Wu B, He Z L. Sonneratia apetala introduction alters methane cycling microbial communities and increases methane emissions in mangrove ecosystems. Soil Biology and Biochemistry, 2020, 144: 107775. |
| [38] |
Chen S Z, Wang P, Liu H D, Xie W, Wan X S, Kao S J, Phelps T J, Zhang C L. Population dynamics of methanogens and methanotrophs along the salinity gradient in Pearl River Estuary: implications for methane metabolism. Applied Microbiology and Biotechnology, 2020, 104(3): 1331-1346. |
| [39] |
沈李东. 湿地亚硝酸盐型厌氧氨氧化和厌氧甲烷氧化微生物生态学研究[D]. 杭州: 浙江大学, 2014.
|
| [40] |
贠娟莉, 王艳芬, 张洪勋. 好氧甲烷氧化菌生态学研究进展. 生态学报, 2013, 33(21): 6774-6785. DOI:10.5846/stxb201207171013 |
| [41] |
王晓琳, 曹爱新, 周传斌, 赵恺凝, 赵国柱. 垃圾填埋场甲烷氧化菌及甲烷减排的研究进展. 生物技术通报, 2016, 32(5): 16-25. |
| [42] |
Yun J L, Wang Y F, Zhang H X. Ecology of aerobic methane oxidizing bacteria (methanotrophs). Acta Ecologica Sinica, 2013, 33(21): 6774-6785. |
| [43] |
Yun J L, Zhuang G Q, Ma A Z, Guo H G, Wang Y F, Zhang H X. Community structure, abundance, and activity of methanotrophs in the zoige wetland of the Tibetan Plateau. Microbial Ecology, 2012, 63(4): 835-843. |
| [44] |
刘洋, 陈永娟, 王晓燕, 许康利. 城市河流沉积物中氮素与好氧甲烷氧化菌群落特征响应关系. 湖泊科学, 2018, 30(5): 1271-1283. |