 2024, Vol. 44
2024, Vol. 44文章信息
- 陈彦君, 张瑜函, 傅博, 潘丽波, 邢世平, 肖能文, 关潇
- CHEN Yanjun, ZHANG Yuhan, FU Bo, PAN Libo, XING Shiping, XIAO Nengwen, GUAN Xiao
- 转Cry1Ah基因抗虫玉米HGK60对田间节肢动物多样性的影响
- Impact of transgenic insect-resistant maize HGK60 with Cry1Ah gene on biodiversity of arthropods in the fields
- 生态学报. 2024, 44(16): 7360-7370
- Acta Ecologica Sinica. 2024, 44(16): 7360-7370
- http://dx.doi.org/10.20103/j.stxb.202309262085
-
文章历史
- 收稿日期: 2023-09-26
- 网络出版日期: 2024-06-25






2. 生态环境部机关服务中心, 北京 100006;
3. 桓台县公园城市管理服务中心, 淄博 256400
2. Government Service Center, Ministry of Ecology and Environment, Beijing 100006, China;
3. Huantai County Park City Management Service Center, Zibo 256400, China
最新报告显示, 全球转基因作物的种植面积已从1996年的170万hm2增加到2022年的2.022亿hm2, 增长了约112倍, 极大地提高了全球农作物生产力[1]。转基因作物在带来显著经济效益的同时, 其可能带来的环境安全性问题也越来越多的受到关注, 种植后是否会对生态环境造成影响还需要进行长期、全方位的安全性测定。迄今为止, 已经报道了一些转基因作物商业种植引发的相关农业的问题[2], 例如, 靶标昆虫抗性的产生[3—5]、“超级杂草”[6]、对非靶标生物的影响[7]等。
节肢动物作为生态系统的重要组成部分, 在食物链构建、生物防治、授粉和有机物分解等方面都发挥了重要作用, 是维持农田生态系统功能的重要参与者[8—10], 对其多样性的评估是转基因作物环境安全评价的重要环节之一。Cry1Ah蛋白是一种来自Bt亚种的杀虫基因, 对鳞翅目昆虫表现出极大的毒性, 杀虫效率高于Cry1Ab、Cry1Ac等, 但是对家蚕等经济昆虫的毒性却相对较低[11]。本研究以转Cry1Ah基因抗虫玉米HGK60为研究对象, 研究了其种植对田间节肢动物的影响, 以期用更加详实的数据为转Cry1Ah基因抗虫玉米HGK60的环境安全评价提供数据支撑, 为生物安全管理的健康发展贡献力量。
1 试验区概况试验区位于河北省廊坊市中国农业科学研究院国际农业高新技术产业园(116°36′34″E, 39°36′10″N), 属暖温带大陆性季风气候, 年均气温约11.9℃, 年平均降水量为554.9 mm, 降水多集中于夏季, 年平均日照时数约2660 h。该基地为农业农村部已认定的农业转基因生物试验基地, 符合《农业转基因生物安全管理通用要求试验基地》(农业部2406号公告-3—2016)相关规定。
2 材料与方法 2.1 供试材料与实验设计选用玉米品种为中国农业科学院生物技术研究所(北京)提供的转Cry1Ah基因抗虫玉米HGK60(简称“HGK60”)及其对照常规玉米郑58(亲本, 简称“郑58”)。以HGK60为试验组, 以郑58为对照组, 采取随机区组设计, 两个处理均设置6个重复(10 m×10 m), 不同处理间设置1 m隔离带, 采用一穴一粒、一穴两粒的循环播种方式进行玉米播种, 保持种植行距60 cm, 株距25 cm, 田间管理同当地玉米常规种植, 种植时间为2019年、2020年和2021年5月初。
2.2 样品的采集与处理采用直接观测法、陷阱调查法和扫网法[12], 连续3年在玉米生长期的苗期、喇叭口期、抽穗期和成熟期, 按照五点采样法进行田间节肢动物样本的采集, 每个生育期分别进行3次。
直接观察法:每点选择15株玉米, 调查植株上节肢动物种类和数量;陷阱调查法:每点埋设5个塑料杯(直径7.5 cm, 间隔0.5 m), 其中放置1/3防冻液(5%), 保持杯口与地面平齐, 24 h后调查杯中节肢动物的种类和数量并记录。扫网法采样时间为16:00—19:00[12], 每个点随机抽检10株玉米, 采用直径30 cm的防虫网, 对玉米植株茎叶自上而下取样10次。将所有标本放入含70%酒精的瓶中, 带回实验室, 进行物种鉴定与统计[13]。按其在食物链中的位置(营养水平)大致分为四个类群:植食性、捕食性、寄生性和中性类群(包括腐生类和对植物无危害、不食其他昆虫的类群)。蛛形纲(Arachnida)的物种被划分为捕食类群[14]。
2.3 数据处理与分析统计分析玉米各生育期田间节肢动物的群落结构与组成, 计算各生态学指数。
(1) Margalef指数: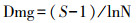
(2) Shannon指数: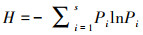
(3) 优势度指数: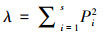
(4) 均匀度指数:
式中:S为物种数;Pi=Ni/N, 其中Ni为第i种物种的个体数, N为总个体数。
群落相似性系数:C′=2W/(a+b), 其中W是HGK60和郑58共有物种数量, a是HGK60的物种总数, b是郑58的物种总数。0.75≤ C′<1.0表示非常相似, 0.5≤C′<0.75表示中等相似, 0.25≤C′<0.5表示中等差异, 0<C′<0.25表示差异较大[15]。
用Nn/Np、Nd/Np、Sn/Sp、Sd/Sp值反应节肢动物群落中天敌类群和中性类群对植食性类群的调控作用。其中, Nn、Np、Nd分别表示天敌类群、植食性类群、中性类群个体数, Sn、Sp、Sd分别代表天敌类群、植食性类群、中性类群物种数[16]。天敌类群包括捕食性、寄生性天敌。
利用Execl 2010软件进行数据整理, 利用SPSS 16.0软件分析两处理差异(t-test)。
3 结果与分析 3.1 田间节肢动物群落组成调查2019—2021年HGK60与郑58玉米田间节肢动物, 共记录11目40科124种48746头(表 1)。比较同一生育期HGK60与郑58玉米田间节肢动物数量, 螟蛾科在2021年抽穗期、完熟期表现为HGK60与郑58差异显著(P<0.05);盲蝽科、姬蝽科、草蛉科仅在2019年抽穗期表现为HGK60显著高于郑58(P<0.05);蝗科仅在2021年苗期表现为二者差异显著(P<0.05);实蝇科仅在2021年抽穗期表现为二者差异显著(P<0.05);食蚜蝇科、园蛛科仅在2021年完熟期表现为二者差异显著(P<0.05)。其他物种及生育期均表现为HGK60与郑58差异不显著(P>0.05)(图 1)。
| 目Orders | 科Families | 种Species |
| 鳞翅目Lepidoptera | 螟蛾科Pyralidae | 玉米螟Pyrausta nubilalis、二化螟Chilo suppressalis、三化螟Scirpophaga incertulas、褐边螟Catagela adjurella、桃蛀螟Conogethes punctiferalis、葡萄卷叶野螟Pyralidae、稻纵卷叶螟Cnaphalocrocis medinalis、二点螟Chilo infuscatellus |
| 弄蝶科Hesperiidae | 小黄斑弄蝶Ampittia dioscorides | |
| 粉蝶科Pieridae | 菜粉蝶Pieris rapae、山楂绢粉蝶Aporia crataegi | |
| 羽蛾科Pterophoridae | Pterophorus monodactylus | |
| 半翅目Hemiptera | 盲蝽科Miridae | 绿盲蝽Apolygus lucorum、华氏卵盲蝽Cyrtorrhinus chinesis、小赤须盲蝽Tenuis、苜蓿盲蝽Adelphocoris lineolatus、三点盲蝽Adelphocoris fsdciaticollis |
| 姬蝽科Nabidae | 华姬蝽Nabis sinoferus、Gorpis brevilineatus | |
| 长蝽科Lygaeidae | 横带红长蝽Lygaeus equestris、红脊长蝽Tropidothorax elegans Distant、大眼长蝽Geocoris pallidipennis | |
| 花蝽科Anthocoridae | Orius similis、东亚小花蝽Orius sauteri、微小花蝽Orius minutus | |
| 猎蝽科Reduviidae | Coranus lativentris、Haematoloecha nigrorufa、Oncocephalus confusus | |
| 缘蝽科Coreidae | 点蜂缘蝽Riptortus pedestris、南瓜缘蝽Cores marginatus、茶色赭缘蝽Ochrochira camelina | |
| 同翅目Homoptera | 飞虱科Delphacidae | 白背飞虱Sogatella furcifera、长绿飞虱Saccharosydne procerus、白条飞虱Terthron albovittata、褐飞虱Nilaparvata lugens、拟褐飞虱Nilaparavata bakeri、伪褐飞虱Nilaparvata muiri、灰飞虱Laodelphax striatellus |
| 叶蝉科Cicadellidae | 大青叶蝉Cicadella viridis、黑尾叶蝉Nephotettix bipunctatus、电光叶蝉Inazuma dorsalis、白翅叶蝉Thaia rubiginosa、一点叶蝉Erythroneura sudra、四点叶蝉Macrosteles quadrimaculata、二点叶蝉Cotton leafhopper、小绿叶蝉Jacobiasca formosana、大白叶蝉Tettigoniella spectra | |
| 蚜科Aphidoidea | 玉米蚜Rhopalosiphum maidis、麦长管蚜Sitobion avenae、桃蚜Myzus persicae、油菜蚜Lipaphis erysimi pseudobrassicae | |
| 木虱科Psyllidae | 中国梨木虱Psyllia chinensis、桑木虱Anomoneura mori Schwarz | |
| 直翅目Orthoptera | 蟋蟀科Gryllidae | 北京油葫芦Teleogryllus emma、大扁头蟋Loxoblemmus doenitzi Stein |
| 蝗科Acrididae | 亚洲小车蝗Oedaleus decorus asiaticus、日本黄脊蝗Patanga japonica、中华蚱蜢Acrida cinerea、东亚飞蝗Locusta migratoria manilensis | |
| 蚱科Tetrigoidea | 花胫绿纹蝗Aiolopus tamulus、菱蝗Tetrigid、平背棘菱蝗Eucriotettix oculatus | |
| 脉翅目Neuroptera | 草蛉科Chrysopidae | 中华通草蛉Chrysoperla sinica、大草蛉Chrysopa pallens、丽草蛉Chrysopa formosa Brauer、多斑草蛉Chrysopa intima |
| 鞘翅目Coleoptera | 步甲科Carabidae | 黄斑青步甲Chlaenius micans Fabricius、中华星步甲Calosoma Chinese、后斑青步甲Chlaenius Posticalis、一棘蝼步甲Scarites terricola、毛婪步甲Harpalus griseus |
| 瓢虫科Staphylinidae Coccinellidae | 龟纹瓢虫Propylaea japonica、异色瓢虫Harmonia axyridis、二星瓢虫Adalia bipunctata、十二星瓢虫Henosepilachna pusillanima、Coccinella septempunctata、Adonia variegata | |
| 隐翅虫科Staphylinidae | Paederus fuscipes | |
| 缨翅目Thysanoptera | 蓟马总Thripoidea | 禾蓟马Frankiniella tenuicornis、西花蓟马Franjliniella occidentalis |
| 双翅目Diptera | 食蚜蝇Syrphidae | Dideopsis aegrotus、Eristalis tenax、Sphaerophoria sp.、Bacch maculata |
| 实蝇科Tephritidae | 瓜实蝇Bactrocera cucurbitae、柑橘大实蝇Bactrocera Minax | |
| 寄蝇科Tachinidae | Exorista civilis、Lydella grisescens、Cuphocera varia | |
| 蝇科Muscidae | Musca domestica、厩腐蝇Muscina stabulans、厩螫蝇Stomoxys calcitrans、夏厕蝇Fannia canicularis | |
| 果蝇科Drosophilidae | 黑腹果蝇Drosophila melanogaster、拟暗果蝇D.pseudoobscura | |
| 麻蝇科Sarcophagidae | 肥须亚麻蝇Parasarcophaga crassipalpis、拟对岛亚麻蝇Parasarcophaga (Kanoisca) kanoi | |
| 蜻蜓目Odonata | 蟌科Coenagrionidae | Ischnura asiatica、Ischnura elegans |
| 膜翅目Hymenoptera | 姬蜂科Ichneumonidae | Charops bicolor、Vulgichneumon leucaniae、Campoletis chlorideae、Coccygomimus disparis、Diplozon laetatorius、Butenyacra picta |
| 赤眼科Trichogrammatidae | Trichogramma ostriniae | |
| 跳小蜂科Encyrtidae | Aphidencyrtus aphidivorus | |
| 青蜂科Chrysididae | Chrysis shanghaiensis | |
| 泥蜂科Sphecidae | Sceliphron deforme、Sphex umbrosus、 | |
| 金小蜂科Pteromalidae | Dibrachys xavus | |
| 缨小蜂科Mymaridae | Lymaenon sp. | |
| 茧蜂科Braconidae | Macrocentrus cingulum | |
| 蜘蛛目Araneae | 园蛛科Araneidae | 大腹园蛛Araneus ventricosus、Araneus ejusmodi、Araneus yuanminensis、银背艾蛛Cyclosa argenteoalba、八瘤艾蛛Cyclosa octotuberculata、黄褐新园蛛Neoscone doenitzi、茶色新圆蛛Neoscona theisi |
| 蟹蛛科Thomisidae | Misumenops tricuspidatus、Xysticus ephippiafus | |
| 跳蛛科Salticidae | Marpissa elongata、Myrmarachne formicaria、Sitticus sinensis |

|
| 图 1 田间节肢动物相对丰度聚类热图 Fig. 1 The cluster heatmap of relative abundance of arthropods in the fields Ⅰ, 苗期Seedling stage; Ⅱ, 喇叭口期Bell stage; Ⅲ, 抽穗期Heading stage; Ⅳ, 完熟期Full ripe sta |
对不同营养类群田间节肢动物的物种数和个体数进行统计分析(图 2)。植食性、捕食性和寄生性类群分析结果显示, 连续3年各生育期HGK60与郑58在物种数和个体数上均没有显著差异(P>0.05)。对于中性类群, HGK60与郑58田间节肢动物的物种数在各种生育期均无显著差异(P>0.05), 其个体数仅在2019年的抽穗期表现为HGK60显著高于郑58(P<0.05)。总体上, HGK60种植对田间节肢动物各营养类群的种类数和个体数均无显著影响(P>0.05)。进一步研究不同营养类群节肢动物物种数与个体数在田间节肢动物中所占比例, 植食性类群的物种数在两种玉米田间节肢动物群落中所占比例最大, 寄生性类群、中性类群的物种数相对较少。各类群节肢动物个体数亦表现为植食性类群最多, 捕食性次之, 寄生性和中性类群相对较少。同一年份同一生育期, HGK60与郑58田间各类群节肢动物物种数和个体数所占比例基本保持一致(图 3)。

|
| 图 2 田间各营养类群节肢动物物种数和个体数 Fig. 2 Species number and individual number of four groups of arthropods in the fields |

|
| 图 3 田间节肢动物物种数和个体数的百分比 Fig. 3 The percentage of the species number and individual number of arthropods |
选取丰富度指数、Shannon指数、优势度指数和均匀度指数对HGK60与郑58田间节肢动物生态指标进行分析, 结果显示2019—2021连续3年的各生育期HGK60和郑58在各指数均未表现出显著差异(P>0.05), 认为HGK60的种植对田间节肢动物各生态指标无显著影响(图 4)。

|
| 图 4 田间节肢动物群落各生态指标 Fig. 4 The biodiversity indexes of arthropods in the fields |
对节肢动物群落相似性研究结果显示(表 2), 3年各生育期HGK60与郑58田间节肢动物群落相似性系数均高于0.7, 除2019年苗期、2020年喇叭口期以及2021年的苗期、抽穗期, 二者在均达到高度相似水平;HGK60与郑58植食性类群相似性亦较高, 二者在3年的各生育期均接近或达到高度相似水平, 其中二者2020年抽穗期的相似性最高, 相似性系数达到0.8;捕食性类群的相似性在2019及2020年各生育期均为高度相似水平, 2021年相似性略低于前两年, 但均达到中度相似并趋向高度相似水平;对寄生性类群的分析结果显示, 二者在3年各生育期的相似性系数均大于0.6, 达到中度相似水平, 其中2019年喇叭口期、完熟期以及2020年完熟期为高度相似水平;2019、2020年完熟期HGK60与郑58田间节肢动物的中性类群为高度相似, 其他生育期均为中度相似。
| 年份 Year |
生育期 Growth period |
节肢动物 Arthropod community |
植食性类群 Herbivorous group |
捕食性类群 Predatory group |
寄生性类群 Parasitic group |
中性类群 Neutral group |
| 2019 | 苗期 | 0.73 | 0.72 | 0.76 | 0.67 | 0.67 |
| 喇叭口期 | 0.75 | 0.73 | 0.78 | 0.83 | 0.60 | |
| 抽穗期 | 0.75 | 0.77 | 0.76 | 0.67 | 0.71 | |
| 完熟期 | 0.78 | 0.79 | 0.76 | 0.76 | 0.80 | |
| 2020 | 苗期 | 0.75 | 0.77 | 0.78 | 0.62 | 0.60 |
| 喇叭口期 | 0.74 | 0.73 | 0.78 | 0.71 | 0.62 | |
| 抽穗期 | 0.76 | 0.80 | 0.75 | 0.70 | 0.67 | |
| 完熟期 | 0.80 | 0.76 | 0.84 | 0.78 | 0.88 | |
| 2021 | 苗期 | 0.72 | 0.74 | 0.75 | 0.68 | 0.71 |
| 喇叭口期 | 0.76 | 0.76 | 0.73 | 0.72 | 0.69 | |
| 抽穗期 | 0.71 | 0.74 | 0.73 | 0.65 | 0.72 | |
| 完熟期 | 0.75 | 0.74 | 0.68 | 0.73 | 0.65 |
同一生育期HGK60与郑58玉米田间节肢动物的群落稳定性差异较小(表 3)。Nn/Np仅在2021年抽穗期和完熟期表现出HGK60与郑58的显著差异(P<0.05);Nd/Np仅在2020年的苗期表现为HGK60与郑58间差异显著(P<0.05);Sn/Sp连续3年各生育期均表现为二者差异不显著(P>0.05), Sd/Sp仅在2021年的完熟期表现为HGK60与郑58差异显著(P<0.05)。总体上, 玉米各生育期田间节肢动物的群落稳定性指标Nn/Np、Nd/Np、Sn/Sp、Sd/Sp相对稳定, 田间节肢动物群落中天敌类群和中性类群对植食性类群的调控作用未因HGK60的种植产生显著变化。
| 年份 Year |
生育期 Growth period |
玉米材料 Maize treatments |
稳定性指数Stability index | |||
| Nn/Np | Nd/Np | Sn/Sp | Sd/Sp | |||
| 2019 | 苗期 | HGK60 | 1.02±0.14 | 0.24±0.03 | 1.13±0.25 | 0.15±0.10 |
| 郑58 | 0.86±0.03 | 0.20±0.05 | 1.18±0.27 | 0.12±0.02 | ||
| 喇叭口期 | HGK60 | 0.92±0.02 | 0.22±0.02 | 1.11±0.18 | 0.15±0.05 | |
| 郑58 | 0.89±0.08 | 0.23±0.05 | 0.99±0.38 | 0.11±0.04 | ||
| 抽穗期 | HGK60 | 1.02±0.05 | 0.28±0.02 | 1.09±0.23 | 0.14±0.02 | |
| 郑58 | 0.92±0.12 | 0.24±0.02 | 1.12±0.22 | 0.11±0.05 | ||
| 完熟期 | HGK60 | 1.14±0.06 | 0.26±0.03 | 1.29±0.29 | 0.22±0.07 | |
| 郑58 | 1.23±0.05 | 0.28±0.02 | 1.24±0.11 | 0.21±0.02 | ||
| 2020 | 苗期 | HGK60 | 0.84±0.08 | 0.19±0.02* | 1.14±0.26 | 0.17±0.04 |
| 郑58 | 0.76±0.08 | 0.16±0.04 | 1.18±0.37 | 0.16±0.05 | ||
| 喇叭口期 | HGK60 | 0.82±0.09 | 0.20±0.03 | 1.02±0.21 | 0.17±0.03 | |
| 郑58 | 0.90±0.06 | 0.17±0.01 | 1.02±0.05 | 0.14±0.02 | ||
| 抽穗期 | HGK60 | 0.93±0.06 | 0.19±0.01 | 1.07±0.29 | 0.19±0.02 | |
| 郑58 | 0.93±0.01 | 0.20±0.03 | 1.09±0.14 | 0.18±0.06 | ||
| 完熟期 | HGK60 | 1.17±0.05 | 0.27±0.03 | 1.05±0.17 | 0.24±0.04 | |
| 郑58 | 1.08±0.04 | 0.25±0.02 | 1.03±0.12 | 0.23±0.04 | ||
| 2021 | 苗期 | HGK60 | 0.52±0.09 | 0.10±0.03 | 0.56±0.12 | 0.12±0.04 |
| 郑58 | 0.44±0.12 | 0.13±0.07 | 0.65±0.16 | 0.15±0.04 | ||
| 喇叭口期 | HGK60 | 0.61±0.11 | 0.09±0.01 | 0.53±0.01 | 0.10±0.01 | |
| 郑58 | 0.61±0.19 | 0.11±0.05 | 0.59±0.10 | 0.13±0.04 | ||
| 抽穗期 | HGK60 | 0.72±0.22* | 0.09±0.04 | 0.54±0.07 | 0.10±0.02 | |
| 郑58 | 0.55±0.22 | 0.12±0.04 | 0.50±0.16 | 0.12±0.04 | ||
| 完熟期 | HGK60 | 0.53±0.24* | 0.11±0.02 | 0.46±0.14 | 0.12±0.02* | |
| 郑58 | 0.77±0.12 | 0.08±0.04 | 0.57±0.05 | 0.07±0.03 | ||
| Nn: 天敌类群个体数the number of individuals of natural group; Np: 植食性类群个体数the number of individuals of herbivorous group; Nd: 中性类群个体数the number of individuals of neutral group; Sn: 天敌类群物种数the number of species of natural group; Sp: 植食性类群物种数the number of species of herbivorous group; Sd: 中性类群物种数the number of species of neutral group; 表中数据(均数±标准差)加上星号表示HGK60和郑58之间经t检验在P<0.05时有显著性差异 | ||||||
随着转基因作物研发工作的不断推进, 农业农村部已陆续发放多个转基因生物安全证书, 转基因作物种植是否会带来环境安全性问题也得到了越来越多的关注, 其可能存在环境安全问题主要包括转基因植物入侵、对非靶标生物的影响、靶标昆虫抗性的产生、次生害虫的爆发等几个方面。节肢动物是农田生物多样性重要的组成部分, 是农田中最丰富的生物类群之一[17—18], 同时节肢动物多样性随环境的变化也是评估农田生态环境质量的重要指标之一[19]。
通过对田间节肢动物的调查发现, 整体上转Cry1Ah基因抗虫玉米HGK60与对照常规玉米郑58在田间节肢动物群落组成与结构上没有显著差异, 在相对丰度、多样性、相似性、稳定性等方面偶尔出现的差异并未持续、重复存在。在前期研究工作中, 我们在其他地区、不同播种季也进行了转Cry1Ah基因抗虫玉米HGK60对田间节肢动物的影响研究, 与此次连续多年的调查结果基本一致, 可以说即使在不同的生态区位上、在各非生物因素的影响下, HGK60种植均未对田间节肢动物产生显著影响[20]。许多专家学者研究亦认为转基因作物种植对土壤动物多样性的影响很小甚至没有[21—23]。对长春地区田间节肢动物调查发现, Bt-799的群落组成、群落结构和对照玉米之间没有显著差异, 季节变化也一致[24]。郭井菲等人连续2年采用直接观察、气盆诱捕器、地面诱捕器和昆虫引诱器4种方法, 得出转基因玉米田间节肢动物数量、多样性指数、优势集中指数和节肢动物均匀度指数与对照无显著差异的结论, 表明转Cry1Ie基因玉米的种植对田间节肢动物群落的结构和多样性没有显著影响[25]。Habustova在ČESKÉBUDĚJOVICE连续3年监测转基因Bt玉米田间节肢动物, 发现转基因Bt作物和对照之间的生物量和物种丰度没有显著差异[26]。Wang等在北京调查了两年转植酸酶玉米田节肢动物群落的组成和丰度, 结果表明, 转植酸酶玉米与对照玉米田间节肢动物在物种数量、种体数量、群落多样性指数、相似系数和群落相对稳定性等方面均无显著差异[12]。此外, 许多对转基因大豆、水稻的生态风险评估也表明, 转基因大豆、水稻等的种植不会对田间节肢动物产生显著影响[27]。亦有专家学者认为, 转基因抗虫作物的种植有效减少了杀虫剂的使用, 降低了杀虫剂对田间节肢动物的影响从而增加了其丰度和群落多样性, 有益于种植区域生态系统稳定性的提高[28—29]。
从营养类群的角度来看, HGK60的种植并未对植食性类群、天敌类群以及中性类群产生持续且显著的影响。目前, 转基因作物种植对田间节肢动物影响的研究主要集中在植食性和天敌两大类群, 主要通过研究“信息链影响植物-植食性昆虫-天敌间的化学信息通讯”等分析对植食性昆虫和天敌的群落组成和种群动态的影响[30]。1999年, John Losey研究发现植食性昆虫黑脉金斑蝶取食拌有转Bt基因抗虫玉米花粉后死亡率升高[7], 在科学界引起风暴, 但后续科研人员针对黑脉金斑蝶开展的研究结果并不支持此结论[31—32]。部分对天敌昆虫的研究认为, 转基因抗虫植物的利用会明显降低靶标害虫的危害, 从而降低虫害诱导植物挥发物(HIPVs)的释放, 最终影响植物对天敌昆虫行为的调控作用[33], 从而一定程度上造成田间天敌昆虫数量的改变[30, 34], 但多数研究与本结果较为一致, 认为转基因抗虫作物的种植对天敌昆虫的影响并不显著[20, 35—36], CrylAb、CrylAc、Cry2Aa、Cry2Ab、CrylAh、CrylF等杀虫蛋白混入饲料后对天敌昆虫的生长发育亦无显著影响[37—39]。目前, 对中性昆虫的影响研究相对较少, 但仅仅分析对植食性与天敌昆虫的影响是不够的[40—41]。生态学意义上, 中性昆虫属基位物种, 是中位、顶位物种的捕食对象之一, 对研究生态系统的物质和能量循环有重要意义[42], 是不可忽视的一个类群。因此, 中性昆虫也将是此后研究的重点。
5 结论从玉米的整个生长阶段来看, 转Cry1Ah基因抗虫玉米HGK60的种植对田间节肢动物的种类、数量、生态指标以及群落相似性与稳定性等均未造成显著影响。偶尔出现的短暂性差异因存在不可重复性, 不能否定HGK60种植对田间节肢动物无显著影响的结论。
尽管本研究认为HGK60种植对田间节肢动物无显著影响, 但是按照“个案”原则[43], 转入不同基因而产生的不同转基因事件其表现可能存在差异, 其他转基因作物对田间节肢动物的影响仍然存在争议。另一方面, 转基因作物的环境安全评估是一个长期的研究过程, 需要在多个地点进行连续多年的检测, 以丰富重复检测结果, 获得更全面和可靠的结论。
| [1] |
李浩辉, 刘彩月, 张海文, 王旭静, 唐巧玲, 王友华. 2022年度全球转基因作物产业化发展现状及趋势分析. 中国农业科技导报, 2023, 25(12): 6-16. |
| [2] |
Nawaz M A, Mesnage R, Tsatsakis A M, Golokhvast K S, Yang S H, Antoniou M N, Chung G. Addressing concerns over the fate of DNA derived from genetically modified food in the human body: A review. Food and Chemical Toxicology, 2019, 124: 423-430. DOI:10.1016/j.fct.2018.12.030 |
| [3] |
Tabashnik B E, Carrière Y. Surge in insect resistance to transgenic crops and prospects for sustainability. Nature Biotechnology, 2017, 35: 926-935. DOI:10.1038/nbt.3974 |
| [4] |
Campos S O, Santana I V, Silva C, Santos-Amaya O F, Guedes R N C, Pereira E J G. Bt-induced hormesis in Bt-resistant insects: theoretical possibility or factual concern?. Ecotoxicology and Environmental Safety, 2019, 183: 109577. DOI:10.1016/j.ecoenv.2019.109577 |
| [5] |
Guan Z J, Wei W, Stewart C N, Tang Z X. Effects of transgenic oilseed rape harboring the Cry1Ac gene on microbial communities in the rhizosphere soil. European Journal of Soil Biology, 2021, 103: 103277. DOI:10.1016/j.ejsobi.2021.103277 |
| [6] |
Mandal A, Sarkar B, Owens G, Thakur J K, Manna M C, Niazi N K, Jayaraman S, Patra A K. Impact of genetically modified crops on rhizosphere microorganisms and processes: a review focusing on Bt cotton. Applied Soil Ecology, 2020, 148: 103492. DOI:10.1016/j.apsoil.2019.103492 |
| [7] |
Losey J E, Rayor L S, Carter M E. Transgenic pollen harms monarch larvae. Nature, 1999, 399: 214. |
| [8] |
戈峰, 欧阳芳, 赵紫华. 基于服务功能的昆虫生态调控理论. 应用昆虫学报, 2014, 51(3): 597-605. |
| [9] |
王尚, 王柏凤, 严杜升, 王军, 武奉慈, 席景会, 宋新元. 转EPSPS基因抗除草剂玉米CC-2对田间节肢动物多样性的影响. 生物安全学报, 2014, 23(4): 271-277. DOI:10.3969/j.issn.2095-1787.2014.04.008 |
| [10] |
任振涛, 薛堃, 周宜君. 转基因作物安全性评价研究的文献计量可视化分析. 生态与农村环境学报, 2021, 37(12): 1503-1513. |
| [11] |
Li X Y, Lang Z H, Zhang J, He K L, Zhu L, Huang D F. Acquisition of insect-resistant transgenic maize harboring a truncated cry1Ah gene via Agrobacterium-mediated transformation. Journal of Integrative Agriculture, 2014, 13(5): 937-944. DOI:10.1016/S2095-3119(13)60531-6 |
| [12] |
Wang M, Guan X. The effects of phytase transgenic maize on the community components and diversity of arthropods. Journal of Asia-Pacific Entomology, 2020, 23(4): 1228-1234. DOI:10.1016/j.aspen.2020.09.001 |
| [13] |
蔡晓明. 昆虫分类学(修订版). 北京: 化学工业出版社, 2017, 528-831. |
| [14] |
Chen Y J, Ren M Y, Pan L B, Liu B, Guan X, Tao J. Impact of transgenic insect-resistant maize HGK60 with Cry1Ah gene on community components and biodiversity of arthropods in the fields. PLoS One, 2022, 17(6): e0269459. DOI:10.1371/journal.pone.0269459 |
| [15] |
马克平, 刘灿然, 刘玉明. 生物群落多样性的测度方法Ⅱβ多样性的测度方法. 生物多样性, 1995(1): 38-43. DOI:10.3321/j.issn:1005-0094.1995.01.007 |
| [16] |
蒋杰贤, 万年峰, 季香云, 淡家贵. 桃园生草对桃树节肢动物群落多样性与稳定性的影响. 应用生态学报, 2011, 22(9): 2303-2308. |
| [17] |
段美春, 刘云慧, 王长柳, Axmacher J C, 李良涛, 宇振荣. 坝上地区不同海拔农田和恢复半自然生境下尺蛾多样性. 应用生态学报, 2012, 23(3): 785-790. |
| [18] |
章家恩. 中国农业生物多样性及其保护. 农村生态环境, 1999(2): 37-41. |
| [19] |
岳天祥. 生物多样性研究及其问题. 生态学报, 2001(3): 462-467. DOI:10.3321/j.issn:1000-0933.2001.03.019 |
| [20] |
陈彦君, 李俊生, 闫冰, 关潇, 陶均. 转Cry1Ah基因抗虫玉米HGK60对生物多样性的影响. 环境科学研究, 2021, 34(4): 964-975. |
| [21] |
Singh A K, Dubey S K. Current trends in Bt crops and their fate on associated microbial community dynamics: a review. Protoplasma, 2016, 253(3): 663-681. DOI:10.1007/s00709-015-0903-5 |
| [22] |
Marvier M, McCreedy C, Regetz J, Kareiva P. A meta-analysis of effects of Bt cotton and maize on nontarget invertebrates. Science, 2007, 316(5830): 1475-1477. DOI:10.1126/science.1139208 |
| [23] |
Farinós G P, Poza M D L, Hernández-Crespo P, Ortego F, Castañera P. Diversity and seasonal phenology of aboveground arthropods in conventional and transgenic maize crops in Central Spain. Biological Control, 2008, 44(3): 362-371. DOI:10.1016/j.biocontrol.2007.11.007 |
| [24] |
尹俊琦, 武奉慈, 周琳, 宋新元. 转Cry1Ac基因抗虫玉米Bt-799对田间节肢动物群落多样性的影响. 生物安全学报, 2017, 26(2): 159-167. DOI:10.3969/j.issn.2095-1787.2017.02.009 |
| [25] |
郭井菲, 张聪, 袁志华, 何康来, 王振营. 转cry1Ie基因抗虫玉米对田间节肢动物群落多样性的影响. 植物保护学报, 2014, 41(4): 482-489. |
| [26] |
Habuštová O S, Svobodová Z, Spitzer L, Doležal P, Hussein H M, Sehnal F. Communities of ground-dwelling arthropods in conventional and transgenic maize: background data for the post-market environmental monitoring. Journal of Applied Entomology, 2015, 139(1-2): 31-45. DOI:10.1111/jen.12160 |
| [27] |
Horak M J, Rosenbaum E W, Phillips S L, Kendrick D L, Carson D, Clark P L, Nickson T E. Characterization of the ecological interactions of Roundup Ready 2 Yield© soybean, MON 89788, for use in ecological risk assessment. GM Crops & Food, 2015, 6(3): 167-182. |
| [28] |
Bhatti M A, Duan J, Head G P, Jiang C J, McKee M J, Nickson T E, Pilcher C L, Pilcher C D. Field evaluation of the impact of corn rootworm (Coleoptera: Chrysomelidae)-protected Bt corn on foliage-dwelling arthropods. Environmental Entomology, 2005, 34(5): 1336-1345. DOI:10.1093/ee/34.5.1336 |
| [29] |
Bitzer R J, Rice M E, Pilcher C D, Pilcher C L, Lam W F. Biodiversity and community structure of epedaphic and euedaphic springtails (Collembola) in transgenic rootworm Bt corn. Environmental Entomology, 2005, 34(5): 1346-1376. DOI:10.1093/ee/34.5.1346 |
| [30] |
刘清松, 李云河, 陈秀萍, 彭于发. 转基因抗虫植物-植食性昆虫-天敌间化学通讯的研究进展. 应用生态学报, 2014, 25(8): 2431-2439. |
| [31] |
Hellmich R L, Siegfried B D, Sears M K, Stanley-Horn D E, Daniels M J, Mattila H R, Spencer T, Bidne K G, Lewis L C. Monarch larvae sensitivity to Bacillus thuringiensis- purified proteins and pollen. Proceedings of the National Academy of Sciences of the United States of America, 2001, 98(21): 11925-11930. |
| [32] |
Sears M K, Hellmich R L, Stanley-Horn D E, Oberhauser K S, Pleasants J M, Mattila H R, Siegfried B D, Dively G P. Impact of Bt corn pollen on monarch butterfly populations: A risk assessment. Proceedings of the National Academy of Sciences of the United States of America, 2001, 98(21): 11937-11942. |
| [33] |
Ibrahim M A, Stewart-Jones A, Pulkkinen J, Poppy G M, Holopainen J K. The influence of different nutrient levels on insect-induced plant volatiles in Bt and conventional oilseed rape plants. Plant Biology, 2008, 10(1): 97-107. DOI:10.1111/j.1438-8677.2007.00013.x |
| [34] |
李保平, 孟玲, 万方浩. 转基因抗虫植物对天敌昆虫的影响. 中国生物防治, 2002(3): 97-105. |
| [35] |
马月, 刘娜, 谢彦博, 梁晋刚, 李飞武. 转基因抗虫玉米对节肢动物多样性影响研究进展. 中国生物防治学报, 2022, 38(5): 1135-1142. |
| [36] |
Guo J F, He K L, Hellmich R L, Bai S X, Zhang T T, Liu Y J, Ahmed T, Wang Z Y. Field trials to evaluate the effects of transgenic cry1Ie maize on the community characteristics of arthropod natural enemies. Scientific Reports, 2016, 6: 22102. DOI:10.1038/srep22102 |
| [37] |
Ali I, Zhang S, Muhammad M S, Iqbal M, Cui J J. Bt proteins have no detrimental effects on larvae of the green lacewing, Chrysopa pallens (Rambur) (Neuroptera: Chrysopidae). Neotropical Entomology, 2018, 47(3): 336-343. DOI:10.1007/s13744-017-0526-y |
| [38] |
李亚荣, 张帅, 雒珺瑜, 朱香镇, 王丽, 崔金杰. Cry2Ab蛋白对龟纹瓢虫的安全性研究. 生物安全学报, 2019, 28(3): 195-199. |
| [39] |
Li Y H, Hu L, Romeis J, Wang Y N, Han L Z, Chen X P, Peng Y F. Use of an artificial diet system to study the toxicity of gut-active insecticidal compounds on larvae of the green lacewing Chrysoperla sinica. Biological Control, 2014, 69: 45-51. DOI:10.1016/j.biocontrol.2013.10.017 |
| [40] |
刘雨芳, 古德祥. 稻田中性昆虫多样性及其生态功能分析. 中国生物防治, 2002(4): 149-152. |
| [41] |
徐建祥, 吴进才. 综论稻田生态系中性昆虫的意义及其调控. 生态学杂志, 1999(5): 6+41-44. |
| [42] |
吴进才, 胡国文, 唐健, 束兆林, 杨金生, 万志农, 任正才. 稻田中性昆虫对群落食物网的调控作用. 生态学报, 1994(4): 381-386. DOI:10.3321/j.issn:1000-0933.1994.04.015 |
| [43] |
刘标, 韩娟, 薛堃. 转基因植物环境监测进展. 生态学报, 2016, 36(9): 2490-2496. |