 2024, Vol. 44
2024, Vol. 44文章信息
- 段成伟, 李希来, 李成一, 杨鹏年, 柴瑜, 徐文印
- DUAN Chengwei, LI Xilai, LI Chengyi, YANG Pengnian, CHAI Yu, XU Wenyin
- 黄河源区高寒草甸裸露斑块形成机理与生态修复关键限制因子
- Formation mechanism of alpine meadow bare patches and key factors limiting their restoration in the Source Zone of the Yellow River
- 生态学报. 2024, 44(16): 6952-6964
- Acta Ecologica Sinica. 2024, 44(16): 6952-6964
- http://dx.doi.org/10.20103/j.stxb.202308051674
-
文章历史
- 收稿日期: 2023-08-05
- 网络出版日期: 2024-06-24






2. 青海大学高原生态与农业国家重点实验室, 西宁 810016
2. State Key Laboratory of Plateau Ecology and Agriculture, Qinghai University, Xining 810016, China
高寒草甸是高原地带性与山地垂直带谱的主要植被类型[1]。高寒草甸所处区域气候恶劣、生存环境严酷、生态系统结构简单、自我更新能力脆弱, 其植被组成为多年生草本植物, 莎草科、禾草科以及杂类草都很丰富, 密丛型短根茎嵩草属为重要组成植物。近年来, 由于气候变化、超载过牧等不合理的人类活动, 青藏高原已有三分之一的天然草地出现不同程度的退化[2—4]。研究表明, 黄河源区高寒草甸的重度退化面积为可利用草地面积的一半以上[5]。过度放牧和牲畜的严重践踏导致大面积高寒草甸草毡层被剥蚀[6], 高原鼠兔(Ochotona curzoniae)等啮齿动物的持续扰动和大量暴发导致高寒草甸植破碎化程度加剧, 造成高寒草甸裸露和斑块的出现[7]。裸露斑块的出现是高寒草甸退化的重要特征, 裸露斑块产生和扩大, 最终导致黑土滩形成, 也称其为斑块化退化。斑块化退化高寒草甸主要特征为草毡层破坏、草地破碎化、鼠害严重[8]。裸露斑块面积是反映草甸退化程度的重要指标, 裸露斑块的出现和毒杂草的蔓延相辅相成, 降低了优良牧草的竞争优势, 导致草地生态系统的组成、结构与功能发生逆行演替, 阻碍了当地畜牧业的发展[9]。
黄河源区高寒草甸的退化和局部难以持续恢复是一个普遍关注的科学问题[10—12]。草地斑块化是青藏高原高寒草甸中重度退化的主要表现[13—14], 裸露斑块的修复一直是退化高寒草甸生态系统修复的关键和难点[15]。为了促进青藏高原黄河源区高寒草甸的生态稳定与可持续发展, 需要对斑块化退化高寒草甸进行恢复治理, 防止进一步退化为“黑土滩”。不同退化斑块的自然演替恢复至少需要15—20 a[15—16], 而人工干预恢复措施, 比如人工施肥、补播优良牧草、镇压土壤及围栏封育能缩短自然演替进程, 加快斑块化退化高寒草甸的恢复速度。目前, 黄河源区高寒草甸退化的机理和恢复研究虽然取得了一些进展[11, 17—19], 但现有的恢复技术没有考虑地形和水热条件的影响, 导致恢复过的生态系统不稳定[20]。而且, 常规的修复治理一般只注重免耕补播, 忽视了播种后的保护措施, 导致这种方式的效果不理想。因此, 探索黄河源区斑块化退化高寒草甸修复的机理并研究加快退化草地修复进程的有效措施, 已成为青藏高原高寒草甸生态建设与畜牧业发展面临的首要问题。
长期过度放牧和牲畜践踏, 很容易导致草甸草毡层的剥蚀[6]。目前已知, 啮齿动物的频繁活动和暴发是导致高寒草甸生态系统退化的主要原因, 特别是高原鼠兔[7]。它们的频繁扰动造成健康高寒草甸裸露斑块的出现和形成, 并使高寒草甸破碎化。加之水蚀、风蚀和冻融作用, 高寒草甸会斑块化退化形成一定深度的塌陷坑, 也就是裸露斑块[16]。同时, 野外调查发现, 地势较低的斑块化退化高寒草甸免耕补播容易成功, 地势相对较高的斑块化退化高寒草甸免耕补播措施治理效果则不甚理想(图 1), 因此初步判断斑块土壤水分可能是影响高原鼠兔扰动形成不同退化斑块生态修复的关键限制因子。在修复高原鼠兔扰动形成的不同退化斑块时, 补播牧草的同时需要进行土壤镇压来保持土壤水分[16]。

|
| 图 1 黄河源区高原鼠兔和高原鼠兔+高原鼢鼠扰动退化斑块的恢复效果 Fig. 1 Recovery effect of interference degraded patches of plateau pikas and plateau pikas + plateau zokors by no-tillage reseeding in the Source Zone of the Yellow River 图中红色部分可以清楚地看到免耕补种的痕迹 |
本研究通过比较不同斑块自然恢复演替阶段(裸露斑块(Bare patches, BP)、短期恢复斑块(Short-term recovered patches, SRP)、长期恢复斑块(Long-term recovered patches, LRP))和健康草甸(Healthy alpine meadows, AM)根-土-石复合体中土壤质地组成, 揭示黄河源区高寒草甸裸露斑块的形成机理。同时, 对高原鼠兔扰动形成的裸露斑块土壤进行室内花盆镇压试验, 试图找出影响斑块化退化高寒草甸生态修复的关键限制因子, 探讨斑块化退化高寒草甸生态恢复技术, 为黄河源区高寒草地生态修复和保护提供理论依据。
1 研究区概况与研究方法 1.1 研究区概况斑块化退化高寒草甸研究所需的土壤样品采集于青海省河南县优干宁镇泽曲河流域克其合滩阳坡滩地(34°40′48″—34°41′0.84″ N, 101°46′2″—101°46′24″ E, 海拔3689 m)(图 2)。样地坡向南偏西30°, 坡度为3°—5°, 地势呈东高西低。草地类型为高寒草甸, 土壤为山地草甸土。研究区高寒草甸已发生重度退化, 高原鼠兔活动频繁, 植被盖度占35%, 裸露秃斑面积占65%, 属于典型的斑块化退化的黑土滩草地。该地区分布着广泛的斑块化退化高寒草甸, 其植被以矮生嵩草(Kobresia humilis)、青藏苔草(Carex moorcroftii)、黄花棘豆(Oxytropis ochrocephala)、垂穗披碱草(Elymus nutans)和冷地早熟禾(Poa crymophila)为优势种, 鹅绒委陵菜(Potentilla anserina)、黄帚橐吾(Ligularia virgaurea)和密花香薷(Elsholtzia densa)为主, 并伴有其他杂草。

|
| 图 2 样地分布图 Fig. 2 Plots and sampling points distribution |
(1) 2022年8月下旬在青海省河南县优干宁镇泽曲河流域克其合滩阳坡滩地, 随机选取50 m×150 m斑块化退化高寒草甸, 然后分成50 m×50 m三个重复样地。根据斑块退化不同自然演替阶段, 将高寒草甸斑块分为4种演替类型:BP、SRP、LRP和AM(图 3)。BP指土壤物理结皮已经形成的裸露塌陷地, 伴有明显高原鼠兔经常扰动的痕迹。SRP是经过控鼠并定居杂类草的恢复演替斑块, 植被盖度小于40%, 植物种类主要以鹅绒委陵菜、密花香薷和黄帚橐吾为主。LRP是通过自然演替并伴有禾本科和莎草科分布的演替斑块, 如垂穗披碱草和青藏苔草、鹅绒委陵菜、密花香薷等, 植被盖度大于40%。AM是未退化的原生健康草甸, 植被类型以矮生嵩草为主。采集单位体积草甸土壤和裸露斑块(BP)、短期恢复斑块(SRP)、长期恢复斑块(LRP)3种类型斑块表层(0—5 cm)土壤, 每种斑块9个重复, 共计36个斑块(图 4)。

|
| 图 3 斑块化退化高寒草甸在自然演替阶段的不同类型斑块 Fig. 3 Different types of patches in the natural restoration succession stages of patchily degraded alpine meadows |

|
| 图 4 研究区不同类型斑块和三个样地的样地分布特征 Fig. 4 Different types of patches in the study area and distribution characteristics of sample plots in three plots |
(2) 2022年5月下旬采集裸露斑块表层(0—20 cm)土壤, 布置室内花盆试验。在草地植物返青前将高原鼠兔扰动形成的裸露斑块表层0—20 cm土壤样品采集运回实验室, 在花盆里进行室内土壤镇压试验。花盆规格为23.5 cm×19 cm×25 cm(上口径×下口径×高), 材质为耐压塑料。具体方法为:将斑块土壤放置花盆中, 与花盆上沿相差2 cm, 进行有机肥+牧草补播配施处理。有机肥和牧草补播量依据前人研究[25]确定, 具体为:有机肥(150 g/m2)+牧草补播(1.5 g/m2同德短芒披碱草(Elymus breviaristatus cv. Tongde)+0.75 g/m2青海中华羊茅(Festuca sinensis cv. Qinghai)+0.75 g/m2青海冷地早熟禾(Poa crymophila cv. Qinghai), 按2∶1∶1比例混播。有机肥+牧草补播配施处理的方式为:将有机肥和补播牧草草种混合后人工撒施在土壤表层, 然后覆土, 并将表层土壤耙平至与花盆上沿齐平。用500 g橡皮锤进行土壤镇压, 镇压深度分别为0 cm(S0)、5 cm(S5)、10 cm(S10)、15 cm(S15), 每个处理重复4次, 共计16个花盆试验。试验在实验室进行, 自然采光, 白天温度控制在(20±5)℃, 夜间温度(15±5)℃, 培养周期为45 d。培养期间, 所有的花盆置于阳面窗户旁边并每天轮流调换花盆位置。同时, 每周浇灌一次水, 每盆水量为300 mL, 水量相当于研究区返青期的降雨量(图 5)。

|
| 图 5 室内花盆镇压试验 Fig. 5 Indoor flowerpot compaction test S0:镇压深度0 cm The depth of compression is 0 cm;S5:镇压深度5 cm The depth of compression is 5 cm;S10:镇压深度10 cm The depth of compression is 10 cm;S15:镇压深度15 cm The depth of compression is 15 cm |
2020年9月初在研究区随机选取的3块50 m×50 m斑块化退化高寒草甸样地进行土壤样品采集。对于BP、SRP、LRP 3种类型斑块进行0—5 cm表层土壤样品采集;对于AM, 根据裸露斑块的深度和大小确定草甸土体积进行采集草甸土块。将3种类型斑块土壤样品和草甸土块带回实验室, 3种类型斑块土壤样品置于通风处自然风干, 用于测试土壤质地组成。将草甸土块选取10 cm×10 cm×10 cm(长×宽×高)进行机械压扁, 挑出较大石块, 以防止损坏石块, 最终成为根-土-石块复合体。对石块、根系和土壤颗粒进行称重, 计算单位体积重量。同时, 测定并计算单位体积石块、根系、土壤颗粒占比。
在室内花盆培养期间, 利用便携式三参仪TDR 350测定土壤体积含水率、电导率和温度日变化。室内花盆培养结束后, 将地上部分植被齐平地面进行刈割并装进信封带称重得到地上鲜重, 以代表地上生物量。用土壤紧实度检测仪测定每个花盆的土壤紧实度, 用环刀在0—10 cm土层垂直剖面水平方向采集土壤样品进行土壤容重和根-土复合体剪切试验测定, 并计算土壤蓄水量。用药勺采集0—10 cm土层土壤样品, 部分新鲜土壤样品进行土壤湿度测定, 剩余新鲜土壤样品在5 mm筛下进行过筛, 过完筛的每个花盆内土样剔除动物残体和小石块, 制备成1个土样置于通风处自然风干, 用于测试土壤pH。
1.4 测定项目与方法(1) 各组分单位重量占比测定
采用称重法测定各组分单位体积重量。
(2) 各组分单位体积占比测定
采用容积体积法测定组分单位体积占比[21]。具体操作为:首先将机械压扁的根-土-石块复合体区分成石块、根系和土壤样品。然后将石块、根系和土壤样品转移到1000 mL容量瓶中用水定容, 得出石块、根系和土壤总体积;再将石块和根系进行洗涤, 将石块和根系转移到500 mL容量瓶中用水定容, 得出石块和根系体积;最后将石块挑出, 将根系转移到250 mL容量瓶中用水定容, 得出根系体积。
(3) 土壤质地组成测定
采用土壤激光粒度分析仪(Winner 2309A)测定土壤质地组成[22]。
(4) 利用铝盒烘干法测定土壤湿度, 计算公式:
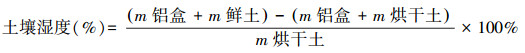
|
(5) 土壤pH采用pH计测定(水土比:1∶2.5)。
(6) 土壤容重采用环刀烘干法测定。
(7) 土壤蓄水量(mm)=土壤容重(g/cm3)×土层深度(cm)×土壤含水量(%)。
(8) 根-土复合体直剪试验:采用ZJ型应变控制式直剪仪进行室内花盆根-土复合体剪切试验。
(9) 土壤紧实度:土壤紧实度检测仪(SC900)。
1.5 数据分析Excel 2010软件用于数据整理和分析, 运用OriginPro 2021(OriginLab, 北安普顿, 马萨诸塞州, 美国)和SigmaPlot 13.0(Systat软件公司, 伊利诺伊州, 芝加哥)进行绘图。采用SPSS 26.0(IBM公司, 纽约州, 美国)进行相关统计分析, 采用Tukey进行差异显著性检验(P < 0.05)。使用线性回归分析确定不同镇压深度下植物地上生物量与土壤性质的关系。
2 结果与分析 2.1 健康草甸组分组成和不同退化斑块间土壤质地组成变化分析由图 6可知, 健康草甸包括土壤颗粒、根系和石块三部分组成。单位体积草甸内各组分重量占比差异显著(P < 0.05), 三种组分单位重量占比从大到小排列为:土壤颗粒(0.65±0.013)>根系(0.11±0.003)>石块(0.02±0.001)。各组分间单位体积占比差异显著(P < 0.05), 三种组分单位体积占比从大到小排列为:根系(0.62±0.003)>土壤颗粒(0.30±0.004)>石块(0.06±0.002)。

|
| 图 6 健康草甸各组分单位重量和体积占比变化 Fig. 6 The changes of unit mass and volume ratio of components in healthy meadow 不同小写字母表示各组分间差异显著(P < 0.05) |
图 7反映了退化斑块自然演替阶段不同类型斑块土壤质地组成变化, 四种类型斑块间土壤质地组成(砂粒、粉粒和黏粒)差异均不显著(P>0.05)。

|
| 图 7 退化斑块自然演替阶段不同类型斑块土壤质地组成 Fig. 7 Soil texture composition of different types in the natural succession stages of degraded patches 不同小写字母表示不同斑块间差异显著(P < 0.05);BP:裸露斑块Bare patches;SRP:短期恢复斑块Short-term recovered patches;LRP:长期恢复斑块Long-term recovered patches;AM:健康草甸Healthy alpine meadows |
不同镇压深度下花盆内植物地上生物量差异显著。与对照(S0)相比, 随着镇压深度的增加植物地上生物量显著提高, 从S0到S15显著增加了73.86%, 且各处理间差异均极显著(P < 0.001)(图 8)。

|
| 图 8 不同镇压深度植物生物量的变化 Fig. 8 The changes of plant biomass at different intensity of compression 不同小写字母表示不同镇压深度间差异显著(P < 0.05) |
不同镇压深度土壤物理性质差异显著。与对照(S0)相比, 镇压深度显著提高土壤湿度, 从S0到S15显著增加了101.54%, 且各处理间差异均显著(P < 0.05);土壤pH不随镇压深度而显著变化;土壤容重随着镇压深度的增加而显著增加(P < 0.05);土壤蓄水量也随着镇压深度的增加而显著增加(P < 0.05)(表 1)。
| 镇压深度 Intensity of suppression |
土壤湿度 Soil moisture/% |
pH | 容重 Bulk density/(g/cm3) |
土壤蓄水量 Soil water storage/mm |
| S0 | 10.35±1.44c | 7.13±0.18a | 1.14±0.02b | 267.28±33.18c |
| S5 | 14.78±0.94b | 7.13±0.13a | 1.13±0.03b | 445.67±40.69b |
| S10 | 16.91±1.17b | 6.91±0.22a | 1.34±0.03a | 534.61±38.48b |
| S15 | 20.86±0.88a | 7.01±0.24a | 1.40±0.05a | 741.24±63.88a |
| 不同小写字母表示不同镇压深度间差异显著(P < 0.05) | ||||
室内花盆培养期间, 不同镇压深度下土壤湿度变化不同。在培养初期(前5 d), 不同镇压深度土壤湿度由高到低排列为:S10>S5>S15>S0;在6—11 d内不同镇压深度土壤湿度由高到低排列为:S10>S15>S5>S0;12—45 d内不同镇压深度土壤湿度由高到低排列为:S15>S10>S5>S0(图 9)。

|
| 图 9 不同镇压深度土壤湿度的变化 Fig. 9 The changes of soil humidity at different intensities of compression |
不同镇压深度下土壤电导率变化不同。在培养初期(前5 d), 不同镇压深度土壤电导率由高到低排列为:S10>S5>S15>S0;在6—45 d内不同镇压深度土壤电导率变化不稳定, 总体上由高到低排列为S15>S10>S5>S0(图 10)。

|
| 图 10 不同镇压深度土壤电导率的变化 Fig. 10 The changes of soil electrical conductivity at different intensities of compression |
不同镇压深度下土壤温度变化不同。在整个培养期间(1—45 d), 不同镇压深度土壤温度由高到低排列为:S15>S10>S5>S0(图 11)。

|
| 图 11 不同镇压深度土壤温度的变化 Fig. 11 The changes of soil temperature at different intensities of compression |
不同镇压深度土壤机械强度变化差异显著。与对照(S0)相比, 镇压深度显著增加土壤紧实度, 从S0到S15显著增加了1316.92%, 且各处理间差异均极显著(P < 0.001)(图 12)。土壤粘聚力随着镇压深度的增加而显著增加(P < 0.001), 但S5和S10间差异不显著(图 13)。根-土复合体抗剪强度随着镇压深度的增加而显著增加, 且各处理间差异均极显著(P < 0.001)(图 14)。

|
| 图 12 不同镇压深度土壤紧实度变化 Fig. 12 The changes of soil compactness at different intensities of compression 不同小写字母表示不同镇压深度间差异显著(P < 0.05) |

|
| 图 13 不同镇压深度土壤粘聚力变化 Fig. 13 The changes of soil cohesion at different intensities of compression 不同小写字母表示不同镇压深度间差异显著(P < 0.05) |

|
| 图 14 不同镇压深度根-土复合体抗剪强度变化 Fig. 14 The changes of shear strength of the root-soil composite at different intensities of compression 不同小写字母表示不同镇压深度间差异显著(P < 0.05) |
土壤性质与植物地上生物量的线性回归分析表明, pH与地上生物量之间不存在显著相关性(P=0.4333), 土壤蓄水量、容重、紧实度、根-土复合体抗剪强度和湿度与地上生物量之间均存在极显著正相关关系(P < 0.001)(图 15)。

|
| 图 15 植物地上生物量与土壤物理性质和力学性质之间的线性关系 Fig. 15 Linear relationship between plant aboveground biomass and soil physical and mechanical properties |
从土壤理化性质对植物地上生物量标准总效应来看, 土壤蓄水量对植物地上生物量的总效应最大, 其次是土壤湿度, 土壤容重的总效应最小, 它们都是正效应。而pH对植物地上生物量为负效应(图 16)。

|
| 图 16 土壤理化性质对植物地上生物量标准总效应 Fig. 16 Total effect of soil physical and chemical properties on plant aboveground biomass |
高寒草甸出现的裸露斑块主要形成于阳坡滩地的退化高寒草甸中, 其土壤剥蚀呈现草毡表层死亡, 加之风蚀引起草毡层土壤颗粒的自然沉降形成的塌陷坑[16, 21]。其发生过程是由于过度放牧和高原鼠兔的挖掘与冻融交替作用的结果[16, 23]。
健康草甸单位体积根-土-石块复合体的研究发现, 健康草甸包括土壤颗粒、根系和石块三种组分。三种组分单位重量占比从大到小排列为:土壤颗粒(0.65±0.013)>根系(0.11±0.003)>石块(0.02±0.001)。三种组分单位体积占比从大到小排列为:根系(0.62±0.003)>土壤颗粒(0.30±0.004)>石块(0.06±0.002)。这表明健康高寒草甸在斑块化形成过程中, 高原鼠兔的持续扰动导致草毡表层发生剥离使得植被根系暴露, 自然风蚀作用加速了根系的脱水和死亡形成大量的根孔, 这部分根系很容易腐烂被风吹走;在放牧、高原鼠兔扰动和冻融交替作用下, 残余植物根系被矿化和微生物分解, 使得草毡表层破碎消失殆尽。没有植物根系的高寒草甸土壤颗粒在重力作用下沉至斑块表层形成塌陷坑, 风蚀作用将微细颗粒物吹走, 造成塌陷坑越来越深。这与曹广民等[21]对三江源地区退化高寒矮嵩草草甸剥蚀坑的成因研究、梁东营等[24]对三江源退化高寒草甸草毡层剥蚀过程及发生机理的初步研究和Duan等[16]对黄河源区不同草甸斑块土壤物理、化学和微生物群落特性分析研究结果一致。此外, 健康草甸中根系体积占比大于土壤颗粒, 石块体积最小, 根系体积为草甸单位总体积的3/5—3/4, 因此, 损失的总体积应为健康草甸表层体积的3/5—3/4, 与野外观测到塌陷坑的深度、面积和体积基本相当, 与曹广民等[21]研究结果表现一致。本研究还发现, BP、SRP、LRP和AM各斑块间土壤质地组成(砂粒、粉粒和黏粒)差异均不显著, 进一步说明健康草甸在过度放牧、啮齿动物(高原鼠兔)的挖掘与冻融交替作用下形成的斑块塌陷坑表层土壤颗粒就是健康草甸土壤颗粒自然沉降的结果。在裸露斑块自然演替阶段, 不同退化斑块土壤质地组成差异均不显著, 表明演替时间不影响裸露斑块自然演替进程中土壤质地的组成[14]。
在高寒草甸草毡层剥蚀阶段, 当草甸斑块形成大量的塌陷坑或裸露斑块后, 高原鼠兔密度就会大幅降低[25]。因此, 高寒草甸的草毡表层是其维持稳定性的策略之一, 采取人工修复斑块的植物多样性达到恢复各斑块的土壤根系组成是高寒草甸斑块化生态修复的必要和关键措施。
3.2 黄河源区高寒草甸裸露斑块生态修复关键限制因子分析本研究表明, 随着花盆试验镇压深度的增加植物地上生物量显著提高, 这与王鑫[26]不同镇压深度对土壤墒情及幼苗的影响研究结果基本一致。镇压通过增加土壤密度和紧实度, 让水分子的运动减弱, 使之不容易从土壤中挥发, 保持了土壤表层土壤含水量, 有利于种子吸收更多的水分并萌发[27]。土壤pH不随镇压深度而显著变化, 土壤容重随着镇压深度的增加而显著增加, 土壤蓄水量也随着镇压深度的增加而显著增加, 这与党红凯[28]和张迪等[27]报道的结果一致, 表明镇压有显著的紧实土壤和保墒作用。室内花盆培养期间, 1—5 d内不同镇压深度土壤湿度由高到低排列为:S10>S5>S15>S0;在6—11 d内不同镇压深度土壤湿度由高到低排列为:S10>S15>S5>S0;12—45 d内不同镇压深度土壤湿度由高到低排列为:S15>S10>S5>S0。表明镇压深度与土壤湿度有关, 镇压深度10—15 cm最有利紧实土壤和保墒作用。1—5 d内不同镇压深度土壤电导率由高到低排列为:S10>S5>S15>S0;在6—45 d内不同镇压深度土壤电导率变化不稳定, 总体上由高到低排列为:S15>S10>S5>S0。土壤电导率变化与湿度基本一致, 这与相关研究相反[29—30]。在整个培养期间(1—45 d), 不同镇压深度土壤温度由高到低排列为:S15>S10>S5>S0。究其原因, 镇压使土壤紧实、孔隙度减小、土壤容重增加, 土壤容积热容量增大, 有利于土壤增温保湿。土壤粘聚力和根-土复合体抗剪强度随着镇压深度的增加而显著增加。产生该现象的主要原因在于, 根系对抗剪强度具有显著性影响, 随着土壤地下生物量的增加, 其粘聚力表现出逐渐增大, 而镇压深度与地上生物量显著正相关, 故其土壤粘聚力增大。此外, 镇压深度越大, 土壤孔隙变小、紧实度增大, 土壤密度愈大导致抗剪强度也就愈强[31]。
土壤理化性质与植物地上生物量的线性回归分析表明, pH与地上生物量之间不存在显著相关性, 土壤蓄水量、容重、紧实度、根-土复合体抗剪强度和湿度与地上生物量之间均存在极显著正相关关系。从土壤理化性质对植物地上生物量标准总效应来看, 土壤蓄水量对植物地上生物量的总效应最大, 其次是土壤湿度, 土壤容重的总效应最小, 它们都是正效应。土壤pH对植物地上生物量为负效应。以上分析表明土壤水分就是影响高原鼠兔扰动形成不同退化斑块生态修复的关键限制因子。裸露斑块的修复需要进行人工镇压或者覆盖遮阴网来给土壤保墒, 提高土壤湿度, 增加土壤蓄水量, 尤其在播种后不久, 以便有利于种子吸收水分并萌发生长[16, 23]。
4 结论高寒草甸裸露斑块形成机理主要表现为, 过度放牧+鼠类暴发, 导致高寒草甸草毡层坏死剥蚀, 出现裸露斑块高寒草甸破碎化, 高原鼠兔暴发的持续扰动干扰, 引起高寒草甸根系暴露土表, 被土壤微生物和土壤动物分解腐烂风蚀殆尽, 裸露斑块土壤颗粒自然沉降形成塌陷坑, 出现裸露斑块。
室内花盆试验验证了野外原位观测结果, 裸露斑块土壤镇压显著提高植物地上生物量、土壤紧实度、土壤湿度、蓄水量、容重、电导率、粘聚力和根-土复合体抗剪强度。人工镇压紧实土壤达到保墒作用维持土壤水分, 为补播的禾本科牧草种子的正常发芽并出苗提供了充足的水分。不同退化斑块生态修复有效镇压深度为10—15 cm。由此得出结论, 土壤水分的匮乏是青藏高原斑块化退化高寒草甸生态修复的关键限制因子, 采取重度机械紧压措施或者覆盖遮阴网/无纺布可以改善和保持退化高寒草甸各类斑块的表层水分, 有利于提高其生态修复的成效, 降低修复成本。
| [1] |
张金霞, 曹广民, 周党卫, 胡启武, 赵新全. 高寒矮嵩草草甸大气-土壤-植被-动物系统碳素储量及碳素循环. 生态学报, 2003, 23(4): 627-634. |
| [2] |
周华坤, 周立, 赵新全, 刘伟, 严作良, 师燕. 江河源区"黑土滩"型退化草场的形成过程与综合治理. 生态学杂志, 2003, 22(5): 51-55. |
| [3] |
Li X L, Perry G L W, Brierley G J, Sun H Q, Li C H, Lu G X. Quantitative assessment of degradation classifications for degraded alpine meadows (Heitutan), Sanjiangyuan, Western China. Land Degradation & Development, 2014, 25(5): 417-427. |
| [4] |
Lu X Y, Yan Y, Sun J, Zhang X K, Chen Y C, Wang X D, Cheng G W. Carbon, nitrogen, and phosphorus storage in alpine grassland ecosystems of Tibet: effects of grazing exclusion. Ecology and Evolution, 2015, 5(19): 4492-4504. DOI:10.1002/ece3.1732 |
| [5] |
赵新全, 周华坤. 三江源区生态环境退化、恢复治理及其可持续发展. 中国科学院院刊, 2005, 20(6): 471-476. DOI:10.3969/j.issn.1000-3045.2005.06.008 |
| [6] |
Sun J, Hou G, Liu M, Fu G, Zhan T Y, Zhou H K, Tsunekawa A, Haregewey N. Effects of climatic and grazing changes on desertification of alpine grasslands, Northern Tibet. Ecological Indicators, 2019, 107: 105647. DOI:10.1016/j.ecolind.2019.105647 |
| [7] |
Liu Y S, Fan J W, Shi Z J, Yang X H, Harris W. Relationships between plateau pika (Ochotona curzoniae) densities and biomass and biodiversity indices of alpine meadow steppe on the Qinghai-Tibet Plateau China. Ecological Engineering, 2017, 102: 509-518. DOI:10.1016/j.ecoleng.2017.02.026 |
| [8] |
Dong Q M, Zhao X Q, Wu G L, Shi J J, Ren G H. A review of formation mechanism and restoration measures of "black-soil-type" degraded grassland in the Qinghai-Tibetan Plateau. Environmental Earth Sciences, 2013, 70(5): 2359-2370. DOI:10.1007/s12665-013-2338-7 |
| [9] |
李世雄, 王玉琴, 王彦龙, 尹亚丽. 黄河源区不同退化阶段高寒草甸植被特征. 青海畜牧兽医杂志, 2020, 50(2): 27-34. |
| [10] |
Dong S K, Shang Z H, Gao J X, Boone R B. Enhancing sustainability of grassland ecosystems through ecological restoration and grazing management in an era of climate change on Qinghai Tibetan Plateau. Agriculture Ecosystems & Environment, 2020, 287: 106684. |
| [11] |
Wang J, Zhang P, Bao J T, Zhao J C, Song G, Yang H T, Huang L, He M Z, Li X R. Comparison of cyanobacterial communities in temperate deserts: A cue for artificial inoculation of biological soil crusts. Science of the Total Environment, 2020, 745: 140970. DOI:10.1016/j.scitotenv.2020.140970 |
| [12] |
尹亚丽, 李世雄, 马玉寿. 人工补播对退化高寒草甸土壤真菌群落特征的影响. 草地学报, 2020, 28(6): 1791-1797. |
| [13] |
Li J S, Shao X Q, Huang D, Shang J Y, Liu K S, Zhang Q, Yang X M, Li H, He Y X. The addition of organic carbon and nitrogen accelerates the restoration of soil system of degraded alpine grassland in Qinghai-Tibet Plateau. Ecological Engineering, 2020, 158: 106084. DOI:10.1016/j.ecoleng.2020.106084 |
| [14] |
Jiang X J, Zhu X A, Yuan Z Q, Li X G, Liu W J, Zakari S. Lateral flow between bald and vegetation patches induces the degradation of alpine meadow in Qinghai-Tibetan Plateau. Science of the Total Environment, 2021, 751: 142338. DOI:10.1016/j.scitotenv.2020.142338 |
| [15] |
宋梓涵, 李希来, 李杰霞, 卡着才让, 马戈亮. 高原鼠兔跑道对高寒草甸退化斑块扩大与连通的影响. 生态学杂志, 2020, 39(10): 3276-3284. |
| [16] |
Duan C W, Li X L, Li C Y, Yang P N, Shi Y, Chai Y, Xu W Y. Analysis on the soil physical, chemical, and microbial community properties of different alpine meadow patches in the Source Zone of the Yellow River, West China. Ecological Indicators, 2022, 144: 109531. DOI:10.1016/j.ecolind.2022.109531 |
| [17] |
孙华方, 李希来, 金立群, 李成一, 张静. 生物土壤结皮对黄河源区人工草地植被与土壤理化性质的影响. 草地学报, 2020, 28(2): 509-520. |
| [18] |
段成伟, 李希来, 马盼盼, 徐文印, 柴瑜, 苏乐乐, 杨鑫光. 人工修复措施对退化高寒草甸土壤养分及酶活性的影响. 西北农业学报, 2022, 31(4): 431-440. |
| [19] |
段成伟, 李希来, 柴瑜, 徐文印, 苏乐乐, 杨鑫光, 马盼盼. 人工修复下退化高寒草甸碳、氮、磷生态化学计量特征. 中国草地学报, 2022, 44(7): 23-32. |
| [20] |
Dai L C, Fu R Y, Guo X W, Du Y G, Lin L, Zhang F W, Li Y K, Cao G M. Long-term grazing exclusion greatly improve carbon and nitrogen store in an alpine meadow on the northern Qinghai-Tibet Plateau. Catena, 2021, 197: 104955. DOI:10.1016/j.catena.2020.104955 |
| [21] |
曹广民, 龙瑞军, 张法伟, 林丽, 李以康, 梁东营. 三江源地区退化高寒矮嵩草草甸剥蚀坑的成因. 草原与草坪, 2010, 30(2): 16-21. |
| [22] |
段世航, 崔若然, 江荣风, 范明生, 袁会敏. 激光衍射法测定土壤粒径分布的研究进展. 土壤, 2020, 52(2): 247-253. |
| [23] |
Duan C W, Li X L, Li C Y, Yang P N, Chai Y, Xu W Y. Positive effects of fungal β diversity on soil multifunctionality mediated by pH in the natural restoration succession stages of alpine meadow patches. Ecological Indicators, 2023, 144: 110122. |
| [24] |
梁东营, 林丽, 李以康, 王溪, 曹广民. 三江源退化高寒草甸草毡表层剥蚀过程及发生机理的初步研究. 草地学报, 2010, 18(1): 31-36. |
| [25] |
段成伟, 李希来, 柴瑜, 徐文印, 苏乐乐, 马盼盼, 杨鑫光. 不同修复措施对黄河源退化高寒草甸植物群落与土壤养分的影响. 生态学报, 2022, 42(18): 7652-7662. |
| [26] |
王鑫. 小麦镇压机不同镇压强度对墒情及苗情的影响. 现代农村科技, 2020, 590(10): 61-62. |
| [27] |
张迪, 王红光, 马伯威, 李瑞奇, 李雁鸣. 播后镇压和冬前灌溉对土壤条件和冬小麦生育特性的影响. 麦类作物学报, 2014, 34(06): 787-794. |
| [28] |
党红凯, 曹彩云, 郑春莲, 马俊永, 郭丽, 王亚楠, 李伟, 李科江. 造墒与播后镇压对小麦冬前耗水和生长发育的影响. 中国生态农业学报, 2016, 24(8): 1071-1079. |
| [29] |
马金玲. 不同灌水量下绿洲棉田土壤水分与盐分变化试验研究[D]. 乌鲁木齐: 中国科学院新疆生态与地理研究所, 2005.
|
| [30] |
杨红, 徐唱唱, 赛曼, 曹舰艇, 曹丽花, 张爱琴, 刘合满. 不同土地利用方式对土壤含水量、pH值及电导率的影响. 浙江农业学报, 2016, 28(11): 1922-1927. |
| [31] |
申紫雁, 刘昌义, 胡夏嵩, 周林虎, 许桐, 李希来, 李国荣. 黄河源区高寒草地不同深度土壤理化性质与抗剪强度关系研究. 干旱区研究, 2021, 38(02): 392-401. |