 2024, Vol. 44
2024, Vol. 44文章信息
- 张亚, 陈超, 于明含, 何莹莹, 张富崇, 张建玲
- ZHANG Ya, CHEN Chao, YU Minghan, HE Yingying, ZHANG Fuchong, ZHANG Jianling
- 模拟降水年际和年内变化对黑沙蒿群落物种多样性和生产力的影响
- Modeling the effects of inter- and intra-annual variability in precipitation on the species diversity and productivity of Artemisia ordosica community
- 生态学报. 2024, 44(15): 6865-6873
- Acta Ecologica Sinica. 2024, 44(15): 6865-6873
- http://dx.doi.org/10.20103/j.stxb.202401120113
-
文章历史
- 收稿日期: 2024-01-12
- 网络出版日期: 2024-05-24





2. 宁夏盐池毛乌素沙地生态系统国家定位观测研究站, 盐池 751500;
3. 水利部沙棘开发管理中心, 北京 100036
2. Yanchi Ecology Research Station of Mu Us Desert, Yanchi 751500, China;
3. Seabuckthorn Development and Management Center, Ministry of Water Resources, Beijing 100036, China
在全球气候变化的背景下, 全球变暖显著改变了全球水分循环, 造成中纬度干旱和半干旱内陆地区降水格局受到影响[1]。现有研究表明, 近30年来中国干旱和半干旱地区不仅仅遭受干旱年和湿润年的频繁交替, 年内降水时间分配也发生变化, 向集中强降水和干旱周期增长的极端降水格局演化[2—3]。由于降水资源是该区域植被生存的主要限制因子[4], 因此, 降水格局的变化对该区域内植被群落结构和功能的影响受到格外关注[5—6]。
物种多样性和生产力是度量群落组成结构和功能复杂性、指示环境承载力大小的重要指标[7—9], 其受降水的影响已被广泛探讨, 其中, 年际降水量变化对群落影响的研究成果最为丰富, 如马文静在内蒙古荒漠草原开展降水控制试验, 得到了随降水量增加物种多样性和植物群落生产力增加的结果[10];Liu等通过Meta分析, 证明物种多样性与群落生产力随降水量变化的同向响应规律[11—12]。另有研究发现, 植物群落除了降水量的直接影响以外, 年内降水的分配也会干扰物种组成和结构[13], 如朱国栋在内蒙古荒漠草原地区的研究表明, 短期极端干旱仍可以显著降低了Margalef丰富度指数、Simpson优势度指数和Shannon-Wiener多样性指数(P < 0.05), 改变了荒漠草原植物群落组成[14];更集中的强降水可以促使荒漠灌草群落向灌木化方向演替, 并带来更高的生产力[15—17]。
现阶段, 多数研究基于年际降水量变化和年内降水时间分配变化对植被群落的独立影响而展开[18], 但二者对植被的影响是否完全独立尚未可知。本研究认为, 在不同的降水量背景下, 降水分配对生态系统影响将具有显著差异, 如丰水年和干旱年的极端强降水对群落生产力的促进作用并不一致[19], 干旱年的极端干旱周期很可能会加剧生产力的降低和物种多样性的丧失[20]。因此, 基于年际降水量和年内降水分配双重变异的降水格局影响亟待探究, 二者交互效应的相关试验和理论有待完善。
本研究以我国毛乌素沙地的典型植被黑沙蒿(Artemisia ordosica)灌草群落为研究对象, 通过设置小区尺度的人工降水控制实验, 探明降水量和降水间隔时间的交互作用对黑沙蒿植物群落多样性和生产力的影响, 以期补充降水变化相关研究理论, 并为未来降水变化下干旱半干旱地区生态系统演变和稳定性预测提供依据。
1 材料与方法 1.1 研究区概况本研究在宁夏回族自治区盐池县毛乌素沙地生态系统国家定位研究站, 站区位于毛乌素沙地的西南部边缘(E107°61′, N37°79′), 该地处于半干旱区, 具有半干旱大陆性季风气候的特点。年均温度为8.1℃, 年平均日照时数为2867.3h, 年均降水量为319mm。土壤类型以风沙土为主, 土壤结构疏松。优势植物种主要有黑沙蒿、赖草(Leymus secalinus)、沙柳(Salix psammophila)、白草(Pennisetum centrasiaticum)等。
1.2 试验设计降水控制实验于2020—2022年生长季(5—9月)进行。根据气象资料统计, 该地区(1990—2019年)平均降水量为338mm, 年最高降水量为449mm, 年最低降水量为212mm。年最高降水量、年最低降水量与年均降水量的差值分别为37%和34%, 同时参考了其他北方降水控制试验的设计, 所以本实验就采用了增减水30%。因此, 以年平均降水量为对照(W), 设置减水30%(W-)、自然降水(W)、增水30%(W+)模拟年际降水量的变化。为了模拟年内降水时间分配, 本研究对降水事件间隔期进行统计。结果表明, 降水间隔为5d内的降水事件发生的概率最大, 约占降水事件的64.01%, 间隔期在15d以上的约占总降水事件的7.59%。故本实验选取5d的降水间隔时间模拟自然降水(T), 15d模拟长降水间隔期(T++)。基于近30年降水的季节分配比例, 对年总降水量进行了逐月分配, 具体降水量和降水频次如表 1所示[21]。通过人工喷洒模拟降水, 按照实验设定的降水处理进行人工降水控水实验。
| 月份 Month |
月平均降水量 Average monthly precipitation/mm |
降水间隔期 Precipitation interval |
平均每次降水量 Average precipitation per event/mm |
月降水频次 Monthly precipitayion frequency |
||
| W- | W | W+ | ||||
| 5 | 33.09 | T | 3.86 | 5.52 | 7.17 | 6 |
| T++ | 11.58 | 16.53 | 21.51 | 2 | ||
| 6 | 41.08 | T | 4.79 | 6.85 | 8.90 | 6 |
| T++ | 14.38 | 20.54 | 26.70 | 2 | ||
| 7 | 72.39 | T | 8.45 | 12.07 | 15.68 | 6 |
| T++ | 25.34 | 36.20 | 47.05 | 2 | ||
| 8 | 63.51 | T | 7.41 | 10.59 | 13.76 | 6 |
| T++ | 22.23 | 31.76 | 41.28 | 2 | ||
| 9 | 52.71 | T | 6.15 | 8.79 | 11.42 | 6 |
| T++ | 18.45 | 26.36 | 34.26 | 2 | ||
| T:降水间隔期为5d Precipitation interval 5 days;T++:降水间隔期为15d Precipitation interval 15 days;W:自然降水Natural precipitation;W-:降水量减少30% Precipitation reduce by 30%;W+:降水量增加30% Precipitation increase by 30% | ||||||
实验采用双因素交叉试验设计开展野外小区降水控制试验, 设置3个降水量梯度和2个降水间隔期梯度, 共设置W-T、WT、W+T、W-T++、WT++、W+T++6个降水处理, 每组处理设置3个重复, 共计18个试验小区。每个小区设置为5m×5m的样方, 样方四周埋设80cm深的PVC隔水板, 确保相邻小区土壤水文之间的独立性。
1.3 地面调查和指标测量 1.3.1 植物群落调查与植物多样性指数计算2022年8月下旬, 本研究采用常规群落调查方法, 对每个5m×5m样方的物种、数量、高度和盖度进行调查。通过计算各物种的相对密度、相对盖度、相对频率计算出物种重要值, 公式如下:
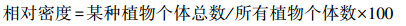
|
(1) |
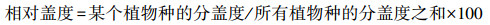
|
(2) |
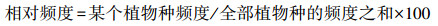
|
(3) |
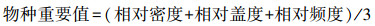
|
(4) |
本研究物种多样性的测定采用广泛应用的α多样性指数, 即Margalef丰富度指数(Ma), Pielou均匀度指数(E), Simpson优势度指数(D)和Shannon-Wiener多样性指数(H)来衡量物种多样性, 各指数计算公式如下:

|
(5) |

|
(6) |

|
(7) |

|
(8) |
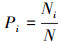
|
(9) |
式中, S为样方内物种数, N为样方中物种个体数, Ni是物种i的个体数, Pi为第i个种的多度比例。
1.3.2 土壤样品采集及理化指标测定本研究区植物根系生物量大部分集中在0—60cm土层内, 生长几乎不依赖地下水, 故试验仅对60cm深度以上的土壤进行取样[22]。2022年8月底, 从每个实验小区随机选择3个土壤采样点。从每个采样点取0—10cm、10—20cm、20—30cm、30—40cm、40—50cm、50—60cm土层的土壤, 通过烘干法测定土壤含水量(SWC)。从0—20cm土层取土样, 风干通过2mm的筛子, 进而测定土壤有机碳、全氮、全磷含量。
1.3.3 地上生产力的测定本研究采用非破坏法测定黑沙蒿地上生产力(Aboveground net primary production, ANPP)。于8月测定黑沙蒿的高度(H)、最大冠幅宽度(CW1)和最小冠幅宽度(CW2)的计数和测量, 进而计算灌木的个体盖度(C)和体积(V)分别为:
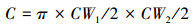
|
(10) |
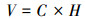
|
(11) |
根据区域已经建立起的黑沙蒿异速生长模型ANPP=193.433×V0.906计算黑沙蒿的ANPP(g/m2)[23]。
1.4 数据处理采用双因素方差分析方法, 用SPSS 26.0软件分析降水量与降水间隔以及两者之间的交互作用对植物群落组成、植物群落多样性的影响, 采用最小显著差异法(LSD)进行多重比较。用Canoco 5.0对多样性和生产力-土壤因子数据进行RDA排序, 利用Origin 2022软件绘图。
2 结果与分析 2.1 降水处理对黑沙蒿群落组成和结构的影响黑沙蒿群落主要植物种及其重要值如表 2所示, 所有降水处理下均有分布的植物种有黑沙蒿、赖草和狗娃花。WT、W+T试验小区出现植物都为11种;W-T++试验小区出现植物7种, 数量最少;WT++、W+T++试验小区分别出现植物12、14种, 物种数最多, 由此可见, 降水量增加将显著增加物种种数, 降水量减少的同时降水间隔期延长将显著影响物种存活数量。
| 功能群类型 Functional groups |
物种名 Species |
重要值Important values | |||||
| W-T | W-T++ | WT | WT++ | W+T | W+T++ | ||
| 灌木 Shrub |
黑沙蒿 Artemisia ordosica |
46.00 | 58.92 | 42.08 | 50.96 | 41.27 | 49.64 |
| 胡枝子 Lespedeza bicolor |
1.47 | - | - | 4.8 | 1.51 | 3.6 | |
| 地稍瓜 Cynanchum thesioides |
1.33 | - | 1.16 | - | 1.24 | 3.42 | |
| 多年生禾草 Perennial grass |
隐子草 Cleistogenes serotina |
8.93 | - | 6.81 | 6.86 | 3.17 | 6.98 |
| 白草 Pennisetum flaccidum |
2.96 | 18.65 | 17.43 | 6.53 | 5.29 | - | |
| 赖草 Leymus secalinus |
22.64 | 1.83 | 10.7 | 5.22 | 29.95 | 8.51 | |
| 狗尾草 Setaria viridis |
4.38 | - | 8.4 | 1.91 | 5.94 | 1.33 | |
| 针茅 Stipa capillata |
- | 1.94 | - | - | - | - | |
| 多年生杂类草 Perennial forbs |
狗娃花 Aster hispidus |
7.57 | 13.55 | 7.03 | 11.31 | 7.64 | 10.84 |
| 甘草 Glycyrrhiza uralensis |
- | - | 2.52 | 3.69 | - | - | |
| 角蒿 Incarvillea sinensis |
- | - | 1.16 | - | 1.27 | - | |
| 草木樨状黄芪 Astragalus melilotoides |
- | - | - | 2.28 | - | - | |
| 牛心朴子 Cynanchum hancockianum |
1.48 | 2.09 | - | 1.77 | - | 1.89 | |
| 披针叶野决明 Thermopsis lanceolata |
- | - | - | 2.99 | - | 2.92 | |
| 二色棘豆 Oxytropis bicolor |
- | 3.02 | 1.54 | - | 1.33 | ||
| 细叶苦荬菜 Ixeris gracilis |
- | - | 1.68 | 1.47 | 2.63 | ||
| 一年生草本 Annuals |
刺藜 Teloxys aristata |
3.24 | - | 1.17 | - | 1.24 | 3.91 |
| 虫实 Corispermum hyssopifolium |
- | - | - | - | - | 1.29 | |
| 灰绿藜 Oxybasis glauca |
- | - | - | - | - | 1.71 | |
| 数据以平均值表示, “-”表示该物种在群落中的重要值小于1或者不存在, W-T、WT、W+T分别表示降水间隔5d降水减少30%、降水间隔5d自然降水、降水间隔5d降水增加30%; W-T++、WT++、W+T++表示降水间隔15d降水减少30%、降水间隔15d自然降水、降水间隔15d降水增加30% | |||||||
不同降水处理下, 黑沙蒿均为群落优势种, 其重要值显著高于其他物种。黑沙蒿重要值高低受降水处理的影响, 在相同降水间隔期处理下, 降水量增加其重要值降低;在相同降水量情境下, 降水间隔期延长将增大黑沙蒿重要值。
按照功能群对物种进行分类(图 1), 在降水量一致的情况下降水间隔期的延长显著提高了灌木和杂草的重要值, 禾草的重要值显著降低。在降水时间间隔为15d的情况下, 增加30%的降水, 一年生草本植物在群落中大量出现。

|
| 图 1 降水格局变化对不同功能群植物重要值百分比 Fig. 1 Changes in precipitation patterns on percentage plant importance values for different functional groups |
不同降水格局下, 黑沙蒿群落多样性发生了显著变化。降水量改变对黑沙蒿群落Simpson优势度指数、Shannon-Wiener多样性指数、Pielou均匀度指数存在显著影响(P < 0.05)。当降水间隔期为15d时, W-、W、W+的Simpson优势度指数表现为W+ < W < W-;当降雨间隔为5d时, 黑沙蒿群落Margalef丰富度指数、Simpson优势度指数、Shannon-Wiener多样性指数、Pielou均匀度指数在不同降水量处理下均无显著影响。降水间隔期改变对Simpson优势度指数、Shannon-Wiener多样性指数存在显著影响(P < 0.05)。当降水量为W-水平时, 黑沙蒿群落Shannon-Wiener多样性指数表现为T++ < T;当降水量为W-水平时, 黑沙蒿群落Simpson优势度指数表现为T++>T。当降水间隔期为15d时, W-、W、W+的Shannon-Wiener多样性指数表现为W- < W+ < W。由此可知, 当降水量减少30%, 延长降水间隔为15d时, 有助于黑沙蒿植物群落优势物种的生存;当降水量减少30%, 降水间隔为5d时, 有助于提高黑沙蒿植物群落的物种多样性, 而延长降水间隔则降低了黑沙蒿植物群落的物种多样性。
不同降水量处理间黑沙蒿ANPP具有显著差异(图 2), 无论降水间隔为5d或15d, 黑沙蒿ANPP均表现为W- < W < W+。降水间隔期延长对ANPP的影响在降水量减少的实验小区存在显著性差异(P < 0.05), 当降水量为W-水平时, 降水间隔的延长显著增加了黑沙蒿ANPP, 在降水量增加的实验小区降水间隔期对ANPP无显著影响。

|
| 图 2 降水格局变化对黑沙蒿群落多样性的影响(平均值±标准误差) Fig. 2 Effects of changing precipitation patterns on community diversity of Artemisia ordosica (mean ± standard error) 相同降水间隔期下不同大写字母表示3个降水量处理之间差异显著(P < 0.05);相同降水量下不同小写字母表示2个降水间隔期处理之间差异显著(P < 0.05), T降水间隔期为5 d;T++降水间隔期为15 d;W自然降水;W-降水量减少30%;W+降水量增加30% |

|
| 图 3 降水格局变化对黑沙蒿地上生产力的影响 Fig. 3 Anatomical images of the stem of Artemisia ordosica with different precipitation treatments 相同降水间隔期下不同大写字母表示3个降水量处理之间差异显著(P < 0.05);相同降水量下不同小写字母表示2个降水间隔期处理之间差异显著(P < 0.05), T降水间隔期为5d;T++降水间隔期为15d;W自然降水;W-降水量减少30%;W+降水量增加30% |
随着土层深度的增加, 土壤含水率呈增加的趋势, 40—60cm深层土壤的含水率最高(表 3)。降水间隔的延长和降水量的增加, 显著增加了40—60cm深层土壤的含水率, 对0—20cm浅层土壤的含水率影响不显著(表 3)。
| 降水处理 Precipitation treatment |
土层Soil layer/cm | |||||
| 0—10 | 10—20 | 20—30 | 30—40 | 40—50 | 50—60 | |
| W-T | 0.016±0.005Ca | 0.019±0.002BCa | 0.021±0.002ABCa | 0.027±0.001ABa | 0.029±0.001Ab | 0.027±0.002ABc |
| W-T++ | 0.014±0.002Ca | 0.022±0.006BCa | 0.028±0.008ABCa | 0.035±0.006ABCa | 0.044±0.008ABab | 0.052±0.004Aa |
| WT | 0.022±0.007Aa | 0.026±0.005Aa | 0.029±0.005Aa | 0.032±0.001Aa | 0.034±0.002Aab | 0.032±0.004Abc |
| WT++ | 0.015±0.001Ca | 0.023±0.004BCa | 0.026±0.004BCa | 0.039±0.002ABa | 0.048±0.003Aab | 0.045±0.005Aab |
| W+T | 0.021±0.006Ca | 0.025±0.004BCa | 0.025±0.003BCa | 0.031±0.003ABCa | 0.039±0.003Aab | 0.036±0.003ABbc |
| W+T++ | 0.015±0.001Ba | 0.025±0.007ABa | 0.033±0.012ABa | 0.042±0.0105ABa | 0.055±0.015Aa | 0.053±0.006Aa |
| 表中数据用平均值±标准误差(n=3)表示; 同行不同大写字母表示在同一降水格局处理下不同土层具有显著性差异(P < 0.05); 同列不同小写字母代表示同一土层不同降水格局处理间具有显著性差异(P < 0.05), T降水间隔期为5d;T++降水间隔期为15d;W自然降水;W-降水量减少30%;W+降水量增加30% | ||||||
Margalef丰富度指数、Simpson优势度指数、生产力与SWC呈显著正相关关系, Margalef丰富度指数、Simpson优势度指数与其他土壤环境因子呈显著负相关关系。Pielou均匀度指数和Shannon-Wiener多样性指数与C∶N、C∶P、SOC、TN、TP、N∶P呈正相关关系(图 4)。

|
| 图 4 降水格局变化下土壤因子与植物群落多样性的RDA分析 Fig. 4 RDA analysis of soil factors and plant community diversity under changing precipitation patterns SWC、SOC、TN、TP、C∶N、C∶P、N∶P分别代表土壤含水量、土壤有机碳、全氮、全磷、C∶N、C∶P、N∶P、Margalef、Shannon-Wiener、Simpson和Pielou分别代表丰富度指数、Shannon-Wiener多样性指数、优势度指数和均匀度指数 |
降水间隔延长与单次降水量增多的极端降水事件在中国西北干旱地区将会更频繁的出现[24]。荒漠生态系统对降水极为敏感, 降水变化会显著改变黑沙蒿群落的物种组成。在本实验中减少降水30%、降水间隔延长为15d时物种数较对照组(WT)减少了4种, 究其原因是由于降水减少、降水事件间干旱周期延长, 导致土壤表层水分减少, 土壤表层干旱化, 使其难以维持耐旱性较弱的浅根植物生长[16, 25—26], 因此导致了浅根系草本植物的死亡并退出群落。在本研究中, 表现为一年生草本(虫实、灰绿藜)以及多年生杂草(甘草、角蒿、草本樨状黄芪、披针叶野决明、细叶苦荬菜)在减少降水量时均没有在群落中出现。
增加降水30%、延长降水间隔为15d时物种数较对照组增加了3种, 本研究认为这是由于降水量增加和降水集中分配使得单次降水量增大, 有效的促进了降水入渗、增加了40cm以下深层土壤的含水量。这种降水情境有助于增加40cm以下土壤层水分的有效性, 为40—60cm深度分布的深根灌木植物的生长提供了良好的条件[27—29], 因此优势种黑沙蒿重要值增加。与此同时, 胡枝子、地稍瓜等半灌木在增加降水量、延长降水间隔处理时物种重要值达到最大, 也充分证明了年内降水集中分配对群落物种组成数量的积极影响[30]。
随降水量的增加, 黑沙蒿群落的Margalef丰富度指数、Shannon-Wiener多样性指数、Pielou均匀度指数均呈现增加的趋势, 而Simpson优势度指数却呈现出降低的趋势。其原因是降水量的增加可以为一年生草本植物的生长提供更为有利的生长条件, 从而促进群落中物种丰富度的增加, 该结果在RDA分析结果中也可以得到佐证(图 4)。与此同时, 草本植物的增多与灌木争夺有限的空间资源, 表现为群落中的黑沙蒿优势度降低(表 2), 即降水的改变可以通过改变物种间的相互作用间接影响植物群落物种多样性[31—32]。不同降水处理下土壤养分对物种多样性指数的影响也在本研究中得到检验, 然而经由养分驱动的群落物种组成变化的机制还需进一步探究。
3.2 降水处理对黑沙蒿生产力的影响干旱、半干旱地区植物的生产力主要受水分的影响[33]。本研究的结果表明, 降水间隔无论5d还是15d, 增加降水都会提高黑沙蒿的生产力。这种降水量与生产力的正相关关系与以往诸多研究结果相一致[34—36]。但值得注意的是, 本研究结果表明, 即使年总降水量保持恒定, 年内降水分配的改变仍然可以影响群落植物的生产力, 在低降水量处理组且降水间隔期为15d时, 黑沙蒿的生产力显著高于间隔期为5d的处理, 说明降水间隔的延长对黑沙蒿的生长具有促进作用, 这与降水年内时间变化导致的土壤水分再分配紧密相关[37], 间隔期延长导致单次降水强度增大, 使深层土壤水分的有效性增加, 有利于深根性的黑沙蒿水分吸收, 这一土壤含水率对生产力的影响从RDA分析图(图 4)二者夹角的显著相关关系中也可以得到证实。
然而, 降水年内时间分配对生产力的影响并非始终存在, 在本研究中降水年内分配对群落的影响受年总降水量的控制, 仅在低降水量时表现出显著性, 由此可以推测, 干旱年的降水分配对荒漠植物群落的影响更为值得关注。鉴于降水量增加, 降水频率减少的降水趋势, 本研究推测未来本区域内黑沙蒿群落中深根灌木植物比重将增加, 而浅根系的草本植物将难以生存, 植物的群落结构将会发生显著变化。
4 结论黑沙蒿灌草群落多样性和生产力受到年际降水量和年内降水时间分配的双重影响。降水量的增加提高了土壤各层次的含水率, 促进了多年生杂草的生长, 提高了群落的物种丰富度和多样性;相反, 降水量的减少降低了群落多样性和均匀度。降水年内分配对群落的影响受年总降水量的控制, 仅在低降水量年份, 降水间隔延长、集中强降水事件通过增加40—60cm的土壤含水率, 促进了深根性黑沙蒿的重要值和生产力, 使得群落整体优势度上升、多样性下降。
| [1] |
薛媛, 薛晓萍. 极端降水与干旱同步频发的研究进展. 海洋气象学报, 2022, 42(1): 61-73. |
| [2] |
Climate change 2013. The Physical Science Basis. Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: University Press, 2014.
|
| [3] |
符淙斌, 马柱国. 全球干旱/半干旱区年代尺度干湿变化研究的进展及思考. 大气科学学报, 2023, 46(4): 481-490. |
| [4] |
王明明, 刘新平, 李玉霖, 车力木格, 罗永清, 孙珊珊, 魏静. 不同植被盖度沙质草地生长季土壤水分动态. 中国沙漠, 2019, 39(5): 54-61. |
| [5] |
Yang H J, Wu M Y, Liu W X, Zhang Z, Zhang N L, Wan S Q. Community structure and composition in response to climate change in a temperate steppe. Global Change Biology, 2011, 17(1): 452-465. DOI:10.1111/j.1365-2486.2010.02253.x |
| [6] |
李长斌, 彭云峰, 赵殿智, 宁祎, 周国英. 降水变化和氮素添加对青藏高原高寒草原群落结构和物种多样性的影响. 水土保持研究, 2016, 23(6): 185-191. |
| [7] |
常学礼, 赵爱芬, 李胜功. 科尔沁沙地固定沙丘植被物种多样性对降水变化的响应. 植物生态学报, 2000, 24(2): 147-151. DOI:10.3321/j.issn:1005-264X.2000.02.004 |
| [8] |
汪殿蓓, 暨淑仪, 陈飞鹏. 植物群落物种多样性研究综述. 生态学杂志, 2001, 20(4): 55-60. |
| [9] |
张蕊, 赵学勇, 王少昆, 左小安, 王瑞雄. 极端干旱对荒漠草原群落物种多样性和地上生物量碳氮的影响. 生态环境学报, 2019, 28(4): 715-722. |
| [10] |
马文静, 张庆, 牛建明, 康萨如拉, 刘朋涛, 何欣, 杨艳, 张艳楠, 邬建国. 物种多样性和功能群多样性与生态系统生产力的关系——以内蒙古短花针茅草原为例. 植物生态学报, 2013, 37(7): 620-630. |
| [11] |
Liu D J, Zhang C, Ogaya R, Fernández-Martínez M, Pugh T A, Peñuelas J. Increasing climatic sensitivity of global grassland vegetation biomass and species diversity correlates with water availability. New Phytologist, 2021, 230(5): 1761-1771. DOI:10.1111/nph.17269 |
| [12] |
韩彬, 樊江文, 钟华平. 内蒙古草地样带植物群落生物量的梯度研究. 植物生态学报, 2006, 30(4): 553-562. |
| [13] |
Knapp A K, Beier C, Briske D D, Classen A T, Luo Y, Reichstein M, Smith M D, Smith S D, Bell J E, Fay P A, Heisler J L, Leavitt S W, Sherry R, Smith B, Weng E. Consequences of more extreme precipitation regimes for terrestrial ecosystems. BioScience, 2008, 58(9): 811-821. |
| [14] |
董亚威, 郭子月, 特尼乌, 邱璧迎, 张志铭, 雒文涛, 钱建强. 内蒙古荒漠草原地下芽库对两种模拟极端干旱的响应. 内蒙古大学学报(自然科学版), 2021, 52(6): 601-608. |
| [15] |
朱国栋, 郭娜, 韩勇军, 吕广一, 王占义, 王成杰. 极端干旱对内蒙古荒漠草原植物群落物种多样性和土壤性质的影响. 中国草地学报, 2021, 43(3): 52-59. |
| [16] |
Gherardi L A, Sala O E. Enhanced precipitation variability decreases grass- and increases shrub-productivity. Proceedings of the National Academy of Sciences, 2015, 112(41): 12735-12740. |
| [17] |
朱雅娟, 吴波, 卢琦. 干旱区对降水变化响应的研究进展. 林业科学研究, 2012, 25(1): 100-106. |
| [18] |
He Y, Yu M, Ding G, Wang C, Zhang F. Precipitation amount and event intervals interact to change plant diversity during dry years in a desert shrubland. Ecological Indicators, 2022, 145: 109701. |
| [19] |
张彬, 朱建军, 刘华民, 潘庆民. 极端降水和极端干旱事件对草原生态系统的影响. 植物生态学报, 2014, 38(9): 1008-1018. |
| [20] |
张腊梅, 刘新平, 赵学勇, 张铜会, 岳祥飞, 云建英. 科尔沁固定沙地植被特征对降雨变化的响应. 生态学报, 2014, 34(10): 2737-2745. |
| [21] |
张富崇, 于明含, 何莹莹, 王春媛, 丁国栋, 唐颖. 毛乌素沙地降水格局变化对油蒿木质部解剖特征的影响. 生态学报, 2023, 43(16): 6661-6669. |
| [22] |
梁香寒. 毛乌素沙地不同固定程度油蒿群落土壤水分变化过程及影响因素研究[D]. 北京: 北京林业大学, 2020.
|
| [23] |
She W W, Zhang Y Q, Qin S G, Wu B, Bai Y X. Increased precipitation and nitrogen alter shrub architecture in a desert shrubland: implications for primary production. Frontiers in Plant Science, 2016, 7: 1908. |
| [24] |
Hoover D L, Hajek O L, Smith M D, Wilkins K, Slette I J, Knapp A K. Compound hydroclimatic extremes in a semi-arid grassland: drought, deluge, and the carbon cycle. Global Change Biology, 2022, 28(8): 2611-2621. |
| [25] |
孙岩, 何明珠, 王立. 降水控制对荒漠植物群落物种多样性和生物量的影响. 生态学报, 2018, 38(7): 2425-2433. |
| [26] |
张加涛. 干旱与氮沉降对内蒙古典型草原群落结构及生产力的影响[D]. 呼和浩特: 内蒙古大学, 2021.
|
| [27] |
Cheng X, An S, Li B, Chen J, Lin G, Liu Y, Luo Y, Liu S. Summer rain pulse size and rainwater uptake by three dominant desert plants in a desertified grassland ecosystem in northwestern China. Plant Ecology, 2006, 184: 1-12. |
| [28] |
Suttle K B, Thomsen M A, Power M E. Species interactions reverse grassland responses to changing climate. Science, 2007, 315(5812): 640-642. |
| [29] |
Báez S, Collins S L, Pockman W T, Johnson J E, Small E E. Effects of experimental rainfall manipulations on Chihuahuan Desert grassland and shrubland plant communities. Oecologia, 2013, 172: 1117-1127. |
| [30] |
Chen Y X, Vogel A, Wagg C, Xu T Y, Iturrate-Garcia M, Scherer-Lorenzen M, Weigelt A, Eisenhauer N, Schmid B. Drought-exposure history increases complementarity between plant species in response to a subsequent drought. Nature Communications, 2022, 13: 3217. |
| [31] |
Klanderud K, Totland ∅. Simulated climate change altered dominance hierarchies and diversity of an alpine biodiversity hotspot. Ecology, 2005, 86(8): 2047-2054. |
| [32] |
Zhong M X, Song J, Zhou Z X, Ru J Y, Zheng M M, Li Y, Hui D F, Wan S Q. Asymmetric responses of plant community structure and composition to precipitation variabilities in a semi-arid steppe. Oecologia, 2019, 191: 697-708. |
| [33] |
赵学勇, 刘良旭, 王玮, 常学礼, 曲浩, 毛伟, 贾昆峰. 降水波动对荒漠草原生产力的影响. 中国沙漠, 2014, 34(6): 1486-1495. |
| [34] |
孙羽, 张涛, 田长彦, 李晓林, 冯固. 增加降水对荒漠短命植物当年牧草生长及群落结构的影响. 生态学报, 2009, 29(4): 1859-1868. |
| [35] |
毕旭, 徐向超, 张立新. 中亚高海拔梯度山盆系统草地净初级生产力时空演变格局及其驱动机制分析. 生态环境学报, 2020, 29(5): 876-888. |
| [36] |
邓强, 刘迪, 时新荣, 袁志友. 黄土高原刺槐人工林林下植物多样性对模拟降水变化的响应. 水土保持研究, 2021, 28(1): 1-10. |
| [37] |
魏兰兰. 黄土高原浅层土壤水分对降水的响应机制研究——以董志塬为例[D]. 兰州: 兰州大学, 2022.
|