 2024, Vol. 44
2024, Vol. 44文章信息
- 程铭昊, 李劲, 张溢, 丛微, 张宇, 蒋军, 黄天鹏, 梅玉娇, 李书航, 张于光
- CHENG Minghao, LI Jin, ZHANG Yi, CONG Wei, ZHANG Yu, JIANG Jun, HUANG Tianpeng, MEI Yujiao, LI Shuhang, ZHANG Yuguang
- 基于DNA条形码技术的神农架国家公园川金丝猴食性研究
- Golden snub-nosed monkey diet research in Shennongjia National Park based on DNA barcoding technology
- 生态学报. 2024, 44(15): 6424-6435
- Acta Ecologica Sinica. 2024, 44(15): 6424-6435
- http://dx.doi.org/10.20103/j.stxb.202401090082
-
文章历史
- 收稿日期: 2024-02-09
- 网络出版日期: 2024-05-24





2. 神农架国家公园管理局, 神农架 442421
2. Shennongjia National Park Administration, Shennongjia 442421, China
食性分析是动物生态学研究的重要组成部分, 有助于揭示动物的饮食结构、觅食策略和栖息地选择偏好, 以及评估物种生存状况和种间关系等[1]。食性研究对于濒危野生动物的保护管理具有了解其生存威胁、制定更有效的保护策略、对同域分布物种合理划分保护区、预测其种群数量等重要意义[2—3]。
传统的食性研究方法主要包括通过观察取食行为的野外扫描取样法、笼养饲喂法、胃内容物法、粪便显微分析法等[3], 其中扫描取样法需要长期对野生动物进行跟踪及采集植物对野生动物进行投喂, 工作效率低且不能得到较完整的食物组成[4];依赖于显微观察的显微分析法主要是观察粪便中残留的食物残渣以鉴别食物种类, 由于动物消化能力的差异及研究者鉴别水平的差异无法较为准确的得到完整的食物组成[5]。而新兴的DNA宏条形码技术利用动物粪便DNA的高通量测序结果进行物种鉴定, 相较于传统的研究方法体现出鉴别效率高、结果准确率高等优势[6]。该技术主要通过序列相似度进行物种鉴别, 这对研究者的植物识别能力要求较低且不受主观因素的干扰[7], 但物种鉴别的准确率很大程度上受参考数据库的影响, 不完整的参考数据库会使得DNA条形码的鉴别能力大幅下降[8]。
DNA宏条形码技术已被广泛应用于野生灵长类动物的食物组成、觅食策略、种间关系分析和物种保护等方面[9—14]。研究发现灵长类动物不单以植物为食, 还会取食节肢动物、水生生物甚至小型哺乳动物, 其中鞍背柽柳猴(Saguinus fuscicollis)会取食蜘蛛类、甲虫类和蚁类等节肢动物以补充体内所需的蛋白质和能量[9], 研究发现日本猕猴(Macaca fuscata)捕食蟹类、褐鳟(Salmo trutta)等其他水生动物[10], 而在非洲的倭黑猩猩(Pan paniscus)的粪便中发现了坦桑尼亚小羚羊(Cephalophus spadix)等哺乳动物的DNA, 之前野外观察的结果也能证实其取食哺乳动物[11]。基于DNA宏条形码技术的食性研究发现, 白脸卷尾猴(Cebus capucinus)不同年龄或不同性别的个体通过取食不同种类的昆虫规避种内竞争[12]。马达加斯加同域分布的10种灵长类动物通过取食树冠层的不同的植物资源和节肢动物, 以实现资源的合理分配及种间共存[13]。通过研究云南怒江金丝猴(Rhinopithecus strykeri)的食性, 对保护区内怒江金丝猴喜食的植物进行重点保护, 以促进其野外种群恢复[14]。
川金丝猴(R. roxellana)是我国特有的灵长类动物, 属于国家一级重点保护野生动物[15], 《中国脊椎动物红色名录》易危(VU)物种[16]。川金丝猴分化为三个亚种, 其中指名亚种(R. roxellana roxellana)分布于四川的岷山山系和邛崃山系, 秦岭亚种(R. roxellana qinlingensis)分布于秦岭山系, 而神农架国家公园则是川金丝猴湖北亚种(R. roxellana hubeiensis)的唯一分布区[17]。目前我国野生川金丝猴总数约为23000只, 其中湖北亚种的野外种群数量最少, 仅有1500余只, 在三个亚种之中面临最大的灭绝风险[17]。神农架金丝猴是神农架国家公园的旗舰物种和指示性物种, 在维系森林群落结构与生态功能的完整性, 动植物协同进化上发挥着极其重要的作用, 保护和研究意义重大[18]。目前对川金丝猴的食性研究已有较多报道, 但研究方法主要为传统的野外观察法及显微分析法等, DNA宏条形码技术尚未应用于川金丝猴的食性研究 [19—21]。
本研究以神农架国家公园的大龙潭野外川金丝猴补食群为研究对象, 通过采集不同季节的川金丝猴新鲜粪便, 利用DNA宏条形码技术分析神农架川金丝猴的食物组成及多样性的季节差异, 为川金丝猴的饲喂繁育和保护管理提供参考价值。
1 研究方法 1.1 研究对象神农架国家公园的大龙潭野外川金丝猴补食群数量约为80只, 该猴群主要活动在大龙潭区域内的龙潭湾、三岔沟、龙林沟、观音洞等地(31.4903°—31.4916°N, 110.2974°—110.2988°E), 平均海拔2243m。该区域的植被类型为巴山冷杉(Abies fargesii)、华山松(Pinus armandii)和红桦(Betula albosinensis)的针阔混交林[22]。为保证该种群的长期生存和开展川金丝猴的野外研究, 在食物短缺的冬季对川金丝猴进行人工补食, 补食食物包括苹果、红薯和胡萝卜等[23]。
1.2 样品采集于2023年冬季(2月)、春季(5月)和秋季(10月)分别采集了川金丝猴补食群的新鲜粪便样品。采集时佩戴一次性无菌手套, 将疑似金丝猴的粪便样品装入含干燥剂的收集管中, 并在收集管上标注采集时间和样品编号。采集时避免在同一地点多次采集, 同时收集大小、颜色不一致的粪便以确保收集到多只个体的粪便。待粪便样品干燥后放入冰箱-40 ℃保存, 野外工作结束后带回实验室处理[24]。共采集疑似川金丝猴粪便样品133份, 其中冬季50份, 春季42份, 秋季41份。
1.3 样品DNA提取及PCR扩增和测序使用Qiagen Stool DNA试剂盒(Qiagen, Valencia, California, USA)提取样品中的DNA。提取完成后使用分光光度计对DNA溶液进行浓度和纯度的测定。
选用Riaz等人设计的脊椎动物通用引物12SV5(F: TAGAACAGGCTCCTCTAG;R: TTAGATACC CCACTATGC)对样品DNA进行扩增用于鉴别粪便的物种来源[25]。PCR反应程序为95 ℃预变性3 min;95 ℃变性30 s, 60 ℃退火30 s, 72 ℃延伸30 s, 循环45次;72 ℃延伸5 min。使用植物叶绿体基因trnL片段(g:GGGCAATCCTGAGCCAA;h:CATTGAGTCTCTGCACCTATC)对粪便样品中的植物DNA进行扩增[26], PCR反应程序为95 ℃预变性10 min;95 ℃变性30 s, 55 ℃退火30 s, 72 ℃延伸30 s, 循环35次; 72 ℃延伸2 min。PCR体系总体积为20 μL:DNA模板(10—50 ng/μL)2 μL;2×Taq Master Mix 10 μL;正反向引物(10μmol/L)各0.5 μL;ddH2O 7 μL。扩增时在正向和反向的5′端各添加8 bp条形码序列, 以区分不同的样品。PCR扩增产物于Illumina HiSeq 2500测序平台进行高通量测序, PCR扩增及测序由广东美格基因科技有限公司完成。
1.4 测序数据处理使用Vsearch v2.22.1对测序结果进行处理, 首先对每个样品的正反向序列进行裁剪、拼接, 将相同季节的样品测序结果合并, 并对来自不同样品的序列添加标签。之后过滤长度过长或过短的序列及累积碱基错误率高于1%的低质量序列, 然后去除重复序列及单一读数序列, 再使用uchime2功能去除包含嵌合体的序列[27]。去除嵌合体序列后以97%的相似度阈值对结果进行聚类, 剔除单一读数序列后将每条序列记为一个分子可操作分类单元(molecular operational taxonomic unit, MOTU)。
1.5 参考数据库的建立及物种注释对每个各样品的12S条形码测序结果与NCBI(national center for biotechnology information)的GeneBank(https://www.ncbi.nlm.nih.gov)中川金丝猴的12S序列进行比对, 进行粪便的物种鉴定。舍弃其他物种的粪便样品, 保留川金丝猴的粪便样品进行食源植物的物种注释。
综合神农架国家公园植物名录及已有神农架川金丝猴食源植物研究结果整理出包括1309种植物的神农架国家公园川金丝猴食源植物参考名录[28—29], 从GeneBank和EMBL(european molecular biology laboratory, https://www.ebi.ac.uk)等数据库共检索到参考名录中1185种植物的trnL序列信息, 使用BLAST+(v2.15.0)软件将检索到的植物序列建立为DNA条形码的参考数据库。
将trnL条形码测序结果与建立的植物数据库进行相似度比对, 物种注释的原则为:(1)与单一物种序列的相似度≥98%, 记为该物种(2)与多个物种序列的相似度≥98%时, 记为能够涵盖所有物种≥98%的最低分类单元。(3)比对结果中, 最高相似度<98%且≥95%时, 相似度最高序列仅有1条, 且与得分次之的序列一致度差距≥1%时, 记为相似度最高序列的上一级分类单元。最高一致度对应有多个物种时, 则记为能够涵盖所有最高相似度的物种最低分类单元。(4)比对结果中, 最高相似度 < 95%时, 认为无法判定该序列的分类单元, 不计入食性结果[30]。
1.6 食性分析将同一季节的川金丝猴粪便样品测序结果合并处理得到该组的食性结果。为判断取样量是否充分, 使用R软件vegan包绘制物种累积曲线图, 同时计算不同组别的Simpson指数D、Shannon指数H、Pielou指数J以比较川金丝猴的食物多样性差异[31]。具体计算方法如下:
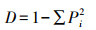
|
(1) |
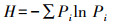
|
(2) |
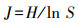
|
(3) |
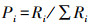
|
(4) |
式中, S为MOTU总数;Pi为某MOTU的相对序列丰度(relative read abundance, RRA/%);Ri为某组中第i个MOTU的序列读数。
2 结果 2.1 物种鉴定采集的133份粪便样品均扩增出目的条带并进行物种鉴定, 鉴定结果显示127份粪便为川金丝猴的粪便, 其中冬季50份, 春季40份, 冬季37份。
2.2 神农架川金丝猴食性分析对上述127份粪便样品进行trnL区域的PCR扩增测序, 分别对3个季节的MOTU进行物种注释, 共得到127个物种类别, 其中79个鉴定到种水平, 占比为62%;38个仅鉴定到属水平, 占比为30%;10个仅鉴定到科水平, 占比为8%。不同季节的MOTU物种注释结果见表 1。
| 分类水平 Classification level |
冬季 Winter |
春季 Spring |
秋季 Autumn |
分类水平 Classification Level |
冬季 Winter |
春季 Spring |
秋季 Autumn |
|
| 种Species | 25 | 47 | 42 | 科Family | 6 | 4 | 5 | |
| 属Genus | 25 | 25 | 23 | MOTU总数Total number of MOTU | 56 | 76 | 70 | |
| MOTU: 分子可操作分类单元Molecular operational taxonomic unit | ||||||||
物种累积曲线显示样品数量大于30时, 鉴定得到的物种数趋于稳定, 因此说明本研究的样品数量足以表征金丝猴粪便中的植物种类(图 1)[32]。

|
| 图 1 不同季节样品的物种累积曲线图 Fig. 1 Species accumulation curves of samples from different seasons |
在科水平上, 食物种类涵盖了62个科, 其中相对序列丰度较高的5个优势科为:蔷薇科(Rosaceae, 37.70%)、猕猴桃科(Actinidiaceae, 11.33%)、茄科(Solanaceae, 8.62%)、无患子科(Sapindaceae, 4.88%)和山茱萸科(Cornaceae, 4.45%)。
在属水平上, 食物种类涵盖了110个属, 其中相对序列丰度较高的5个优势属为:花楸属(Sorbus, 25.27%)、地海椒属(Archiphysalis, 8.73%)、藤山柳属(Clematoclethra, 8.63%)、委陵菜属(Potentilla, 8.13%)和槭属(Acer, 4.94%)。
在种水平上, 共鉴定出79种食物。其中相对序列丰度较高的5个优势种为:地海椒(18.20%)、藤山柳(Clematoclethra scandens)(17.99%)、委陵菜(Potentilla chinensis)(16.95%)、菱叶茴芹(Pimpinella rhomboidea)(8.89%)和勾儿茶(Berchemia sinica)(7.51%)。
神农架川金丝猴不同季节的食物组成详见附表 1。
| 科 Family |
属 Genus |
种 Species |
冬季 Winter |
春季 Spring |
秋季 Autumn |
| 松科Pinaceae | 冷杉属Abies | 巴山冷杉Abies fargesii | 有 | 有 | 无 |
| 毛茛科Ranunculaceae | 乌头属Aconitum | 花莛乌头Aconitum scaposum | 无 | 有 | 有 |
| 毛茛科Ranunculaceae | 类叶升麻属Actaea | 升麻Actaea cimicifuga | 无 | 有 | 有 |
| 禾本科Poaceae | 剪股颖属Agrostis | 小花剪股颖Agrostis micrantha | 无 | 无 | 有 |
| 伞形科Apiaceae | 峨参属Anthriscus | 峨参Anthriscus sylvestris | 无 | 有 | 无 |
| 毛茛科Ranunculaceae | 耧斗菜属Aquilegia | 无距耧斗菜Aquilegia ecalcarata | 无 | 有 | 有 |
| 茄科Solanaceae | 地海椒属Archiphysalis | 地海椒Archiphysalis sinensis | 有 | 无 | 无 |
| 禾本科Poaceae | 野古草属Arundinella | 野古草Arundinella hirta | 无 | 有 | 无 |
| 马兜铃科Aristolochiaceae | 细辛属Asarum | 单叶细辛Asarum himalaicum | 无 | 无 | 有 |
| 虎耳草科Saxifragaceae | 落新妇属Astilbe | 落新妇Astilbe chinensis | 无 | 有 | 无 |
| 小檗科Berberidaceae | 小檗属Berberis | 单花小檗Berberis candidula | 无 | 有 | 无 |
| 小檗科Berberidaceae | 小檗属Berberis | 假豪猪刺Berberis soulieana | 无 | 有 | 无 |
| 鼠李科Rhamnaceae | 勾儿茶属Berchemia | 勾儿茶Berchemia sinica | 有 | 有 | 有 |
| 桑科Moraceae | 构属Broussonetia | 构树Broussonetia papyrifera | 无 | 无 | 有 |
| 山茶科Theaceae | 山茶属Camellia | 茶树Camellia sinensis | 有 | 无 | 无 |
| 十字花科Brassicaceae | 碎米荠属Cardamine | 大叶碎米荠Cardamine macrophylla | 无 | 有 | 无 |
| 莎草科Cyperaceae | 薹草属Carex | 基花薹草Carex basiflora | 无 | 无 | 有 |
| 莎草科Cyperaceae | 薹草属Carex | 柄果薹草Carex stipitinux | 无 | 无 | 有 |
| 石竹科Caryophyllaceae | 卷耳属Cerastium | 鄂西卷耳Cerastium wilsonii | 无 | 有 | 无 |
| 连香树科Cercidiphyllaceae | 连香树属Cercidiphyllum | 连香树Cercidiphyllum japonicum | 有 | 无 | 无 |
| 杜鹃花科Ericaceae | 喜冬草属Chimaphila | 喜冬草Chimaphila japonica | 有 | 无 | 无 |
| 菊科Asteraceae | 菊属Chrysanthemum | 甘菊Chrysanthemum lavandulifolium | 无 | 无 | 有 |
| 虎耳草科Saxifragaceae | 金腰属Chrysosplenium | 舌叶金腰 Chrysosplenium glossophyllum | 有 | 有 | 有 |
| 柳叶菜科Onagraceae | 露珠草属Circaea | 高原露珠草Circaea alpina subsp. imaicola | 无 | 有 | 有 |
| 菊科Asteraceae | 蓟属Cirsium | 刺儿菜Cirsium arvense var. integrifolium | 无 | 有 | 无 |
| 芸香科Rutaceae | 柑橘属Citrus | 宜昌橙Citrus cavaleriei | 有 | 无 | 有 |
| 毛茛科Ranunculaceae | 铁线莲属Clematis | 毛蕊铁线莲Clematis lasiandra | 无 | 有 | 有 |
| 猕猴桃科Actinidiaceae | 藤山柳属Clematoclethra | 藤山柳Clematoclethra scandens | 有 | 有 | 无 |
| 唇形科Lamiaceae | 风轮菜属Clinopodium | 风轮菜Clinopodium chinense | 无 | 有 | 无 |
| 罂粟科Papaveraceae | 紫堇属Corydalis | 神农架紫堇Corydalis ternatifolia | 无 | 有 | 有 |
| 禾本科Poaceae | 鸭茅属Dactylis | 鸭茅 Dactylis glomerata | 无 | 有 | 有 |
| 蔷薇科Rosaceae | 金露梅属Dasiphora | 金露梅Dasiphora fruticosa | 无 | 无 | 有 |
| 禾本科Poaceae | 野青茅属Deyeuxia | 野青茅Deyeuxia pyramidalis | 无 | 有 | 无 |
| 薯蓣科Dioscoreaceae | 薯蓣属Dioscorea | 穿龙薯蓣Dioscorea nipponica | 无 | 无 | 有 |
| 蔷薇科Rosaceae | 蛇莓属Duchesnea | 蛇莓Duchesnea indica | 无 | 无 | 有 |
| 柳叶菜科Onagraceae | 柳叶菜属Epilobium | 柳叶菜Epilobium hirsutum | 有 | 有 | 有 |
| 牻牛儿苗科Geraniaceae | 老鹳草属Geranium | 老鹳草Geranium wilfordii | 无 | 有 | 有 |
| 唇形科Lamiaceae | 活血丹属Glechoma | 活血丹 Glechoma longituba | 无 | 有 | 无 |
| 豆科Fabaceae | 皂荚属Gleditsia | 皂荚Gleditsia sinensis | 有 | 无 | 有 |
| 豆科Fabaceae | 大豆属Glycine | 野大豆Glycine soja | 有 | 有 | 有 |
| 青荚叶科Helwingiaceae | 青荚叶属Helwingia | 中华青荚叶Helwingia japonica | 有 | 无 | 无 |
| 金丝桃科Hypericaceae | 金丝桃属Hypericum | 元宝草Hypericum sampsonii | 无 | 有 | 无 |
| 凤仙花科Balsaminaceae | 凤仙花属Impatiens | 神农架凤仙花Impatiens shennongensis | 无 | 无 | 有 |
| 无患子科Sapindaceae | 栾属Koelreuteria | 栾树 Koelreuteria paniculata | 无 | 无 | 有 |
| 忍冬科Caprifoliaceae | 忍冬属Lonicera | 忍冬Lonicera japonica | 有 | 有 | 无 |
| 灯芯草科Juncaceae | 地杨梅属Luzula | 散序地杨梅 Luzula effusa | 无 | 有 | 无 |
| 报春花科Primulaceae | 珍珠菜属Lysimachia | 显苞过路黄Lysimachia rubiginosa | 有 | 有 | 无 |
| 天门冬科Asparagaceae | 舞鹤草属Maianthemum | 管花鹿药Maianthemum henryi | 无 | 有 | 无 |
| 禾本科Poaceae | 乱子草属Muhlenbergia | 多枝乱子草Muhlenbergia ramosa | 无 | 有 | 无 |
| 石竹科Caryophyllaceae | 齿缀草属Odontostemma | 神农架齿缀草 Odontostemma shennongjiaense |
无 | 有 | 无 |
| 酢浆草科Oxalidaceae | 酢浆草属Oxalis | 白花酢浆草Oxalis acetosella | 无 | 有 | 无 |
| 泡桐科Paulowniaceae | 泡桐属Paulownia | 毛泡桐Paulownia tomentosa | 有 | 无 | 无 |
| 景天科Crassulaceae | 费菜属Phedimus | 费菜 Phedimus aizoon | 无 | 有 | 无 |
| 芸香科Rutaceae | 黄檗属Phellodendron | 川黄檗Phellodendron chinense | 有 | 无 | 有 |
| 绣球花科Hydrangeaceae | 山梅花属Philadelphus | 山梅花Philadelphus incanus | 无 | 有 | 无 |
| 绣球花科Hydrangeaceae | 山梅花属Philadelphus | 绢毛山梅花Philadelphus sericanthus | 无 | 有 | 无 |
| 荨麻科Urticaceae | 冷水花属Pilea | 粗齿冷水花Pilea sinofasciata | 无 | 无 | 有 |
| 伞形科Apiaceae | 茴芹属Pimpinella | 菱叶茴芹Pimpinella rhomboidea | 无 | 有 | 无 |
| 蔷薇科Rosaceae | 委陵菜属Potentilla | 委陵菜 Potentilla chinensis | 无 | 有 | 有 |
| 胡桃科Juglandaceae | 枫杨属Pterocarya | 枫杨Pterocarya stenoptera | 无 | 无 | 有 |
| 壳斗科Fagaceae | 栎属Quercus | 栓皮栎Quercus variabilis | 有 | 无 | 无 |
| 鼠李科Rhamnaceae | 猫乳属Rhamnella | 猫乳Rhamnella franguloides | 有 | 无 | 有 |
| 茶藨子科Grossulariaceae | 茶藨子属Ribes | 茶藨子Ribes janczewskii | 有 | 有 | 有 |
| 十字花科Brassicaceae | 蔊菜属Rorippa | 蔊菜Rorippa indica | 无 | 有 | 无 |
| 茜草科Rubiaceae | 茜草属Rubia | 茜草Rubia cordifolia | 无 | 有 | 有 |
| 蔷薇科Rosaceae | 悬钩子属Rubus | 喜阴悬钩子Rubus mesogaeus | 有 | 有 | 无 |
| 荚蒾科Viburnaceae | 接骨木属Sambucus | 接骨木Sambucus williamsii | 有 | 无 | 有 |
| 五味子科Schisandraceae | 五味子属Schisandra | 毛叶五味子Schisandra pubescens | 有 | 无 | 无 |
| 五味子科Schisandraceae | 五味子属Schisandra | 红花五味子Schisandra rubriflora | 无 | 无 | 有 |
| 禾本科Poaceae | 三毛草属Sibirotrisetum | 湖北三毛草Sibirotrisetum henryi | 有 | 有 | 有 |
| 石竹科Caryophyllaceae | 繁缕属Stellaria | 箐姑草Stellaria vestita | 无 | 有 | 有 |
| 蔷薇科Rosaceae | 红果树属Stranvaesia | 波叶红果树 Stranvaesia davidiana var. undulata |
无 | 无 | 有 |
| 漆树科Anacardiaceae | 漆树属Toxicodendron | 漆树Toxicodendron vernicifluum | 无 | 无 | 有 |
| 百合科Liliaceae | 油点草属Tricyrtis | 黄花油点草Tricyrtis pilosa | 无 | 无 | 有 |
| 忍冬科Caprifoliaceae | 缬草属Valeriana | 缬草Valeriana officinalis | 无 | 有 | 无 |
| 荚蒾科Viburnaceae | 荚蒾属Viburnum | 球核荚蒾Viburnum propinquum | 有 | 有 | 无 |
| 堇菜科Violaceae | 堇菜属Viola | 鸡腿堇菜Viola acuminata | 无 | 无 | 有 |
| 堇菜科Violaceae | 堇菜属Viola | 深圆齿堇菜Viola davidii | 无 | 无 | 有 |
| 堇菜科Apiaceae | 堇菜属Viola | 乌泡连Viola vaginata | 无 | 有 | 有 |
| 无患子科Sapindaceae | 槭属Acer | 有 | 有 | 有 | |
| 猕猴桃科Actinidiaceae | 猕猴桃属Actinidia | 有 | 有 | 有 | |
| 桔梗科Campanulaceae | 沙参属Adenophora | 无 | 有 | 无 | |
| 菊科Asteraceae | 蒿属Artemisia | 有 | 有 | 有 | |
| 菊科Asteraceae | 紫菀属Aster | 有 | 有 | 有 | |
| 豆科Fabaceae | 黄芪属Astragalus | 无 | 无 | 有 | |
| 桦木科Betulaceae | 桦木属Betula | 有 | 有 | 有 | |
| 桦木科Betulaceae | 鹅耳枥属Carpinus | 有 | 有 | 有 | |
| 卫矛科Celastraceae | 南蛇藤属Celastrus | 有 | 有 | 有 | |
| 桔梗科Campanulaceae | 党参属Codonopsis | 无 | 有 | 无 | |
| 山茱萸科Cornaceae | 山茱萸属Cornus | 有 | 有 | 有 | |
| 蓝果树科Nyssaceae | 珙桐属Davidia | 无 | 无 | 有 | |
| 五加科Araliaceae | 五加属Eleutherococcus | 有 | 有 | 无 | |
| 卫矛科Celastraceae | 卫矛属Euonymus | 有 | 有 | 有 | |
| 壳斗科Fagaceae | 水青冈属Fagus | 有 | 无 | 无 | |
| 禾本科Poaceae | 箭竹属Fargesia | 有 | 有 | 有 | |
| 绣球花科Hydrangeaceae | 绣球属Hydrangea | 有 | 有 | 有 | |
| 唇形科Lamiaceae | 香茶菜属Isodon | 无 | 有 | 无 | |
| 桑科Moraceae | 桑属Morus | 有 | 无 | 无 | |
| 藜芦科Melanthiaceae | 重楼属Paris | 无 | 无 | 有 | |
| 松科Pinaceae | 云杉属Picea | 有 | 无 | 无 | |
| 松科Pinaceae | 松属Pinus | 有 | 有 | 无 | |
| 杨柳科Salicaceae | 杨属Populus | 有 | 无 | 有 | |
| 蔷薇科Rosaceae | 李属Prunus | 有 | 有 | 无 | |
| 鼠李科Rhamnaceae | 鼠李属Rhamnus | 有 | 无 | 无 | |
| 杜鹃花科Ericaceae | 杜鹃属Rhododendron | 有 | 无 | 无 | |
| 蔷薇科Rosaceae | 蔷薇属Rosa | 无 | 无 | 有 | |
| 清风藤科Sabiaceae | 清风藤属Sabia | 无 | 无 | 有 | |
| 杨柳科Salicaceae | 柳属Salix | 有 | 有 | 有 | |
| 伞形科Apiaceae | 变豆菜属Sanicula | 无 | 有 | 无 | |
| 菊科Asteraceae | 风毛菊属Saussurea | 有 | 有 | 无 | |
| 五味子科Schisandraceae | 五味子属Schisandra | 无 | 有 | 无 | |
| 菝葜科Smilacaceae | 菝葜属Smilax | 无 | 有 | 有 | |
| 蔷薇科Rosaceae | 花楸属Sorbus | 有 | 有 | 有 | |
| 蔷薇科Rosaceae | 绣线菊属Spiraea | 无 | 有 | 有 | |
| 毛茛科Ranunculaceae | 唐松草属Thalictrum | 无 | 有 | 有 | |
| 锦葵科Malvaceae | 椴属Tilia | 有 | 无 | 有 | |
| 松科Pinaceae | 铁杉属Tsuga | 有 | 无 | 无 | |
| 伞形科Apiaceae | 有 | 有 | 无 | ||
| 马兜铃科Aristolochiaceae | 无 | 无 | 有 | ||
| 菊科Asteraceae | 有 | 有 | 无 | ||
| 唇形科Lamiaceae | 有 | 无 | 无 | ||
| 樟科Lauraceae | 有 | 有 | 无 | ||
| 木樨科Oleaceae | 有 | 无 | 无 | ||
| 禾本科Poaceae | 无 | 无 | 有 | ||
| 蔷薇科Rosaceae | 无 | 无 | 有 | ||
| 虎耳草科Saxifragaceae | 无 | 有 | 有 | ||
| 葡萄科Vitaceae | 有 | 无 | 有 |
不同季节川金丝猴的食物组成也有一定差异, 川金丝猴在春季的偏好食物为花楸属一种(27.34%)和藤山柳(16.56%);秋季的偏好食物是委陵菜(22.12%)和花楸属一种(16.52%);冬季的偏好食物为地海椒(33.49%)和花楸属一种(28.13%)不同季节的食物组成见图 2。

|
| 图 2 不同季节川金丝猴的食物组成 Fig. 2 Food composition of golden snub-nosed monkeys in different seasons |
从不同季节川金丝猴食物种类venn图中可以看出不同季节的食物重叠情况, 3个季节的共有食物有20种, 冬季的特有食物有16种, 春季有29种, 秋季有27种, 分别占植物总数的15.75%、12.60%、22.83%和21.26%。三个季节的共有食物包括勾儿茶、喜阴悬钩子(Rubus mesogaeus)、湖北三毛草(Sibirotrisetum henryi)、花楸属一种、槭属一种(Acer sp.)和山茱萸属一种等, 冬季特有食物主要有地海椒、青荚叶(Helwingia japonica)、连香树(Cercidiphyllum japonicum)、云杉属一种(Picea sp.)和鼠李属一种(Rhamnus sp.)等, 春季特有食物主要包括菱叶茴芹、鄂西卷耳(Cerastium wilsonii)、活血丹(Glechoma longituba)和五味子属一种(Schisandra sp.), 秋季特有食物包括构树(Broussonetia papyrifera)、栾树(Koelreuteria paniculata)、枫杨(Pterocarya stenoptera)和漆树(Toxicodendron vernicifluum)等。春季和秋季之间的重叠食物种类最多有舌叶金腰(Chrysosplenium glossophyllum)、委陵菜和茶藨子(Ribes janczewskii)等35种, 春季和冬季的重叠食物种类有巴山冷杉(Abies fargesii)、勾儿茶和藤山柳等32种而冬季和秋季重叠食物种类最少仅有宜昌橙(Citrus cavaleriei)、皂荚(Gleditsia sinensis)和接骨木(Sambucus williamsii)等28种, 分别占总数的27.56%和22.83%(图 3)。

|
| 图 3 不同季节食物种类重叠分布Venn图 Fig. 3 Venn diagram of food overlapping distribution in different seasons |
使用R软件vegan程序包计算不同季节川金丝猴食物的多样性指数, 比较发现川金丝猴在春季的食物多样性最高, 秋季次之, 冬季最低。而春季的Pielou均匀度指数最高即食物种类分布最均匀, 冬季均匀度指数最低, 在3个季节中分布最不均匀(表 2)。
| 季节 Seasons |
Simpson指数 Simpson |
Shannon指数 Shannon |
均匀度指数 Pielou |
| 冬季Winter | 0.787 | 1.996 | 0.496 |
| 春季Spring | 0.877 | 2.760 | 0.637 |
| 秋季Autumn | 0.868 | 2.447 | 0.576 |
研究印证了传统的神农架川金丝猴食性研究结果[33], 并进一步丰富了川金丝猴的食物组成。研究结果的食源植物种类虽小于传统食性研究的结果, 但在科(62)和属(110)水平上均高于传统食性研究结果, 这表明DNA条形码技术可能较传统的方法鉴定出更多的食源植物类群。食性结果中的地海椒、委陵菜和菱叶茴芹等草本植物在已有的川金丝猴食性研究结果中并未出现, 可能是因为显微分析法等难以鉴别粪便中残留的草木植物, 而野外观察法鉴定草木植物的识别率不高, 使得传统的食性结果中草木植物在川金丝猴的食物组成中占比较低[33]。发现了神农架川金丝猴食物组成中未记录过的食物, 如箭竹属植物, 但在四川白河川金丝猴和白马雪山的滇金丝猴(R. bieti)食物中均发现过箭竹属植物, 因此推测神农架川金丝猴可能取食箭竹属植物的叶片或者取食其竹笋[34—35]。在梅花鹿(Cervus nippon)和豹猫(Prionailurus bengalensis)等动物的食性研究中也发现基于DNA条形码研究方法比传统的研究方法能够得到更丰富的食物组成[36—37]。
食性研究结果表明神农架国家公园川金丝猴补食群的食物组成存在季节差异, 冬季食物多样性维持在最低水平, 春季食物多样性明显升高, 秋季时又开始下降, 这与神农架植物群落物候特性是一致的[38]。在冬季, 川金丝猴喜食植物的叶片、花和果实已经凋落, 而且降雪过后林下的草本植物也被覆盖, 川金丝猴的可获得食物数量锐减。除此之外, 植物的营养成分随季节的变化也对川金丝猴的食物组成造成了影响, 从冬季至春季神农架川金丝猴栖息地内的植物种类和营养成分含量不断变化, 致使川金丝猴的食物组成也发生变化[39]。
建立了川金丝猴食物DNA条形码参考数据库, 包括了神农架国家公园分布的1185种植物参考序列。但是数据库中缺失松萝属植物及神农架部分特有种的trnL序列片段, 因此食性结果中未鉴别出金丝猴经常取食的松萝(Usnea diffracta)。表明建立本地植物的序列数据库相比于使用公共数据库具有更高的物种鉴别率, 特别是在种和属的级别上[40—41]。但由于序列信息均来自于公共数据库, 其中存在的错误序列也会对鉴别结果造成一定影响, 因此建立本地化的物种序列数据库仍是非常重要的[42]。
DNA条形码的选择也是影响物种鉴定分辨率的重要因素, 选用的trnL为叶绿体DNA片段, 因此仅能用于识别含叶绿体的部分植物, 由于trnL条形码在同属近缘种之间的鉴别成功率较低, 因此部分植物仅鉴定到属水平。鉴定出的127种食物中, 有60%左右的食物鉴定到物种级别, 这与所选择的DNA条形码鉴别率有很大关系。用于植物鉴别的条形码包括trnL、ITS、rbcL、matK等, 不同条形码的鉴别率有所差异[43]。表明条形码组合使用有更高的识别成功率, 如rbcL和matK组合使用对物种的识别率高达98%[44]。因此, 未来的食性研究中应选用多种条形码进行物种鉴别以提高物种鉴别精确度。
使用DNA条形码技术得到了神农架川金丝猴不同季节的食物组成, 证实了DNA条形码技术在野生灵长类动物食性研究中的可行性, 为植食性动物的食性研究提供了方法借鉴。基于食性结果, 可以为川金丝猴野外种群的补食和笼养群的人工饲养制定更科学的食谱, 同时对川金丝猴主要取食的植物重点保护, 以促进其栖息地的恢复和野外种群数量的增长。本研究仍存在一定的不足, 虽然在样本采集过程中通过大小、形态及采集地点尽可能地收集不同个体的粪便, 但是仍可能存在来自同一个体的粪便样本, 这会对研究结果的准确性造成影响。因此, 使用本方法时应对粪便样本进行个体识别, 在食性分析时排除来自同一个体的粪便样本。样品采集时间仅选取了, 春季、秋季和冬季三个时间段, 缺少川金丝猴食物种类可能最为丰富的夏季。川金丝猴取食昆虫和脊椎动物已有报道[45—46], 还有待对其粪便中存在的动物DNA进行分析, 以便得到更为完整的川金丝猴食物组成。
| [1] |
郭艳萍, 张浩, 赵新钢, 罗海玲, 张英俊. DNA宏条形码技术在食草动物食性研究中的应用. 生物技术通报, 2021, 37(3): 252-260. |
| [2] |
Galán-Acedo C, Arroyo-Rodríguez V, Andresen E, Arasa-Gisbert R. Ecological traits of the world's Primates. Scientific Data, 2019, 6(1): 55. DOI:10.1038/s41597-019-0059-9 |
| [3] |
程铭昊, 张于光. DNA宏条形码技术在灵长类动物食性研究中的应用. 陆地生态系统与保护学报, 2022, 2(3): 80-86. |
| [4] |
Cui Z W, Wang Z L, Zhang S Q, Wang B S, Lu J Q, Raubenheimer D. Living near the limits: effects of interannual variation in food availability on diet and reproduction in a temperate primate, the Taihangshan macaque (Macaca mulatta tcheliensis). American Journal of Primatology, 2020, 82(1): e23080. DOI:10.1002/ajp.23080 |
| [5] |
Matthews J K, Ridley A, Kaplin B A, Grueter C C. A comparison of fecal sampling and direct feeding observations for quantifying the diet of a frugivorous primate. Current Zoology, 2020, 66(4): 333-343. DOI:10.1093/cz/zoz058 |
| [6] |
Pertoldi C, Schmidt J B, Thomsen P M, Nielsen L B, de Jonge N, Iacolina L, Muro F, Nielsen K T, Pagh S, Lauridsen T L, Andersen L H, Yashiro E, Lukassen M B, Nielsen J L, Elmeros M, Bruhn D. Comparing DNA metabarcoding with faecal analysis for diet determination of the Eurasian otter (Lutra lutra) in Vejlerne, Denmark. Mammal Research, 2021, 66(1): 115-122. DOI:10.1007/s13364-020-00552-5 |
| [7] |
Pansu J, Hutchinson M C, Anderson T M, Te Beest M, Begg C M, Begg K S, Bonin A, Chama L, Chamaillé-Jammes S, Coissac E, Cromsigt J P G M, Demmel M Y, Donaldson J E, Guyton J A, Hansen C B, Imakando C I, Iqbal A, Kalima D F, Kerley G I H, Kurukura S, Landman M, Long R A, Munuo I N, Nutter C M, Parr C L, Potter A B, Siachoono S, Taberlet P, Waiti E, Kartzinel T R, Pringle R M. The generality of cryptic dietary niche differences in diverse large-herbivore assemblages. Proceedings of the National Academy of Sciences of the United States of America, 2022, 119(35): e2204400119. |
| [8] |
刘山林. DNA条形码参考数据集构建和序列分析相关的新兴技术. 生物多样性, 2019, 27(5): 526-533. |
| [9] |
Lyke M M, Di Fiore A, Fierer N, Madden A A, Lambert J E. Metagenomic analyses reveal previously unrecognized variation in the diets of sympatric Old World monkey species. PLoS One, 2019, 14(6): e0218245. DOI:10.1371/journal.pone.0218245 |
| [10] |
Milner A M, Wood S A, Docherty C, Biessy L, Takenaka M, Tojo K. Winter diet of Japanese macaques from Chubu Sangaku National Park, Japan incorporates freshwater biota. Scientific Reports, 2021, 11(1): 23091. DOI:10.1038/s41598-021-01972-2 |
| [11] |
Hofreiter M, Kreuz E, Eriksson J, Schubert G, Hohmann G. Vertebrate DNA in fecal samples from bonobos and gorillas: evidence for meat consumption or artefact?. PLoS One, 2010, 5(2): e9419. DOI:10.1371/journal.pone.0009419 |
| [12] |
Mallott E K, Garber P A, Malhi R S. Integrating feeding behavior, ecological data, and DNA barcoding to identify developmental differences in invertebrate foraging strategies in wild white-faced capuchins (Cebus capucinus). American Journal of Physical Anthropology, 2017, 162(2): 241-254. DOI:10.1002/ajpa.23113 |
| [13] |
Rowe A K, Donohue M E, Clare E L, Drinkwater R, Koenig A, Ridgway Z M, Martin L D, Nomenjanahary E S, Zakamanana F, Randriamanandaza L J, Rakotonirina T E, Wright P C. Exploratory analysis reveals arthropod consumption in 10 lemur species using DNA metabarcoding. American Journal of Primatology, 2021, 83(6): e23256. DOI:10.1002/ajp.23256 |
| [14] |
Yang Y, Groves C, Garber P, Wang X W, Li H, Long Y C, Li G S, Tian Y P, Dong S H, Yang S Y, Behie A, Xiao W. First insights into the feeding habits of the Critically Endangered black snub-nosed monkey, Rhinopithecus strykeri (Colobinae, Primates). Primates, 2019, 60(2): 143-153. DOI:10.1007/s10329-019-00717-0 |
| [15] |
国家林业和草原局, 农业农村部. 国家重点保护野生动物名录(2021年2月1日修订). 野生动物学报, 2021, 42(2): 605-640. |
| [16] |
蒋志刚, 江建平, 王跃招, 张鹗, 张雁云, 李立立, 谢锋, 蔡波, 曹亮, 郑光美, 董路, 张正旺, 丁平, 罗振华, 丁长青, 马志军, 汤宋华, 曹文宣, 李春旺, 胡慧建, 马勇, 吴毅, 王应祥, 周开亚, 刘少英, 陈跃英, 李家堂, 冯祚建, 王燕, 王斌, 李成, 宋雪琳, 蔡蕾, 臧春鑫, 曾岩, 孟智斌, 方红霞, 平晓鸽. 中国脊椎动物红色名录. 生物多样性, 2016, 24(5): 500-551, 615. |
| [17] |
Yu Y, He G, Li D Y, Zhao X M, Chang J, Liu X C, Xiang Z F, Li B G, Li M. Climate change challenge, extinction risk, and successful conservation experiences for a threatened primate species in China: golden snub-nosed monkey (Rhinopithecus roxellana). Zoological Research, 2022, 43(6): 940-944. DOI:10.24272/j.issn.2095-8137.2022.198 |
| [18] |
曹国斌, 朱兆泉, 杨敬元, 姚辉, 谭文赤, 刘胜祥, 余辉亮. 湖北川金丝猴现状及保护研究. 野生动物学报, 2019, 40(3): 602-609. |
| [19] |
Fang G, Li M, Liu X J, Guo W J, Jiang Y T, Huang Z P, Tang S Y, Li D Y, Yu J, Jin T, Liu X G, Wang J M, Li S, Qi X G, Li B G. Preliminary report on Sichuan golden snub-nosed monkeys (Rhinopithecus roxellana roxellana) at Laohegou Nature Reserve, Sichuan, China. Scientific Reports, 2018, 8(1): 16183. DOI:10.1038/s41598-018-34311-z |
| [20] |
Hou R, Chapman C A, Jay O, Guo S, Li B, Raubenheimer D. Cold and hungry: combined effects of low temperature and resource scarcity on an edge-of-range temperate primate, the golden snub-nose monkey. Ecography, 2020, 43(1): 1672-1682. |
| [21] |
Liu X C, Li F, Jiang J, Wang X J, Li Y M. Age-sex analysis for the diet of Sichuan snub-nosed monkeys (Rhinopithecus roxellana) in Shennongjia National Nature Reserve, China. Primates, 2016, 57(4): 479-487. DOI:10.1007/s10329-016-0535-1 |
| [22] |
李亭亭, 汪正祥, 龚苗, 雷耘, 林丽群, 戴璨, 任雁. 神农架国家级自然保护区川金丝猴栖息地的植物群落分类及特征. 植物科学学报, 2016, 34(4): 563-574. |
| [23] |
刘峻杉, 朱遵燕, 姚辉, 杨万吉, 禹洋, 向左甫. 川金丝猴非母亲照料行为的影响因素及其母亲获益——以神农架川金丝猴投食群为例. 兽类学报, 2021, 41(2): 144-155. |
| [24] |
周芸芸, 张宇, 杨万吉, 蒋军, 余辉亮, 李迪强, 张于光. 不同保存方法对川金丝猴粪便DNA提取效果的影响. 现代生物医学进展, 2018, 18(1): 43-47. |
| [25] |
Riaz T, Shehzad W, Viari A, Pompanon F, Taberlet P, Coissac E. ecoPrimers: inference of new DNA barcode markers from whole genome sequence analysis. Nucleic Acids Research, 2011, 39(21): e145. DOI:10.1093/nar/gkr732 |
| [26] |
Quéméré E, Hibert F, Miquel C, Lhuillier E, Rasolondraibe E, Champeau J, Rabarivola C, Nusbaumer L, Chatelain C, Gautier L, Ranirison P, Crouau-Roy B, Taberlet P, Chikhi L. A DNA metabarcoding study of a primate dietary diversity and plasticity across its entire fragmented range. PLoS One, 2013, 8(3): e58971. DOI:10.1371/journal.pone.0058971 |
| [27] |
Rognes T, Flouri T, Nichols B, Quince C, Mahé F. VSEARCH: a versatile open source tool for metagenomics. PeerJ, 2016, 4(1): e2584. |
| [28] |
谢丹, 王玉琴, 张小霜, 吴玉, 杨敬元, 张代贵. 神农架国家公园植物采集史及模式标本名录. 生物多样性, 2019, 27(2): 211-218. |
| [29] |
铁军, 张晶, 彭林鹏, 杨林森, 胡德夫, 张志翔. 神农架川金丝猴栖息地优势树种生态位及食源植物. 植物生态学报, 2009, 33(3): 482-491. |
| [30] |
邵昕宁, 宋大昭, 黄巧雯, 李晟, 姚蒙. 基于粪便DNA及宏条形码技术的食肉动物快速调查及食性分析. 生物多样性, 2019, 27(5): 543-556. |
| [31] |
宫爽, 张萍, 栾雪, 包衡, 姜广顺. 粪便显微分析和DNA宏条形码技术在野猪食性分析中的应用和比较. 野生动物学报, 2022, 43(3): 645-653. |
| [32] |
郑佳鑫, 左清秋, 王刚, 韦旭, 翁晓东, 王正寰. 藏狐的食物组成及其季节差异. 兽类学报, 2023, 43(4): 398-411. |
| [33] |
Li J Y, Zhang J, Tie J, Wu C K, Zhang Z X. Abundance and distribution of the diet of the Sichuan snub-nosed monkey (Rhinopithecus roxellana) in the Shennongjia Nature Reserve in China. Acta Theriologica Sinica, 2015, 35(1): 14-28. |
| [34] |
李沂韦. 四川白河川金丝猴(Rhinopithecus roxellana)的生态学研究-家域利用与食物选择[D]. 南充: 西华师范大学, 2017.
|
| [35] |
黎大勇, 任宝平, 和鑫明, 胡刚, 李保国, 李明. 白马雪山自然保护区响古箐滇金丝猴的食性. 兽类学报, 2011, 31(4): 338-346. |
| [36] |
黄乃莹. 基于分子粪便法和粪便显微法的梅花鹿和狍食性比较研究[D]. 哈尔滨: 东北林业大学, 2023.
|
| [37] |
滕扬, 盖立新, 李建, 房新民, 杨南, 鲍伟东. 两种分析方法在豹猫食物构成中的应用比较. 兽类学报, 2023, 43(1): 50-58. |
| [38] |
胡铮铮, 肖飞, 冯奇, 王立辉, 王宙. 东秦巴山地自然保护地样区植被物候的过渡特性研究. 自然保护地, 2023, 3(2): 26-33. |
| [39] |
铁军, 张晶, 彭林鹏, 张志翔. 神农架川金丝猴冬春季节取食影响因素. 生态学杂志, 2011, 30(11): 2562-2569. |
| [40] |
Kolter A, Gemeinholzer B. Plant DNA barcoding necessitates marker-specific efforts to establish more comprehensive reference databases. Genome, 2021, 64(3): 265-298. DOI:10.1139/gen-2019-0198 |
| [41] |
吴峰, 温佩颖, 唐志珍, 李青鉴, 朱頔, 王天明, 葛剑平, 王红芳. 东北虎豹国家公园植物DNA微条形码数据库. 北京师范大学学报: 自然科学版, 2023, 59(4): 623-628. |
| [42] |
Jin S, Kim K Y, Kim M S, Park C. An assessment of the taxonomic reliability of DNA barcode sequences in publicly available databases. Algae, 2020, 35(3): 293-301. DOI:10.4490/algae.2020.35.9.4 |
| [43] |
Tercel M P T G, Symondson W O C, Cuff J P. The problem of omnivory: a synthesis on omnivory and DNA metabarcoding. Molecular Ecology, 2021, 30(10): 2199-2206. DOI:10.1111/mec.15903 |
| [44] |
魏亚男, 王晓梅, 姚鹏程, 陈小勇, 李宏庆. 比较不同DNA条形码对中国海岸带耐盐植物的识别率. 生物多样性, 2017, 25(10): 1095-1104. |
| [45] |
Yang B, Zhang P, Garber P A, Hedley R, Li B G. Sichuan snub-nosed monkeys (Rhinopithecus roxellana) consume cicadas in the Qinling Mountains, China. Folia Primatologica; International Journal of Primatology, 2016, 87(1): 11-16. DOI:10.1159/000443733 |
| [46] |
朱遵燕, 黄天鹏, 姚辉, 杨万吉, 向左甫. 神农架川金丝猴捕食矛纹草鹛雏鸟简报. 兽类学报, 2019, 39(5): 585-589. |