 2024, Vol. 44
2024, Vol. 44文章信息
- 周鑫, 吕青昕, 桑珠扎西, 扎西, 周密, 黎勇, 孟秀祥
- ZHOU Xin, LÜ Qingxin, SANGZHU Zhaxi, ZHA Xi, ZHOU Mi, LI Yong, MENG Xiuxiang
- 青藏高原东北缘濒危马麝迁地保育种群繁殖成效与社会性和社会网络的关系
- Reproduction success and the relationships to sociality and social networks in ex-situ conserved Alpine musk deer in the east-northern edging area of the Qinghai-Tibet Plateau
- 生态学报. 2024, 44(15): 6417-6423
- Acta Ecologica Sinica. 2024, 44(15): 6417-6423
- http://dx.doi.org/10.20103/j.stxb.202312222784
-
文章历史
- 收稿日期: 2023-12-22
- 网络出版日期: 2024-05-30





2. 西藏大学生态环境学院, 拉萨 850000;
3. 四川逢春麝业集团有限公司, 成都 610030;
4. 西昌学院动物科技学院, 西昌 615000
2. School of Ecology and Environment, Tibet University, Lasa 850000, China;
3. Sichuan Fengchun Musk Deer Technology Group Company Limited, Chengdu 610030, China;
4. School of Animal Science, Xichang University, Xichang 615000, China
动物个性是动物个体间跨时间和情景的稳定行为差异或倾向, 社群环境中的动物个性发育与社会结构和个体间社会行为关系紧密[1]。社会网络分析方法(Social Network Analysis, SNA)是研究社群环境下动物个性的重要工具, 可以通过分析个体间的社会相互作用及群体的社会结构来研究动物个性与社群特征的关系[2]。SNA已成为动物个性研究领域的重要手段[3—4]。
“社会性”是动物的重要个性之一, 表征动物社群中个体的合群倾向及与社群个体合作的程度[5]。动物的社会性与其适合度(fitness)密切相关。动物社群中个体的社会性与其对食物资源和配偶资源的占有有关, 与个体的免疫力和健康度等也关系紧密[6—8]。在群居性哺乳动物中, 动物的社会性影响个体适应性及繁殖策略等[9]。在狒狒(Papio cynocephalus ursinus)[10]、山雀(Mandrillus sphinx)[11]和野马(Equus caballus)[12]等物种, 个体的社会性和个体间的社会联系可以显著提高个体繁殖成效。
通过动物行为计测动物个性已被广泛认可[13—15]。社会网络分析可以通过动物个体层面的互动以及群体层面的社会结构全面研究动物个性[2], 是动物个性研究的重要工具[16—17]。迄今为止, 社会网络分析以及社会性与繁殖成功之间的研究主要侧重于强社会性动物, 而缺乏对在群居驯养环境中表现出更强独居性的哺乳动物的相关研究。
马麝是分布于青藏高原及周边地区的濒危资源性动物, 成年雄麝分泌的麝香被广泛用于传统医药及国际香水业[18]。因生境丧失、质量下降及历史上的过度利用等原因, 马麝已极度濒危, 被列为国家一级保护动物。除基于野生种群和栖息地保护的就地保育(in-situ conservation)之外, 马麝迁地保育(ex-situ conservation)是濒危麝类动物及药用麝香可持续供给的的重要方式[19], 但前提是深入了解其迁地保育种群的行为格局及个性特征及对迁地保育环境的响应方式。
野生马麝是典型小型森林型独栖有蹄类动物, 个体的社会相互作用极为稀少, 而且多发生在繁殖交配等有限生理阶段[20]。在现代群养性迁地保育环境下, 个体所处的人工环境为社群条件。深入了解迁地保育的马麝社群的个体社会性和社会网络特征及与繁殖成效间的关系, 是对濒危马麝进行成功繁育、迁地保育及野外放归建立基础种群准备的基础。
本研究对青藏高原东北缘甘肃平凉马麝繁育中心的迁地保育马麝种群的社会行为、社会网络、粪样类固醇激素和繁殖成效及相互关系开展研究, 其结果可为濒危麝类迁地保育及药用麝香的可持续供给提供重要参考。
1 实验方法 1.1 研究区域及实验动物本研究于2022年7月1日至8月31日在甘肃平凉马麝繁育中心(以下简称麝场)开展。麝场地处青藏高原东北缘, 属祁连山东延余脉区和六盘山西麓, 场地海拔1800 m, 四季分明。年平均气温8.1 ℃, 其中7月平均温度最高, 1月最低;年降水量489 mm, 多集中在夏季(7—9月);年均无霜期142 d。麝场的马麝圈舍设置统一, 均由一个10 m × 10 m的活动场(天然泥质基底)和中央的2 m × 3 m × 2 m露台组成, 一侧具有4个小室。麝场采用统一的管理模式, 每日饲喂两次(08:00、17:00), 饲喂同时进行圈舍清扫, 其余时间马麝不受干扰。马麝均佩戴耳号可进行个体识别。实验共涉及7个圈舍(1雄3雌圈)内的21头成年自繁雌麝(2—5岁)。
麝类幼仔在1月龄前死亡概率最大[21], 本文雌麝繁殖成效定义:雌麝分娩且哺育其仔麝存活至1月龄, 即为繁殖成功;雌麝空怀、流产、死胎(still birth)或其自然哺乳仔麝在1月龄前死亡, 则为繁殖失败。圈群的繁殖成功率指各圈群雌麝中繁殖成功的雌麝所占比, 即圈群繁殖成效。
1.2 行为取样和数据分析采用焦点动物取样法(Focal Sampling Method)和所有事件记录法(All Occurrence Recording)[22]进行数据采集。研究者在圈舍20 m外的掩体处进行行为记录, 用10 × 22°望远镜协助进行个体识别, 记录各行为的行为发出个体、行为指向个体及持续时间等行为变量;每个样本持续时间为10 min;每周进行两天, 每天进行两次行为取样, 取样时间为每日晨间和黄昏(06:00—08:00、18:00—20:00)[23]。
参照报道的马麝的社会行为[24—25], 定义马麝社会行为:互饰(mutual grooming)指两个及以上个体相互舔毛及互饰等;接触(mutual contact)指一个马麝个体非侵犯性地用头部接触其它个体的身体;趋近(closeness)马麝个体无攻击意向地向另一个体靠近至一个身体距离单元以内。
整理行为取样数据, 计算个体的社会性强度(展现社会行为的持续时间/行为取样单元%)。根据圈养马麝社会性均值, 将样本动物分为高社会性组(大于等于均值)及低社会性组(小于均值)。
将马麝个体间的社会行为关系数据转为二值邻接矩阵, 用Ucinet 6.0[26]分析社会网络属性特征, 计算社会网络密度, 计算公式为:
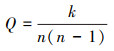
|
(1) |
其中, Q表示网络密度, 社会性行为有指向, n(n-1)表示网络中马麝个体间能够发生最多不同种类的社会性行为关系的总数, k表示网络中马麝个体间已经发生的社会行为总数。
1.3 粪样收集和粪便类固醇激素代谢物浓度测定在对马麝个体进行行为取样时同期收集粪样, 粪样采集后立即装入保鲜袋密封, 并记录个体、日期, 放入-20℃冰箱内储存待测。
参照Wasser等[27]和孙太福等[28]的方法, 将冷冻粪便样品解冻后, 用电热鼓风干燥箱60 ℃烘干12 h至恒重, 粉碎研磨干燥粪样;用万分之一天平称取0.1 g粪便于离心管中, 加入80%的乙醇1 mL;涡旋振荡1 min, 70 ℃水浴15 min, 再涡旋振荡1 min, 冷却至4℃, 样品离心(4000 r/min)15 min, 取上清液至另一离心管中;残渣加入0.5 mL的80%乙醇重新悬浮, 涡旋振荡1 min, 4000 r/min再次离心15 min;合并上清液, 将含有乙醇的上清液70 ℃水浴空气吹干;加600 μL的0.01 M PBS(磷酸钠盐缓冲液)复溶, 复溶完全后4 ℃保存待测。
采用放射免疫分析药盒(北京北方生物技术研究所), 通过放射免疫分析法(RIA), 用γ-计数器测定各管沉淀的放射性计数, 计算剂量反应标准曲线方程, 检测粪样中皮质醇、雌二醇代谢物含量。
1.4 数据处理与分析采用Kolmogorov-Smirnov检验数据的正态性, 并采用Leven′s test对数据进行方差齐次性检验。根据检验结果, 采用Pearson或者Spearman相关分析检验繁殖成效、社会性强度、社会网络密度及粪样类固醇激素代谢物水平间的相关性。并进一步使用t test或Mann-Whitney U test检验繁殖成功与繁殖失败雌麝的社会性强度、粪样类固醇激素代谢物浓度的差异;按社会性平均值将马麝个体分为高社会性组和低社会性组, 并进一步使用t test或Mann-Whitney U test检验高社会性组与低社会性组的粪便类固醇激素代谢物浓度差异。结果均以均值±标准差表示, 显著性水平设置为P=0.05。所有数据分析均在SPSS 26.0环境下进行。
2 结果 2.1 圈养马麝的繁殖成效与社会性雌麝的社会性强度均值为(16.30±1.44)% (n=21), 范围为7.02%—32.00%。个体的社会性强度与繁殖成效显著相关(r=0.551, P=0.010 < 0.05);繁殖失败雌麝的社会性((12.38±1.46)%, n=10)显著低于繁殖成功雌麝((19.86±1.88)%, n=11) (T=-3.095, P=0.006 < 0.05)。
2.2 繁殖成效与社会网络密度雌麝圈群的社会网络密度均值为0.77±0.03 (n=7), 范围为0.67—1.00。圈群繁殖成效均值为(52.29±17.62)% (n=7)。圈群繁殖成效随网络密度升高而提高, 两者间相关显著(r=0.762, P=0.046 < 0.05)。
2.3 繁殖成效与粪样类固醇代谢物水平的关系雌麝的粪样皮质醇代谢物水平均值为(12.02±0.95) ng/g, 范围为6.18—24.82 ng/g。繁殖失败雌麝的皮质醇代谢物水平((13.89±1.60) ng/g, n=10)略高于繁殖成功雌麝((10.31±0.86) ng/g, n=11), 差异呈边界显著(t=2.20, P=0.058>0.05), 两者间存在一定的相关性(r=-0.420, P=0.058)。
雌麝的粪样雌二醇代谢物水平均值为(6.01±0.56) ng/g (n=21), 范围为2.14—11.10 ng/g。繁殖失败雌麝的粪样雌二醇代谢物水平((4.78±0.76) ng/g, n=10)与繁殖成功雌麝((6.63±0.90) ng/g, n=11)没有显著差异(t=-1.55, P=0.139>0.05), 两者间相关也不显著(r=0.299, P=0.188>0.05) (图 1)。

|
| 图 1 不同繁殖结果雌麝粪样类固醇激素代谢物浓度图 Fig. 1 Plot of fecal steroid hormone metabolite concentrations in female musk with different reproductive outcomes |
个体的社会性随雌麝粪样皮质醇代谢物的浓度的升高而下降, 两者间呈显著负相关(r=-0.437, P=0.047 < 0.05), 方程y=-0.29x+16.70可较好地拟合皮质醇代谢物水平与社会性之间的关系;社会性与粪样雌二醇代谢物浓度相关不显著(r=0.066, P=0.776>0.05)所示, 方程y=0.028x+5.29可较好地拟合雌二醇水平与社会性之间的关系(图 2)。

|
| 图 2 皮质醇、雌二醇代谢物浓度随社会性变化图 Fig. 2 Plot of cortisol and estradiol metabolite concentrations with sociability |
将个体以社会性均值为基准分为强弱2类, 强社会性马麝的皮质醇代谢物浓度((10.75±0.99) ng/g, n=12)低于弱社会性马麝((13.70±1.68) ng/g, n=9), 但两者间无显著差异(t=1.594, P=0.127>0.05);强社会性马麝的雌二醇代谢物浓度(6.16±0.79) ng/g, n=12)大于弱社会性马麝((5.20±0.99) ng/g, n=9), 两者间无显著差异(t=-0.758, P=0.458>0.05)。社会网络密度与粪样皮质醇代谢物浓度、粪样雌二醇代谢物浓度相关均不显著(r=-0.355, P=0.114>0.05;r=-0.347, P=0.123>0.05)。
3 讨论动物社群内的社会性表现为个体倾向于合群和与其他个体合作。个体的社会性互动构建起社会网络, 并在信息传递、资源获取和个体交往中发挥重要作用[29]。群居性较强动物社群和群居性较弱动物社群间的差异较大。马鹿(Cervus canadensis)的平均社会行为持续时间占比超过60%[30], 而本研究中迁地保育马麝的社会行为持时间占比为16.3%左右, 最高也仅为32.0%, 表明马麝的社会性相对较弱。这与学者们对野生麝类动物的研究一致。野生麝独居性较强, 个体间仅存在松散的联系[31—32]。虽然迁地保育马麝生活在群居驯养模式下, 但个体之间的结构仍然相对松散, 社会性不太明显[33]。
雌性马麝在社会群体中的繁殖成效与其社会性程度密切相关。随着雌性马麝的社会性增加, 以及社会网络密度升高, 它们的繁殖成效也相应提高。较强的社会性表明雌性马麝在迁地保育社群内进行社交互动的时间较长。此外, 社群内更高的社会网络密度表示个体之间社交互动的频率和频次更多。长期以来, 学者们基于对野生麝的独居性强等生态习性, 经验性地认为麝类迁地保育应该模仿野生麝, 在独居环境进行[34—35], 本研究的结果则提出了不同的观点。
雌性马麝表现出较强的社会性以及较高的圈群社会网络密度, 这有助于顺利完成其繁殖过程, 进而提升繁殖成效。在发情和交配期间, 社群内雌雄马麝之间的社会接触越频繁, 有助于顺利完成发情和交配过程, 进而提高雌性马麝的受孕率, 减少雌性马麝的空怀率[36]。此外, 社会性较高的个体表现出相对较少的侵犯行为, 也较少成为侵犯行为的目标, 较少卷入与其他个体的冲突。与此同时, 社会网络密度较大的社群中侵犯行为的频率也较低, 社群更加稳定[24]。这些因素共同减少了社群内雌性马麝个体由于冲突和争斗引起的妊娠终止、流产和死胎的风险。在育幼哺乳阶段, 社会性较高的雌性马麝与其新生仔麝相处时间更长, 展现出更多的亲代哺养行为, 包括更频繁和持续的哺乳行为。这显著提高了仔麝的哺乳成功率[37]。因此, 如研究结果所示, 迁地保育雌性马麝的社会性水平越高, 社群的社会网络密度越大, 越有利于提高其繁殖成效。
马麝的粪样皮质醇代谢物水平与其社会性之间存在密切关联。皮质醇是一种应激激素, 通常在动物表现出更多攻击性行为时, 其体内皮质醇及其代谢物浓度会升高, 这一现象在其他动物如猫[38]和羚羊(Gazella dorcas)[39]中也被发现。然而, 由于马麝通常表现出相对较低的社会性, 因此在马麝群体内, 尤其是在群居社群环境中, 个体之间容易发生冲突[40]。然而, 随着迁地保育时间的延长, 马麝社群内部逐渐形成了更为稳定的社会结构, 这导致个体的社会性增强, 从而降低了攻击性行为如打斗等的发生概率。因此, 不再需要引发皮质醇水平升高以应对应激状态, 这解释了皮质醇水平与社会性强度之间呈现负相关的现象。此外, 社会性行为的增加还有助于缓解压力, 产生生理上的镇静作用[41]。在这方面, 与其他社会性动物的观察结果一致, 即社会性越高的马麝, 其皮质醇水平越低[10]。
高水平的粪样皮质醇代谢物与马麝的繁殖成效之间存在负相关关系。大量研究已经表明, 皮质醇对动物的繁殖成效有不利影响, 例如, 它可能抑制母羊的正常发情, 降低其生育能力[42]。此外, 皮质醇还可能抑制卵泡的发育, 减少食欲, 导致奶牛的繁殖成效下降[43]。因此, 高水平的皮质醇可能会降低马麝怀孕的机会。在本研究中, 对于繁殖成功的雌麝, 它们处于育幼哺乳期。一些研究表明, 在哺乳期的雌性动物中, 下丘脑-垂体-肾上腺轴会受到抑制, 从而减少皮质醇的分泌[44]。另一方面, 对于那些繁殖失败的雌麝, 有研究表明, 在雄性存在下的混合小型反刍类动物群中, 未能成功进入育幼期的雌性会承受更大的生殖压力[45]。因而本研究中成功进入育幼期的雌麝的皮质醇代谢物水平稍低于未成功繁殖的雌麝。
4 结论及建议本研究发现, 随圈养马麝的社会性和社会网络密度升高, 其繁殖成效也相应提高, 而粪样皮质醇代谢物水平则相对降低, 皮质醇水平升高展现出对马麝的繁殖成效的负面影响。
在迁地保育过程中, 构建麝群时需充分考虑社会性对繁殖成效的影响。在以迁地保育和野化训练为目的的马麝驯养中, 为了实现高繁殖成效, 可以首先采用群居驯养模式来扩大种群规模, 然后过渡到独居驯养模式, 以维持马麝的序位结构及较强的独居性;而在以繁育和麝香生产为目标的商业性养殖, 可采用群居驯养模式, 以促进马麝的社会性, 马麝较高的社会性可提升其繁殖成效和生产效益, 并有助于成功的麝类驯养和群体驯化。
| [1] |
Sih A, Bell A, Johnson J C. Behavioral syndromes: an ecological and evolutionary overview. Trends in Ecology & Evolution, 2004, 19(7): 372-378. |
| [2] |
Croft D P, Krause J, Darden S K, Ramnarine I W, Faria J J, James R. Behavioural trait assortment in a social network: patterns and implications. Behavioral Ecology and Sociobiology, 2009, 63(10): 1495-1503. DOI:10.1007/s00265-009-0802-x |
| [3] |
Fewell J H. Social insect networks. Science, 2003, 301(5641): 1867-1870. DOI:10.1126/science.1088945 |
| [4] |
Croft D P, Krause J, James R. Social networks in the guppy (Poecilia reticulata). Proceedings Biological Sciences, 2004, 271(Suppl 6): S516-S519. |
| [5] |
Wilson E O. Sociobiology: The new synthesis. The United States: Harvard University Press, 2000.
|
| [6] |
Nunn C L, Craft M E, Gillespie T R, Schaller M, Kappeler P M. The sociality-health-fitness nexus: synthesis, conclusions and future directions. Philosophical Transactions of the Royal Society of London Series B, Biological Sciences, 2015, 370(1669): 20140115. DOI:10.1098/rstb.2014.0115 |
| [7] |
Schaller M, Murray D R, Bangerter A. Implications of the behavioural immune system for social behaviour and human health in the modern world. Philosophical Transactions of the Royal Society of London Series B, Biological Sciences, 2015, 370(1669): 20140105. DOI:10.1098/rstb.2014.0105 |
| [8] |
Capitanio J P, Cole S W. Social instability and immunity in rhesus monkeys: the role of the sympathetic nervous system. Philosophical Transactions of the Royal Society of London Series B, Biological Sciences, 2015, 370(1669): 20140104. DOI:10.1098/rstb.2014.0104 |
| [9] |
Silk J B. The adaptive value of sociality in mammalian groups. Philosophical Transactions of the Royal Society of London Series B, Biological Sciences, 2007, 362(1480): 539-559. DOI:10.1098/rstb.2006.1994 |
| [10] |
Silk J B, Alberts S C, Altmann J. Social bonds of female baboons enhance infant survival. Science, 2003, 302(5648): 1231-1234. DOI:10.1126/science.1088580 |
| [11] |
Charpentier M J E, Huchard E, Widdig A, Gimenez O, Sallé B, Kappeler P, Renoult J P. Distribution of affiliative behavior across kin classes and their fitness consequences in mandrills. Ethology, 2012, 118(12): 1198-1207. DOI:10.1111/eth.12026 |
| [12] |
Cameron E Z, Setsaas T H, Linklater W L. Social bonds between unrelated females increase reproductive success in feral horses. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(33): 13850-13853. |
| [13] |
Travnik I C, Sant'Anna A C. Do you see the same cat that I see? Relationships between Qualitative Behaviour Assessment and indicators traditionally used to assess temperament in domestic cats. Animal Welfare, 2021, 30(2): 211-223. DOI:10.7120/09627286.30.2.211 |
| [14] |
Weiss A, Yokoyama C, Hayashi T, Inoue-Murayama M. Personality, subjective well-being, and the serotonin 1a receptor gene in common marmosets (Callithrix jacchus). PLoS One, 2021, 16(8): e0238663. DOI:10.1371/journal.pone.0238663 |
| [15] |
Payne E, Sinn D L, Spiegel O, Leu S T, Gardner M G, Godfrey S S, Wohlfeil C, Sih A. Consistent after all: behavioural repeatability in a long-lived lizard across a 6-year field study. Animal Behaviour, 2021, 174: 263-277. DOI:10.1016/j.anbehav.2021.01.025 |
| [16] |
Lusseau D, Newman M E J. Identifying the role that animals play in their social networks. Proceedings Biological Sciences, 2004, 271(Suppl 6): S477-S481. |
| [17] |
Krause J, Croft D P, James R. Social network theory in the behavioural sciences: potential applications. Behavioral Ecology and Sociobiology, 2007, 62(1): 15-27. DOI:10.1007/s00265-007-0445-8 |
| [18] |
Goh C L, Kwok S F. A simple method of qualitative analysis for musk ambrette, musk ketone and musk xylene in cologne. Contact Dermatitis, 1986, 14(1): 53-56. DOI:10.1111/j.1600-0536.1986.tb01153.x |
| [19] |
王静, 孙军平, 徐涛, 祁军, 张远林, 张学炎, 孟秀祥. 甘肃兴隆山保护区野生马麝种群动态研究. 生态学报, 2020, 40(21): 7997-8004. |
| [20] |
Zhao Y J, Wang J, Li Y, Zhou M, Weladji R B, Bonoan J T, Meng X X. Temporal pattern of parturition in captive Alpine Musk Deer (Moschus sifanicus). Biologia, 2020, 75(2): 259-266. DOI:10.2478/s11756-019-00293-0 |
| [21] |
徐正强, 徐宏发. 饲养林麝的种群特征和幼麝的存活研究. 兽类学报, 2003, 23(1): 17-20. |
| [22] |
Altmann J. Observational study of behavior: sampling methods. Behaviour, 1974, 49(3): 227-267. |
| [23] |
孟秀祥, 杨奇森, 冯祚建, 夏霖, 王培民, 蒋应文, 白正清, 李国林. 圈养马麝夏秋冬活动格局的比较. 兽类学报, 2002, 22(2): 87-97. |
| [24] |
Meng X, Cody N, Gong B, Xiang L. Stable fight strategies in endangered male musk deer (Moschus sifanicus). Animal Science Journal, 2012, 83: 617-622. DOI:10.1111/j.1740-0929.2011.01007.x |
| [25] |
褚梦真, 曾凡刚, 邓益娟, 王静, 黎勇, 周密, 汉尚春, 白守林, 盛岩, 孟秀祥. 甘肃庄浪圈养马麝的社群性及其影响因素. 生态学杂志, 2020, 39(12): 4172-4177. |
| [26] |
Borgatti S P, Everett M G, Freeman L C. UCINET 6.0 Version 1.00. Natick: Analytic Technologies, 1999.
|
| [27] |
Wasser S K, Monfort S L, Wildt D E. Rapid extraction of faecal steroids for measuring reproductive cyclicity and early pregnancy in free-ranging yellow baboons (Papio cynocephalus cynocephalus). Journal of Reproduction and Fertility, 1991, 92(2): 415-423. DOI:10.1530/jrf.0.0920415 |
| [28] |
孙太福, 王静, 蔡永华, 程建国, 黎勇, 周密, 盛岩, 孟秀祥. 圈养林麝粪样类固醇激素水平与麝香分泌的关系. 生态学报, 2020, 40(24): 9245-9251. DOI:10.5846/stxb202001210162 |
| [29] |
Pike T W, Samanta M, Lindström J, Royle N J. Behavioural phenotype affects social interactions in an animal network. Proceedings Biological Sciences, 2008, 275(1650): 2515-2520. |
| [30] |
Vander Wal E, Paquet P C, Messier F, McLoughlin P D. Effects of phenology and sex on social proximity in a gregarious ungulate. Canadian Journal of Zoology, 2013, 91(9): 601-609. DOI:10.1139/cjz-2012-0237 |
| [31] |
刘志霄, 盛和林. 我国麝的生态研究与保护问题概述. 动物学杂志, 2000, 35(3): 54-57. |
| [32] |
Yang Q S, Meng X X, Xia L, Feng Z J. Conservation status and causes of decline of Musk Deer (Moschus spp.) in China. Biological Conservation, 2003, 109(3): 333-342. DOI:10.1016/S0006-3207(02)00159-3 |
| [33] |
Meng X X, Zhou C Q, Hu J C, Li C, Meng Z B, Feng J C, Zhou Y J, Zhu Y J. Musk Deer farming in China. Animal Science, 2006, 82(1): 1-6. DOI:10.1079/ASC200516 |
| [34] |
Parry J R, Wu J Y. Musk Deer Farming as a Conservation Tool in China. Hong Kong: Trafic, East Asia. 2001.
|
| [35] |
Zhang B L. Musk deer their capture, domestication and care according to Chinese experience and methods. Unasylva, 1983, 35. |
| [36] |
Meng X X, Yang Q S, Xia L, Feng Z J, Jiang Y W, Wang P M. The temporal estrous patterns of female alpine musk Deer in captivity. Applied Animal Behaviour Science, 2003, 82(1): 75-85. DOI:10.1016/S0168-1591(03)00013-3 |
| [37] |
Meng X, Yang Q, Feng Z, Feng J, Perkins G, Xu H. Using behavioral data to predict reproductive success in captive female musk deer. North-Western Journal of Zoology, 2008, 4(2): 263-269. |
| [38] |
Finkler H, Terkel J. Cortisol levels and aggression in neutered and intact free-roaming female cats living in urban social groups. Physiology & Behavior, 2010, 99(3): 343-347. |
| [39] |
Salas M, Temple D, Abáigar T, Cuadrado M, Delclaux M, Enseñat C, Almagro V, Martínez-Nevado E, Quevedo M Á, Carbajal A, Tallo-Parra O, Sabés-Alsina M, Amat M, Lopez-Bejar M, Fernández-Bellon H, Manteca X. Aggressive behavior and hair cortisol levels in captive Dorcas gazelles (Gazella dorcas) as animal-based welfare indicators. Zoo Biology, 2016, 35(6): 467-473. DOI:10.1002/zoo.21323 |
| [40] |
Zhou X, Lv Q, Qin Y, Yuan N, Li Y, Zhou M, Meng X. Effects of social stress on the welfare of captive male Alpine musk deer: Stereotypic behavior, fecal cortisol, and musk secretion. Applied Animal Behaviour Science, 2023, 258: 105828. DOI:10.1016/j.applanim.2022.105828 |
| [41] |
Feh C, de Mazières J. Grooming at a preferred site reduces heart rate in horses. Animal Behaviour, 1993, 46(6): 1191-1194. DOI:10.1006/anbe.1993.1309 |
| [42] |
Dobson H, Fergani C, Routly J E, Smith R F. Effects of stress on reproduction in ewes. Animal Reproduction Science, 2012, 130(3/4): 135-140. |
| [43] |
De Rensis F, Scaramuzzi R J. Heat stress and seasonal effects on reproduction in the dairy cow—a review. Theriogenology, 2003, 60(6): 1139-1151. DOI:10.1016/S0093-691X(03)00126-2 |
| [44] |
Douglas A J, Brunton P J, Bosch O J, Russell J A, Neumann I D. Neuroendocrine responses to stress in mice: hyporesponsiveness in pregnancy and parturition. Endocrinology, 2003, 144(12): 5268-5276. DOI:10.1210/en.2003-0461 |
| [45] |
Delgadillo J A, Martin G B. Alternative methods for control of reproduction in small ruminants: a focus on the needs of grazing industries. Animal Frontiers, 2015, 5(1): 57-65. DOI:10.2527/af.2015-0009 |