 2024, Vol. 44
2024, Vol. 44文章信息
- 丁慧芳, 卞庆嘉, 陈青, 王亚芳, 徐志文, 余定坤, 王文娟
- DING Huifang, BIAN Qingjia, CHEN Qing, WANG Yafang, XU Zhiwen, YU Dingkun, WANG Wenjuan
- 鄱阳湖白鹤越冬期不同阶段食物组成
- Food compositions of Siberia cranes at Poyang Lake during different wintering periods
- 生态学报. 2024, 44(15): 6436-6444
- Acta Ecologica Sinica. 2024, 44(15): 6436-6444
- http://dx.doi.org/10.20103/j.stxb.202307241581
-
文章历史
- 收稿日期: 2023-07-24
- 网络出版日期: 2024-05-24





2. 江西鄱阳湖湿地保护与恢复国家长期科研基地 江西鄱阳湖湿地生态系统国家定位观测研究站, 南昌 330031;
3. 南昌大学生命科学学院, 南昌 330031;
4. 江西鄱阳湖国家级自然保护区管理局, 南昌 330038;
5. 北京林业大学生态与自然保护学院, 北京 100083;
6. 北京林业大学东亚-澳大利西亚候鸟迁徙研究中心, 北京 100083
2. Jiangxi Poyang Lake Wetland Conservation and Restoration National Permanent Scientific Research Base, National Ecosystem Research Station of Jiangxi Poyang Lake Wetland, Nanchang 330031, China;
3. School of Life Science, Nanchang University, Nanchang 330031, China;
4. Jiangxi Poyang Lake National Nature Reserve Administration, Nanchang 330038, China;
5. School of Ecology and Nature Conservation, Beijing Forestry University, Beijing 100083, China;
6. Center for East Asian-Australasian Flyway Studies, Beijing Forestry University, Beijing 100083, China
食物是鸟类赖以生存的基础, 也是影响鸟类数量和分布的重要因素[1]。对鸟类食性的研究能在一定程度上揭示鸟类的活动范围及栖息地选择, 为鸟类保护政策的制定提供科学依据, 也为物种间关系的研究提供理论基础[2]。近年来, 全球气候变化、人类干扰及栖息地退化等因素导致自然湿地的食物资源不断减少, 大量水鸟被迫寻找新的觅食地, 甚至转向农业用地觅食[3—4]。目前, 农业用地已成为众多水鸟的重要觅食地。
粪便显微镜检法是动物食性研究的传统方法, 该方法主要通过显微镜观察粪便中未被消化的角质碎片细胞结构来鉴定动物的食物组成。然而, 该方法操作繁琐, 费时费力, 且很多消化程度较高的食物无法被检测到, 因此结果的可靠性较低[5]。DNA宏条形码方法结合DNA条形码和高通量测序技术, 通过提取粪便或胃容物中的DNA, 对特定DNA序列(DNA条形码)进行PCR扩增和高通量测序, 并依据参考文库进行比对, 从而鉴定动物的食物组成[5]。该方法具有快速、准确、高效等优点, 不仅能够鉴定出消化程度较高的食物, 而且准确度较高, 能同时处理大量样本, 现已成为动物食性研究的主流方法[5]。
白鹤(Leucogeranus leucogeranus)隶属鹤形目鹤科, 为IUCN极危物种[6], 国家一级重点保护动物[7]。目前白鹤有三个种群, 它们均繁殖于俄罗斯北部, 但分别越冬于中国(东部种群)、印度(中部种群)和伊朗(西部种群)[7]。目前, 白鹤中部种群已经灭绝, 西部种群仅剩1只[7]。东部种群是最大的白鹤种群, 最新统计结果表明东部种群数量为5, 616只[8]。白鹤东部种群主要在俄罗斯雅库特繁殖, 迁徙途径我国东北地区后, 飞至鄱阳湖和黄河三角洲越冬, 部分个体在蒙古度夏[9]。
鄱阳湖是白鹤最重要的越冬地, 约85.7%的个体在鄱阳湖越冬[8]。白鹤在鄱阳湖以取食行为为主[10], 其中沉水植物苦草冬芽(Vallisneria spp.) 是白鹤最主要的传统食物[11—12]。20世纪70年代和80年代, 沉水植被在长江中下游流域湖泊中占主导地位[13], 然而, 近年来由于鄱阳湖湿地生态环境质量恶化, 沉水植被退化严重[14]。1983—2013年的植被调查结果显示, 鄱阳湖沉水植被面积减少了37.7%[13]。此外, 苦草冬芽密度和生物量也呈下降趋势, 2010年、2012年、2015—2016年冬季, 其密度和生物量基本降为零[15]。食物资源的减少, 使得白鹤被迫离开传统觅食的自然湿地, 前往稻田、藕塘等农业用地(亦是人工湿地)觅食[15—16]。
鄱阳湖周边有大面积稻田, 秋季收割后散落的稻谷吸引了大量白鹤、灰鹤(Grus grus)等越冬水鸟在此觅食[17]。上饶市余干县插旗州更是购买了约66.7hm2未收割稻田, 南昌市五星白鹤保护小区种植了约66.7hm2莲藕供白鹤等越冬水鸟食用[18]。近年来插旗州和五星白鹤保护小区已成为白鹤最主要的栖息地, 2020—2021年越冬季两个地点记录到的白鹤最大数量均超过全球总数量的50%, 这表明农业用地对鄱阳湖白鹤的保护发挥了至关重要的作用[19]。
笔者课题组的前期研究表明, 鄱阳湖沉水植被退化后白鹤的主要食物由苦草冬芽等转变为水稻(Oryza sativa)、莲藕(Nelumbo nucifera)和蓼子草(Polygonum criopolitanum)等[20—21], 然而目前对白鹤越冬期不同阶段的食物组成变化尚不清楚。白鹤主要食物的丰富度存在较大的年内波动, 这可能导致白鹤的食物组成也存在一定的年内变化。因此, 本研究采用DNA宏条形码方法研究越冬期不同阶段白鹤的食物组成及其变化。本研究有助于揭示白鹤的觅食策略, 为白鹤保护政策的制定提供科技支撑。
1 研究地区与研究方法 1.1 研究区概括鄱阳湖(115°48′—116°44′E, 28°25′—29°45′N)位于江西省北部, 是我国最大的淡水湖, 每年有超过40万只水鸟在此越冬, 包括白鹤、白头鹤(Grus monacha)、白枕鹤(Grus vipio)、鸿雁(Anser cygnoides)、白额雁(Anser albifrons)、东方白鹳(Ciconia boyciana)等珍稀濒危鸟类[22]。鄱阳湖是一个典型的季节性、吞吐型湖泊, 每年的4月至9月为丰水期, 湖泊面积超过3000 km2;10月至次年3月为枯水期, 湖泊面积缩减到不足1000 km2[23]。秋冬季节水位的下降, 使得大面积草洲、泥滩、浅水等生境出露, 为鸟类提供了丰富的食物及良好的栖息环境。此外, 湖区旁边大面积的稻田、藕塘等农业用地也成为了越冬水鸟潜在的觅食生境。
2 研究方法 2.1 白鹤粪便样品采集根据鄱阳湖洲滩植物的生长周期变化, 将越冬期分为三个阶段:越冬前期(10月—11月)、越冬中期(12月—1月)、越冬后期(2月—3月)[24]。笔者于2020/2021和2021/2022年越冬季前往鄱阳湖白鹤主要栖息地采集粪便样品, 采样点包括南昌市五星垦殖场、成新农场和方洲分场, 上饶市余干县插旗洲等白鹤的主要觅食地点, 采样生境包括稻田、藕塘、草洲等白鹤的主要觅食生境。为了完善课题组已构建的鄱阳湖植物DNA参考文库[21], 在采集粪便的过程中, 对其觅食地植物也进行采样。采集粪便样品时, 主要选取数量较多的白鹤群体进行采集, 本研究采样的群体大小平均为198(88—700)只个体。为了确定粪便样品的来源, 一般选取仅由白鹤组成的单一群体进行采样。采样过程中尽量采取新鲜的粪便, 此外, 为了避免交叉污染, 在同一个地点采样的过程中需要佩戴橡胶手套, 并用无菌镊子夹取粪便, 将其储存于10 mL的离心管中, 随后将样品置于液氮罐中冷冻, 到实验室后转移至-80℃冰箱保存。
觅食生境调查地点:1蚌湖、2沙湖、3朱市湖、4常湖池、5大湖池、6梅西湖、7南昌市朱港农场、8南昌市成新农场、9战备湖、10常湖、11白沙湖、12凤尾湖、13三泥湾、14南深湖、15余干县康山乡、16余干县插旗洲、17南昌市五星垦殖场;越冬前期采样点:18南昌市方洲分场、19王罗湖;越冬中期采样点:20南昌市五星垦殖场、21王罗湖、22余干县插旗洲;越冬后期采样点:23南昌市成新农场、24南昌市五星垦殖场
本研究共采集粪便样品45份, 其中, 越冬前期10份, 采样地点为王罗湖和南昌市方洲分场, 采样生境为草洲和稻田;越冬中期15份, 采样地点为王罗湖、南昌市五星垦殖场和余干县插旗洲, 采样生境为草洲和稻田;越冬后期20份, 采样地点为南昌市五星垦殖场和成新农场, 采样生境为稻田(图 1)。

|
| 图 1 白鹤粪便样品采样点和觅食生境调查点 Fig. 1 Fecal sampling sites and habitat survey sites of Siberian cranes |
于2021/2022年越冬季(11月—3月)每个月对鄱阳湖白鹤的生境利用情况进行调查。借助于SWAROVSKI(20—80 ×)单筒望远镜, 采用样点法和样线法相结合的方法调查主要分布区的白鹤数量, 并记录其生境类型(稻田、藕塘、自然湿地)。调查地点包括南昌市五星垦殖场、成新农场和朱港农场, 余干县插旗洲和康山乡, 鄱阳湖国家级自然保护区的常湖池、沙湖、蚌湖、朱市湖、梅西湖和大湖池, 南矶湿地国家级自然保护区的常湖、战备湖、南深湖、三泥湾、白沙湖和凤尾湖(图 1)。随后对不同越冬期白鹤数量进行汇总, 将不同生境白鹤数量除以总数量, 得到越冬期不同阶段白鹤在不同生境的数量占比。
2.3 鄱阳湖植物DNA参考文库完善在湿地植物中叶绿体trnL基因具有较高的扩增效率和物种区分能力[25], 因此本研究选用trnL基因作为分子标记。本课题组在前期研究中已构建鄱阳湖植物DNA trnL基因参考文库[21], 然而该文库并不完善, 缺少一些关键植物的序列信息。为完善该DNA参考文库, 本实验采用通用植物试剂盒(天根)提取植物的总DNA。使用引物c(5′-3′CGAAATCGGTAGACGCTACG)和h(5′-3′CCATTGAGTCTCTGCACCTATC)进行PCR扩增[26], 扩增体系为25 μL。反应条件如下:首先在PCR仪器上94℃预变性4 min, 之后进行33个循环(94 ℃变性30 s, 56 ℃退火30 s, 72 ℃延伸45 s), 最后72℃延伸10 min, 随后将其放置于4 ℃冰箱中保存。在1%琼脂糖凝胶上通过电泳检测PCR产物, 将扩增效果较好的PCR产物送到上海生工生物公司进行测序, 本实验采用Sanger测序法借助于ABI 3730xl DNA测序仪进行测序。经补充后的DNA参考文库共含有68种鄱阳湖湿地植物的trnL序列信息。
2.4 粪便样品DNA提取、扩增和测序基于采集的45份粪便样品, 采用改良的CTAB法分别提取每个样品的总DNA[21], 随后使用紫外分光光度计测量DNA的浓度和质量, 将质量较好的DNA保存在-20℃冰箱中备用。使用引物c和h扩增trnL基因。为了混样测序, 在引物前端添加了特异性标签(tag)用来区分样品。扩增体系、反应程序与DNA参考数据库构建过程相同。为了避免实验过程中的污染, 粪便样品DNA提取和PCR扩增在专门的实验室进行。此外, 考虑到PCR扩增过程的随机性, 每个样品设置3个重复, 并用1%的琼脂糖凝胶电泳检测扩增结果, 随后使用分光光度计测量其浓度, 筛选出扩增效果较好的样品在Illumina HiSeq 2500平台上测序, 所有样品测序均在诺禾致源生物公司(天津)进行。
2.5 宏条形码数据处理对于高通量测序得到的原始数据, 需要将其进行拼接、过滤、修剪和组装, 以得到高质量序列用于后续数据分析。首先将测序数据拆分成各个样品数据, 去除tag和引物序列, 引物和标签不能完全匹配的序列均去除;使用FLASH V1.2.7软件[27]将reads进行拼接, 获得原始数据, 然后将数据进行质量过滤;使用QIIME V1.7软件[28]对拼接数据进行过滤(将错误率>25%的reads过滤掉);然后使用Usearch软件对高质量数据进行去重复和OTU(可操作分类单元)聚类, 将相似度大于97%的序列合并为OTUs序列[29];最后用本地Blast软件将OTU序列和已构建的鄱阳湖植物DNA参考文库进行比对, 鉴定食物种类[21]。识别阈值为:序列覆盖度>97%, 序列一致性>97%, e-value < 1.0e-50。当OTU与某个分类单元相匹配时, 则将该OTU划分为该分类单位;当OTU与两个或者两个以上的分类单元都匹配时, 将该序列归为包含所有分类单元的更高分类级别。
2.6 食物组成变化分析将序列数据转换成各食物所占比例数据RRA(Relative read abundance), 用以衡量白鹤的食物组成[30]。公式如下:
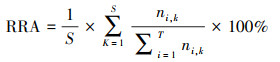
|
式中, ni, k表示食物种类i在样品k中的序列条数;T表示食物种类数;S表示样本数量。采用非度量多维尺度分析(NMDS)展示越冬期不同阶段白鹤的食物组成差异。采用相似性分析(ANOSIM)检验不同阶段的食物组成是否存在显著差异。使用R的vegan[31]包进行NMDS和ANOSIM分析, 距离算法采用Bray-Curtis距离。将水稻和莲藕归为农业用地食物, 其他植物归为自然生境食物, 计算农业用地和自然生境食物在白鹤越冬期不同阶段的占比。
为分析越冬期不同阶段白鹤的食物组成多样性, 笔者计算了不同阶段白鹤的Shannon-Wiener指数H′、食物均匀度指数J′和生态位宽度指数B。具体公式如下:

|
式中, Pi表示每种食物在所有食物中所占的比例;S表示食物种类数。
3 研究结果 3.1 白鹤的生境利用每个月调查的白鹤总数量超过1153±1330(SD)只, 因此能较好地代表白鹤的生境利用情况。在整个越冬期, 白鹤主要聚集在稻田、藕塘等农业用地觅食。其中在越冬前期, 白鹤主要聚集在藕塘觅食(93.06%);在越冬中期, 稻田中白鹤数量大幅增加(79.79%), 藕塘中的白鹤数量大幅减少(18.06%);在越冬后期, 在农业用地觅食的白鹤数量逐渐减少, 在自然湿地(草洲、水体和泥滩)觅食的白鹤数量逐渐增加(11.97%)(图 2)。

|
| 图 2 白鹤在越冬期不同阶段的生境利用情况 Fig. 2 The habitat use of Siberian cranes in different wintering periods |
白鹤在鄱阳湖共取食29种植物, 其中1种植物鉴定到科, 6种植物鉴定到属, 22种植物鉴定到种(图 3)。常见的食物有蓼子草、水稻、莲藕、蓼属(Polygonum spp.)、日本看麦娘(Alopecurus aequalis)、禾本科(Gramineae)、苔草(Carex spp.)、下江委陵菜(Potentilla limprichtii)、虉草(Phalaris arundinacea)、芡实(Euryale ferox)、南荻(Triarrhena lutarioriparia)。

|
| 图 3 白鹤在越冬期不同阶段的食物组成 Fig. 3 The food compositions of Siberian cranes in different wintering periods |
通过对越冬期不同阶段白鹤的食物组成及占比进行分析发现, 在越冬前期, 白鹤共取食13种植物, 其中2种植物鉴定到属, 11种植物鉴定到种, 莲藕(47.93±45.15)%和蓼属(47.63±43.43)%为主要食物。在越冬中期, 白鹤共取食14种植物, 其中2种植物鉴定到属, 12种植物鉴定到种, 蓼属(49.22±33.83)%、莲藕(26.20±32.58)%和水稻(19.08±35.18)%为主要食物。在越冬后期, 白鹤共取食28种植物, 其中1种植物鉴定到科, 5种植物鉴定到属, 22种植物鉴定到种, 蓼子草(23.94±16.75)%、水稻(20.82±17.09)%、莲藕(19.08±37.42)%和禾本科(13.07±15.18)%为主要食物。
NMDS分析的应力值(stress)为0.08, 表示结果可靠程度较高。不同阶段白鹤的食物组成形成不同的聚类, 表明不同阶段的食物组成差异较大, 尤其是越冬后期与其它两个时期的食物组成差异较大(图 4)。ANOSIM分析结果表明白鹤的食物组成在越冬前期与中期(P=0.034)、前期与后期(P=0.001)、以及中期与后期(P=0.001)之间均存在显著差异。

|
| 图 4 白鹤在越冬期不同阶段食物组成的非度量多维尺度分析(NMDS)结果 Fig. 4 The nonmetric multidimensional scaling (NMDS) plots of the diet compositions of Siberian crane in different wintering periods |
在越冬前期和越冬中期, 白鹤在自然生境和农业用地的食物占比差别不大。在越冬前期, 自然生境和农业用地的食物占比分别为50.64%和48.75%;在越冬中期, 两者的占比分别为43.35%和55.65%。然而, 在越冬后期, 白鹤主要取食自然生境食物(58.26%), 农业用地食物占比较小(39.90%)。
3.3 食性生态位宽度白鹤在越冬后期的食性生态位宽度最宽(4.72±0.78), 取食的食物种类最多, 均匀度指数(0.56±0.04)和多样性指数(1.86±0.12)也最大(表 1)。白鹤在越冬中期的生态位宽度(2.81±1.66)、均匀度指数(0.34±0.07)和多样性指数(0.94±0.28)均低于越冬后期, 但高于越冬前期。在越冬前期, 白鹤食性生态位宽度最窄(1.18±0.04), 均匀度指数(0.14±0.03)和多样性指数(0.37±0.10)也最低。
| 越冬季 Wintering period |
越冬前期 Early winter |
越冬中期 Mid-winter |
越冬后期 Late winter |
| 食物种类Type of foods | 13 | 14 | 28 |
| Shannon-Wiener多样性指数H′ Shannon-Wiener diversity H′ | 0.37±0.10 | 0.94±0.28 | 1.86±0.12 |
| 均匀度指数J′ Evenness index J′ | 0.14±0.03 | 0.34±0.07 | 0.56±0.04 |
| 生态位宽度指数B Dietary niche breadth B | 1.18±0.04 | 2.81±1.66 | 4.72±0.78 |
食物是动物维持生存和繁殖的必备条件, 动物通过觅食保证能量的摄入。对于候鸟来说, 越冬地的食物质量和丰富度不仅影响鸟类的数量和分布, 还会影响鸟类的身体状况、迁徙时间、繁殖成功率等[32—33]。以前白鹤在鄱阳湖主要分布在鄱阳湖保护区的蚌湖、沙湖和大湖池以及共青城的南湖等, 它们主要以沉水植物苦草冬芽为食[11—12]。近年来, 水质恶化、极端气候频发等导致鄱阳湖沉水植被退化严重, 这也使得白鹤的食物组成和觅食生境发生了改变[20—21]。例如, 2010年夏季鄱阳湖洪涝导致沉水植物生长受阻, 苦草冬芽短缺, 白鹤由自然湿地浅水区和泥滩转移至草洲寻找下江委陵菜块根和老鸦瓣(Tulipa edulis)球茎为食[11, 34]。2015年之后, 沉水植被退化导致数千只白鹤从自然湿地转移至稻田和藕塘觅食, 农业用地已成为白鹤的重要觅食生境[20—21]。笔者的调查结果同样表明稻田和藕塘是白鹤重要的觅食生境。
由于越冬水鸟在觅食的过程中, 会衡量觅食斑块的质量, 因而能量摄入的多少会影响水鸟对觅食地的选择[30]。白鹤的生境利用调查结果表明, 在越冬前期白鹤主要在五星白鹤保护小区的藕塘取食;在越冬中期随着藕塘资源的不断减少, 大量白鹤前往余干插旗洲的预留稻田觅食;在越冬后期, 五星白鹤小区藕塘和余干插旗洲稻田的食物逐渐减少, 大量白鹤开始前往撂荒稻田及自然生境草洲觅食。
白鹤的食物组成变化与生境变化基本一致。在越冬前期, 白鹤主要以莲藕和蓼属植物为食。虽然生境利用结果表明藕塘是越冬前期白鹤最主要的觅食生境, 但白鹤栖息的五星白鹤保护小区藕塘与鄱阳湖仅一堤之隔, 笔者观察到白鹤常在藕塘和鄱阳湖草洲轮番觅食。此时鄱阳湖水露滩出, 草洲长出了大片蓼子草[18], 这可能是蓼属植物也在白鹤食物中占比较高的原因。在越冬前期, 白鹤的食性生态位最窄, 食物多样性指数最低。在越冬中期, 由于鄱阳湖气温较低, 大多植物停止生长。此外, 随着藕塘食物资源的消耗, 在藕塘觅食的白鹤数量逐渐减少, 为了满足自身生存需求, 大量白鹤选择在余干插旗洲稻田中觅食。因此在越冬中期白鹤主要以莲藕、水稻以及自然生境蓼属植物为食。在越冬后期, 藕塘和稻田的食物逐渐减少, 此时随着气温回升, 许多滩涂植物开始陆续生长, 植物种类也在增加, 大量白鹤选择在自然生境草滩觅食, 因此在越冬后期, 自然湿地中食物的占比增加, 白鹤的食性生态位宽度最宽, 多样性指数最高。以上结果表明白鹤在越冬季的食物组成受外界环境中食物资源的种类和丰富度影响较大, 且随着时间变化在不断改变。与本研究结果类似, 升金湖越冬白头鹤在越冬前期和中期主要在食物资源丰富的稻田取食, 随着稻谷丰富度的逐渐减少, 白头鹤于越冬后期转移至自然湿地草洲取食[35]。
4.2 白鹤保护建议本研究结果表明, 在整个越冬期, 农业用地都是白鹤重要的觅食生境。近年来, 受自然湿地退化和土地利用方式改变的影响, 农业用地已成为很多水鸟的重要栖息地[4, 36]。鄱阳湖周边农业用地是白鹤、小天鹅、鸿雁等越冬水鸟的主要觅食地, 大量水鸟聚集在此取食[16, 19]。陈青等[17]在2020年越冬季对五星垦殖场藕塘觅食的白鹤数量调查中发现, 在此觅食的白鹤数量高达2, 900只。笔者在对余干插旗洲稻田中水鸟数量的调查中发现, 2021年12月15日在余干插旗洲稻田觅食的水鸟数量高达8, 273只, 其中白鹤1, 639只、小天鹅4, 600只、灰鹤1, 940只、白枕鹤94只。大量的水鸟聚集在一起觅食, 可能增加疾病传播的风险[34]。对此, 建议避免大片食物预留地, 而是分散预留小片农业用地, 减少大量水鸟聚集觅食的现象。
农业生产会使用大量农药, 而农药的使用可能会对稻田中觅食的水鸟产生危害[37]。已有研究表明, 大量农药的使用可能会对白鹤的健康构成威胁[38]。因此, 建议预留的稻田和藕塘限制农药尤其是高危农药的使用。此外, 鄱阳湖人为干扰也较为严重, 受人为干扰的影响, 鸟类的警戒行为比例会增加[39], 惊飞距离会变短[40]。五星白鹤保护小区是白鹤的重要觅食地, 每天有很多摄影爱好者和游客来此拍照, 人为干扰较为严重。因此, 在藕塘中觅食白鹤的警戒行为比例(25.02%)明显高于自然生境(11.94%)[18, 41]。因此, 建议加大食物预留地的巡护力度, 尤其是越冬前期和中期等白鹤利用程度较高时期的巡护力度, 减少人为干扰对白鹤觅食和安全造成的负面影响。
鉴于农业用地觅食对白鹤安全构成了潜在威胁, 因此开展鄱阳湖沉水植被恢复, 让白鹤回归自然湿地, 对于该物种的保护至关重要。近年来, 鄱阳湖苦草冬芽恢复工作正在逐步开展, 但尚未取得突破性进展, 加上全球气候变化引起的洪涝、干旱等极端气候频发, 给沉水植被恢复工作带来巨大挑战。鄱阳湖正在规划的水利枢纽工程也将深刻改变湖泊的水文节律, 这将进一步加大白鹤自然湿地食物资源的不确定性。因此, 可以预测在未来很长一段时间里白鹤仍将依赖农业用地觅食。此外, 本研究的分析结果表明蓼子草、下江委陵菜等洲滩食物也是白鹤的重要食物, 因此, 后期在开展鄱阳湖植被恢复及栖息地管理时, 也可将重点放在这些植物的扩繁上, 以增加自然湿地中白鹤的食物资源储量。
| [1] |
Wang X, Fox A D, Cong P H, Cao L. Food constraints explain the restricted distribution of wintering Lesser White-fronted Geese Anser erythropusin China. Ibis, 2013, 155(3): 576-592. DOI:10.1111/ibi.12039 |
| [2] |
张梅清, 赵运林, 徐正刚, 廖嘉欣, 李立, 黄田, 段酬苍. 洞庭湖越冬小天鹅食性研究. 西南林业大学学报, 2016, 36(4): 121-125. |
| [3] |
Fox A D, Cao L, Zhang Y, Barter M, Zhao M J, Meng F J, Wang S L. Declines in the tuber-feeding waterbird guild at Shengjin Lake National Nature Reserve, China-a barometer of submerged macrophyte collapse. Aquatic Conservation: Marine and Freshwater Ecosystems, 2011, 21(1): 82-91. DOI:10.1002/aqc.1154 |
| [4] |
Czech H A, Parsons K C. Agricultural wetlands and waterbirds: A review. Waterbirds, 2002, 25: 56-65. DOI:10.1675/1524-4695(2002)025[0056:BEOWTI]2.0.CO;2 |
| [5] |
Pompanon F, Deagle B E, Symondson W O C, Brown D S, Jarman S N, Taberlet P. Who is eating what: diet assessment using next generation sequencing. Molecular Ecology, 2012, 21(8): 1931-1950. DOI:10.1111/j.1365-294X.2011.05403.x |
| [6] |
BirdLife International. Species factsheet: Leucogeranus leucogeranus. Downloaded from http://datazone.birdlife.org/species/factsheet/siberian-crane-leucogeranus-leucogeranus on 22/12/2023.
|
| [7] |
国家林业和草原局, 农业农村部. 国家重点保护野生动物名录. 2021. https://www.forestry.gov.cn/c/www/lczc/10746.jhtml
|
| [8] |
温立嘉, 王榄华, 丁洪安, 李剑志, 郭玉民. 无人机调查白鹤种群数量和越冬分布. 生态学报, 2023, 43(18): 7693-7700. DOI:10.20103/j.stxb.202205041234 |
| [9] |
Yi K P, Zhang J J, Batbayar N, Higuchi H, Natsagdorj T, Bysykatova I P. Using tracking data to identify gaps in knowledge and conservation of the critically endangered Siberian crane (leucogeranus leucogeranus). Remote Sensing, 2022, 14(20): 5101. DOI:10.3390/rs14205101 |
| [10] |
刘洁, 黎敏灵, 刘继超, 吴志勇, 程彬彬, 孔凡前, 张微微. 取食生境对白鹤越冬行为的影响. 四川动物, 2023, 42(2): 152-160. |
| [11] |
Burnham J, Barzen J, Pidgeon A M, Sun B T, Wu J D, Liu G H, Jiang H X. Novel foraging by wintering Siberian Cranes Leucogeranus leucogeranus at China's Poyang Lake indicates broader changes in the ecosystem and raises new challenges for a critically endangered species. Bird Conservation International, 2017, 27(2): 204-223. DOI:10.1017/S0959270916000150 |
| [12] |
Li F S, Wu J D, Harris J, Burnham J. Number and distribution of cranes wintering at Poyang Lake, China during 2011-2012. Chinese Birds, 2012, 3(3): 180-190. DOI:10.5122/cbirds.2012.0027 |
| [13] |
胡振鹏, 林玉茹. 鄱阳湖水生植被30年演变及其驱动因素分析. 长江流域资源与环境, 2019, 28(8): 1947-1955. |
| [14] |
Li Y K, Zhong Y F, Shao R Q, Yan C, Jin J F, Shan J H, Li F S, Ji W, Bin L, Zhang X Y, Cao K Q, Shen J H. Modified hydrological regime from the Three Gorges Dam increases the risk of food shortages for wintering waterbirds in Poyang Lake. Global Ecology and Conservation, 2020, 24: e01286. DOI:10.1016/j.gecco.2020.e01286 |
| [15] |
Hou J J, Liu Y F, Fraser J D, Li L, Zhao B, Lan Z C, Jin J F, Liu G H, Dai N H, Wang W J. Drivers of a habitat shift by critically endangered Siberian cranes: evidence from long-term data. Ecology and Evolution, 2020, 10(20): 11055-11068. DOI:10.1002/ece3.6720 |
| [16] |
王文娟, 王榄华, 侯谨谨. 人工生境已成为鄱阳湖越冬白鹤的重要觅食地. 野生动物学报, 2019, 40(1): 133-137. |
| [17] |
陈青, 汪志如, 汤文超, 丁慧芳, 廖劲扬, 王亚芳, 王文娟. 鄱阳湖周边稻田对白鹤的承载力. 生态学报, 2022, 42(13): 5340-5347. DOI:10.5846/stxb202108102197 |
| [18] |
邵明勤, 张聪敏, 戴年华, 徐宁, 植毅进, 卢萍. 越冬小天鹅在鄱阳湖围垦区藕塘生境的时间分配与行为节律. 生态学杂志, 2018, 37(3): 817-822. |
| [19] |
底明晓, 翁晓东, 卢萍, 陆远鸿, 戴年华. 2020—2021年越冬季鄱阳湖两处垦殖场农田白鹤数量动态. 湿地科学与管理, 2022, 18(1): 23-26. |
| [20] |
侯谨谨, 王亚芳, 金斌松, 王榄华, 王文娟. 鄱阳湖越冬白鹤在农业用地的食物组成. 动物学杂志, 2019, 54(1): 15-21. |
| [21] |
Hou J J, Li L, Wang Y F, Wang W J, Zhan H Y, Dai N H, Lu P. Influences of submerged plant collapse on diet composition, breadth, and overlap among four crane species at Poyang Lake, China. Frontiers in Zoology, 2021, 18(1): 24. DOI:10.1186/s12983-021-00411-2 |
| [22] |
Li Y K, Qian F, Silbernagel J, Larson H. Community structure, abundance variation and population trends of waterbirds in relation to water level fluctuation in Poyang Lake. Global Ecology and Conservation, 2020, 24: e01286. DOI:10.1016/j.gecco.2020.e01286 |
| [23] |
Feng L, Hu C M, Chen X L, Cai X B, Tian L Q, Gan W X. Assessment of inundation changes of Poyang Lake using MODIS observations between 2000 and 2010. Remote Sensing of Environment, 2012, 121: 80-92. DOI:10.1016/j.rse.2012.01.014 |
| [24] |
Zhao M J, Cao L, Klaassen M, Zhang Y, Fox A D. Avoiding competition? site use, diet and foraging behaviours in two similarly sized geese wintering in China. Ardea, 2015, 103(1): 27-101. DOI:10.5253/arde.v103i1.a3 |
| [25] |
Yang Y Z, Zhan A B, Cao L, Meng F J, Xu W B. Selection of a marker gene to construct a reference library for wetland plants, and the application of metabarcoding to analyze the diet of wintering herbivorous waterbirds. PeerJ, 2016, 4: e2345. DOI:10.7717/peerj.2345 |
| [26] |
Taberlet P, Coissac E, Pompanon F, Gielly L, Miquel C, Valentini A, Vermat T, Corthier G, Brochmann C, Willerslev E. Power and limitations of the chloroplast trn L (UAA) intron for plant DNA barcoding. Nucleic Acids Research, 2007, 35(3): e14. DOI:10.1093/nar/gkl938 |
| [27] |
MagočT, Salzberg S L. FLASH: fast length adjustment of short reads to improve genome assemblies. Bioinformatics, 2011, 27(21): 2957-2963. DOI:10.1093/bioinformatics/btr507 |
| [28] |
Caporaso J G, Kuczynski J, Stombaugh J, Bittinger K, Bushman F D, Costello E K, Fierer N, Peña A G, Goodrich J K, Gordon J I, Huttley G A, Kelley S T, Knights D, Koenig J E, Ley R E, Lozupone C A, McDonald D, Muegge B D, Pirrung M, Reeder J, Sevinsky J R, Turnbaugh P J, Walters W A, Widmann J, Yatsunenko T, Zaneveld J, Knight R. QⅡME allows analysis of high-throughput community sequencing data. Nature Methods, 2010, 7(5): 335-336. DOI:10.1038/nmeth.f.303 |
| [29] |
Edgar R C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics, 2010, 26(19): 2460-2461. DOI:10.1093/bioinformatics/btq461 |
| [30] |
Deagle B E, Thomas A C, McInnes J C, Clarke L J, Vesterinen E J, Clare E L, Kartzinel T R, Eveson J P. Counting with DNA in metabarcoding studies: how should we convert sequence reads to dietary data?. Molecular Ecology, 2019, 28(2): 391-406. DOI:10.1111/mec.14734 |
| [31] |
Oksanen J, Blanchet F G, Kindt R, Legendre P, Minchin P R, O'hara R, Simpson G L, Solymos P, Stevens M H, Wagner H. Vegan: Community Ecology Package. 2013. http://CRAN.R-project.org/package=vegan
|
| [32] |
Holmes R T, Sherry T W. Thirty-year bird population trends in an unfragmented temperate deciduous forest: importance of habitat change. The Auk, 2001, 118(3): 589-609. DOI:10.1093/auk/118.3.589 |
| [33] |
Newton I. Population limitation in migrants. Ibis, 2004, 146(2): 197-226. DOI:10.1111/j.1474-919X.2004.00293.x |
| [34] |
Jia Y F, Jiao S W, Zhang Y M, Zhou Y, Lei G C, Liu G H. Diet shift and its impact on foraging behavior of Siberian crane (Grus leucogeranus) in Poyang Lake. PLoS One, 2013, 8(6): e65843. DOI:10.1371/journal.pone.0065843 |
| [35] |
Zheng M, Zhou L Z, Zhao N N, Xu W B. Effects of variation in food resources on foraging habitat use by wintering Hooded Cranes (Grus monacha). Avian Research, 2015, 6: 11. DOI:10.1186/s40657-015-0020-3 |
| [36] |
Huang C, Zhou K W, Huang Y J, Fan P F, Liu Y, Lee T M. Insights into the coexistence of birds and humans in cropland through meta-analyses of bird exclosure studies, crop loss mitigation experiments, and social surveys. PLoS Biology, 2023, 21(7): e3002166. DOI:10.1371/journal.pbio.3002166 |
| [37] |
Vallon M, Dietzen C, Laucht S, Ludwigs J D. Focal species candidates for pesticide risk assessment in European rice fields: a review. Integrated Environmental Assessment and Management, 2018, 14(5): 537-551. DOI:10.1002/ieam.4054 |
| [38] |
陈春玲, 周立志, 江浩, 吴治安, 邓磊, 汪维平, 杨荣景, 童康. 有机氯农药在东方白鹳和白鹤羽毛中的残留分析. 动物学研究, 2008, 29(2): 159-164. |
| [39] |
Luo J M, Wang Y J, Yang F, Liu Z J. Effects of human disturbance on the Hooded Crane (Grus monacha) at stopover sites in northeastern China. Chinese Birds, 2012, 3(3): 206-216. DOI:10.5122/cbirds.2012.0024 |
| [40] |
王彦平, 陈水华, 丁平. 惊飞距离——杭州常见鸟类对人为侵扰的适应性. 动物学研究, 2004, 25(3): 214-220. |
| [41] |
袁芳凯, 李言阔, 李凤山, 李佳, 缪泸君, 谢光勇. 年龄、集群、生境及天气对鄱阳湖白鹤越冬期日间行为模式的影响. 生态学报, 2014, 34(10): 2608-2616. DOI:10.5846/stxb201307291975 |