 2024, Vol. 44
2024, Vol. 44文章信息
- 马锦峰, 赵梦颖, 李雨欣, 谢婉玉, 纪洋, 冯兆忠, 冯彦房
- MA Jinfeng, ZHAO Mengying, LI Yuxin, XIE Wanyu, JI Yang, FENG Zhaozhong, FENG Yanfang
- 不同种类秸秆添加对稻田NH3挥发的影响机制
- The influence mechanism of different straw types on NH3 volatilization in paddy fields
- 生态学报. 2024, 44(15): 6696-6707
- Acta Ecologica Sinica. 2024, 44(15): 6696-6707
- http://dx.doi.org/10.20103/j.stxb.202305100975
-
文章历史
- 收稿日期: 2023-05-10
- 网络出版日期: 2024-05-24





2. 江苏省农业科学院农业资源与环境研究所, 农业农村部长江下游平原农业环境重点实验室, 南京 210014
2. Key Laboratory of Agro-Environment in Downstream of Yangtze Plain, Ministry of Agriculture and Rural Affairs, Institute of Agricultural Resources and Environment, Jiangsu Academy of Agricultural Sciences, Nanjing 210014, China
水稻是世界上三大粮食作物之一, 全球约50%的人口以水稻为主食[1]。随着人口不断增长, 对水稻等主要粮食作物的需求也随之增加, 而氮肥投入的大幅增加提高了水稻的产量, 对保障粮食供应与安全发挥了重要作用。全球农田氮肥近几十年用量与20世纪60年代相比增加了7倍左右[2]。氮肥投入的增加虽然可提高水稻产量, 但过量使用增加了农田氮素损失。稻田NH3挥发约占施氮量的10%—40%[3—4]。在NH3挥发损失中90%的损失将直接进入大气, 大量氨的传输和沉积导致整个生态系统的氮超负荷, 进而引起一系列环境问题, 如土壤酸化[5]、雾霾[6]、水体富营养化等[7]。稻田NH3挥发过程是发生在土-水-气界面, 与稻田NH3挥发相关的化学平衡为NH4+(代换性)经过一系列转换最终转换成为NH3(气相)的过程[8], 也是田面水中的铵态氮转换为气态的NH3后经水面挥发到空气中的过程, 与田面水铵态氮浓度、施氮量和田面水pH等密切相关[9], 也受稻田土壤、水稻植株、水分、环境因素以及各种田间管理措施的影响[10]。
在诸多田间管理措施中, 秸秆还田不仅可以培肥地力、固碳增产[11—12], 还可以改变土壤中氮固定相关微生物的多样性和丰度[13—14], 进而影响氮素循环以及NH3挥发作用强度[15—17]。当前有关秸秆还田对农田NH3挥发影响的报道尚未达成一致结论。有研究表明, 秸秆还田增加农田土壤NH3挥发损失。汪军等[18]研究认为, 秸秆还田相比单施氮肥处理稻田土壤NH3挥发损失增加了19.8%—20.6%。张刚等[16]研究认为, 秸秆全量还田与不还田相比稻田土壤NH3挥发损失增加18.2%。也有研究表明, 秸秆还田降低了农田土壤NH3挥发损失, 并增加氮肥利用率[19—20]。Zhang等[21]研究认为, 秸秆全量还田和半量还田与无秸秆施加相比分别降低了稻田土壤年积累NH3挥发量35.1%和16.1%, 其原因可能为秸秆还田促进了土壤无机氮的固定。在不同种类秸秆还田试验中, 有研究表明, 相同秸秆还田量下玉米秸秆还田处理的稻田土壤氨挥发最大日平均值显著高于小麦秸秆还田处理的氨挥发最大日平均值[22];Xu等[23]和Sun等[24]研究发现小麦秸秆还田较相同添加量下水稻秸秆还田显著增加稻田土壤NH3挥发量。以上研究均为不同条件下单一秸秆种类施用或不同秸秆种类同一添加量施用对稻田土壤NH3挥发的影响, 关于不同秸秆种类及不同添加量对稻田土壤NH3挥发的影响差异及机制研究较少。
基于以上国内外研究现状, 本研究科学假设是, 在相同环境条件及施肥措施下, 不同种类作物秸秆和不同添加量通过影响稻田土壤中与NH3挥发相关的生物因子和非生物因子, 促使不同种类秸秆和添加量下稻田土壤NH3挥发产生差异。针对上述假设, 本研究以南粳46水稻品种为材料, 太湖地区典型单季稻田原状土(江苏宜兴)为研究对象, 设置温室盆栽试验, 综合利用连续气流封闭法[25]和分子生物学技术, 同步监测三种作物秸秆(RS:水稻;WS:小麦;MS:玉米)两种添加量(0.5%、0.8%)下水稻各生长期NH3挥发通量、土壤理化因子及水稻产量, 并将上述因子与氮循环相关的功能微生物进行耦合, 以揭示稻田土壤NH3挥发对不同种类作物秸秆和不同添加量的响应机制, 并筛选出常规秸秆种类还田下稻田氨减排效果最佳的秸秆还田种类及施用量, 从而为农田生态系统氨减排提供理论依据和数据支撑。
1 材料与方法 1.1 盆栽试验设计水稻盆栽试验在江苏省农业科学院现代化温室开展, 于2020年6月移栽, 同年11月收获。供试土壤为单季稻田湖白土, 由0—20、20—40和40—60cm三个深度的土壤各自混合均匀后得到, 共称取35kg, 依次装入半径15cm、有效高度50cm的土柱内。供试土壤原始基本理化性质为:有机质29.2g/kg, pH6.38 (土水比为1 ∶ 2.5), CEC 22.61cmol/kg, TN 1.72g/kg, 有效磷23.09g/kg, 速效钾159.28mg/kg。试验共设置3种秸秆施用, 分别是水稻秸秆RS、小麦秸秆WS和玉米秸秆MS, 施用秸秆基本性质如图 1所示。每种秸秆的添加量分别为0.5%(0.5%每克干土, 相当于田间施用量为5t/hm2)和0.8%(0.8%每克干土, 相当于田间施用量为8t/hm2), 采用尼龙网袋法处理秸秆还田, 即将秸秆粉碎至1—2cm后装入350目的尼龙网袋(15cm×20cm)中密封, 根据不同施用量施用至盆栽中间、距离土面5—10cm处。试验共设置7个处理:对照(CK)、水稻秸秆0.5%施用量(RS-0.5)、水稻秸秆0.8%施用量(RS-0.8)、小麦秸秆0.5%施用量(WS-0.5)、小麦秸秆0.8%施用量(WS-0.8)、玉米秸秆0.5%施用量(MS-0.5)、玉米秸秆0.8%施用量(MS-0.8), 每个处理设置3个重复。

|
| 图 1 供试秸秆基本性质 Fig. 1 Basic properties of tested straw types RS:水稻秸秆;WS:小麦秸秆:MS:玉米秸秆;TN:总氮;TP:总磷;TK:总钾;TC:总碳;C/N:碳氮比;图中不同字母表示处理间差异显著(P < 0.05) |
综合当地水稻种植管理措施, 本试验盆栽施肥管理措施为:整个生育期氮肥施入量为240kg/hm2, 按照2 ∶ 1 ∶ 1的比例于基肥、分蘖肥和穗肥分别施入, 三个肥期的时间为2020年6月30日、7月15日和8月13日。磷肥(P2O5)和钾肥(K2O)作为基肥一次性施用, 施用量分别为96kg/hm2和192kg/hm2。水分管理措施为:间隙灌溉模式, 分为四个阶段, 即前期淹水, 土面保持有水层(6月30日—7月29日);中期烤田, 通常土壤含水量达到50%—60%田间最大持水量时结束烤田(7月30日—8月12日);干湿交替, 约每周灌溉一次, 期间土面出现落干状态(8月12日—10月14日);排水落干, 土面落干至水稻收获(10月15日—11月9日)。
1.2 样品采集与测定分别于水稻生长的基肥期、蘖肥期和穗肥期(施肥后1—7d)连续测定田面水pH(pH 3310 SET 2;Germany), 并采集田面水水样, 过滤后通过SKALAR SAN++ SYSTEM流动分析仪(Skalar, the Netherlands)测定田面水NH4+-N和NO3--N含量。同时在水稻生长的三个肥期(基肥期、蘖肥期和穗肥期)原位采集0—10cm土壤样品, 于-20℃条件下保存用于土壤微生物分析。
本试验采用封闭式酸吸收法[26]计算每日NH3挥发通量, 将溴甲酚、甲基红指示剂、乙醇和80mL 2.0%硼酸混合物用作NH3吸收剂, 在高度20cm和内径15cm的有机玻璃瓶中捕获盆栽中挥发的NH3于广口瓶中。分别于水稻生长的基肥期、蘖肥期和穗肥期施肥后7d内于每天早上8:00开始采集。
NH3挥发速率计算公式为:
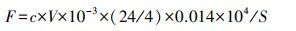
|
式中, F为NH3挥发速率, kg hm-2 d-1;c为硫酸的标准浓度, mol/L;V为滴定用去的标准酸的体积, mL;24/4为24h与日氨挥发收集时间4h的比值;0.014表示氮原子的相对原子质量, kg/mol;S为捕获装置的横切面积, m2;104表示面积转换系数。
NH3挥发肥期排放总量=测定时期内每次收集的NH3挥发通量之和。
1.3 土壤微生物碳氮及微生物群落丰度分析分别于水稻生长的三个肥期(基肥期、蘖肥期和穗肥期)采集CK处理和0.8%施用量秸秆处理表土, 一部分使用Fast DNA SPIN KIT试剂盒(MP Biomedicals, Eschwege, Germany)提取DNA, 利用定量PCR技术分析土壤氨氧化古菌(AOA)、氨氧化细菌(AOB)和细菌的群落丰度[27];一部分采用氯仿熏蒸浸提-TOC分析仪[28]测定土壤微生物碳(MBC)和微生物氮(MBN)含量。
同时称取各肥期表土0.5g, 用Fast DNA SPIN KIT试剂盒(MP Biomedicals, Eschwege, Germany)。根据说明书操作步骤提取总DNA, DNA纯度和浓度检测方法均为NanoDrop2000, DNA完整性检测方法为琼脂糖凝胶电泳。将样品在冰上融化后, 充分混匀并离心, 取适量样品进行检测。将提取得到的微生物总DNA在电压5V/cm条件下通过1% 琼脂糖凝胶电泳检测, 检测时间为20min, 稀释10倍后进行下游实验。
使用Lighe Cycler ST300(Roche Diagnostics, Germany)定量PCR仪分别对成功提取的AOA、AOB和细菌16S rRNA基因的特定DNA片段进行扩增。AOA的扩增引物为Arch-amoAF/Arch-amoAR, AOB的扩增引物为amoA1F/amoA2R[29], 16S rRNA基因的扩增引物为Ba519f和Ba907r[30]。使用CFX96C100TM热循环器(Bio-Rad, Hercules, CA)进行扩增。AOA和AOB的扩增反应程序为:95℃变性3min;然后35个循环为95℃变性45s, 53℃(AOA)或55℃(AOB)退火1min, 72℃延伸1min;最后72℃处理7min[31]。细菌16S rRNA的扩增反应程序为:95℃预变形5min, 94℃变性30s, 55℃退火30s, 72℃延伸30s, 30个循环[32]。AOA的扩增效率为96.50%, R2为0.9977;AOB的扩增效率为104.12%, R2为0.9938;16S rRNA的扩增效率为98.48%, R2为0.9984。标准曲线制备流程为10倍梯度稀释构建好的各质粒, 90μL稀释液+10μL质粒, 一般做4—6个点, 通过预实验分别选取标准品的10-3—10-8用于制备标准曲线。
1.4 数据处理与分析采用Microsoft Excel 2019进行数据统计, 采用SPSS 26.0进行单因素方差分析(one-way ANOVA)、LSD法进行多重比较、Pearson法在0.001、0.01和0.05的显著水平下双尾检验进行相关性分析, 采用Origin2021绘制图形, 采用Amos24.0构建结构方程模型(SEM模型)。
2 结果分析 2.1 稻田土壤不同施肥期NH3挥发通量如图 2所示, 不同施肥期相比, 各处理基肥期和蘖肥期的NH3挥发通量均显著高于穗肥期。在基肥期, 施加秸秆处理的NH3挥发通量均显著高于CK处理, 在施肥后3—4天出现峰值, 且各个处理峰值出现时间存在差异。在蘖肥期, 各处理NH3挥发通量在施肥后第1—2天出现峰值, 随后下降, 除WS-0.8和MS-0.5处理外施加秸秆处理NH3挥发通量均显著低于CK处理, 其中0.8%添加量下, RS和MS处理的NH3挥发通量显著低于WS处理下。与基肥期和蘖肥期不同, 穗肥期各处理的NH3挥发通量较低且趋于稳定。

|
| 图 2 水稻不同施肥期NH3挥发通量 Fig. 2 Variation in NH3 volatilization fluxes in different fertilization periods CK:空白对照;RS-0.5:水稻秸秆0.5%施用量;RS-0.8:水稻秸秆0.8%施用量;WS-0.5:小麦秸秆0.5%施用量;WS-0.8:小麦秸秆0.8%施用量;MS-0.5:玉米秸秆0.5%施用量;MS-0.8:玉米秸秆0.8%施用量 |
如图 3所示, 秸秆种类对NH3挥发产生显著影响(P < 0.05), 添加量对NH3挥发影响不显著(P>0.05), 秸秆种类与添加量交互作用下对NH3挥发产生显著影响(P < 0.05)。整个生长季NH3挥发主要集中在基肥期和蘖肥期, 而穗肥期的NH3挥发总量相对较少。在基肥期, 与CK相比, 施加秸秆处理NH3挥发总量增幅8.2%—73.0%, 其中RS-0.5、WS-0.5、WS-0.8、MS-0.5以及MS-0.8处理的NH3挥发总量与CK处理相比均有显著性差异(P < 0.05)。在蘖肥期, 与CK相比, 施加秸秆处理NH3挥发总量的变化幅度在-55.9%—19.6%, 其中RS-0.8、WS-0.5、WS-0.8以及MS-0.8处理的NH3挥发总量与CK处理相比均有显著性差异(P < 0.05)。在穗肥期, 除MS-0.8外, 其余施加秸秆处理的NH3挥发总量与CK相比降低20.1%—33.3%, 且各个处理间无显著性差异(P>0.05)。

|
| 图 3 水稻不同施肥期NH3累积排放量 Fig. 3 Cumulative emissions of NH3 in different fertilization periods 不同大写字母表示同一时期不同处理间差异显著(P < 0.05), 不同小写字母表示同一处理不同时期的差异显著(P < 0.05), *表示秸秆还田处理与CK差异显著(P < 0.05) |
整个生育期的NH3挥发总量如表 1所示, 与CK相比, WS-0.8、MS-0.5处理分别显著增加稻田土壤NH3挥发总量3.5和2.8kg N/hm2, 增幅分别为27.3%和21.1%;然而, RS-0.8处理显著降低NH3挥发总量4.2kg N/hm2, 降幅为33.2%(P < 0.05)。不同秸秆种类相比, 0.5%添加量下, WS处理下的NH3挥发总量最低;0.8%添加量下, RS处理下的NH3挥发总量最低。不同添用量相比, RS和MS处理在0.8%添加量下的NH3挥发总量显著低于0.5%添加量, 降低幅度为31.2%—32.8%(P < 0.05);WS处理呈现相反规律, 0.8%添加量较0.5%添加量显著增加NH3挥发总量28.1%(P < 0.05)。
| 处理 Treatments |
累计NH3挥发 Cumulative NH3 volatilization/(kg/hm2) |
水稻产量 Rice yield/(t/hm2) |
单位产量NH3挥发量 NH3 volatilization per yield/(g/kg) |
田面水pH Field surface water pH |
||
| 基肥期 Basal fertilizer |
蘖肥期 Tillering fertilizer |
穗肥期 Panicle fertilizer |
||||
| CK | 12.95±0.14b | 13.78±0.56b | 0.94±0.10ab | 7.72±0.03a | 7.76±0.02a | 7.21±0.01a |
| RS-0.5 | 12.88±0.03b | 17.13±1.51a | 0.76±0.06abc | 7.53±0.02b | 7.60±0.01b | 7.19±0.02a |
| RS-0.8 | 8.65±0.14c | 16.93±0.43a | 0.51±0.09d | 7.54±0.02b | 7.62±0.01b | 7.18±0.01a |
| WS-0.5 | 12.86±0.19b | 17.61±1.25a | 0.74±0.15bc | 7.59±0.03ab | 7.56±0.01b | 7.22±0.02a |
| WS-0.8 | 16.48±0.21a | 17.35±1.20a | 0.96±0.18a | 7.61±0.03ab | 7.52±0.01b | 7.17±0.02a |
| MS-0.5 | 15.68±0.10a | 17.53±0.28a | 0.90±0.06ab | 7.61±0.02ab | 7.62±0.01b | 7.16±0.02a |
| MS-0.8 | 10.79±0.06b | 15.67±0.63ab | 0.69±0.03cd | 7.58±0.02ab | 7.58±0.01b | 7.18±0.02a |
| 同列不同字母表示不同处理间差异显著(P < 0.05);CK:空白对照;RS-0.5:水稻秸秆0.5%施用量;RS-0.8:水稻秸秆0.8%施用量;WS-0.5:小麦秸秆0.5%施用量;WS-0.8:小麦秸秆0.8%施用量;MS-0.5:玉米秸秆0.5%施用量;MS-0.8:玉米秸秆0.8%施用量 | ||||||
由表 1可知, 与CK相比, 施加秸秆均显著提高水稻产量, 增产幅度为13.7%—27.8%(P < 0.05)。不同秸秆种类处理与CK相比, WS秸秆处理增产比例最高, 高低两种添加量下分别显著增产25.9%和27.8%(P < 0.05);其次RS秸秆处理, 分别显著增产22.9%和24.4%(P < 0.05);MS秸秆增产比例最低, 分别增产13.7%(P>0.05)和27.2%(P < 0.05), 其中除MS-0.8处理外, 其余处理的水稻产量与CK相比呈显著性差异(P < 0.05)。相同秸秆种类下0.5%添加量较0.8%添加量增加水稻产量1.2%—3.5%, 其中仅MS-0.5与MS-0.8之间无显著性差异(P>0.05)。
与CK相比, 施加秸秆处理下的单位产量NH3挥发量变化幅度介于-45.5%—1.9%, 其中, 高添加量下RS和MS处理较CK处理显著降低单位产量NH3挥发量(P < 0.05)。与0.5%添加量相比, 0.8%添加量下RS和MS处理的单位产量NH3挥发量分别降低23.0%和32.2%, 且呈显著性差异(P < 0.05), 而WS处理呈现相反趋势, 0.8%添加量显著增加单位产量NH3挥发量29.9%(P < 0.05)。
2.4 水稻生长期间田面水pH动态变化如图 4所示, 整个生长季来看, 秸秆施用处理田面水pH值均低于CK处理。在基肥期, 各个处理间pH值在施肥后3—4天出现峰值, 且各个处理之间峰值出现的时间存在差异, 其中RS-0.5、RS-0.8处理的pH值显著低于CK处理(P < 0.05)。在蘖肥期, 各个处理pH值均呈现下降趋势, 施加秸秆处理的pH值均显著低于CK处理(P < 0.05), 其中在0.8%添加量下, RS处理pH值高于WS和MS处理(P>0.05)。在穗肥期, 各处理均在施肥后的第2天趋于稳定, 且各处理间pH值均无显著性差异(P>0.05)。

|
| 图 4 水稻生长期田面水pH季节变化 Fig. 4 Seasonal variations in pH concentrations in floodwater during rice growing season |
整个生长季来看, 三个肥期各个处理田面水TN浓度均呈下降趋势, 且田面水NH4+-N浓度整体高于田面水NO3--N浓度(图 5)。整个生长季来看, 与CK相比, 秸秆施用处理下田面水TN浓度降低8.8%—21.1%(图 5)。

|
| 图 5 水稻生长期田面水NH4+-N、NO3--N和TN浓度季节变化 Fig. 5 Seasonal variations in NH4+-N、NO3--N and TN concentrations in floodwater during rice growing season |
在基肥期, 各个处理田面水NH4+-N浓度在施肥后2—3天达到峰值;在蘖肥期和穗肥期, 田面水NH4+-N浓度变化趋势基本相似, 均呈下降趋势。整个生长季来看, 与CK相比, 除WS-0.8%处理田面水NH4+-N浓度降低10.5%外, 各秸秆处理的田面水NH4+-N浓度均增幅1.9%—7.1%, 其中RS和MS处理田面水NH4+-N浓度均随秸秆添加量的增加而增加, 而WS处理田面水NH4+-N浓度则呈现相反规律(图 5)。
在基肥期, 各处理田面水NO3--N浓度均在施肥后4—5天达到峰值, 且各处理峰值出现时间存在差异;在蘖肥期, 各个处理田面水NO3--N浓度均呈上升趋势;在穗肥期, 虽然NO3--N浓度变化趋势波动较大, 但是各个处理均第2天出现峰值后呈现下降趋势。整个生长季来看, 与CK相比, 各个处理田面水NO3--N浓度的变化幅度-25.8%—44.1%, 其中RS、WS和MS处理下田面水NO3--N浓度均随着秸秆添加量的增加而降低(图 5)。
2.6 水稻生育期土壤功能微生物丰度的动态变化荧光定量PCR检测结果表明, 本试验中稻田土壤氨氧化古菌(AOA)丰度较氨氧化细菌(AOB)高2个数量级, 可知本试验中水稻土壤环境更利于AOA生长。整个生育期来看, 施加秸秆处理AOA、AOB和细菌群落丰度均随生长期呈现先升高后降低的趋势, 而CK处理为逐渐升高的趋势。
与CK相比, 施加秸秆处理下AOA群落丰度升高18.7%—85.1%(图 6);其中, RS-0.8处理下AOA群落丰度显著高于CK和MS-0.8处理下AOA群落丰度(P < 0.05);RS-0.8处理下不同水稻生育期AOA群落丰度变化幅度较大, 且呈显著性差异(P < 0.05)。与CK相比, 施加秸秆处理下AOB群落丰度升高26.4%—166.8%(图 6);其中, RS-0.8处理下AOB群落丰度显著高于CK和WS-0.8处理下AOB群落丰度(P < 0.05);RS-0.8处理下不同水稻生育期AOB群落丰度变化幅度较大, 且呈显著性差异(P < 0.05)。

|
| 图 6 水稻生长期土壤微生物丰度以及微生物碳、氮含量变化 Fig. 6 Changes in soil microbial abundance and microbial carbon and nitrogen contents during rice growth period |
与CK相比, 施加秸秆处理下细菌16S rRNA基因拷贝数增幅介于9.0%—111.6%(图 6);秸秆施加处理之间无显著差异;不同生育时期比较, 蘖肥期RS-0.8处理细菌丰度显著高于其他处理(P < 0.05);而其他两个肥期(基肥期、穗肥期)各个处理间无显著性差异(P>0.05)。
2.7 水稻生育期土壤微生物碳氮的动态变化整个生育期来看, 所有处理土壤微生物量碳(MBC)含量均随水稻生长呈现下降趋势(图 6), 且各个处理间无显著性差异(P>0.05)。与CK相比, 施加秸秆处理下MBC含量变化幅度介于-10.5%—7.1%;相较于低添加量处理, 高添加量处理下MBC含量增加4.4%—13.9%, 其中CK、RS、WS处理的基肥期MBC含量均显著高于其他两个生育期(P < 0.05)。
整个生育期来看, 所有处理下土壤微生物量氮(MBN)含量均随着生长期的变化并无显著性差异(P>0.05)。各处理在整个生长季的MBN含量并无显著性差异(P>0.05), 与CK相比, 施加秸秆处理下MBN含量变化幅度介于-32.8%—17.0%;相较于低添加量处理, 高添加量处理下MBN含量增加4.7%—74.1%(图 6)。
各处理土壤微生物量碳氮含量比值(MBC/MBN)随水稻生长呈下降趋势(图 6), 其中基肥期比值显著高于其他两个时期(P < 0.05)。与CK相比, 施加秸秆处理下MBC/MBN比值变化幅度介于-29.2%—10.4%;相较于低添加量处理, 高添加量处理下MBC/MBN比值变化幅度介于-31.3%—2.5%。
2.8 稻田土壤NH3挥发与土壤生物和非生物因子的相关性分析由图 7可知, 从整个生长季来看, 不同施肥期稻季NH3挥发累积量与土壤MBC含量、MBC/MBN、田面水pH、NH4+-N和TN浓度在P < 0.01水平上极显著正相关, 与田面水NO3--N浓度在P < 0.05水平上显著正相关;土壤MBC、MBC/MBN、田面水pH、田面水TN和NH4+-N浓度指标两两交互下在P < 0.01水平上均呈显著正相关性;相关性数据表明, 田面水pH(0.72)是稻田土壤NH3挥发最主要的影响因素。

|
| 图 7 稻田土壤NH3挥发和土壤理化因子的相关性 Fig. 7 Correlation between soil NH3 volatilization and soil physicochemical factors 不同颜色表示相关性的大小;*表示在P < 0.05水平上显著相关;* *表示在P < 0.01水平上显著相关;* * *表示在P < 0.001水平上显著相关;NH3:稻田土壤氨挥发量;NO3--N:田面水硝态氮浓度;NH4+-N:田面水铵态氮浓度;TN:田面水总氮浓度;pH:田面水pH;MBC:土壤微生物量碳含量;MBN:土壤微生物量氮含量;MBC/MBN:土壤微生物量碳与土壤微生物量氮的比值 |
图 8为以稻田NH3挥发为中心、水稻三个施肥期土壤理化和微生物为影响因素构建的结构方程模型(图 8), 通过建立路径分析模型研究各生物因子和非生物因子对稻田NH3挥发量的直接或间接影响(图 8)。由于稻田土壤NH3挥发过程的复杂性, 不同土壤因子可能产生协同或抑制作用, 因此采用结构方程模型可以使不同土壤因子对稻田土壤NH3挥发的影响方向和强度更加直观[33]。由图 8所示, 稻田土壤MBC含量(0.376)、田面水pH(-0.592)、田面水NH4+-N(1.790)和TN(-1.262)浓度对NH3挥发均具有极显著影响(P < 0.001), 细菌16S rRNA对稻田NH3挥发无显著直接影响。依于结构方程模型模型, 以上土壤因子对稻田NH3挥发影响的总效应分别为:pH(-0.796)、TN(-0.784)、NH4+-N(-0.080)、细菌16S rRNA(0.137)、MBC(0.336), 表明田面水pH、田面水TN以及土壤MBC含量是导致不同秸秆种类还田下稻田土壤NH3挥发产生差异的主要影响因素。

|
| 图 8 稻田土壤理化因子和微生物丰度对NH3挥发影响的结构方程模型和各因子的直接、间接和总效应 Fig. 8 Structural equation model of influence of soil Physicochemical factors and microbial abundance on NH3 volatilization and direct, indirect and total effects of each factor 路径旁边显示标准化的路径系数;实箭头和虚箭头分别表示正相关关系和负相关关系;*代表P < 0.05, * * *代表P < 0.001; Chi-square代表卡方值,DF代表自由度,Chi/DF GFI代表拟合优度指数,AGFI代表修正拟合优度指数 |
本试验中水稻蘖肥期土壤NH3挥发量最高。有研究表明, 施肥后一周内为稻田NH3挥发的主要时期, NH3挥发排放峰值一般出现在施肥后1—3d[34]。本试验中稻季NH3挥发排放峰值主要集中在施肥后2—3d, 与前人研究一致。由于蘖肥期光照强、温度高, 相较于其他两个肥期(基肥期、穗肥期)更有利于稻田NH3挥发, 因此本试验稻田NH3挥发主要集中在蘖肥期;而穗肥期相较于其他两个肥期(基肥期、蘖肥期), 经历干湿交替和排水落干两个时期, 不利于NH3挥发, 因此穗肥期的NH3挥发排放量最低[35], 这与前人研究结论基本一致。
不同秸秆种类还田对稻田土壤NH3挥发的影响存在差异。常菲等[36]和Cao等[37]发现在高添加量下, 玉米秸秆还田处理稻田土壤NH3挥发量低于小麦秸秆还田处理NH3挥发量, 本试验发现类似结果, 0.8%添加量下玉米秸秆处理的NH3挥发总量低于小麦秸秆处理34.5%。此外, 也有研究发现小麦秸秆还田的稻田NH3挥发量高于水稻秸秆还田NH3挥发量[23—24], 本试验中, 0.8%添加量下, 水稻秸秆施用较小麦秸秆显著降低稻田NH3挥发总量90.4%, 且与CK相比也显著降低稻田NH3挥发, 而小麦和玉米添加并未显著降低氨挥发总量, 原因可能是由于不同秸秆种类碳氮比值和养分含量对土壤MBC与MBC/MBN的影响存在差异[38]。李伟等[39]研究发现, 稻田土壤MBC和NH3挥发呈显著正相关, 其原因可能是MBC加速土壤有机氮的分解, 导致NH3挥发底物增加, 进而促进NH3排放[40]。本研究中, 结构方程模型显示稻田田面水pH对土壤NH3挥发呈现显著负效应, 这与Sun等[3]研究结果相同, 原因可能是随着田面水pH升高, 土壤氮素转化中的氨氧化反应得到促进。Bru等[41]和Yao等[42]提出农田土壤pH值与AOA、AOB群落丰度高度相关, Wu等[43]也提出水稻秸秆还田可以增加土壤AOA、AOB丰度, 是由于水稻矿化速率低, 可持续为土壤供应氮源[44], 促进土壤AOA、AOB群落丰度增长[45], 而NH3为AOA、AOB可消耗底物[46—47], 随着土壤AOA、AOB群落丰度的增加, 稻田NH3排放量降低[3]。本试验中, 与小麦和玉米秸秆处理相比, 水稻秸秆处理(RS)降低蘖肥期土壤MBC含量(P>0.05), 减少有机氮分解;增加田面水pH值(P>0.05), 促进氨氧化反应;其中RS-0.8处理显著增加土壤AOA、AOB群落丰度(P < 0.05), 促进NH3消耗, 综合效应导致水稻秸秆高添加量下土壤NH3挥发量最低。
不同秸秆添加量对稻田土壤NH3挥发的影响亦存在差异。Zhang等[21]研究认为, 秸秆全量还田和半量还田与无秸秆施加相比分别降低了农田土壤NH3挥发量35.1%和16.1%, 赵政鑫等[15]基于Meta分析发现当秸秆施用量高于6000kg/hm2时将降低农田土壤NH3挥发。本试验发现类似结果, 0.8%添加量下, 水稻和玉米秸秆, 处理的NH3挥发量显著低于0.5%秸秆添加量处理, 原因可能是当秸秆施用量较高时, 可促进微生物活性, 提高了土壤有机氮的固持与土壤TN储量, 降低田面水TN[36—37], 从而抑制稻田NH3挥发。本试验发现不同种类秸秆的高添加量相较于低添加量显著降低田面水TN含量2.3%—13.5%, 进而导致高添加量下土壤NH3挥发降低, 同上述结果一致。水稻秸秆高添加量处理相较于其他处理显著增加土壤AOA、AOB群落丰度并显著降低田面水TN含量。基于上述因素, 水稻秸秆高添加量处理显著降低稻田土壤NH3累计排放量。
4 结论田面水pH、TN浓度、土壤MBC含量、土壤AOA和AOB群落丰度是影响稻田土壤NH3挥发的主要因素;水稻秸秆高添加量(RS-0.8)处理相较于其他处理显著增加土壤AOA、AOB群落丰度促进NH3消耗, 显著降低田面水TN含量, 综合效应下显著减少稻田土壤NH3挥发量33.2%, 并显著增加水稻产量22.9%。综合稻田土壤NH3挥发量和水稻产量, 秸秆施用水稻种植体系中水稻(RS)秸秆以0.8%施用量模式下氨减排效果最佳。
| [1] |
Durand-Morat A, Nalley L L, Thoma G. The implications of red rice on food security. Global Food Security, 2018, 18: 62-75. DOI:10.1016/j.gfs.2018.08.004 |
| [2] |
Luo Y L, Li Q Q, Shen J, Wang C Q, Li B, Yuan S, Zhao B, Li H X, Zhao J W, Guo L K, Li S, He Y T. Effects of agricultural land use change on organic carbon and its labile fractions in the soil profile in an urban agricultural area. Land Degradation & Development, 2019, 30(15): 1875-1885. |
| [3] |
Sun X, Zhong T, Zhang L, Zhang K S, Wu W X. Reducing ammonia volatilization from paddy field with rice straw derived biochar. Science of the Total Environment, 2019, 660: 512-518. DOI:10.1016/j.scitotenv.2018.12.450 |
| [4] |
Yu S, Xue L H, Feng Y F, Liu Y, Song Z Z, Mandal S, Yang L Z, Sun Q Y, Xing B S. Hydrochar reduced NH3 volatilization from rice paddy soil: Microbial-aging rather than water-washing is recommended before application. Journal of Cleaner Production, 2020, 268(11): 122233. |
| [5] |
Van Der Graaf S C, Kranenburg R, Segers A J, Schaap M, Erisman J W. Satellite-derived leaf area index and roughness length information for surface-atmosphere exchange modelling: a case study for reactive nitrogen deposition in north-western Europe using LOTOS-EUROS v2.0. Geoscientific Model Development, 2020, 13(5): 2451-2474. DOI:10.5194/gmd-13-2451-2020 |
| [6] |
Li Y, Schwandner F M, Sewell H J, Zivkovich A, Tigges M, Raja S, Holcomb S, Molenar J V, Sherman L, Archuleta C, Lee T, Collett J L Jr. Observations of ammonia, nitric acid, and fine particles in a rural gas production region. Atmospheric Environment, 2014, 83: 80-89. DOI:10.1016/j.atmosenv.2013.10.007 |
| [7] |
Bosch-Serra À D, Yagüe M R, Teira-Esmatges M R. Ammonia emissions from different fertilizing strategies in Mediterranean rainfed winter cereals. Atmospheric Environment, 2014, 84(1): 204-212. |
| [8] |
杨国英, 郭智, 刘红江, 王鑫, 陈留根. 稻田氨挥发影响因素及其减排措施研究进展. 生态环境学报, 2020, 29(9): 1912-1919. |
| [9] |
徐珊珊, 侯朋福, 范立慧, 薛利红, 杨林章, 王绍华, 李刚华. 生活污水灌溉对麦秸还田稻田氨挥发排放的影响. 环境科学, 2016, 37(10): 3963-3970. |
| [10] |
卢丽丽, 吴根义. 农田氨排放影响因素研究进展. 中国农业大学学报, 2019, 24(1): 149-162. |
| [11] |
杨帆, 李荣, 崔勇, 包耀贤, 段英华. 我国南方秸秆还田的培肥增产效应. 中国土壤与肥料, 2011, 47(1): 10-14. |
| [12] |
裴鹏刚, 张均华, 朱练峰, 禹盛苗, 胡志华, 金千瑜. 秸秆还田对水稻固碳特性及产量形成的影响. 应用生态学报, 2014, 25(10): 2885-2891. |
| [13] |
李凤博, 牛永志, 刘金根, 卞新民, 章秀福. 秸秆填埋对水稻土表层水三氮动态的影响. 农业环境科学学报, 2009, 28(3): 513-517. |
| [14] |
孔维栋, 刘可星, 廖宗文. 有机物料种类及腐熟水平对土壤微生物群落的影响. 应用生态学报, 2004, 15(3): 487-492. |
| [15] |
赵政鑫, 王晓云, 田雅洁, 王锐, 彭青, 蔡焕杰. 基于Meta分析的不同生产条件下秸秆还田对土壤氨挥发的影响. 环境科学, 2022, 43(3): 1678-1687. |
| [16] |
张刚, 王德建, 俞元春, 王灿, 庄锦贵. 秸秆全量还田与氮肥用量对水稻产量、氮肥利用率及氮素损失的影响. 植物营养与肥料学报, 2016, 22(4): 877-885. |
| [17] |
杨坤宇, 王美慧, 王毅, 尹黎明, 李勇, 吴金水. 不同农艺管理措施下双季稻田氮磷径流流失特征及其主控因子研究. 农业环境科学学报, 2019, 38(8): 1723-1734. |
| [18] |
汪军, 王德建, 张刚, 王远. 麦秸全量还田下太湖地区两种典型水稻土稻季氨挥发特性比较. 环境科学, 2013, 34(1): 27-33. |
| [19] |
张刚. 太湖地区稻麦两熟制农田秸秆还田综合效应研究[D]. 南京: 南京林业大学, 2020.
|
| [20] |
许仁良. 麦秸还田对水稻产量、品质及环境效应的影响[D]. 扬州: 扬州大学, 2010.
|
| [21] |
Zhang B W, Zhou M H, Lin H Y, Ntacyabukura T, Wang Y Q, Zhu B. Effects of different long-term crop straw management practices on ammonia volatilization from subtropical calcareous agricultural soil. Atmospheric and Oceanic Science Letters, 2020, 13(3): 232-239. |
| [22] |
何莹, 张玉钧, 尤坤, 高彦伟, 赵南京, 刘文清. 基于红外光谱技术的秸秆还田环境下土壤氨挥发特征分析. 红外与毫米波学报, 2017, 36(4): 397-402. |
| [23] |
Xu S S, Hou P F, Xue L H, Wang S H, Yang L Z. Treated domestic sewage irrigation significantly decreased the CH4, N2O and NH3 emissions from paddy fields with straw incorporation. Atmospheric Environment, 2017, 169: 1-10. |
| [24] |
Sun L Y, Wu Z, Ma Y C, Liu Y L, Xiong Z Q. Ammonia volatilization and atmospheric N deposition following straw and urea application from a rice-wheat rotation in southeastern China. Atmospheric Environment, 2018, 181: 97-105. |
| [25] |
Feng Y F, Sun H J, Xue L H, Liu Y, Gao Q, Lu K P, Yang L Z. Biochar applied at an appropriate rate can avoid increasing NH3 volatilization dramatically in rice paddy soil. Chemosphere, 2017, 168: 1277-1284. |
| [26] |
韩晨, 侯朋福, 薛利红, 冯彦房, 余姗, 杨林章. 麦秸水热炭及其改良产物对水稻产量和稻田氨挥发排放的影响. 环境科学, 2021, 42(7): 3451-3457. |
| [27] |
Kolb S, Knief C, Dunfield P F, Conrad R. Abundance and activity of uncultured methanotrophic bacteria involved in the consumption of atmospheric methane in two forest soils. Environmental Microbiology, 2005, 7(8): 1150-1161. |
| [28] |
Vance E D, Brookes P C, Jenkinson D S. An extraction method for measuring soil microbial biomass C. Soil Biology and Biochemistry, 1987, 19(6): 703-707. |
| [29] |
Long X E, Chen C R, Xu Z H, Oren R, He J Z. Abundance and community structure of ammonia-oxidizing bacteria and Archaea in a temperate forest ecosystem under ten-years elevated CO2. Soil Biology and Biochemistry, 2012, 46: 163-171. |
| [30] |
Stubner S. Enumeration of 16S rDNA of Desulfotomaculum lineage 1 in rice field soil by real-time PCR with SybrGreen detection. Journal of Microbiological Methods, 2002, 50(2): 155-164. |
| [31] |
Li M, Cao H L, Hong Y G, Gu J D. Spatial distribution and abundances of ammonia-oxidizing Archaea (AOA) and ammonia-oxidizing bacteria (AOB) in mangrove sediments. Applied Microbiology and Biotechnology, 2011, 89(4): 1243-1254. |
| [32] |
Muyzer G, De Waal E C, Uitterlinden A G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Applied and Environmental Microbiology, 1993, 59(3): 695-700. |
| [33] |
Eisenhauer N, BowkerM A, Grace J B, Powell J R. From patterns to causal understanding: Structural equation modeling (SEM) in soil ecology. Pedobiologia, 2015, 58(2): 65-72. |
| [34] |
侯朋福, 薛利祥, 俞映倞, 薛利红, 范立慧, 杨林章. 缓控释肥侧深施对稻田氨挥发排放的控制效果. 环境科学, 2017, 38(12): 5326-5332. |
| [35] |
曹金留, 田光明, 任立涛, 蔡祖聪, 何任红. 江苏南部地区稻麦两熟土壤中尿素的氨挥发损失. 南京农业大学学报, 2000, 23(4): 51-54. |
| [36] |
常菲, 红梅, 武岩, 李艳勤, 赵巴音那木拉, 德海山. 灌溉方式和改良措施对河套灌区盐渍土氨挥发的影响. 中国土壤与肥料, 2019(2): 38-45. |
| [37] |
Cao Y S, Sun H F, Zhang J N, Chen G F, Zhu H T, Zhou S, Xiao H Y. Effects of wheat straw addition on dynamics and fate of nitrogen applied to paddy soils. Soil and Tillage Research, 2018, 178: 92-98. |
| [38] |
潘孝晨, 唐海明, 肖小平, 李超, 汤文光, 程凯凯, 郭立君, 黄桂林, 汪柯. 不同耕作和秸秆还田模式对紫云英-双季稻土壤微生物生物量碳、氮含量的影响. 生态环境学报, 2019, 28(8): 1585-1595. |
| [39] |
李伟, 袁亮, 赵秉强. 增值尿素的氨挥发特征及其对土壤微生物量碳和脲酶活性的影响. 腐植酸, 2013, 155(6): 15-20. |
| [40] |
靳红梅, 常志州, 郭德杰, 吴华山. 追施猪粪沼液对菜地氨挥发的影响. 土壤学报, 2012, 49(1): 86-95. |
| [41] |
Bru D, Ramette A, Saby N P A, Dequiedt S, Ranjard L, Jolivet C, Arrouays D, Philippot L. Determinants of the distribution of nitrogen-cycling microbial communities at the landscape scale. The ISME Journal, 2011, 5(3): 532-542. |
| [42] |
Yao H Y, Gao Y M, Nicol G W, Campbell C D, Prosser J I, Zhang L M, Han W Y, Singh B K. Links between ammonia oxidizer community structure, abundance, and nitrification potential in acidic soils. Applied and Environmental Microbiology, 2011, 77(13): 4618-4625. |
| [43] |
Wu Y, Lu L, Wang B, Lin X, Zhu J, Cai Z, Yan X, Jia Z. Long-term field fertilization significantly alters community structure of ammonia-oxidizing bacteria rather than archaea in a paddy soil. Soil Science Society of America Journal, 2011, 75(4): 431-1439. |
| [44] |
宋大利, 侯胜鹏, 王秀斌, 梁国庆, 周卫. 中国秸秆养分资源数量及替代化肥潜力. 植物营养与肥料学报, 2018, 24(1): 1-21. |
| [45] |
Schauss K, Focks A, Leininger S, Kotzerke A, Heuer H, Thiele-Bruhn S, Sharma S, Wilke B M, Matthies M, Smalla K, Munch J C, Amelung W, Kaupenjohann M, Schloter M, Schleper C. Dynamics and functional relevance of ammonia-oxidizing Archaea in two agricultural soils. Environmental Microbiology, 2009, 11(2): 446-456. |
| [46] |
Jung M Y, Sedlacek C J, Kits K D, Mueller A J, Rhee S K, Hink L, Nicol G W, Bayer B, Lehtovirta-Morley L, Wright C, de la Torre J R, Herbold C W, Pjevac P, Daims H, Wagner M. Ammonia-oxidizing Archaea possess a wide range of cellular ammonia affinities. The ISME Journal, 2022, 16(1): 272-283. |
| [47] |
Taylor A E, Taylor K, Tennigkeit B, Palatinszky M, Stieglmeier M, Myrold D D, Schleper C, Wagner M, Bottomley P J. Inhibitory effects of C2 to C10 1-alkynes on ammonia oxidation in two Nitrososphaera species. Applied and Environmental Microbiology, 2015, 81(6): 1942-1948. |