 2024, Vol. 44
2024, Vol. 44文章信息
- 李祖婵, 玄锦, 王秋雪, 彭书婷, 孙耿敏, 黄柳菁
- LI Zuchan, XUAN Jin, WANG Qiuxue, PENG Shuting, SUN Gengmin, HUANG Liujing
- 闽江江心洲不同生境植物叶功能性状差异及其对土壤特性的响应
- Differences of plant leaf functional traits in different habitats and their responses to soil characteristics in central bar of Minjiang River
- 生态学报. 2024, 44(14): 6301-6316
- Acta Ecologica Sinica. 2024, 44(14): 6301-6316
- http://dx.doi.org/10.20103/j.stxb.202311192515
-
文章历史
- 收稿日期: 2023-11-19
- 网络出版日期: 2024-05-11

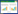




植物功能性状表征植物长期适应环境变化而形成的形态及生理的生态策略, 反映了植物适生策略的生态系统功能特征[1]。植物的功能性状之间存在着内在的联系和权衡关系, 其中最为常见的就是权衡关系, 这种权衡关系是在植物长期进化的过程中, 通过自然环境的筛选而产生的一系列功能性状的组合, 也被称为“生态策略”[2—3]。Wright等[4]基于全球范围内175个样点的2548种植物的叶功能性状进行研究, 首次在全球尺度上揭示了一系列叶性状组合之间的相互联系和协同变化规律。研究发现, 叶寿命较短的物种具有高叶氮/磷含量、高光合速率、快速呼吸速率和小比叶重的特点, 即“快速投资收益型”;反之, 叶寿命较长的物种则具有低叶氮/磷含量、低光合速率、缓慢呼吸速率和大比叶重的特点, 即“缓慢投资收益型”, 这表明植物在利用资源时采取了权衡策略。这些功能性状组合共同构成了全球叶片经济学谱, 反映了物种沿环境梯度的分布情况。通过性状组合的动态变化, 植物能够适应其所处的生境, 从而实现生存和繁殖。
根据学者的研究, 通过将个体水平的植物功能性状值进行加权赋值计算得到的植物功能性状加权平均值(CWM), 极大促进了对植物群落与环境因子之间关系的研究[5—7]。在植物功能性状中, 叶片与环境的接触最为密切, 对环境变化有高度敏感性, 叶功能性状会随环境的改变而产生表型可塑性的差异性特征, 各性状间协同进化, 直观体现了其对环境变化的适应机制[8—9]。相关研究表明, 在大尺度上, 气候因子是影响植物功能性状的决定性因素, 而在小尺度上, 土壤因子的影响更大, 土壤与植物有着密切的物质转化与互作关系[10—11]。
江心洲是指在河流和河床的相互作用下, 由心滩逐渐淤高发育而成的, 是一种特殊形式的湿地类型[12—13]。江心洲具有净化水质、涵养水源、调节微气候、维持生物多样性、调节生态服务功能和防止水土流失的生态系统服务功能。然而, 由于江心洲的边界隔离效应, 能量获取主要依靠河流, 生态敏感而又脆弱, 鉴于此, 江心洲的生态系统一旦受到干扰破坏后, 就难于恢复[14]。据调查显示, 闽江福州河段的厚美沙洲、中房洲和古山洲等江心洲, 有较大部分土地转换为牧场、耕地和园林绿化用地等, 其原有植被受到不同程度的破坏。在河流水动力、潮汐及洪水侵蚀的多重影响下, 土壤水分、养分分布不均匀导致闽江福州河段江心洲形成了不同的土壤生境类型, 各生境间的土壤异质性大, 进而影响了江心洲植物群落的分布。然而, 目前对闽江江心洲的研究主要集中在土壤重金属污染等方面, 而对闽江江心洲尺度上的植物群落叶功能性状与土壤环境之间的关系有待深入探究。
因此, 本研究以闽江福州河段5个江心洲的植物群落为研究对象, 探讨不同生境类型(洲岸沙滩、洲岸泥滩和洲中央林地)植物群落叶功能性状与土壤理化性质的关系, 以期回答以下问题:(1)江心洲不同生境类型的土壤理化性质和植物群落叶功能性状是否存在差异?(2)江心洲不同生境类型的植物群落功能性状对土壤会有怎样的响应机制?研究结果可为江心洲植被的修复、重建和管理, 维护敏感而又脆弱的湿地生态系统提供理论依据。
1 研究区概况与研究方法 1.1 研究区概况闽江流域位于中国东南沿海福建省境内(116°23′—119°35′E, 25°23′—28°16′N), 为福建省内的第一大入海河流, 长577 km, 流域面积60 092 km2, 上游三大支流沙溪、建溪和富屯溪在南平汇合, 进入闽江干流, 流经福州市区后经马尾自西向东流入东海[15]。闽江福州河段位于闽江下游, 长120 km, 流域面积约8000 km2, 地处中亚热带和南亚热带过渡区, 气候暖热潮湿, 雨热同期, 年均气温19 ℃, 年降水量达1300 mm, 湿地生态系统发育良好[16]。全年受到河口水动力和海洋潮汐的共同作用, 潮汐类型属于典型的正规半日潮, 全河段属于感潮河道, 受潮汐影响极大, 冬春季节潮位较低, 夏秋季节潮位较高, 平均潮差4.1 m(图 1), 5—6月份是闽江流域的主汛期, 降水强度为全年最大[17]。

|
| 图 1 古山洲涨潮与退潮的对比图 Fig. 1 Comparison between rising tide and ebbing tide in Gushan Bar |
本研究沿闽江福州河段上游至下游方向选取5个江心洲(图 2), 即中房洲、绿洲、古山洲、厚美沙洲和大洋鹭洲, 其面积等详细信息, 如表 1所示。其中, 选取人为干预较小或无人为干预的, 能较好反映江心洲植物群落基本特征的原生自然植被区域作为调查样地[18—19]。

|
| 图 2 研究区地理位置和样地分布示意图 Fig. 2 Schematic diagram of the geographical location and distribution of sample plots in the study area |
| 编号 No. |
江心洲 Central bar |
面积 Area/m2 |
经度(E) Longitude |
纬度(N) Latitude |
| 1 | 中房洲 | 655 082 | 119°11′25″—119°12′24″ | 26°06′30″—26°06′55″ |
| 2 | 绿洲 | 208 276 | 119°12′36″—119°12′59″ | 26°06′20″—26°06′38″ |
| 3 | 古山洲 | 506 532 | 119°12′56″—119°13′42″ | 26°06′09″—26°06′28″ |
| 4 | 厚美沙洲 | 722 637 | 119°12′24″—119°13′04″ | 26°04′57″—26°05′46″ |
| 5 | 大洋鹭洲 | 353 216 | 119°13′08″—119°13′13″ | 26°13′07″—26°37′59″ |
调研结果显示, 闽江福州河段江心洲植物群落较为简单, 植被种类单一, 主要的乔木种类有枫杨(Pterocarya stenoptera)、乌桕(Triadica sebifera)、楝(Melia azedarach)和榕树(Ficus macrocarpa)等;主要的灌木种类有小蜡(Ligustrum sinense)、枫杨和构(Broussonetia papyrifera)等;主要的草本种类有水蓼(Persicaria hydropiper)马兰(Aster indicus)、荩草(Arthraxon hispidus )和艾(Artemisia argyi)等。
1.2 研究方法 1.2.1 生境划分根据江心洲土壤的发育程度[20]以及土壤是否直接受到河流水动力和潮汐的侵蚀作用, 并结合实地勘察, 闽江福州河段江心洲主要有3种生境类型(图 3)如下, (1)洲岸沙滩:土壤为较新淤积、发育较浅的沙质土壤, 且直接受到河流水动力和潮汐的侵蚀作用;(2)洲岸泥滩:土壤为较新淤积、发育较浅的泥质土壤, 且直接受到河流水动力和潮汐的侵蚀作用;(3)洲中央林地:土壤为发育较成熟的耕作土壤, 且没有直接受到河流水动力和潮汐的侵蚀作用。

|
| 图 3 闽江福州河段江心洲的3种生境类型 Fig. 3 Three habitat types of central bar in Fuzhou Reach of Minjiang River |
于2022年5月至6月, 采用样方调查法调查闽江福州河段5个江心洲的植物群落。沿闽江福州河段上游至下游方向, 分别在中房洲、绿洲、古山洲、厚美沙洲和大洋鹭洲设置样地, 每个江心洲分别设置上述3种不同的生境样地, 每个生境样地设置3个(符合同一生境条件)不同样方, 每个样方至少间隔20米及以上, 共设置15个样地和45个样方(图 2)。在每个样方中, 根据其主要的植物群落情况设置乔木样方(面积10 m×10 m)、灌木样方(面积5 m×5 m, 在有灌木分布的乔木样方内沿着对角线随机选取1或2个灌木样方)和草本样方(面积1 m×1 m, 在有草本分布的灌木样方内沿着对角线随机选取1或2个草本样方), 累计设置乔木样方38个, 灌木样方37个, 草本样方81个。在乔木样方内调查胸径大于等于1 cm的乔木, 记录乔木的种名、数量、高度和胸径;在灌木样方内调查记录所有灌木的种名、数量、高度和基茎;在草本样方内调查记录所有草本的种名、数量、高度和盖度。其中, 高度采用测高仪(精度1 cm)或卷尺(精度0.1 cm)测量, 胸径采用围尺(精度0.1 cm)测量, 盖度参考文献[21]中的方法测量。采用GPS定位记录样地的经度、纬度、海拔和小生境等指标。
参考《中国植物志》[22]《福建植物志》[23]对江心洲植物进行鉴定和分类, 参考《中国入侵植物名录》[24]确定外来入侵种, 采用恩格勒系统对江心洲植物进行科属种的统计分析。
1.2.3 样品采集(1) 土壤样品采集
在每个样方的中心位置清除表层枯落物后, 用环刀法取0—20 cm的土壤样品, 即在地面平整处将体积为100 cm3环刀垂直压入土内, 当环刀充满土壤后, 用铁铲将环刀取出, 用削土刀将环刀两侧多余的土壤削去, 然后, 将环刀土样装入铝盒保存, 带回实验室测定土壤容重和土壤含水量。在每个样方内的5个1 m×1 m草本样方的4个顶点和中心位置, 在清除表层枯落物后, 用直径10 cm的土钻采集0—20 cm的土壤样品, 将同一样方的5份土层样品混合为1个待测土钻土样, 每个样方取土约1 kg左右, 将土钻土样装入密封袋, 带回实验室。
(2) 植物叶片样品采集
选取与物种的生长速率、养分循环和生态策略等密切相关的8个叶功能性状, 详细计算公式和生态意义等信息, 如表 2所示。分乔木层、灌木层和草本层3个不同的植物层次, 选取样方内重要值大于0.2的优势种进行叶片采集, 每个优势种选择3—5株生长成熟、健康良好的植株, 每株采集各个方向健康成熟、生长均匀、完全展开的4—6片未受损叶片, 最终挑选15—20片。将采集的植物样品装入密封袋内, 喷洒自来水, 并放在装有冰袋的保温袋内保鲜, 然后带回实验室放入冰箱冷藏保存, 并在24 h内测定相关指标。
| 编号 No. |
叶功能性状 Leaf functional traits |
计算公式及单位 Formula and unit |
生态意义 Ecological meaning |
| 1 | LA | LA(cm2) | 直接反映植物对不同生境的实验特征和植物获取资源的能力 |
| 2 | SLA | SLA(cm2/g)=叶面积(cm2)/叶片干重(g) | 反映植物对碳获取的策略以及表征环境资源匮乏的权衡表现 |
| 3 | SLW | SLW(g/cm2)=叶片干重(g)/叶面积(cm2) | 叶片对资源利用率和环境适应性的表现 |
| 4 | LDMC | LDMC(g/g)=叶片干重(g)/叶片鲜重(g) | 表征植物对养分的保有能力, 直观反映植物获取利用有效资源略 |
| 5 | LWC | LWC(%)=[叶片鲜重(g)-叶片干重(g)/叶片鲜重(g)×100% | 反映叶片的容量, 影响植物生长发育、光合作用能力 |
| 6 | LTC | LTC(g/kg)=叶片全碳(g)/叶片干重(kg) | 叶片组织的构建成本, 与植物的生长发育有关 |
| 7 | LTN | LTN(g/kg)=叶片全氮(g)/叶片干重(kg) | 判断叶片光合作用强弱的决定性因素 |
| 8 | LTP | LTP(g/kg)=叶片全磷(g)/叶片干重(kg) | 与植物的资源获取、代谢和生长发育有关 |
| LA:叶面积Leaf area;SLA:比叶面积Specific leaf area;SLW:比叶重Specific leaf weight;LDMC:叶干物质含量Leaf dry matter content;LWC:叶片含水量Leaf water content;LTC:叶全碳含量Total carbon content of leaves;LTN:叶全氮含量Leaf total nitrogen content;LTP:叶全磷含量Leaf total nitrogen and phosphorus content | |||
(1) 土壤指标测定
1) 土壤容重和土壤含水量的测定[25]
把环刀土样转移至重量为W1的铝盒, 环刀土样的体积记为V, 用精度为0.001的电子天平称重, 记为W2, 再将铝盒开盖后放入105 ℃的烘干机内烘干至恒重, 然后将铝盒拿出来, 待土壤温度降至与室内温度一致后称重, 记为W3。
土壤容重(SBD, g/cm3)的计算如下:
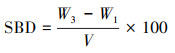
|
(1) |
土壤含水量(SWC, %)的计算如下:

|
(2) |
2) 土壤温度、土壤含盐率和土壤电导率的测定
用土壤温湿度计(TZS-2X-G)测量20 cm深度的土壤温度、土壤含盐率和土壤电导率, 其中土壤温度在上午9点至11点之间进行测定。
3) 土壤pH值的测定[26]:采用电位法测定。
4) 土壤全碳含量、土壤全氮含量和土壤全磷含量的测定[27]
土壤全碳含量和土壤全氮含量的测定:采用碳氮元素分析仪测定(德国elementar vario Macro cube元素分析仪);
土壤全磷含量的测定:采用钼锑抗比色法测定。
(2) 叶功能性状指标测定
1) 叶面积的测定[26]
首先, 用扫描仪(CanoScan LIDE300)进行叶片扫描, 接着用ImageJ软件进行测量, 得到叶面积(LA)。
2) 比叶面积、比叶重、叶干物质含量和叶片含水量的测定[28—29]
将新鲜叶片从自封袋中取出, 清理叶片表面的杂质后, 利用精度为0.001的电子天平称量叶片鲜重(Wf), 再将叶片放入信封内, 置于105 ℃的烘干机中杀青30 min后, 然后置于65 ℃的烘干机中烘干至恒重, 最后, 称量叶片干重(Wd)。
比叶面积的计算如下:
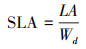
|
(3) |
比叶重的计算如下:
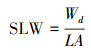
|
(4) |
叶干物质含量的计算如下:
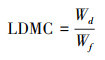
|
(5) |
叶片含水量的计算如下:
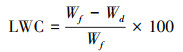
|
(6) |
(3) 叶全碳含量、叶全氮含量和叶全磷含量的测定[30]
烘干的植物叶片, 待其冷却后, 经研磨、过筛(0.149 mm)后, 置于自封袋中密封保存, 用于养分测定, 测定的具体方法如下:
叶全碳含量(LTC)的测定:采用碳氮元素分析仪测定(德国elementar vario Macro cube元素分析仪);
叶全氮含量(LTN)的测定:采用碳氮元素分析仪测定(德国elementar vario Macro cube元素分析仪);
叶全磷含量(LTP)的测定:采用钼锑抗比色法测定。
1.2.5 数据计算(1) 重要值计算
计算闽江福州河段5个江心洲45个样方中各乔木层、灌木层和草本层植物的重要值, 如下:
乔木的重要值[13]:

|
(7) |
灌木和草本植物的重要值[31]:

|
(8) |
(2) 植物群落叶功能性状值的计算
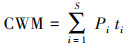
|
(9) |
式中, S为样方内重要值大于0.2的植物物种数;Pi为物种i在群落中的重要值;ti为物种i在群落中的植物叶功能性状值。
1.2.6 数据分析本研究中, 数据处理、分析以及绘图均在Excel 2016、SPSS 26.0和Canoco 5.0软件中完成。
(1) 分析江心洲不同生境植物群落的土壤理化性质特征以及植物群落叶功能性状的差异
利用SPSS 26.0软件, 采用单因素方差分析法分析江心洲不同生境植物群落的土壤理化性质特征以及植物群落叶功能性状的差异, 并采用LSD法(最小显著差异法)进行多重比较, 并在Excel 2016中制表和绘图。
(2) 分析不同生境植物群落叶功能性状对土壤的响应
利用Canoco 5.0软件对江心洲不同生境的植物群落叶功能性状与土壤理化性质进行冗余分析(RDA), 并在Canoco 5.0中绘图。
2 结果与分析 2.1 不同生境的植物群落叶功能性状差异将闽江福州河段5个江心洲3种不同生境植物群落叶功能性状, 即叶面积(LA)、比叶面积(SLA)、比叶重(SLW)、叶干物质含量(LDMC)、叶片含水量(LWC)、叶全碳含量(LTC)、叶全氮含量(LTN)和叶全磷含量(LTP), 共8个叶功能性状进单因素方差分析, 如表 3所示。江心洲不同生境的植物群落叶功能性状存在显著差异(P<0.05)。在乔木层上, 8个叶功能性状在不同生境中均无显著性差异(P>0.05);在灌木层上, 洲中央林地的叶面积显著(P<0.05)高于洲岸泥滩和洲岸沙滩;在草本层上, 洲中央林地的比叶面积存在显著(P<0.05)高于洲岸沙滩, 而洲岸泥滩的比叶面积与洲中央林地和洲岸沙滩均无显著性差异(P>0.05)。
| 叶功能性状 Leaf functional traits |
洲岸沙滩 Sandy beach of bank |
洲岸泥滩 Mudflat of bank |
洲中央林地 Forest land of central bar |
叶功能性状 Leaf functional traits |
洲岸沙滩 Sandy beach of bank |
洲岸泥滩 Mudflat of bank |
洲中央林地 Forest land of central bar |
|
| 乔木层 Arbor layer |
LWC | 0.668±0.047a | 0.690±0.076a | 0.705±0.061a | ||||
| LA | 33.460±40.798a | 34.668±29.147a | 39.099±47.062a | LTC | 446.863±26.025a | 442.117±44.649a | 433.933±35.356a | |
| SLA | 182.601±59.473a | 225.35±101.269a | 189.960±35.584a | LTN | 29.288±6.336a | 28.130±7.420a | 33.819±6.758a | |
| SLW | 0.006±0.002a | 0.005±0.001a | 0.006±0.001a | LTP | 2.832±0.846a | 2.662±0.729a | 3.091±0.979a | |
| LDMC | 0.393±0.072a | 0.391±0.088a | 0.333±0.046a | 草本层 Herb layer |
||||
| LWC | 0.607±0.072a | 0.680±0.195a | 0.667±0.046a | LA | 12.132±11.333a | 24.831±15.866a | 26.134±20.303a | |
| LTC | 448.572±33.146a | 491.339±110.509a | 449.783±31.492a | SLA | 384.974±70.808b | 496.254±105.157ab | 535.484±171.201a | |
| LTN | 28.040±3.614a | 32.248±9.283a | 26.990±6.095a | SLW | 0.003±0.001a | 0.002±0.001a | 0.002±0.001a | |
| LTP | 2.356±0.486a | 2.354±0.802a | 2.267±0.641a | LDMC | 0.206±0.043a | 0.186±0.030a | 0.168±0.039a | |
| 灌木层 Shrub layer |
LWC | 0.794±0.043a | 0.814±0.030a | 0.832±0.039a | ||||
| LA | 14.280±12.290b | 31.685±42.061b | 80.935±53.139a | LTC | 417.157±17.645a | 400.213±34.686a | 423.97±20.457a | |
| SLA | 226.763±71.274a | 283.698±141.777a | 305.999±149.374a | LTN | 42.885±10.519a | 39.517±4.380a | 40.456±7.817a | |
| SLW | 0.005±0.001a | 0.004±0.002a | 0.004±0.0001a | LTP | 4.215±1.011a | 3.732±0.727a | 4.238±0.798a | |
| LDMC | 0.332±0.047a | 0.310±0.076a | 0.295±0.061a | |||||
| 同行不同小写字母表示差异显著(P<0.05) | ||||||||
将闽江福州河段5个江心洲3种不同生境植物群落45个样方中的土壤pH值、土壤容重(SBD)、土壤含水量(SWC)、土壤温度(ST)、土壤含盐率(SS)、土壤电导率(EC)、土壤全碳含量(TC)、土壤全氮含量(TN)和土壤全磷含量(TP), 共9个土壤因子进行单因素方差分析, 如表 4所示。
| 土壤因子 Soil factors |
洲岸沙滩 Sandy beach of bank |
洲岸泥滩 Mudflat of bank |
洲中央林地 Forest land of central bar |
| pH | 6.92±0.08a | 6.35±0.32b | 7.01±1.04a |
| SBD/(g/cm3) | 1.08±0.06a | 0.91±0.07c | 0.98±0.07b |
| SWC/% | 16.39±5.84b | 40.70±11.02a | 16.06±8.51b |
| ST/(℃) | 27.70±2.0a | 28.10±2.0a | 28.60±2.5a |
| SS/(mg/L) | 5.50±5.0b | 32.70±25.7a | 21.80±16.0a |
| EC/(μm/cm) | 10.60±9.2c | 63.50±46.0a | 39.70±27.6b |
| TC/(g/kg) | 7.02±7.73b | 18.53±7.17a | 17.36±11.28a |
| TN/(g/kg) | 0.69±0.53b | 1.69±0.57a | 1.55±0.91a |
| TP/(g/kg) | 0.35±0.13b | 0.90±0.20a | 0.79±0.29a |
| SBD:容重Soil bulk density;SWC:含水量Soil moisture conten;ST:温度Soil temperature;SS:含盐率Soil salinity rate;EC:电导率Electrical conductivity;TC:全碳含量Total carbon content;TN:全氮含量Total nitrogen content;TP:全磷含量Total phosphorus content | |||
除土壤温度外, 江心洲不同生境的其余土壤理化性质存在显著差异(P<0.05)。与洲岸沙滩相比, 洲岸泥滩和洲中央林地的土壤含盐率、土壤全碳含量、土壤全氮含量和土壤全磷含量显著(P<0.05)偏高。两两生境间的土壤电导率差异显著(P<0.05), 表现为洲岸泥滩>洲中央林地>洲岸沙滩。两两生境间的土壤容重差异显著(P<0.05), 表现为洲岸沙滩>洲中央林地>洲岸泥滩。洲中央林地的土壤呈弱碱性, 而洲岸沙滩和洲岸泥滩的土壤呈弱酸性。
2.3 不同生境的植物群落叶功能性状与土壤因子的相关关系将闽江福州河段5个江心洲3种不同生境的植物群落叶功能性状, 即叶面积(LA)、比叶面积(SLA)、比叶重(SLW)、叶干物质含量(LDMC)、叶片含水量(LWC)、叶全碳含量(LTC)、叶全氮含量(LTN)和叶全磷含量(LTP), 共8个叶功能性状作为响应变量, 与土壤pH值(pH)、土壤容重(SBD)、土壤含水量(SWC)、土壤温度(ST)、土壤含盐率(SS)、土壤电导率(EC)、土壤全碳含量(TC)、土壤全氮含量(TN)和土壤全磷含量(TP), 共9个土壤因子作为解释变量, 进行冗余分析(RDA)。
2.3.1 洲岸沙滩的植物群落叶功能性状与土壤因子的相关关系如图 4和表 5所示, 在乔木层上, 前两个排序轴对洲岸沙滩乔木层植物群落叶功能性状的解释率分别为61.28%、3.08%, 累计解释率达64.36%, 前两轴能较好反映了乔木层植物群落叶功能性状和土壤因子的相关性, 且主要由RDA1轴决定, 排序结果科学。蒙特卡洛检验表明, 土壤全碳含量的P值小于0.05, 是显著(P<0.05)影响洲岸沙滩乔木层植物群落叶功能性状变化的土壤因子。土壤全碳含量与叶片含水量、叶全氮含量和叶全磷含量等8个叶功能性状均呈负相关关系。

|
| 图 4 洲岸沙滩的植物群落叶功能性状与土壤因子的RDA排序图 Fig. 4 RDA ordination map of leaf functional traits of plant community in sandy beach of bank and soil factors LA:叶面积Leaf area;SLA:比叶面积Specific leaf area;SLW:比叶重Specific leaf weight;LDMC:叶干物质含量Leaf dry matter content;LWC:叶片含水量Leaf water content;LTC:叶全碳含量Total carbon content of leaves;LTN:叶全氮含量Leaf total nitrogen content;LTP:叶全磷含量Leaf total nitrogen and phosphorus content;SBD:容重Soil bulk density;SWC:含水量Soil moisture conten;ST:温度Soil temperature;SS:含盐率Soil salinity rate;EC:电导率Electrical conductivity;TC:全碳含量Total carbon content;TN:全氮含量Total nitrogen content;TP:全磷含量Total phosphorus content;RDA:冗余分析Redundancy Analysis |
| 生境 Habitat |
土壤因子 Soil factor |
解释变异量 Explain the amount of variation |
贡献率/% Contribution/% |
伪F统计量 Pseudo F statistic |
P |
| 乔木层 | pH | 9.3 | 14.3 | 2.1 | 0.180 |
| Arbor layer | SBD | 1.2 | 1.8 | 0.2 | 0.766 |
| SWC | 12.7 | 19.6 | 2.7 | 0.106 | |
| ST | 0.6 | 0.9 | < 0.1 | 0.848 | |
| SS | 2.6 | 4 | 0.5 | 0.496 | |
| EC | 4.3 | 6.6 | 1 | 0.336 | |
| TC | 30.1 | 46.4 | 5.6 | 0.018 | |
| TN | 2.6 | 4 | 0.6 | 0.520 | |
| TP | 1.4 | 2.2 | 0.3 | 0.666 | |
| 灌木层 | pH | 0.3 | 0.5 | < 0.1 | 0.934 |
| Shrub layer | SBD | 7.6 | 13.1 | 1.6 | 0.210 |
| SWC | 3.9 | 6.8 | 0.8 | 0.430 | |
| ST | 26.5 | 45.7 | 4.7 | 0.034 | |
| SS | 3.3 | 5.8 | 0.7 | 0.438 | |
| EC | 2.8 | 4.8 | 0.5 | 0.498 | |
| TC | 12.6 | 21.7 | 2.5 | 0.120 | |
| TN | 0.3 | 0.5 | < 0.1 | 0.946 | |
| TP | 0.7 | 1.1 | 0.1 | 0.844 | |
| 草本层 | pH | 9.2 | 13.4 | 1.5 | 0.242 |
| Herb layer | SBD | 17.3 | 25.1 | 2.7 | 0.132 |
| SWC | 3 | 4.3 | 0.6 | 0.478 | |
| ST | 5.3 | 7.7 | 1.1 | 0.280 | |
| SS | 15.2 | 22.1 | 3.1 | 0.088 | |
| EC | 6.6 | 9.6 | 1.4 | 0.282 | |
| TC | 0.5 | 0.7 | < 0.1 | 0.834 | |
| TN | 2.8 | 4 | 0.4 | 0.558 | |
| TP | 9.0 | 13.1 | 1.5 | 0.232 |
在灌木层上, 前两个排序轴对洲岸沙滩灌木层植物群落叶功能性状的解释率分别为56.20%、1.70%, 累计解释率达57.90%, 前两轴能较好反映了灌木层植物群落叶功能性状和土壤因子的相关性, 且主要由RDA1轴决定, 排序结果科学。蒙特卡洛检验表明, 土壤温度的P值小于0.05, 是显著(P<0.05)影响洲岸沙滩灌木层植物群落叶功能性状变化的土壤因子, 土壤温度与叶片含水量和叶干物质含量等8个叶功能性状均呈正相关关系。
在草本层上, 前两个排序轴对洲岸沙滩草本层植物群落叶功能性状的解释率分别为67.64%、1.27%, 累计解释率达68.91%, 前两轴能较好反映草本层植物群落叶功能性状和土壤因子的相关性, 且主要由RDA1轴决定, 排序结果科学。土壤容重的解释变异量最高, 对洲岸沙滩草本层植物群落叶功能性状的影响最大, 其次为土壤含盐率>土壤pH值。土壤容重与比叶面积和叶全磷含量等8个叶功能性状均呈负相关关系。
综上, 土壤全碳含量、土壤温度、土壤容重是影响洲岸沙滩植物群落叶功能性状的主要土壤因子。
2.3.2 洲岸泥滩的植物群落叶功能性状与土壤因子的相关关系如图 5和表 6所示, 在乔木层上, 前两个排序轴对洲岸泥滩乔木层植物群落叶功能性状的解释率分别为82.78%、2.51%, 累计解释率达85.29%, 前两轴能很好反映了乔木层植物群落叶功能性状和土壤因子的相关性, 且主要由RDA1轴决定, 排序结果科学。蒙特卡洛检验表明, 土壤含水量和土壤全碳含量的P值均小于0.05, 是显著(P<0.05)影响洲岸泥滩乔木层植物群落叶功能性状变化的土壤因子。土壤含水量、土壤全碳含量与比叶面积、叶全碳含量和叶干物质含量等8个叶功能性状均呈负相关关系。

|
| 图 5 洲岸泥滩的植物群落叶功能性状与土壤因子的RDA排序图 Fig. 5 RDA ordination map of leaf functional traits of plant community in mudflat of bank and soil factors |
| 生境 Habitat |
土壤因子 Soil factor |
解释变异量 Explain the amount of variation |
贡献率/% Contribution |
伪F统计量 Pseudo F statistic |
P |
| 乔木层 | pH | 3.2 | 3.7 | 1.5 | 0.284 |
| Arbor layer | SBD | 8.9 | 10.4 | 3.7 | 0.058 |
| SWC | 43.9 | 50.9 | 10.2 | 0.022 | |
| ST | 1 | 1.1 | 0.4 | 0.618 | |
| SS | 1 | 1.1 | 0.4 | 0.680 | |
| EC | 1.6 | 1.9 | 0.7 | 0.458 | |
| TC | 20.8 | 24.1 | 7.1 | 0.028 | |
| TN | 1.9 | 2.2 | 0.6 | 0.518 | |
| TP | 3.9 | 4.5 | 1.7 | 0.224 | |
| 灌木层 | pH | 1.2 | 2.2 | 0.2 | 0.786 |
| Shrub layer | SBD | 11.2 | 19.7 | 1.8 | 0.192 |
| SWC | 8.2 | 14.5 | 1.5 | 0.194 | |
| ST | 1.9 | 3.3 | 0.3 | 0.748 | |
| SS | 2.6 | 4.7 | 0.5 | 0.590 | |
| EC | 1 | 1.7 | 0.1 | 0.874 | |
| TC | 15.4 | 27.2 | 2.4 | 0.132 | |
| TN | 12.1 | 21.4 | 2.2 | 0.146 | |
| TP | 3.0 | 5.2 | 0.5 | 0.588 | |
| 草本层 | pH | 1.0 | 1.0 | 0.9 | 0.450 |
| Herb layer | SBD | 70.1 | 74.6 | 30.4 | 0.002 |
| SWC | 8.3 | 8.9 | 4.6 | 0.046 | |
| ST | 0.5 | 0.5 | 0.4 | 0.672 | |
| SS | 1.8 | 1.9 | 1.8 | 0.194 | |
| EC | 1.2 | 1.2 | 1 | 0.366 | |
| TC | 2.9 | 3.1 | 1.7 | 0.226 | |
| TN | 7.6 | 8.1 | 6.9 | 0.012 | |
| TP | 0.6 | 0.7 | 0.6 | 0.558 |
在灌木层上, 前两个排序轴对洲岸泥滩灌木层植物群落叶功能性状的解释率分别为52.76%、3.64%, 累计解释率达56.40%, 前两轴能较好反映灌木层植物群落叶功能性状和土壤因子的相关性, 且主要由RDA1轴决定, 排序结果科学。土壤全碳含量的解释变异量最高, 对洲岸泥滩植物群落叶功能性状的影响最大, 其次为土壤全氮含量>土壤容重。土壤全碳含量与比叶面积、叶全氮含量和叶干物质含量等8个叶功能性状均呈正相关关系。
在草本层上, 前两个排序轴对洲岸泥滩草本层植物群落叶功能性状的解释率分别为91.36%、2.52%, 累计解释率达93.88%, 前两轴很好反映了草本层植物群落叶功能性状和土壤因子的相关性, 且主要由RDA1轴决定, 排序结果科学。蒙特卡洛检验表明, 土壤全氮含量、土壤容重和土壤含水量的P值小于0.05, 是显著(P<0.05)影响洲岸泥滩草本层植物群落叶功能性状变化的土壤因子。土壤全氮含量、土壤容重与比叶面积、叶全氮含量等8个叶功能性状均呈正相关关系;土壤含水量与叶面积和比叶面积等8个叶功能性状均呈负相关关系。
综上, 土壤容重、土壤含水量、土壤全碳含量和土壤全氮含量是影响洲岸泥滩植物群落叶功能性状的主要土壤因子。
2.3.3 洲中央林地的植物群落叶功能性状与土壤因子的相关关系如图 6和表 7所示, 在乔木层上, 前两个排序轴对洲中央林地乔木层植物群落叶功能性状的解释率分别为44.69%、5.36%, 累计解释率达50.05%, 前两轴能较好反映乔木层植物群落叶功能性状和土壤因子的相关性, 且主要由RDA1轴决定, 排序结果科学。土壤全碳含量的解释变异量最高, 对洲中央林地乔木层植物群落叶功能性状的影响最大, 其次为土壤容重>土壤电导率。土壤全碳含量与比叶面积、叶全氮含量和叶全磷含量呈负相关关系。

|
| 图 6 洲中央林地的植物群落叶功能性状与土壤因子的RDA排序图 Fig. 6 RDA ordination map of leaf functional traits of plant community in forest land of central bar and soil factors |
| 生境 Habitat |
土壤因子 Soil factor |
解释变异量 Explain the amount of variation |
贡献率/% Contribution |
伪F统计量 Pseudo F statistic |
P |
| 乔木层 | pH | 0.4 | 0.7 | < 0.1 | 0.958 |
| Arbor layer | SBD | 7.7 | 14.9 | 0.9 | 0.436 |
| SWC | 5.1 | 9.9 | 0.7 | 0.436 | |
| ST | 6.1 | 11.9 | 0.8 | 0.374 | |
| SS | 3.7 | 7.2 | 0.5 | 0.568 | |
| EC | 6.6 | 12.8 | 0.9 | 0.394 | |
| TC | 14.4 | 28.0 | 1.8 | 0.182 | |
| TN | 3.3 | 6.4 | 0.4 | 0.652 | |
| TP | 4.3 | 8.3 | 0.5 | 0.492 | |
| 灌木层 | pH | 0.7 | 1.1 | 0.1 | 0.822 |
| Shrub layer | SBD | 21.4 | 30.7 | 4.2 | 0.040 |
| SWC | 3.6 | 5.2 | 0.7 | 0.452 | |
| ST | 3.6 | 5.1 | 0.7 | 0.412 | |
| SS | 17.7 | 25.4 | 2.8 | 0.100 | |
| EC | 4.4 | 6.3 | 0.9 | 0.414 | |
| TC | 2.9 | 4.2 | 0.6 | 0.516 | |
| TN | 6.3 | 9 | 1.3 | 0.262 | |
| TP | 9.1 | 13.0 | 1.9 | 0.188 | |
| 草本层 | pH | 2.5 | 3.6 | 0.6 | 0.516 |
| Herb layer | SBD | 3.6 | 5.2 | 0.9 | 0.378 |
| SWC | 3.4 | 4.9 | 0.8 | 0.410 | |
| ST | 43.7 | 62.6 | 10.1 | 0.010 | |
| SS | 0.4 | 0.6 | < 0.1 | 0.888 | |
| EC | 2.1 | 3.1 | 0.5 | 0.514 | |
| TC | < 0.1 | < 0.1 | < 0.1 | 1.000 | |
| TN | 9.1 | 13 | 2.3 | 0.128 | |
| TP | 4.9 | 7.0 | 1.3 | 0.288 |
在灌木层上, 前两个排序轴对洲中央林地灌木层植物群落叶功能性状的解释率分别为51.46%、15.74%, 累计解释率达67.20%, 前两轴能较好反映了灌木层植物群落叶功能性状和土壤因子的相关性, 且主要由RDA1轴决定, 排序结果科学。蒙特卡洛检验表明, 土壤容重的P值小于0.05, 是显著(P<0.05)影响洲中央林地灌木层植物群落叶功能性状变化的土壤因子。土壤容重与比叶面积、叶全磷含量等8个叶功能性状均呈负相关关系。
在草本层上, 前两个排序轴对洲中央林地草本层植物群落叶功能性状的解释率分别为68.70%、0.86%, 累计解释率达69.56%, 前两轴能较好反映了草本层植物群落叶功能性状和土壤因子的相关性, 且主要由RDA1轴决定, 排序结果科学。蒙特卡洛检验表明, 土壤温度的P值小于0.05, 是显著(P<0.05)影响洲中央林地植物草本层群落叶功能性状变化的土壤因子。土壤温度与叶干物质含量、叶面积和比叶重呈正相关关系, 与比叶面积、叶片含水量、叶全氮含量和叶全磷含量呈负相关关系。
综上, 土壤容重和土壤温度是影响洲中央林地植物群落叶功能性状的主要土壤因子。
3 讨论 3.1 不同生境植物群落叶功能性状的变化规律本研究结果表明, 在乔木层上, 不同生境的8个群落叶功能性状均无显著性差异(P>0.05), 这表明江心洲的生境差异对乔木层植物群落叶功能选择影响不大。在灌木层上, 洲中央林地植物群落的叶面积显著(P<0.05)高于洲岸泥滩和洲岸沙滩。具有较大叶面积的植物有助于加强植物冠层截留雨水的能力, 表明洲中央林地采取“快速投资收益型策略”, 进而增强叶片保水能力以抵抗干旱胁迫[34]。这表明洲中央林地的灌木层植物群落更倾向于增大叶面积以减少叶厚度碳结构而增加叶面积碳投资的策略, 进而为合成碳水化合物提供能量[35], 同时也提高了截留雨水的能力以适应干旱胁迫的生境。这与焦亮等[36]对内陆河湿地芦苇叶功能性状的研究结果保持一致。在草本层上, 洲中央林地草本层植物群落的比叶面积显著(P<0.05)高于洲岸沙滩, 而洲岸泥滩的比叶面积与这两者均无显著(P>0.05)差异。绝大多数草本植物属于林下植物, 对光照等资源的响应更为敏感, 而乔木层的空间郁闭度较高, 导致草本层植物缺乏光照, 在一定程度上抑制了草本层植物的生长[37—38]。这表明洲中央林地草本层植物群落通过提高比叶面积, 即采取“快速投资收益型策略”以应对光照资源较为缺乏的环境, 此结果与前人的研究结果相似[39]。
3.2 不同生境的植物群落叶功能性状对土壤的响应叶功能性状受到植物种类、环境条件等多方面的综合影响, 因而影响植物叶功能性状的主导因子会有所差异[40]。科儿沁沙地植物群落叶功能性状的变化受到不同生境异质性土壤的影响[41], 在不同生境条件下, 影响植物群落叶功能性状变化的主导因子有所不同, 本研究结果也得出了相同的结论。在同一生境中, 植物之间存在空间、资源等竞争, 不同层次的植物对土壤水分、养分的需求也不尽相同[42], 因而植物叶功能性状对土壤的响应机制也会有差异。在本研究中, 影响同一生境江心洲乔灌草层植物群落叶功能性状的主要土壤因子也有所差异。
在洲岸沙滩, 土壤全碳含量是显著(P<0.05)影响洲岸沙滩乔木层植物群落叶功能性状变化的土壤因子, 其与叶片含水量、叶全氮含量和叶全碳含量等叶功能性状均呈负相关关系。洲岸沙滩的土壤全碳含量显著(P<0.05)低于洲岸泥滩和洲中央林地, 这表明洲岸沙滩乔木层植物通过增加叶片含水量、叶全氮含量和叶全碳含量等群落叶功能性状, 以适应土壤碳元素含量较低的生境[35]。此外, 土壤温度是显著(P<0.05)影响洲岸沙滩灌木层植物群落叶功能性状变化的土壤因子, 这与张丽苗等[43]对重庆主城河岸带植物群落功能性状的研究结果一致。洲岸沙滩虽靠近闽江流域, 但其土壤为沙土, 土壤温度日变化明显, 因此洲岸沙滩灌木层群落叶功能性状受到土壤温度的影响较大。土壤温度与叶片含水量和叶干物质含量等8个群落叶功能性状均呈正相关关系, 表明洲岸沙滩灌木层植物通过增加叶干物质含量、叶全碳含量、叶全氮含量和叶全磷含量等群落叶功能性状, 以适应该生境。
在洲岸泥滩, 土壤含水量和土壤全碳含量是显著(P<0.05)影响洲岸泥滩乔木层植物群落叶功能性状变化的土壤因子, 土壤全氮含量、土壤容重和土壤含水量是显著(P<0.05)影响洲岸泥滩草本层植物群落叶功能性状变化的土壤因子。这表明土壤含水量、土壤容重和土壤养分对洲岸泥滩植物群落叶功能性状的影响较大, 这与张丽苗等[43]的研究结果一致。土壤含水量越高, 土壤颗粒之间的间隙就越大, 土壤的疏松程度就越高, 土壤容重就越小[44]。与洲岸沙滩和洲中央林地相比, 洲岸泥滩的土壤含水量最大, 土壤容重最小, 土壤的紧实度也就最高, 因而土壤容重和土壤含水量对洲岸泥滩植物群落叶功能性状影响较大。此外, 植物群落叶功能性状与土壤养分存在很大关系[45], 土壤碳氮元素对植物的生长发育有调控作用[46]。在本研究中, 洲岸泥滩的土壤全碳含量和全氮含量虽然处于最高水平, 但洲岸泥滩的主要优势种为小蜡和枫杨, 不耐瘠薄, 因而对土壤养分的需求较高。因此, 土壤全碳含量、全氮含量对洲岸泥滩植物群落叶功能性状的影响较大。有研究表明, 在高养分土壤中生长的植物通常会表现出比叶面积增大的特征[4]。本研究发现, 洲岸泥滩土壤养分较高, 乔木层植物表现为较高的比叶面积, 即采取“快速投资收益型策略”;而草本植物表现为较低的比叶面积和叶面积, 即采取“缓慢投资收益型策略”, 这表明在同一生境的不同生活型的植物会采取不同的适生策略。
在洲中央林地, 土壤容重和土壤温度分别是显著(P<0.05)影响灌木层和草本层植物群落叶功能性状变化的主要土壤因子。土壤容重是测定土壤紧实度的敏感指标, 与土壤的疏松程度密切相关, 与土壤含水量极显著(P<0.01)负相关[44]。据调查显示, 洲中央林地部分土地受到人类、牛和羊等圈养动物的踩踏, 导致部分土壤板结情况严重。相对草本植物而言, 灌木植物的根系更深, 因而土壤容重成为显著(P<0.05)影响洲中央林地灌木层植物群落叶功能性状变化的主要土壤因子。洲中央林地灌木层植物通过降低比叶面积、叶全碳含量和叶全磷含量等群落叶功能性状, 以适应土壤紧实和透气性差的生境。此外, 土壤温度升高, 会导致土壤水分蒸发, 进一步影响植物的生长发育[47], 而草本植物的根系一般较浅, 更易受到土壤温度影响。与洲岸沙滩和洲岸泥滩相比, 洲中央林地相对远离闽江流域, 土壤表层温度受太阳光照等影响较大, 其土壤温度最高且变化最大, 因而成为影响洲中央林地草本层植物群落叶功能性状的主导因子。洲中央林地的草本层植物通过采取增加叶面积和叶干物质含量, 降低叶片含水量、叶全氮含量和叶全磷含量, 以适应土壤温度较高的生境。
综上所述, 在局域小尺度上, 显著(P<0.05)影响不同生境植物群落叶功能性状变化的主要土壤因子有所差异。土壤全碳含量和土壤温度是显著(P<0.05)影响洲岸沙滩植物群落叶功能性状变化的土壤因子, 土壤含水量、土壤容重、土壤全碳含量和土壤全碳含量是显著(P<0.05)影响洲岸泥滩植物群落叶功能性状变化的土壤因子, 土壤容重和土壤温度是显著(P<0.05)影响洲中央林地植物群落叶功能性状变化的土壤因子, 这反映了植物对不同生境的适生策略。
4 结论本研究以闽江福州河段5个江心洲的植物群落为研究对象, 探讨洲岸沙滩、洲岸泥滩和洲中央林地的植物群落叶功能性状的差异及其对土壤的响应机制。结果表明, 江心洲3种不同生境的土壤理化性质存在显著差异, 与洲岸泥滩和洲中央林地相比, 受河流水动力和潮汐侵蚀作用最大的洲岸沙滩的土壤含盐率、土壤电导率、土壤全碳含量、土壤全氮含量和土壤全磷含量显著减少, 而土壤容重显著增加。江心洲不同生境的植物群落叶功能性状存在显著差异, 灌木层中央林地的叶面积和草本层的比叶面积显著高于洲岸洲泥滩和洲岸沙滩, 而乔木层的群落叶功能性状不存在显著差异, 这表明, 在局域小尺度上, 江心洲不同生境湿地植物群落的物种多样性及叶功能性状与土壤的关系不会保持恒定, 可能会随着环境因子的变化而改变, 这多与环境因子的筛选作用有关。本研究揭示了江心洲不同生境植物群落叶功能性状和土壤理化性质的差异, 这对于进一步了解江心洲湿地植物群落的驱动机制以及物种的适生策略具有重要意义。此外, 该研究还为江心洲植被的修复、重建和管理提供理论依据, 有助于维护敏感而又脆弱的湿地生态系统。
| [1] |
罗恬, 俞方圆, 练琚愉, 王俊杰, 申健, 吴志峰, 叶万辉. 冠层垂直高度对植物叶片功能性状的影响: 以鼎湖山南亚热带常绿阔叶林为例. 生物多样性, 2022, 30(5): 4-17. |
| [2] |
刘晓娟, 马克平. 植物功能性状研究进展. 中国科学: 生命科学, 2015, 45(4): 325-339. |
| [3] |
Sun S C, Jin D M, Shi P L. The leaf size-twig size spectrum of temperate woody species along an altitudinal gradient: an invariant allometric scaling relationship. Annals of Botany, 2006, 97(1): 97-107. DOI:10.1093/aob/mcj004 |
| [4] |
Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T L, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets Ü, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428: 821-827. DOI:10.1038/nature02403 |
| [5] |
Garnier E, Cortez J, Billès G, Navas M L, Roumet C, Debussche M, Laurent G, Blanchard A, Aubry D, Bellmann A, Neill C, Toussaint J P. Plant functional markers capture ecosystem properties during secondary succession. Ecology, 2004, 85(9): 2630-2637. DOI:10.1890/03-0799 |
| [6] |
Gong H D, Cui Q J, Gao J. Latitudinal, soil and climate effects on key leaf traits in northeastern China. Global Ecology and Conservation, 2020, 22: e00904. DOI:10.1016/j.gecco.2020.e00904 |
| [7] |
何雅琴, 史晓洁, 陈国杰, 赖敏英, 曾纪毅, 魏凯, 邓传远. 滨柃叶功能性状对环境因子的响应. 生态学报, 2022, 42(6): 2418-2429. DOI:10.5846/stxb202101180189 |
| [8] |
Wright I, Reich P, Cornelissen J, Falster D, Groom P, Hikosaka K, Lee W, Lusk C, Oleksyn J, Osada N, Poorter H, Warton D, Westoby M. Modulation of leaf economic traits and trait relationships by climate. Global Ecology and Biogeography, 2005, 14: 411. DOI:10.1111/j.1466-822x.2005.00172.x |
| [9] |
Westoby M. A leaf-height-seed (LHS) plant ecology strategy scheme. Plant and Soil, 1998, 199(2): 213-227. DOI:10.1023/A:1004327224729 |
| [10] |
Xi N X, Adler P B, Chen D X, Wu H Y, Catford J A, van Bodegom P M, Bahn M, Crawford K M, Chu C J. Relationships between plant-soil feedbacks and functional traits. Journal of Ecology, 2021, 109(9): 3411-3423. DOI:10.1111/1365-2745.13731 |
| [11] |
Joswig J S, Wirth C, Schuman M C, Kattge J, Reu B, Wright I J, Sippel S D, Rüger N, Richter R, Schaepman M E, van Bodegom P M, Cornelissen J H C, Díaz S, Hattingh W N, Kramer K, Lens F, Niinemets Ü, Reich P B, Reichstein M, Römermann C, Schrodt F, Anand M, Bahn M, Byun C, Campetella G, Cerabolini B E L, Craine J M, Gonzalez-Melo A, Gutiérrez A G, He T H, Higuchi P, Jactel H, Kraft N J B, Minden V, Onipchenko V, Peñuelas J, Pillar V D, Sosinski Ê, Soudzilovskaia N A, Weiher E, Mahecha M D. Climatic and soil factors explain the two-dimensional spectrum of global plant trait variation. Nature Ecology & Evolution, 2022, 6(1): 36-50. |
| [12] |
邓绶林. 地学辞典. 石家庄: 河北教育出版社, 1992: 465.
|
| [13] |
卢洋. 漓江江心洲植被演替及其修复机制研究[D]. 北京: 北京林业大学, 2016.
|
| [14] |
Saunders A, Norton D A. Ecological restoration at Mainland Islands in New Zealand. Biological Conservation, 2001, 99(1): 109-119. DOI:10.1016/S0006-3207(00)00192-0 |
| [15] |
郭晓英, 陈兴伟, 陈莹, 王跃峰. 气候变化与人类活动对闽江流域径流变化的影响. 中国水土保持科学, 2016, 14(2): 88-94. |
| [16] |
高会, 李家兵, 何涛, 孙志高, 范爱连, 祝贺, 任鹏, 翟水晶. 闽江河口典型植被群落带及交错带植物-土壤体系中硅素的分布特征. 水土保持学报, 2017, 31(1): 279-285. |
| [17] |
傅赐福, 董剑希, 刘秋兴, 于福江. 闽江感潮河段潮汐-洪水相互作用数值模拟. 海洋学报, 2015, 37(7): 15-21. |
| [18] |
漆光超. 漓江水陆交错带植物群落分类的研究[D]. 桂林: 广西师范大学, 2018.
|
| [19] |
丁月萍. 桂林市叠彩区湿地植物群落生态学研究[D]. 桂林: 广西师范大学, 2020.
|
| [20] |
余健, 廖媛媛, 张平究, 高超. 长江下游江心洲土壤颗粒特征及分形规律. 生态学杂志, 2012, 31(3): 626-631. |
| [21] |
中华人民共和国环境保护部. 生物多样性观测技术导则陆生维管植物: HJ 710.1—2014[S]. 北京: 中国环境科学出版社, 2015: 2.
|
| [22] |
中国植物志编辑委员会. 中国植物志. 北京: 科学出版社, 2004.
|
| [23] |
福建植物志编辑委员会. 福建植物志. 福州: 科学技术出版社, 1982.
|
| [24] |
马金双, 李惠茹. 中国外来入侵植物名录. 北京: 高等教育出版社, 2018: 2-176.
|
| [25] |
田岳梨. 秦岭山脊油松针叶功能性状特征及其对土壤和海拔的响应[D]. 杨凌: 西北农林科技大学, 2021.
|
| [26] |
段永峰. 贺兰山东麓植物叶片性状对环境因子的响应[D]. 银川: 宁夏大学, 2021.
|
| [27] |
张增可, 吴雅华, 黄柳菁, 刘兴诏. 海岛森林不同演替阶段土壤和植物的碳、氮、磷化学计量特征. 西北植物学报, 2019, 39(5): 925-934. |
| [28] |
李俐蓉. 甘南高寒草甸不同演替阶段植物叶功能性状变化特征及其与环境因子的关系[D]. 西北师范大学, 2019.
|
| [29] |
黄端, 王冬梅, 任远, 覃云斌, 吴林川. 漓江水陆交错带植物叶性状对水淹胁迫的响应及经济谱分析. 生态学报, 2017, 37(3): 750-759. DOI:10.5846/stxb201508281789 |
| [30] |
张增可, 郑心炫, 林华贞, 林欣, 黄柳菁. 海岛植物不同演替阶段植物功能性状与环境因子的变化规律. 生态学报, 2019, 39(10): 3749-3758. DOI:10.5846/stxb201807051468 |
| [31] |
苏闯, 张芯毓, 马文红, 赵利清, 梁存柱. 贺兰山灌丛群落物种多样性海拔格局及环境解释. 山地学报, 2018, 36(5): 699-708. |
| [32] |
高杰. 中国东北部森林群落功能性状与生物多样性的时空变化[D]. 北京: 北京林业大学, 2020.
|
| [33] |
龚时慧. 环境与系统发育背景对延河流域植物群落功能性状的影响[D]. 杨凌: 西北农林科技大学, 2012.
|
| [34] |
郄亚栋, 蒋腊梅, 吕光辉, 杨晓东, 王恒方, 滕德雄. 温带荒漠植物叶片功能性状对土壤水盐的响应. 生态环境学报, 2018, 27(11): 2000-2010. |
| [35] |
McDowell N, Pockman W T, Allen C D, Breshears D D, Cobb N, Kolb T, Plaut J, Sperry J, West A, Williams D G, Yepez E A. Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought?. The New Phytologist, 2008, 178(4): 719-739. DOI:10.1111/j.1469-8137.2008.02436.x |
| [36] |
焦亮, 关雪, 刘雪蕊, 董小刚, 李方. 内陆河湿地芦苇叶功能性状特征及其对土壤环境因子的响应. 干旱区研究, 2020, 37(1): 202-211. |
| [37] |
高钰, 李旭, 苏兰莛, 高润梅. 山西省二郎山国家森林公园主要森林群落植物多样性研究. 林业资源管理, 2019(5): 150-156. |
| [38] |
曹晓栋, 杨波, 黄梅, 余爽, 余登利, 白新祥. 贵州省宽阔水国家级自然保护区草本植物区系及物种多样性研究. 西北植物学报, 2021, 41(9): 1559-1569. |
| [39] |
Hulshof C M, Violle C, Spasojevic M J, McGill B, Damschen E, Harrison S, Enquist B J. Intra-specific and inter-specific variation in specific leaf area reveal the importance of abiotic and biotic drivers of species diversity across elevation and latitude. Journal of Vegetation Science, 2013, 24(5): 921-931. |
| [40] |
魏晨, 张小平, 罗子渝, 曹建军, 冯明铭, 赵慧君, 李空明, 李光栋. 兰州市南山和北山3种乔木叶片生态化学计量特征的对比研究. 生态学报, 2021, 41(6): 2460-2470. DOI:10.5846/stxb201909061861 |
| [41] |
Zhang J, Zuo X A, Zhou X, Lv P, Lian J, Yue X Y. Long-term grazing effects on vegetation characteristics and soil properties in a semiarid grassland, Northern China. Environmental Monitoring and Assessment, 2017, 189(5): 216. |
| [42] |
苏天成, 向琳, 陈聪琳, 喻静, 张瀚文, 李婧, 王琴, 王芳, 郝建锋. 营林措施对成都绕城高速路域杨树人工林物种多样性和土壤理化性质的影响. 生态学杂志, 2023, 42(1): 58-66. |
| [43] |
张丽苗, 谭雪, 董智, 郑杰, 袁中勋, 李昌晓. 重庆主城河岸带植物群落功能性状与土壤理化性质的关系. 生态学报, 2023, 43(5): 1892-1901. DOI:10.5846/stxb202202160368 |
| [44] |
梁士楚, 苑晓霞, 卢晓明, 许桂芬, 覃艳, 姜勇. 漓江水陆交错带土壤理化性质及其分布特征. 生态学报, 2019, 39(8): 2752-2761. DOI:10.5846/stxb201805091026 |
| [45] |
孙乐, 王毅, 李洋, 孙建. 青藏高原高寒草地群落叶片功能性状对降水的非线性响应. 生态学报, 2023, 43(2): 756-767. DOI:10.5846/stxb202111023080 |
| [46] |
戚德辉, 温仲明, 杨士梭, 王红霞, 郭茹. 黄土丘陵区森林草原带不同退耕年限植物群落和土壤N, P化学计量特征. 草地学报, 2016, 24(2): 322-329. |
| [47] |
李永华, 罗天祥, 卢琦, 田晓娅, 吴波, 杨恒华. 青海省沙珠玉治沙站17种主要植物叶性因子的比较. 生态学报, 2005, 25(5): 994-999. |