 2024, Vol. 44
2024, Vol. 44文章信息
- 刘颖, 宋蕊, 王效科, 高艺芹, 冯美琪, 翟欣月, 万五星
- LIU Ying, SONG Rui, WANG Xiaoke, GAO Yiqin, FENG Meiqi, ZHAI Xinyue, WAN Wuxing
- 罗马直立生菜生长发育及营养指标对臭氧浓度升高和持续熏蒸的响应
- Response of Lactuca sativa var. roman growth and nutritional indicators to elevated ozone concentration and continuous fumigation
- 生态学报. 2024, 44(14): 6276-6287
- Acta Ecologica Sinica. 2024, 44(14): 6276-6287
- http://dx.doi.org/10.20103/j.stxb.202311162492
-
文章历史
- 收稿日期: 2023-11-16
- 网络出版日期: 2024-05-11





2. 中国科学院生态环境研究中心城市与区域生态国家重点实验室, 北京 100085
2. State Key Laboratory of Urban and Regional Ecology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China
地面O3是挥发性有机物、一氧化碳、氮氧化物、甲烷(VOCs、CO、NOX、CH4)等经过复杂的光化学反应形成的一种二次空气污染物[1]。由于人类活动和工业化的增加, 地面O3浓度在过去的几十年以来不断增加。地面O3被认为是一个严重的问题, 它会通过破坏各种生理过程和植物生长, 对植被造成直接影响[2]。目前, 中国的地面O3浓度远远高于美国和欧盟等发达国家[3]。在过去的几十年里, 随着空气质量控制的加强, 北美和欧洲的人为O3排放已经下降, 但是随着中国工业、交通和城市化的快速发展, O3前体(NOX、VOC等)的排放量迅速增加[4]。监测数据显示, 2013—2017年, 31个省会城市8 h的O3平均浓度从61.6 nmol/mol增加到76.0 nmol/mol[5]。由于我国的O3浓度正在逐年升高, 对于保护环境和探讨O3对植物的伤害已经成为了一个不可避免的话题。已有研究表明, 高O3浓度诱导敏感的植物产生各种有害反应, 如叶片可见伤害, 光合作用降低[6], 气孔开闭迟缓, 代谢障碍, 细胞程序化死亡[7], 加速衰老和削弱对病虫害的防御能力等[8-9]。高浓度O3使油菜(Brassica napus)出现失水、褪绿、枯斑、叶脉扭曲、叶片凹凸不平、植株矮化等伤害症状[10], 油菜的株高、叶面积、光合速率、经济产量和生物产量随O3浓度升高呈下降趋势[11]。在O3对冬小麦(Triticum aestivum)、水稻(Oryza sativa)和油菜的影响研究中发现, 冬小麦的相对产量与O3暴露量之间存在高度相关性, 水稻和油菜为中度相关性[12]。地面O3浓度升高显著降低了油菜的种子产量和出油率[13]。O3浓度升高对冬小麦和大豆(Glycine max)的各项生长指标均具有明显的抑制作用, 而且随着O3熏气浓度的升高和熏气时间的持续加重, 作物的株高、叶面积和生物量降低[14]。植物长期暴露在高浓度O3中, O3可以通过气孔进入叶片, 再与质外体接触后分子降解为活性氧(ROS)[15]。这些高反应性的ROS会攻击脂质和蛋白质等细胞成分, 并诱导一系列涉及多种植物生长调节剂的信号通路[16-17]。生物量产量减少、叶片萎蔫和坏死等症状以及代谢改变是植物对O3暴露的典型反应。在本研究中, 通过O3熏蒸实验, 探讨O3浓度升高及暴露时间延长对罗马直立生菜形态学及营养指标的影响, 以期为O3污染加剧背景下我国蔬菜的优质高产乃至食品安全提供科学依据和理论基础。
1 材料与方法 1.1 实验地点与供试植物本实验研究地点是山东省淄博市(36°9′N, 118°22′E), 位于山东省的中部, 淄博地处暖温带, 属于半湿润半干旱的大陆性气候。供试作物为罗马直立生菜(Lactuca sativa var. roman), 罗马直立生菜是生菜的一个品种, 该品种具有生长速度快、产量高、抗逆性强、易于栽培、口感好的特点, 所以被广泛种植。2021年8月10日, 将生菜种子人工种植在育苗盆内, 每穴2—3粒种子, 进行前期育苗;8月21日, 生菜开始育苗;9月10日, 将长势良好、均一的罗马直立生菜幼苗移栽至开顶式气室(OTC);9月30日, 试熏气, 进行设备调试;10月6日, 开始熏蒸, 每天熏蒸8 h(9:00—17:00);11月6日, 熏蒸结束, 累计熏气天数32 d(10月9日和10月25日因下雨所以不熏蒸)。整个O3熏蒸期间定时定量给生菜浇水, 并控制病虫害。
1.2 OTC气室本实验采用OTC、通风系统、O3发生和浓度控制系统、O3自动监测系统组成的O3熏蒸系统。一共设置4种O3浓度, 分别是未过滤的环境空气(NF)、环境空气+40 nmol/mol(NF40)、环境空气+80 nmol/mol(NF80)、环境空气+120 nmol/mol(NF120), 每个处理设置了3个重复组, 12个OTC气室[18]。
1.3 监测及采样 1.3.1 叶片取样及株高测定实验开始熏蒸后, 每天对各OTC内植株进行观察直到第一次观察到叶片出现黄色斑点, 症状通常在近轴面且不会穿透叶片组织[19], 其叶脉不受影响且无病虫害[20]。伤害症状首次出现后, 每7 d记录植株叶片的伤害情况并用相机记录下伤害症状, 明显观测到不同处理组间受伤害的特征变化。在实验期间, 在熏蒸第10天、20天、30天对生菜株高进行测量。
1.3.2 生物量和蔬菜营养指标测定在O3熏气过程中, 分别在熏气10 d、20 d、30 d时随机选取正常生长的3株植株进行生物量取样, 计算各叶片样本的含水量。
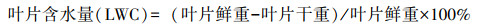
|
(1) |
在O3熏蒸32 d结束时, 采集4种O3熏蒸浓度处理组的生菜叶片进行营养指标检测, 其中测定的营养指标有脂肪、蛋白质、磷(P)、钙(Ca)、钠(Na)、钾(K)、铁(Fe)、锌(Zn)、镁(Mg), 检测依据分别为食品安全国家标准GB 5009.6—2016、GB 5009.5—2016(第一法)、GB 5009.87—2016(第一法)、GB 5009.92—2016(第一法)、GB 5009.91—2017(第一法)、GB 5009.91—2017(第一法)、GB 5009.90—2016(第一法)、GB 5009.14—2017(第三法)、GB 5009.241—2017(第一法)。
1.3.3 叶片解剖结构观察本研究采用指甲油印迹法采集气孔印迹样品, 自然风干后用显微镜进行观察、拍照。拍照时, 随机选择3个视野, 每个视野拍3张照片。使用ImageJ软件记录气孔数量和测量气孔的长度(a)、气孔宽度(b), 气孔长度为平行于保卫细胞的最长值, 气孔宽度为垂直于保卫细胞的最宽值[21]。计算气孔密度(SD), 气孔密度即为气孔数(M, mm2), 气孔开度(SA)按以下公式计算[22]。
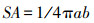
|
(2) |
式中, a为气孔长度,b为气孔宽度。
将采集的新鲜叶片用含70%乙醇的FAA混合固定液(主要由乙醇、乙酸、甲醛组成)固定后, 制成蜡封装片。用OPLYMUS CX 31光学显微镜观察叶片纵切面并拍摄显微图像, 图像在ImageJ软件中进行分析, 并测量以下指标:叶片厚度(BT)、栅栏组织厚度(PT)和海绵组织厚度(ST)。每个样品测量3个样本, 每个样本测量9个视野, 计算平均值。按以下公式计算栅栏组织与海绵组织的比值(P/S)。

|
(3) |
O3暴露剂量AOT40(小时O3浓度大于40 nmol/mol的累积O3暴露值)的计算公式如下:
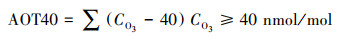
|
(4) |
式中, CO3为太阳总辐射大于50 w/m2时的小时平均O3浓度。
应用Excel 2010和SPSS 26.0进行数据统计和分析, 用SPSS 26.0进行单因素方差分析(One-way ANOVA)和显著性检验。计算各采样时间的O3暴露剂量AOT40(每小时O3浓度超过40 nmol/mol阈值的累计值)[23]。利用R4.3.2软件对叶片解剖结构指标的影响因素进行主成分分析(PCA), 并通过Factoextra程序包的fviz_pca_var函数进行可视化图绘制。采用GraphPad Prism 8建立O3暴露剂量AOT40与各指标的响应关系模型, 以此来评价不同浓度O3对罗马直立生菜影响随暴露时间延长的变化。
2 结果与分析 2.1 熏蒸实验O3条件整个熏蒸期间开顶式气室OTC内的O3日平均浓度变化如下, 其中NF组平均O3浓度为35.23 nmol/mol, NF40组平均O3浓度为73.88 nmol/mol, NF80组平均O3浓度为107.30 nmol/mol, NF120组平均O3浓度为139.31 nmol/mol。NF、NF40、NF80、NF120实验组中的AOT40浓度分别为1.34 μmol mol-1 h-1、9.56 μmol mol-1 h-1、18.58 μmol mol-1 h-1、27.27 μmol mol-1 h-1(图 1)。

|
| 图 1 实验期间O3实际浓度及暴露剂量AOT40 Fig. 1 Actual concentration of O3 and AOT40 during the experiment NF: 环境O3浓度;NF40: 环境O3浓度+40 nmol/mol;NF80: 环境O3浓度+80 nmol/mol;NF120: 环境O3浓度+120 nmol/mol; AOT40: 每小时O3浓度超过40 nmol/mol阈值的累计值 |
O3引起的植物叶片损伤程度是检测地区O3污染状况有效的手段。通过OTC熏蒸实验发现, 罗马直立生菜叶片的O3伤害症状特征是首先在叶片尖端出现浅棕色或黄色斑点, 随着O3浓度的增加, 斑点向叶片内部蔓延, 受害面积同时扩大, 但是叶片的叶脉仍然保持绿色, 无伤害症状。随着熏蒸时间的延长, NF80和NF120实验组叶片出现褪绿, 伴随大面积的坏死斑块(图 2), 这些叶片伤害症状表明O3熏蒸会对植物产生伤害, 长时间的O3熏蒸还会产生累积效应, 加速植株衰老。

|
| 图 2 O3胁迫下的罗马直立生菜叶片可见伤害症状 Fig. 2 Visible injury symptoms on Lactuca sativa var. roman leaves under O3 stress |
随着O3浓度的增加, 罗马直立生菜叶片的受伤害程度也会增加, NF组未观察到受伤害叶片, NF120组的受伤害叶片占比数量最多(表 1)。分析表明, 随着熏蒸时间延长, NF120组的受伤害叶片数量逐渐增多, 在熏蒸28 d后NF120组的受伤害叶片占比为79.79%, 是熏蒸第7天和第14天的6.4倍和1.6倍, 说明高浓度的O3的熏蒸对叶片的伤害有累积效应。
| 时间 Time/d |
受害叶片占比Percentage of injured foliar/% | |||
| NF | NF40 | NF80 | NF120 | |
| 7 | 0±0b | 0.00±0b | 9.54±2.99a | 12.44±3.57a |
| 14 | 0±0c | 0.00±0c | 46.53±1.76b | 50.79±1.61a |
| 28 | 0±0d | 11.59±1.60c | 37.41±2.23b | 79.79±1.14a |
| NF: 环境O3浓度;NF40: 环境O3浓度+40 nmol/mol;NF80: 环境O3浓度+80 nmol/mol;NF120: 环境O3浓度+120 nmol/mol; 表格数据为平均值±标准差(N=3), 同行数值后不同小写字母表示差异显著(P<0.05), 反之则差异不显著 | ||||
不同O3浓度对生菜株高的影响和随熏蒸时间的变化如图 3所示, O3污染抑制罗马直立生菜的生长。生菜在实验初期NF、NF40、NF80、NF120中的株高分别为18.75 cm、15.21 cm、12.94 cm、12.63 cm, 在实验熏蒸30 d结束时株高分别为32.35 cm、27.82 cm、26.70 cm、25.13 cm。实验结束时, NF40、NF80、NF120组的生菜株高分别比NF组下降了14.0%、17.5%、22.3%(P < 0.05)。研究表明, O3浓度升高抑制植物的生长。

|
| 图 3 O3对株高的影响 Fig. 3 Effects of O3 on plant height 不同小写字母表示差异显著(P<0.05) |
O3通过扩散过程通过气孔进入叶片, 损害细胞膜造成细胞内物质外渗[24]。O3胁迫下叶片气孔密度随熏蒸时间的延长产生变化, NF、NF40、NF80组气孔密度伴随熏蒸时间延长逐渐增加, NF120组气孔密度伴随熏蒸时间延长逐渐减少;随O3熏蒸浓度的增加气孔密度逐渐增加(图 4)。O3胁迫下叶片气孔开度发生变化, 在熏气10 d时发现气孔开度随着熏气浓度的升高而增加, 在熏气20 d和30 d时气孔开度都随着熏气浓度的升高而减小(图 4)。在熏气30 d时NF40、NF80、NF120的气孔开度分别比NF下降14.8%、20.6%、32.3%, NF与NF120组存在显著差异(P<0.05)。从整个熏蒸时期来看, NF和NF40组气孔开度随熏气时间的延长变大。NF80和NF120实验组的气孔开度随熏气时间延长先减小再增加, 但不超过熏气10 d时的气孔开度, 表明叶片对O3胁迫的适应有一定范围, 对O3的敏感性随暴露时间延长而降低。

|
| 图 4 O3胁迫对气孔密度和气孔开度的影响 Fig. 4 Effects of O3 stress on stomatal density and stomatal opening |
植物的叶片对空气中水分、温度、污染物等环境因子的变化比其他器官敏感, 叶片最能体现出植物对环境变化的响应[25]。对罗马直立生菜叶片纵切面结构的分析表明, 栅栏组织由2—3层均匀排列的长柱形细胞组成, 细胞间的空隙较大, 海绵组织细胞形状多成不规则排布的圆形或椭圆形。随O3浓度的增加, 栅栏细胞排列结构出现变化, 排列疏松。这种变化先从栅栏薄壁组织内层开始, 然后向周围蔓延。随O3浓度的升高, 在叶肉内部还可看到出现了大面积的空区, 周围有降解的细胞壁, 叶绿体的数量也会减少(图 5)。

|
| 图 5 生菜叶片纵切图 Fig. 5 Longitudinal section of Lactuca sativa var. roman leaf |
叶片厚度在整个熏蒸期间的变化, 在熏蒸30 d结束时NF组叶肉厚度为47.02 μm, 与NF40、NF80、NF120相比均有差异显著水平(P < 0.05), 减少了10.12%、28.69%、28.22%(表 2)。
| 熏蒸天数 Fumigation days/d |
NF | NF40 | NF80 | NF120 | |
| 10 | 47.81±2.80a | 41.34±1.21b | 32.65±3.40c | 34.12±2.90c | |
| BT/μm | 20 | 37.58±2.29a | 35.61±1.10a | 35.54±6.06a | 37.74±2.75a |
| 30 | 47.02±0.62a | 42.26±7.70ab | 33.53±3.14b | 33.75±1.27b | |
| 10 | 23.40±1.84a | 18.59±0.24b | 13.73±1.58c | 13.41±1.31c | |
| PT/μm | 20 | 15.85±0.76a | 16.31±0.99a | 15.67±3.71a | 15.89±1.52a |
| 30 | 20.45±0.02a | 19.18±3.98a | 15.48±1.23a | 15.27±0.89a | |
| 10 | 24.42±1.41a | 22.76±1.08a | 18.91±1.83b | 20.71±1.88ab | |
| ST/μm | 20 | 21.72±1.54a | 19.30±0.13a | 19.87±5.33a | 21.86±1.40a |
| 30 | 26.57±0.60a | 23.09±3.75ab | 18.05±2.00b | 18.48±0.42b | |
| 10 | 0.96±0.07a | 0.82±0.04b | 0.72±0.02bc | 0.65±0.05c | |
| P/S | 20 | 0.73±0.02b | 0.85±0.05a | 0.80±0.03ab | 0.73±0.05b |
| 30 | 0.77±0.02a | 0.82±0.05a | 0.86±0.05a | 0.83±0.03a | |
| BT:叶片厚度Blade thickness;PT:栅栏组织厚度Palisade tissue thickness;ST:海绵组织厚度Sponge tissue thickness;P/S: 栅栏组织/海绵组织Palisade tissue/Sponge tissue;表格数据为平均值±标准差(N=3);同行数值后不同小写字母表示差异显著(P<0.05) | |||||
随着O3暴露剂量的增加, 叶片厚度、栅栏组织厚度、海绵组织厚度逐渐减小, 叶片厚度、海绵组织与AOT40间成显著负相关关系(P<0.05), 栅栏组织与AOT40间关系不显著(图 6)。

|
| 图 6 叶片厚度、栅栏组织厚度、海绵组织厚度与AOT40的关系 Fig. 6 Relationship between blade thickness, palisade tissue thickness, spongy tissue thickness and AOT40 |
通过KMO和Barlett检验得到, KMO的值为0.539(>0.5), 显著性水平P < 0.05, 表明这些变量之间存在共线性问题, 可以进行主成分分析。通过图 7可以知道2个主成分的贡献率达到了78.3%, 已经可以较好的概括了各个指标的大部分的信息。绝对载荷值的大小反应了它对主成分贡献的多少和代表性的强弱[26]。本次研究第一主成分的特征值为3.125, 贡献率是55.4%, 其中绝对载荷值排在前三位的是PT、BT、ST(表 3), 叶肉细胞内栅栏组织中含有丰富的叶绿体进行光合作用, 主成分1主要反应了O3气体进入细胞内部通过影响光合作用来抵抗O3浓度的变化。第二主成分的特征值2.289, 贡献率是22.9%, 其中绝对载荷值前两位的是SA、P/S, 表明气孔是O3进入叶肉细胞的通道。

|
| 图 7 O3对叶片结构指标影响的主成分分析 Fig. 7 Principal component analysis of the effect of O3 on leaf structure indexes |
| 指标 Index |
主成分Principal component | 指标 Index |
主成分Principal component | |||
| 主成分1 | 主成分2 | 主成分1 | 主成分2 | |||
| PT | 0.986 | 0.019 | SA | -0.266 | 0.711 | |
| ST | 0.823 | 0.524 | SD | -0.606 | 0.221 | |
| BT | 0.950 | 0.281 | 特征值Feature value | 3.125 | 2.289 | |
| P/S | 0.579 | -0.681 | 贡献率Contribution/% | 55.421 | 22.872 | |
| SA:气孔开度Stomatal aperture;SD:气孔密度Stomatal density | ||||||
PCA分析图可观察到PT和BT在x轴的投影较大, 在y轴上投影较小, 说明这两种指标对第一个主成分轴的贡献高, 而PT和BT的夹角较小, 说明它们之间密切相关且是正相关关系。综合上述分析可看出, 抵抗O3胁迫最主要的场所是叶肉组织, 叶肉组织的分化程度也可用P/S来评估(图 7)。
2.5 生物量对O3浓度升高及暴露时间延长的响应气孔在吸收O3后叶片内部的氧化会导致细胞死亡, 生态系统生产力和碳储存量会随着时间的推移而降[27], 会损失植物的生物量和产量[28-29]。对植株生物量的研究发现, 随O3浓度升高, 罗马直立生菜各器官的生物量都呈下降趋势(图 8)。与NF组相比较, 熏气30 d时, NF40、NF80、NF120组根干重分别降低了1.29%、22.13%、28.37%, 无显著差异;茎叶干重降低了8.32%、16.96%、36.98%, 其中NF与NF120组存在显著差异(P < 0.05)。研究表明, O3浓度越高植物生长受到的危害越严重。

|
| 图 8 各器官生物量随O3浓度和熏蒸时间的变化 Fig. 8 Changes in biomass of each organ with O3 concentration and fumigation time |
10 d时, 茎叶含水量随O3浓度升高而减少, 20 d时含水量先增加后减少, 30 d时无显著变化;随O3熏蒸时间增加, NF、NF40、NF80、NF120组的茎叶含水量都是增加的趋势(图 9)。

|
| 图 9 O3对茎叶含水量的影响 Fig. 9 Effect of O3 on water content in stems and leaves |
罗马直立生菜叶片中脂肪含量随浓度的升高而增加, 与NF相比, NF80、NF120组分别增长了1.3倍、2.1倍。蛋白质含量随浓度的升高先增加后减少, 与NF相比, NF40、NF80、NF120组的含量变化为19.6%、14.9%、12.5%, 无显著差异。Na含量随浓度的增加而减少, NF80与NF相比降低了23.6%, 无显著差异, NF120组降低了50.8%(P < 0.05)。与NF相比, NF40、NF80、NF120组的Ca含量变化为-16.5%、12.8%、-37.3%;K含量变化为-5.50%、53.3%、3.50%, 无显著差异;Fe含量变化为-29.4、-10.5%、-31.3%, 无显著差异;Zn含量变化为-9.10%、4.7%、-35.9%(P < 0.05);Mg含量变化为-14.1%;-8.50%;-34.2%(P < 0.05)(表 4)。
| 脂肪 Fat (mg/g) |
蛋白质 Protein/(mg/g) |
钙 Ca/(mg/kg) |
钠 Na/(mg/kg) |
钾 K/(mg/g) |
铁 Fe/(mg/kg) |
锌 Zn/(mg/kg) |
镁 Mg/(mg/kg) |
|
| NF | 1.3±0.47b | 8.47±0.78a | 711.33±92.92ab | 17.80±2.26a | 1.99±0.43a | 42.33±7.88a | 5.16±0.37a | 308.00±33.26a |
| NF40 | 1.3±0.47b | 10.13±0.87a | 594.33±8.18ab | 17.80±1.93a | 1.88±0.48a | 29.87±3.83a | 4.69±1.05ab | 264.67±33.26ab |
| NF80 | 3.0±0.82a | 9.73±1.20a | 802.67±220.40a | 13.60±2.08a | 3.05±0.92a | 37.90±7.59a | 5.40±0.87a | 281.67±54.74ab |
| NF120 | 4.0±0.82a | 9.53±1.77a | 446.33±38.66b | 8.76±1.89b | 2.06±0.35a | 29.10±2.66a | 3.31±0.32b | 202.67±18.91b |
| 表格数据为平均值±标准差(N=3);同列数值后不同小写字母表示差异显著(P < 0.05) | ||||||||
伴随社会经济发展, O3已经成为重要的空气污染物[30], 虽然植物对于大气污染物具有一定的脱毒能力, 但城市大气环境中较高的污染物也会影响其生理和形态[31]。了解植物的应对机制, 先从容易观察的表观叶片特征开始探讨。叶片是O3对植物毒性作用的第一个目标, 这种气体污染物通过开放的气孔进入叶片表面, 已有研究针叶树物种的O3损伤表现为轻度烧伤或者褪绿斑驳, 褪绿斑驳为黄色或浅绿色的离散斑块[32]。本实验罗马直立生菜叶片出现褐色点状斑点, 黄色斑块, 随时间延长症状越明显, 这可能是O3在叶片内质外体中迅速离解, 导致活性氧过量[33-35]。株高可以直接反映O3对罗马直立生菜作物生长的影响, 本实验高浓度O3对生菜株高产生抑制作用且这种抑制作用随熏蒸时间延长会减小。
气孔虽然不是O3进入作物的唯一途径, 但却是最主要的途径[37]。在熏气过程中, NF120组的气孔密度总是高于其他实验组, 这可能是植物一开始应对高浓度O3胁迫产生的应激反应, 高浓度O3会导致大范围叶片损伤, 更大的叶片O3损伤与更大的气孔密度呈正相关[38], 这可能增加气孔导度使O3进入到叶片中[39]。本实验在熏气10 d时发现气孔开度随着熏气浓度的升高而增加, 这可能是植物一开始应对高浓度O3胁迫产生的应激反应, 在熏气20 d和30 d时气孔开度都随着熏气浓度的升高而减小, 这可能是随熏蒸时间延长叶片气孔为了减少O3气体的进入, 导致开度减小。高浓度O3引发的气孔密度的增加和气孔开度的减少可以被视为一种规避策略, 旨在减少O3通过气孔进入叶片的流量, 保护光合机制免受氧化应激的影响[40]。O3优先影响栅栏组织细胞, 可能因为这些细胞比海绵组织细胞表现更高的表面积比, 较大的栅栏细胞在细胞膜水平上可能代表O3破坏作用的较大目标[41]。
O3胁迫对于作物生菜生产力的抑制是多种因素共同作用的结果。Ainsworth的气室综合分析表明[42], 与过滤空气相比, 62 nmol/mol的O3浓度使水稻产量降低了约15%(n=18)。另有报告称O3浓度每增加10 nmol/mol, 水稻产量就会下降4.5%[43]。其他作物也有研究, 例如将油菜暴露于高浓度O3环境下6 h, 发现O3暴露增加了种子败育和豆荚内的早熟萌发, 减少了成熟种子数量[44], O3处理降低了茄子(Solanum melongena)的鲜重和干重, O3诱导的生物量减少可能是降低了作物对养分和水分的吸收能力[45]。对流层O3对蓖麻(Ricinus communis)品种的荚膜产量、种子产量和收获指数均有负面影响[46]。在本研究中, 与NF组相比较, NF40、NF80、NF120组生物量下降了5.90%、14.99%、39.21%(P < 0.05), 与之前的研究结果一致。
除了作物形态与产量方面的负面影响外, 探讨暴露在高浓度O3对作物营养指标的影响也非常重要, 因为这与人类的身体健康息息相关。在O3浓度增加的情况下, 水稻大部分金属元素含量呈现增加趋势, 可能是作物抗逆响应[47]。O3暴露显著增加了小麦籽粒中N、P、K、Ca、Mn和Zn元素浓度[48]。本研究中, 罗马直立生菜叶片Ca、Na、Fe、Zn、Mg含量下降, 而K含量增加, 影响脂肪和蛋白质含量增加。在O3胁迫的情况下, Pleijel等[48]证实了这一原理, 他报告了暴露在O3中的小麦的粮食产量和谷物蛋白质浓度之间的负相关性, 作物产量的减少导致合成蛋白质增多。
4 结论随着O3浓度升高, 罗马直立生菜的生长发育受到了抑制, 叶片受伤害程度明显加剧, 株高降低及平均增长率减小, 生物量下降。O3对叶肉中的栅栏组织破坏较明显, 其次是海绵组织, 叶肉厚度、海绵组织厚度与O3暴露剂量AOT40间呈显著负相关关系(P < 0.05)。生菜中营养元素含量的变化是其面对O3产生的调节与适应。基于这个OTC实验, 得出结论, 尽管暴露在高O3浓度下的生菜中, 蛋白质和脂质这两种重要的宏观营养素的浓度增加, 但O3污染对人类营养的总体影响是负面的。这是因为在大多数情况下, 粮食产量的损失抵消了营养指标的增加。
| [1] |
Cao T H, Wang H C, Li L, Lu X, Liu Y M, Fan S J. Fast spreading of surface ozone in both temporal and spatial scale in Pearl River Delta. Journal of Environmental Sciences: China, 2024, 137: 540-552. DOI:10.1016/j.jes.2023.02.025 |
| [2] |
Emberson L D, Pleijel H, Ainsworth E A, van den Berg M, Ren W, Osborne S, Mills G, Pandey D, Dentener F, Büker P, Ewert F, Koeble R, Van Dingenen R. Ozone effects on crops and consideration in crop models. European Journal of Agronomy, 2018, 100: 19-34. DOI:10.1016/j.eja.2018.06.002 |
| [3] |
易睿, 王亚林, 张殷俊, 史宇, 李名升. 长江三角洲地区城市臭氧污染特征与影响因素分析. 环境科学学报, 2015, 35(8): 2370-2377. |
| [4] |
Feng Z Z, Hu E Z, Wang X K, Jiang L J, Liu X J. Ground-level O3 pollution and its impacts on food crops in China: A review. Environmental Pollution, 2015, 199: 42-48. DOI:10.1016/j.envpol.2015.01.016 |
| [5] |
Zeng Y Y, Cao Y F, Qiao X, Seyler B C, Tang Y. Air pollution reduction in China: Recent success but great challenge for the future. The Science of the Total Environment, 2019, 663: 329-337. DOI:10.1016/j.scitotenv.2019.01.262 |
| [6] |
Chaudhary I J, Rathore D. Effects of ambient and elevated ozone on morphophysiology of cotton (Gossypium hirsutum L.) and its correlation with yield traits. Environmental Technology & Innovation, 2022, 25: 102146. |
| [7] |
Mina U, Smiti K, Yadav P. Thermotolerant wheat cultivar (Triticum aestivum L. var. WR544) response to ozone, EDU, and particulate matter interactive exposure. Environmental Monitoring and Assessment, 2021, 193(6): 318. DOI:10.1007/s10661-021-09079-x |
| [8] |
Fiscus E L, Booker F L, Burkey K O. Crop responses to ozone: uptake, modes of action, carbon assimilation and partitioning. Plant, Cell & Environment, 2005, 28(8): 997-1011. |
| [9] |
Kangasjärvi J, Jaspers P, Kollist H. Signalling and cell death in ozone-exposed plants. Plant, Cell & Environment, 2005, 28(8): 1021-1036. |
| [10] |
Han Y J, Gharibeshghi A, Mewis I, Förster N, Beck W, Ulrichs C. Plant responses to ozone: Effects of different ozone exposure durations on plant growth and biochemical quality of Brassica campestris L. ssp. chinensis. Scientia Horticulturae, 2020, 262: 108921. DOI:10.1016/j.scienta.2019.108921 |
| [11] |
白月明, 王春乙, 刘玲, 郭建平, 温民. O3浓度增加对油菜影响的诊断试验研究. 应用气象学报, 2002, 13(3): 364-370. |
| [12] |
姚芳芳, 王效科, 逯非, 冯宗炜, 欧阳志云. 臭氧对农业生态系统影响的综合评估: 以长江三角洲为例. 生态毒理学报, 2008, 3(2): 189-195. |
| [13] |
De Bock M, Op de Beeck M, De Temmerman L, Guisez Y, Ceulemans R, Vandermeiren K. Ozone dose-response relationships for spring oilseed rape and broccoli. Atmospheric Environment, 2011, 45(9): 1759-1765. DOI:10.1016/j.atmosenv.2010.12.027 |
| [14] |
曹嘉晨, 郑有飞, 赵辉, 徐静馨. 地表臭氧浓度升高对冬小麦和大豆生长和产量的影响. 生态毒理学报, 2017, 12(2): 129-136. |
| [15] |
Bohler S, Sergeant K, Lefèvre I, Jolivet Y, Hoffmann L, Renaut J, Dizengremel P, Hausman J F. Differential impact of chronic ozone exposure on expanding and fully expanded poplar leaves. Tree Physiology, 2010, 30(11): 1415-1432. DOI:10.1093/treephys/tpq082 |
| [16] |
Pang J, Kobayashi K, Zhu J G. Yield and photosynthetic characteristics of flag leaves in Chinese rice (Oryza sativa L.) varieties subjected to free-air release of ozone. Agriculture, Ecosystems & Environment, 2009, 132(3/4): 203-211. |
| [17] |
Ansari N, Agrawal M, Agrawal S B. An assessment of growth, floral morphology, and metabolites of a medicinal plant Sida cordifolia L. under the influence of elevated ozone. Environmental Science and Pollution Research, 2021, 28(1): 832-845. DOI:10.1007/s11356-020-10340-y |
| [18] |
高艺芹, 王效科, 万五星, 冯美琪, 张丹红, 耿春梅, 殷宝辉, 张楠, 宋蕊, 刘颖. 臭氧浓度升高对小白菜叶片生理特性的影响随暴露时间的变化研究. 农业环境科学学报, 2023, 42(7): 1454-1464. |
| [19] |
Singh A A, Fatima A, Mishra A K, Chaudhary N, Mukherjee A, Agrawal M, Agrawal S B. Assessment of ozone toxicity among 14 Indian wheat cultivars under field conditions: growth and productivity. Environmental Monitoring and Assessment, 2018, 190(4): 190. DOI:10.1007/s10661-018-6563-0 |
| [20] |
Yang L X, Wang Y L, Shi G Y, Wang Y X, Zhu J G. Responses of rice growth and development to elevated near-surface layer ozone (O3) concentration: a review. Ying Yong Sheng Tai Xue Bao=the Journal of Applied Ecology, 2008, 19(4): 901-910. |
| [21] |
李品, 冯兆忠, 尚博, 袁相洋, 代碌碌, 徐彦森. 6种绿化树种的气孔特性与臭氧剂量的响应关系. 生态学报, 2018, 38(8): 2710-2721. DOI:10.5846/stxb201705220945 |
| [22] |
陈波. 臭氧胁迫对4种树木生长状况和生理特性的影响[D]. 北京: 北京林业大学, 2018.
|
| [23] |
Fuhrer J, Skärby L, Ashmore M R. Critical levels for ozone effects on vegetation in Europe. Environmental Pollution, 1997, 97(1/2): 91-106. |
| [24] |
Laisk A, Kull O, Moldau H. Ozone concentration in leaf intercellular air spaces is close to zero. Plant Physiology, 1989, 90(3): 1163-1167. |
| [25] |
纪若璇, 于笑, 常远, 沈超, 白雪卡, 夏新莉, 尹伟伦, 刘超. 蒙古莸叶片解剖结构的地理种源变异及其对环境变化响应的意义. 植物生态学报, 2020, 44(3): 277-286. |
| [26] |
李娟霞, 田青, 李娇珍, 朱星臻, 何洪盛, 杨秀秀. 兰州市不同空气环境下9种园林植物叶片解剖结构特征. 生态环境学报, 2020, 29(11): 2189-2198. |
| [27] |
Ainsworth E A, Yendrek C R, Sitch S, Collins W J, Emberson L D. The effects of tropospheric ozone on net primary productivity and implications for climate change. Annual Review of Plant Biology, 2012, 63: 637-661. |
| [28] |
McGrath J M, Betzelberger A M, Wang S W, Shook E, Zhu X G, Long S P, Ainsworth E A. An analysis of ozone damage to historical maize and soybean yields in the United States. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(46): 14390-14395. |
| [29] |
Tai A P K, Martin M V, Heald C L. Threat to future global food security from climate change and ozone air pollution. Nature Climate Change, 2014, 4: 817-821. |
| [30] |
栗泽苑, 杨雷峰, 华道柱, 方镜尧, 黄伟, 孙雷, 王琛. 2013-2018年中国近地面臭氧浓度空间分布特征及其与气象因子的关系. 环境科学研究, 2021, 34(9): 2094-2104. |
| [31] |
杜忠, 陶玲, 任珺, 侯培强. 不同污染区对绿化树种可溶性糖含量的影响. 环境科学与技术, 2007, 30(9): 21-23. |
| [32] |
Sicard P, Hoshika Y, Carrari E, De Marco A, Paoletti E. Testing visible ozone injury within a Light Exposed Sampling Site as a proxy for ozone risk assessment for European forests. Journal of Forestry Research, 2021, 32(4): 1351-1359. |
| [33] |
Sandermann H Jr, Ernst D, Heller W, Langebartels C. Ozone: an abiotic elicitor of plant defence reactions. Trends in Plant Science, 1998, 3(2): 47-50. |
| [34] |
Wohlgemuth H, Mittelstrass K, Kschieschan S, Bender J, Weigel H J, Overmyer K, Kangasjärvi J, Sandermann H, Langebartels C. Activation of an oxidative burst is a general feature of sensitive plants exposed to the air pollutant ozone. Plant, Cell & Environment, 2002, 25(6): 717-726. |
| [35] |
Singh A A, Ghosh A, Agrawal M, Agrawal S B. Secondary metabolites responses of plants exposed to ozone: an update. Environmental Science and Pollution Research, 2023, 30(38): 88281-88312. |
| [36] |
姚芳芳, 王效科, 陈展, 冯兆忠, 郑启伟, 段晓男, 欧阳志云, 冯宗炜. 农田冬小麦生长和产量对臭氧动态暴露的响应. 植物生态学报, 2008, 32(1): 212-219. |
| [37] |
Ferdinand J A, Fredericksen T S, Kouterick K B, Skelly J M. Leaf morphology and ozone sensitivity of two open pollinated genotypes of black cherry (Prunus serotina) seedlings. Environmental Pollution, 2000, 108(2): 297-302. |
| [38] |
Wieser G, Havranek W M. Ozone uptake in the sun and shade crown of spruce: quantifying the physiological effects of ozone exposure. Trees, 1993, 7(4): 227-232. |
| [39] |
Castagna A, Ranieri A. Detoxification and repair process of ozone injury: from O3 uptake to gene expression adjustment. Environmental Pollution, 2009, 157(5): 1461-1469. |
| [40] |
Giacomo B, Forino L M C, Tagliasacchi A M, Bernardi R, Durante M. Ozone damage and tolerance in leaves of two poplar genotypes. Caryologia, 2010, 63(4): 422-434. |
| [41] |
Ainsworth E A. Rice production in a changing climate: a meta-analysis of responses to elevated carbon dioxide and elevated ozone concentration. Global Change Biology, 2008, 14(7): 1642-1650. |
| [42] |
Van D T H, Ishii S, Oanh N T K. Assessment of ozone effects on local rice cultivar by portable ozone fumigation system in Hanoi, Vietnam. Environmental Monitoring and Assessment, 2009, 155(1): 569-580. |
| [43] |
Black V, Stewart C, Roberts J, Black C. Timing of exposure to ozone affects reproductive sensitivity and compensatory ability in Brassica campestris. Environmental and Experimental Botany, 2011, 75: 225-234. |
| [44] |
Li Y, Woo S Y, Lee J K, Kwak M J, Khaine I, Jang J H, Kim H N, Kim J E, Park S H, Kim H D, Lim Y J. Effects of elevated ozone on physiological, biochemical and morphological characteristics of eggplant. Horticulture, Environment, and Biotechnology, 2019, 60(6): 809-820. |
| [45] |
Rathore D, Chaudhary I J. Ozone risk assessment of castor (Ricinus communis L. ) cultivars using open top chamber and ethylenediurea (EDU). Environmental Pollution, 2019, 244: 257-269.
|
| [46] |
方笑堃, 罗小三, 张丹, 吴礼春, 邱丹, 陈志炜, 赵朕. 臭氧污染对水稻生长、产量及矿质金属元素含量的影响. 环境科学, 2020, 41(8): 3797-3803. |
| [47] |
Broberg M C, Hayes F, Harmens H, Uddling J, Mills G, Pleijel H. Effects of ozone, drought and heat stress on wheat yield and grain quality. Agriculture, Ecosystems & Environment, 2023, 352: 108505. |
| [48] |
Pleijel H. Grain protein accumulation in relation to grain yield of spring wheat (Triticum aestivum L.) grown in open-top chambers with different concentrations of ozone, carbon dioxide and water availability. Agriculture, Ecosystems & Environment, 1999, 72(3): 265-270. |