 2024, Vol. 44
2024, Vol. 44文章信息
- 赵欣鑫, 刘普, 高捷, 李鹏飞, 齐文博, 翟翊博, 刘智峰
- 基于文献计量的细菌-生物炭协同修复重金属污染土壤研究进展
- 生态学报. 2024, 44(13): 5882-5892
- http://dx.doi.org/10.20103/j.stxb.202401170149
-
文章历史
- 收稿日期: 2024-01-17
- 采用日期: 2024-05-01

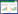




在土壤环境中, 典型重金属污染物包括汞、镉、铅、铜、铬、砷、镍、锰、锌等。土壤中重金属污染物含量超出土壤自净能力后, 会引发严重的土壤污染问题。土壤重金属污染不仅危害农作物的正常生长, 还会诱导地表水和地下水重金属污染, 严重威胁生态环境安全。因此, 重金属污染土壤的有效修复, 对生态系统的持续稳定发展具有重要意义。
土壤作为复杂的环境介质, 基于土壤自净作用对重金属污染物的净化能力相对有限。因此, 为了实现重金属污染土壤的有效修复, 需要人为干预。客土/换土法、稀释法、淋溶法等基于人为干预的传统重金属污染土壤修复技术的修复效能有限, 存在经济成本高、工程难度大、修复周期长等不足。以往研究表明, 在土壤生物修复技术中, 细菌介导的微生物原位修复技术具有低成本、见效快、环境友好等优势[1]。然而, 在细菌介导重金属污染土壤原位修复过程中, 细菌的定殖及土壤修复效率易受土壤含水率、pH、重金属浓度、细菌营养底物等土壤环境条件的影响[2]。因此, 为了提高重金属污染土壤的细菌原位修复效能, 寻求有利于细菌定殖及其对重金属污染土壤有效修复的载体极为必要。
生物炭为秸秆等生物质经高温炭化制备的环境功能材料, 其具有比表面积大、多孔性等特性, 在土壤重金属的吸附和固定中极具潜力[3]。以往研究表明, 生物炭可为细菌提供营养底物和生存载体, 还可调节土壤微环境, 进而为细菌提供适宜的生存环境[4]。因此, 细菌与生物炭对重金属污染土壤的联合修复备受关注。细菌协同生物炭修复重金属污染土壤是一种利用生物炭和细菌的协同作用, 将土壤中的重金属污染物吸附、转化或稳定, 从而降低重金属对环境和生态系统危害的修复技术。该技术具有成本低、效果好、环境友好等优势, 已经成为重金属污染土壤修复领域的研究热点。但是, 细菌-生物炭协同修复重金属污染土壤技术尚未成熟, 细菌与生物炭的相互作用机制及组合形式仍需进一步深入探究。
本研究综述了细菌与生物炭修复重金属污染土壤的研究现状, 分析了细菌-生物炭-重金属的相互作用机制, 阐述了细菌与生物炭在重金属污染土壤修复的应用, 探讨了细菌与生物炭协同修复重金属污染土壤存在的问题及其发展趋势, 提出了强化细菌与生物炭组分调控及细菌与生物炭协同修复技术亟待优化的未来展望, 研究结果可为重金属污染土壤的有效修复及科学管理提供参考。
1 文献计量简述为深入了解细菌-生物炭协同修复重金属污染土壤的研究现状及未来趋势, 本研究以“细菌-生物炭”为研究主题, 以Web of Science核心数据库为数据来源, 设检索式为: (Ts=(biochar)) AND (Ts=(“bacteria” OR “bacterial” OR “bacterium” OR “microbe”)), 检索时间跨度为2018—2023年, 获得国际相关论文2894篇。基于发文量和关键词等检索数据整合分析, 归纳全球对于细菌与生物炭协同修复重金属污染土壤研究的现状与进展。计量分析表明, 在Web of Science收录的文献呈现逐年增长趋势(图 1), 表明细菌-生物炭协同修复重金属污染土壤在世界范围内引起了越来越多的关注。研究论文的被引频次通常可指示研究成果具有的重要学术价值和影响力。计量分析显示, 从2018年至2023年, 相关论文被引频次已经从285上涨至21427, 表明菌炭协同作用在重金属污染土壤修复中的巨大应用潜力。其中, 中国发文量最多(2006篇), 其次为美国(273篇)、澳大利亚(104篇)、巴基斯坦(93篇)、印度(95篇)等(图 2), 表明中国为该领域研究的主体力量, 在细菌协同生物炭修复重金属污染土壤方面取得显著成果。

|
| 图 1 生物炭-细菌的年度研究论文发文量与被引频次 Fig. 1 The number and citation frequency of annual research papers on biochar-bacteria |

|
| 图 2 发文量前十国家数量及占比 Fig. 2 The number and proportion of the top ten countries in terms of publication volume |
基于web of science核心数据库中与细菌-生物炭相关的2894篇论文, 使用VOSviewer确定了生物炭、细菌、重金属、土壤微生物群落、吸附作用、机制等20个关键词(图 3)。生物炭与细菌的相互作用可以促进植物生长, 强化土壤重金属稳定化以减轻作物的重金属胁迫。此外, 作物废弃物可以通过转化为生物炭以可持续的方式进行循环利用, 并可作为土壤肥力的缓效肥料。生物炭与细菌的协同作用可以降低土壤重金属的迁移, 改善土壤微生物群落结构, 促进有益微生物的生长, 提高土壤酶活性, 增强土壤养分循环, 强化重金属污染土壤的有效修复。但是, 关于细菌-生物炭复合体对重金属污染土壤的可持续性修复效能及机制尚未明晰, 仍需进一步深入探究。

|
| 图 3 在Web of Science核心数据库中以细菌-生物炭为主题发表的论文20个关键词 Fig. 3 Twenty keywords were published in the core database of Web of Science with the theme of bacteria-biochar |
生物炭是一种在缺氧或限氧条件下经高温炭化的富含碳的多孔生物质材料, 具有丰富的活性有机官能团和碳芳烃结构, 以及较大的比表面积和负的表面电荷[5—7]。生物炭原料来源广泛, 如农产品、作物残渣、林业残渣、木材废料和城市固体废物等, 主要通过物理吸附、静电吸附、离子交换、络合、沉淀和氧化还原等作用机制固定土壤中的重金属污染物(表 1, 图 4)[8—9]。
| 反应类型 Reaction type |
机制 Mechanism |
应用 Application |
| 物理吸附 Physical adsorption |
在范德华力作用下, 重金属会被吸附在生物炭表面或孔隙中 | 稻秆生物炭在700℃下对Cd和Ni有较好的吸附能力[10] |
| 静电吸附 Electrostatic adsorption |
生物炭的表面电荷通过静电相互作用吸附和固定重金属 | 表面积较大、表面负电荷较低的改性生物炭对Cr(Ⅳ)的去除率较高[11] |
| 离子交换 Ion exchange |
生物炭表面的盐离子可以用重金属离子代替, 对重金属进行固定 | 离子交换在动物源生物炭固定Cd和Cu中占主导地位[12] |
| 络合 Complexation |
生物炭表面官能团提供重金属结合位点, 形成固定重金属的配合物 | 采用巯基乙醇催化酯化法制备巯基改性秸秆生物炭, 其对Cd的吸附量提高了3倍[13—14] |
| 沉淀 Precipitation |
生物炭灰分中的矿物成分可与重金属析出, 起到固定重金属的作用 | 秸秆生物炭中的C2O42-和CO32-可与Pb形成PbC2O4和Pb3(CO3)2(OH)2沉淀, 这是固定Pb的主要机制[15—16] |
| 氧化还原 Oxidation-reduction |
生物炭与重金属之间的电子得失改变了重金属的价态, 降低了重金属的毒性和迁移率 | 生物炭的酚羟基可以作为Cr(Ⅵ)的电子供体, 被氧化成醌基并与吸附的Cr(Ⅵ)络合[17] |

|
| 图 4 生物炭与土壤中重金属的相互作用机理 Fig. 4 The interaction mechanism between biochar particles and heavy metals in soil |
细菌对重金属固定化途径主要包括生物吸附、生物蓄积和生物沉淀等[18—21]。以往研究报道, 分离于工业废水灌溉土壤中的苏云金芽孢杆菌OSM29, 对Cd、Cu、Cr、Ni和Pb具有突出的生物吸附性能[22]。与生物吸附相比, 生物蓄积作用对重金属的吸收周期更长, 该过程与细菌的生化特征、内部结构、遗传和生理能力以及环境条件密切相关[23]。大多蜡样芽孢杆菌、巨型芽孢杆菌等细菌能产生金属硫蛋白, 对重金属具有突出生物积累能力[24]。此外, 细菌可通过催化氧化和还原作用将游离重金属离子转化为生物沉淀, 降低重金属生物利用度和毒性。De等人发现, 抗铅细菌(B. iodinium GP13和B. pumilus S3)诱导铅以硫化铅(PbS)形式沉淀[25]。
细菌修复重金属污染土壤的主要生物机制如下(图 5):(1)细胞外屏障:细胞壁、质膜或荚膜阻止金属离子进入细胞;(2)金属离子的主动转运:从细胞质中输出有毒金属;(3)细胞外螯合作用:细菌在周质、外膜中积累重金属离子或重金属离子作为不溶性化合物发生络合作用;(4)细胞内隔离:基于细胞质内非生物利用形式的重金属的积累[26];(5)离子交换机制。

|
| 图 5 离子交换机制和ATP酶对金属离子的主动转运 Fig. 5 Ion exchange mechanism and active transport of metal ions by ATPase Mn+:重金属阳离子;ATP:腺苷三磷酸;ADP:腺苷二磷酸;P:磷酸基团 |
在生物炭修复重金属土壤实际应用中, 环境中微生物扮演着重要的角色[27]。生物炭与细菌的相互作用主要包括协同、拮抗和相加作用。细菌-生物炭协同修复技术在重金属污染土壤治理中具有重要的应用前景。生物炭具有较大比表面积和丰富的多孔结构, 可以提供大量的吸附位点, 不仅可以作为土壤细菌生存繁殖的碳源, 还可为细菌提供栖息地以减弱细菌种群因定殖场所限制而产生的生存竞争(图 6)。生物炭载体可促进细菌的生长和繁殖, 进而强化生物炭对土壤重金属的捕获和固定。此外, 生物炭可增加土壤的保水性和通气性等, 这有助于细菌的生长和代谢活动, 进而提高土壤的肥力和养分循环。生物炭和细菌的协同作用可以改善土壤环境, 促进植物的生长和发育。植物通过根系吸收养分和水分, 同时也可以吸收固定在生物炭和细菌上的重金属, 从而降低其对植物的毒性。生物炭具有较长的使用寿命, 可以在土壤中长期存在。因此, 生物炭与细菌的协同作用具有可持续修复重金属污染土壤的巨大潜力, 可大大降低重金属的二次污染风险。此外, 鞘氨醇单胞菌等具有重金属降解能力的功能细菌在土壤环境中广泛存在, 但其自身的降解能力有限, 因此, 这些细菌与生物炭联用, 可在重金属固定化中具有相加作用或者协同增效作用的潜力, 进而强化重金属污染土壤的修复。在重金属污染土壤修复中, 生物炭与细菌相加作用的相关报道较少, 其相关作用机制仍有待探索。然而, 也有报道指出, 生物炭的粒径、形态等物理特性会损害细菌细胞, 也会抑制细菌细胞的分裂和生长速度[28]。以往研究基于空间尺度、生物尺度和环境尺度设计实验, 综合研究了生物炭对生物的影响, 结果表明, 较小颗粒(<0.5 mm)的慢热解木屑生物炭可能对土壤某些生物群具有亚致死毒性。此外, 在细菌与吸附材料复合过程中, 细菌可能会占据吸附剂表面的活性反应位点, 进而降低吸附剂的吸附性能, 故而, 在生物炭与细菌联用修复重金属污染土壤中, 细菌与生物炭也可能出现拮抗作用[28], 具体相互作用类型可能因生物炭和细菌的种类、环境条件等因素而异。因此, 在细菌联合生物炭修复重金属污染土壤的实际应用中, 需要考虑生物炭和细菌的使用量及组合方式, 强化生物炭和细菌的协同修复作用, 同时极力削弱或避免细菌与生物炭之间的拮抗作用[29]。

|
| 图 6 生物炭与细菌的协同过程 Fig. 6 The synergistic process of biochar and bacteria |
细菌和生物炭联合修复重金属污染土壤具有良好的环境效益和经济效益。单一生物炭或细菌在重金属修复中的效率相对较低[30], 细菌协同生物炭可以充分结合两者优势, 强化细菌和生物炭对重金属的作用。生物炭-细菌对重金属作用机制主要包括离子交换、固定化作用、氧化还原作用、迁移作用等(图 7)。

|
| 图 7 生物炭-细菌对重金属固定作用机制 Fig. 7 Fixation mechanism of heavy metals by biochar-bacteria a. 离子交换;b. 固定作用;c. 氧化还原作用;d. 迁移作用;Mn+:金属离子;-:电荷;EPS:胞外聚合物 |
细菌和生物炭可以与重金属离子进行离子交换反应, 包括阳离子交换和阴离子交换[31]。细菌表面含有能与重金属进行离子交换的离子, 固定在生物炭上的细菌具有更大的离子交换能力[32]。同样, 生物炭也具有较高的阳离子和阴离子交换能力, 可以介导Na、K、Mg和Ca等碱金属离子和Cd(Ⅱ)、Cr(Ⅲ)、Pb(Ⅱ)带正电的重金属进行离子交换作用[33]。此外, 带负电荷的Cr(Ⅵ)可以与生物炭表面的硫酸根离子进行离子交换反应[34]。
重金属阳离子可以和存在于生物炭和细菌表面的羟基、羧基、磷酰基、氨基等电负性官能团发生络合作用[35—36]。细菌-生物炭对重金属的吸附是生物炭的被动吸附和细菌的主动吸附共同作用的结果[32]。细菌可以分泌多糖、蛋白质和脂质等胞外聚合物, 这些聚合物通常以荚膜或粘液的形式附着在细菌细胞表面, 对重金属具有突出的结合能力[37]。生物炭具有较高的表面电荷, 产生的静电引力和范德华力是生物炭物理截留的重要途径[38]。细菌产生的胞外聚合物可覆盖在生物炭表面, 在静电引力作用下, 吸附在生物炭上的重金属会转移到细菌体内, 强化重金属固定。
在细菌主动吸收或吸附重金属过程中, 通常伴随着氧化还原反应介导的重金属价态的变化[39—41]。以往研究报道, 生物炭表面含有丰富的对苯二酚官能团和持久性自由基, 它们可将Cr(Ⅵ)还原为容易吸附在生物炭表面的Cr(Ⅲ)[42]。在重金属污染土壤环境中, 生物炭可为细菌介导的氧化还原过程提供有利场所, 且持久性自由基可作为电子穿梭体促进细菌对重金属的氧化还原作用[43]。Liu等研究报道, 小麦秸秆生物炭可以促进异养铁还原菌株希瓦氏菌MR-1介导Fe(Ⅲ)生物还原生成Fe(Ⅱ), 随后在生物炭表面形成生物铁矿, 强化Cr(Ⅵ) 的固定[44]。
重金属的迁移性是重金属污染土壤修复的关键影响因素[45]。细菌可产生乙酸、柠檬酸、苹果酸和葡萄糖酸等有机酸, 以及表面活性物质, 这些物质可与重金属发生螯合作用, 从而提高其生物有效性[46]。此外, 甲基杆菌属、假单胞菌属和芽孢杆菌属等功能细菌可促进Hg、Se、Pb等重金属的甲基化, 通过挥发作用释放重金属[47]。
细菌对重金属的迁移作用通常涉及电子的转移, 而且生物炭可充当电子穿梭体促进电子的转移。以往研究报道, 油菜秸秆生物炭促进了细菌诱导的Fe(Ⅲ)还原, 并提高了土壤中硒和镉的迁移性[48]。此外, 生物炭可抑制Cd对沙雷氏菌SNB6的侵害, 同时还可促进金属富集植物(金沙草)的生长, 从而强化植物对富Cd土壤中Cd的生物富集[46]。
4 细菌-生物炭协同修复重金属污染土壤的应用在生态修复工程实际应用中, 基于细菌-生物炭协同修复技术的菌炭联合施用显著优于单一生物炭或单一细菌的修复效能。在细菌协同生物炭修复重金属污染土壤过程中, 生物炭可为细菌生长提供适宜的微环境, 直接或间接影响细菌种群的变化, 从而影响重金属的生物利用度[49]。同时, 细菌可通过改变土壤理化性质, 促进生物炭对土壤重金属的吸附、钝化、稳定等作用。因此, 细菌-生物炭协同修复技术在重金属污染土壤的治理中具有重要的应用前景。以往研究依托中国内蒙古自治区包头市某Pb、Zn、Cr严重超标的重金属污染区块试验田[50], 针对中国北方高盐碱地土壤背景, 利用生物炭协同微生物矿化技术联合修复重金属污染土壤, 结果表明, 土壤中Pb、Zn、Cr的浸出率显著降低, 土壤pH、酶活性、养分含量等土壤性质得到明显改善, 细菌与生物炭的协同作用极大强化了土壤Pb、Zn、Cr的稳定化, 为中国北方高盐碱土壤背景下的复合重金属污染修复提供了科学案例。
细菌-生物炭协同作用不仅可以固定土壤中的重金属, 还可以促进植物生长。以往研究基于芽孢杆菌W25, 发现菌株W25可降低土壤重金属的生物利用度和生菜可食用组织中Cd和铅的含量[51—52], 并且生物炭可促进菌株W25的生长和繁殖, 调节土壤pH值, 进一步提高了菌株W25介导的土壤中Cd和Pb的固定效能。此外, 菌株W25-生物炭-番茄的联合应用, 显著提高了土壤酶活性和链霉菌、芽孢杆菌等重金属固定细菌的相对丰度, 降低了土壤中Cd和Pb的含量以及番茄果实中Cd的积累[53]。还有研究证实了细菌Alcaligenes anensis sp. Nov(NCCP-650)协同生物炭修复镉、铅污染土壤的潜力, 研究指出, 细菌与生物炭的协同作用不仅降低了土壤Cd和Pb的生物有效组分, 而且对植物生长和土壤性质具有积极的促进作用[54]。Wang等人利用玉米秸秆和猪粪生物炭作为枯草芽孢杆菌(B38)突变基因型的载体, 联合细菌与生物炭修复重金属污染土壤, 结果表明, 生物炭显著降低了重金属的生物利用度;生物炭与菌株B38的结合, 强化了生物炭对土壤重金属的稳定化效能;菌株B38和猪粪衍生生物炭的同时施用对促进植物生长和土壤重金属的固定具有优异效果, 植物生物量增加了37.9%, 生菜可食部分中重金属浓度降低了69.9%—96.1%[55]。Wei等人探究了阿氏肠杆菌G3(Enterobacter G3)和肠杆菌I12的生理生化性质及其与生物炭对Cd、Pb复合污染土壤的联合修复效能, 结果表明, 菌株G3和I12均对Cd、Pb具有高度抗性, 且对植物生长具有显著促生作用[56]。Shi等人通过负载耐重金属菌株(Burkholderia contaminans ZCC)于生物炭上, 制备细菌-生物炭复合材料, 结果表明, 施用细菌-生物炭复合材料显着提高了珠芽景天的Cd和Zn积累量, 分别提高了230.13%和381.27%;同时, 细菌-生物炭复合材料通过减少氧化损伤、增加叶绿素和抗氧化酶活性减轻重金属对珠芽景天的毒性[57]。此外, 混合菌与生物炭的协同修复技术对重金属污染土壤的修复也极具潜力。Ji等人采用物理吸附法和海藻酸钠包封法合成了两种负载混合细菌的生物炭, 探究了混合载菌生物炭对土壤As、Pb和Cd稳定化的应用潜力, 结果表明, 混合细菌可在生物炭上定殖, 细菌-生物炭联用技术可通过共沉淀、氧化还原和吸附等作用修复重金属污染土壤[58]。生物炭、细菌作为单一修复剂, 在重金属土壤修复中具有极大局限性, 因此, 发展细菌-生物炭协同联用技术, 对强化重金属污染土壤修复效能和提高修复稳定性能具有重要意义和应用价值。
细菌-生物炭复合体在重金属污染土壤修复的应用中, 细菌的生物状况对细菌协同生物炭修复重金属污染土壤具有重要作用[59]。生物炭通常可为细菌提供碳源和定殖场所, 从而促进细菌的生长和繁殖, 但有些生物炭所携带的有毒物质(多环芳烃、呋喃等)以及高温热解产生的持久性自由基会对微生物产生毒性, 损害细菌细胞, 进而抑制细菌的生长[60—61]。因此, 细菌生长的监测对细菌协同生物炭修复重金属污染土壤具有重要意义。土壤环境中细菌生长情况及变化趋势通常可通过细菌菌落计数、细菌生长曲线绘制、细菌生物量测定、生理指标分析(酶活性、代谢产物等)、分子生物学分析等表征技术手段进行评估。
5 结论与展望生物炭作为一种绿色可持续的环保材料, 原料来源广泛、成本低, 主要通过物理吸附、静电吸附、离子交换、络合、沉淀和氧化还原等重要途径固定土壤中重金属[62]。土壤环境中广泛分布的功能细菌在重金属污染土壤修复中也具有巨大作用, 可基于植物间接修复土壤重金属污染, 也可通过自身的生物吸附、生物蓄积和生物沉淀等途径直接固定重金属。但是, 单一生物炭或细菌对重金属污染土壤的修复存在较大局限性。本研究强调了细菌-生物炭协同修复技术的优势和可行性, 综述了细菌与生物炭通过离子交换、固定作用、氧化还原作用和迁移作用等重要机制实现重金属污染土壤的有效修复, 揭示了细菌-生物炭协同作用在重金属污染土壤修复中的巨大应用价值。但是, 在重金属污染土壤的实际修复中, 通常会面临复杂的复合重金属污染。多环芳烃降解菌和耐砷高效石油烃降解菌等有机物降解菌兼具着突出的重金属固定性能[63—64], 其与生物炭的协同作用对重金属与有机物复合污染土壤的有效修复效能值得进一步深入探究。现有报道中, 大多为单一种类的生物炭与单一细菌的联用, 对土壤重金属的吸附、钝化和稳定等作用存在局限。因此, 优化细菌-生物炭组合形式, 发展混合细菌与多种类生物炭协同技术[65—67], 在复合重金属污染土壤修复中具有极为广阔的应用前景。
诸多研究表明, 细菌与生物炭的协同作用对重金属污染土壤的可持续修复极具潜力, 但生物炭的潜在生态和健康风险也不能忽视, 如, 生物炭合成原料自身富集的重金属、多环芳烃、呋喃等有毒有害物质会威胁土壤微生物群落及土壤环境生态;生物炭高温热解伴生的持久性自由基对土壤微生物的毒害等[68—71]。为规避生物炭生产和应用中的潜在生态健康风险, 生物炭原料的预处理、生物炭制备技术优化及其环境健康风险评估等应对途径亟需探究。此外, 在细菌协同生物炭修复重金属污染土壤的实际应用中, 细菌与生物炭的耦合作用机制、生物炭负载技术优化、菌炭协同效能强化及其影响因素等技术研究, 新型功能细菌选育、生物炭新原料开发、新型细菌与生物炭复合体研发等产品创新研究, 以及细菌与生物炭协同修复技术的应用示范等仍需进一步深入探究。
| [1] |
Dhaliwal S S, Singh J, Taneja P K, Mandal A. Remediation techniques for removal of heavy metals from the soil contaminated through different sources: a review. Environmental Science and Pollution Research International, 2020, 27(2): 1319-1333. DOI:10.1007/s11356-019-06967-1 |
| [2] |
Wu J T, Fu X F, Zhao L, Lv J, Lv S D, Shang J, Lv J X, Du S X, Guo H J, Ma F. Biochar as a partner of plants and beneficial microorganisms to assist in-situ bioremediation of heavy metal contaminated soil. Science of the Total Environment, 2024, 923: 171442-171442. DOI:10.1016/j.scitotenv.2024.171442 |
| [3] |
Yang Q, Wang Y J, Zhong H. Remediation of mercury-contaminated soils and sediments using biochar: a critical review. Biochar, 2021, 3(1): 23-35. DOI:10.1007/s42773-021-00087-1 |
| [4] |
Brichi L, Fernandes J V M, Silva B M, Vizú J D F, Junior J N G, Cherubin M. Organic residues and their impact on soil health, crop production and sustainable agriculture: a review including bibliographic analysis. Soil Use and Management, 2023, 39: 686-706. DOI:10.1111/sum.12892 |
| [5] |
孙进博, 胡玲燕, 李博, 陈建军, 施翠仙, 祖艳群, 李元, 湛方栋. 石灰与生物炭对污染土壤理化性质、万寿菊生长与镉铅含量的影响. 环境化学, 2024, 1(1): 1-11. |
| [6] |
Li X, Zeng J Y, Zuo S, Lin S T, Chen G X. Preparation, modification, and application of biochar in the printing field: a review. Materials, 2023, 16(14): 5081. DOI:10.3390/ma16145081 |
| [7] |
Bashir S, Hussain Q, Akmal M, Riaz M, Hu H Q, Ijaz S S, Iqbal M, Abro S, Mehmood S, Ahmad M. Sugarcane bagasse-derived biochar reduces the cadmium and chromium bioavailability to mash bean and enhances the microbial activity in contaminated soil. Journal of Soils and Sediments, 2018, 18(3): 874-886. DOI:10.1007/s11368-017-1796-z |
| [8] |
Beesley L, Marmiroli M. The immobilisation and retention of soluble arsenic, cadmium and zinc by biochar. Environmental Pollution, 2011, 159(2): 474-480. DOI:10.1016/j.envpol.2010.10.016 |
| [9] |
Al-Wabel M I, Usman A R A, El-Naggar A H, Aly A A, Ibrahim H M, Elmaghraby S, Al-Omran A. Conocarpus biochar as a soil amendment for reducing heavy metal availability and uptake by maize plants. Saudi Journal of Biological Sciences, 2015, 22(4): 503-511. DOI:10.1016/j.sjbs.2014.12.003 |
| [10] |
Deng Y Y, Huang S, Laird D A, Wang X G, Meng Z W. Adsorption behaviour and mechanisms of cadmium and nickel on rice straw biochars in single- and binary-metal systems. Chemosphere, 2019, 218: 308-318. DOI:10.1016/j.chemosphere.2018.11.081 |
| [11] |
Dong H R, Deng J M, Xie Y K, Zhang C, Jiang Z, Cheng Y J, Hou K J, Zeng G M. Stabilization of nanoscale zero-valent iron (nZVI) with modified biochar for Cr(Ⅵ) removal from aqueous solution. Journal of Hazardous Materials, 2017, 332: 79-86. DOI:10.1016/j.jhazmat.2017.03.002 |
| [12] |
Nazari S, Rahimi G, Khademi Jolgeh Nezhad A. Effectiveness of native and citric acid-enriched biochar of Chickpea straw in Cd and Pb sorption in an acidic soil. Journal of Environmental Chemical Engineering, 2019, 7(3): 103064. DOI:10.1016/j.jece.2019.103064 |
| [13] |
Lei S C, Shi Y, Qiu Y P, Che L, Xue C. Performance and mechanisms of emerging animal-derived biochars for immobilization of heavy metals. Science of the Total Environment, 2019, 646: 1281-1289. DOI:10.1016/j.scitotenv.2018.07.374 |
| [14] |
Fan J J, Cai C, Chi H F, Reid B J, Coulon F, Zhang Y C, Hou Y W. Remediation of cadmium and lead polluted soil using thiol-modified biochar. Journal of Hazardous Materials, 2020, 388: 122037. DOI:10.1016/j.jhazmat.2020.122037 |
| [15] |
Shen Z T, Hou D Y, Jin F, Shi J X, Fan X L, Tsang D C W, Alessi D S. Effect of production temperature on lead removal mechanisms by rice straw biochars. Science of the Total Environment, 2019, 655: 751-758. DOI:10.1016/j.scitotenv.2018.11.282 |
| [16] |
Liang Y, Cao X D, Zhao L, Arellano E. Biochar- and phosphate-induced immobilization of heavy metals in contaminated soil and water: implication on simultaneous remediation of contaminated soil and groundwater. Environmental Science and Pollution Research, 2014, 21(6): 4665-4674. DOI:10.1007/s11356-013-2423-1 |
| [17] |
Mohan D, Rajput S, Singh V K, Steele P H, Pittman C U. Modeling and evaluation of chromium remediation from water using low cost bio-char, a green adsorbent. Journal of Hazardous Materials, 2011, 188(1-3): 319-333. DOI:10.1016/j.jhazmat.2011.01.127 |
| [18] |
Efe D. Potential plant growth-promoting bacteria with heavy metal resistance. Current Microbiology, 2020, 77(12): 3861-3868. DOI:10.1007/s00284-020-02208-8 |
| [19] |
Verma C, Singh P, Kumar R. Isolation and characterization of heavy metal resistant PGPR and their role in enhancement of growth of wheat plant under metal (cadmium) stress condition. Archives of Applied Science Research, 2015, 7: 37-43. |
| [20] |
Glick B R. Using soil bacteria to facilitate phytoremediation. Biotechnology Advances, 2010, 28(3): 367-374. DOI:10.1016/j.biotechadv.2010.02.001 |
| [21] |
Burd G I, Dixon D G, Glick B R. Plant growth-promoting bacteria that decrease heavy metal toxicity in plants. Canadian Journal of Microbiology, 2000, 46(3): 237-245. DOI:10.1139/w99-143 |
| [22] |
赵雪, 赵欣鑫, 赵佐平, 杨超, 邱若琪, 丁聪, 杨乐, 陈旭杰. 秸秆生物质炭处理重金属废水研究现状及展望. 生态毒理学报, 2024, 19(1): 31-39. |
| [23] |
Wróbel M, S'liwakowski W, Kowalczyk P, Kramkowski K, Dobrzyński J. Bioremediation of heavy metals by the genus Bacillus. International Journal of Environmental Research and Public Health, 2023, 20(6): 4964. DOI:10.3390/ijerph20064964 |
| [24] |
Murthy S, Geetha B, Sarangi SK. Effect of lead on metallothionein concentration in lead-resistant bacteria Bacillus cereus isolated from industrial effluent. African Journal of Biotechnology, 2011, 10(71): 15966-15972. |
| [25] |
De J, Ramaiah N, Vardanyan L. Detoxification of toxic heavy metals by marine bacteria highly resistant to mercury. Marine Biotechnology, 2008, 10(4): 471-477. DOI:10.1007/s10126-008-9083-z |
| [26] |
González Henao S, Ghneim-Herrera T. Heavy metals in soils and the remediation potential of bacteria associated with the plant microbiome. Frontiers in Environmental Science, 2021, 9: 604216. DOI:10.3389/fenvs.2021.604216 |
| [27] |
刘领, 马宜林, 悦飞雪, 乔鑫鑫, 尹飞, 王艳芳. 生物炭对褐土旱地玉米季氮转化功能基因、丛枝菌根真菌及N2O释放的影响. 生态学报, 2021, 41(7): 2803-2815. |
| [28] |
Prodana M, Silva C, Gravato C, Verheijen F G A, Keizer J J, Soares A M V M, Loureiro S, Bastos A C. Influence of biochar particle size on biota responses. Ecotoxicology and Environmental Safety, 2019, 174: 120-128. DOI:10.1016/j.ecoenv.2019.02.044 |
| [29] |
Ouyang P, Narayanan M, Shi X J, Chen X P, Li Z L, Luo Y M, Ma Y. Integrating biochar and bacteria for sustainable remediation of metal-contaminated soils. Biochar, 2023, 5(1): 63. DOI:10.1007/s42773-023-00265-3 |
| [30] |
Zheng X M, Xu W H, Dong J, Yang T, Shangguan Z C, Qu J, Li X, Tan X F. The effects of biochar and its applications in the microbial remediation of contaminated soil: A review. Journal of Hazardous Materials, 2022, 438: 129557. DOI:10.1016/j.jhazmat.2022.129557 |
| [31] |
Qu J H, Shi J J, Wang Y H, Tong H, Zhu Y J, Xu L S, Wang Y F, Zhang B, Tao Y, Dai X, Zhang H, Zhang Y. Applications of functionalized magnetic biochar in environmental remediation: A review. Journal of Hazardous Materials, 2022, 434: 128841. DOI:10.1016/j.jhazmat.2022.128841 |
| [32] |
Huang F, Li K, Wu R R, Yan Y J, Xiao R B. Insight into the Cd2+ biosorption by viable Bacillus cereus RC-1 immobilized on different biochars: Roles of bacterial cell and biochar matrix. Journal of Cleaner Production, 2020, 272: 122743-122743. DOI:10.1016/j.jclepro.2020.122743 |
| [33] |
Wu J W, Wang T, Wang J W, Zhang Y S, Pan W P. A novel modified method for the efficient removal of Pb and Cd from wastewater by biochar: enhanced the ion exchange and precipitation capacity. Science of the Total Environment, 2021, 754: 142150. DOI:10.1016/j.scitotenv.2020.142150 |
| [34] |
Zhang H, Xiao R, Li R H, Ali A, Chen A L, Zhang Z Q. Enhanced aqueous Cr(Ⅵ) removal using chitosan-modified magnetic biochars derived from bamboo residues. Chemosphere, 2020, 261: 127694. DOI:10.1016/j.chemosphere.2020.127694 |
| [35] |
Ahmad Z, Gao B, Mosa A, Yu H W, Yin X Q, Bashir A, Ghoveisi H, Wang S S. Removal of Cu(Ⅱ), Cd(Ⅱ) and Pb(Ⅱ) ions from aqueous solutions by biochars derived from potassium-rich biomass. Journal of Cleaner Production, 2018, 180: 437-449. DOI:10.1016/j.jclepro.2018.01.133 |
| [36] |
Wang L W, O'Connor D, Rinklebe J, Ok Y S, Tsang D C W, Shen Z T, Hou D Y. Biochar aging: Mechanisms, physicochemical changes, assessment, and implications for field applications. Environmental Science & Technology, 2020, 54(23): 14797-14814. |
| [37] |
Wang Y, Narayanan M, Shi X J, Chen X P, Li Z L, Natarajan D, Ma Y. Plant growth-promoting bacteria in metal-contaminated soil: Current perspectives on remediation mechanisms. Frontiers in Microbiology, 2022, 13: 966226. DOI:10.3389/fmicb.2022.966226 |
| [38] |
Hu F N, Xu C Y, Ma R T, Tu K, Yang J Y, Zhao S W, Yang M Y, Zhang F B. Biochar application driven change in soil internal forces improves aggregate stability: Based on a two-year field study. Geoderma, 2021, 403: 115276. DOI:10.1016/j.geoderma.2021.115276 |
| [39] |
Xia X, Wu S J, Zhou Z J, Wang G J. Microbial Cd(Ⅱ) and Cr(Ⅵ) resistance mechanisms and application in bioremediation. Journal of Hazardous Materials, 2021, 401: 123685. DOI:10.1016/j.jhazmat.2020.123685 |
| [40] |
Xia X, Wu S J, Li N H, Wang D, Zheng S X, Wang G J. Novel bacterial selenite reductase CsrF responsible for Se(Ⅳ) and Cr(Ⅵ) reduction that produces nanoparticles in Alishewanella sp. WH16-1. Journal of Hazardous Materials, 2018, 342: 499-509. DOI:10.1016/j.jhazmat.2017.08.051 |
| [41] |
Giovanella P, Cabral L, Bento F M, Gianello C, Camargo F A O. Mercury (Ⅱ) removal by resistant bacterial isolates and mercuric (Ⅱ) reductase activity in a new strain of Pseudomonas sp. B50A. New Biotechnology, 2016, 33(1): 216-223. DOI:10.1016/j.nbt.2015.05.006 |
| [42] |
Zhang K K, Sun P, Zhang Y R. Decontamination of Cr(Ⅵ) facilitated formation of persistent free radicals on rice husk derived biochar. Frontiers of Environmental Science & Engineering, 2019, 13(2): 22. |
| [43] |
Narayanan M, Ma Y. Influences of biochar on bioremediation/Phytoremediation potential of metal-contaminated soils. Frontiers in Microbiology, 2022, 13: 929730. DOI:10.3389/fmicb.2022.929730 |
| [44] |
Liu L C, Liu G F, Zhou J T, Jin R F. Interaction between hexavalent chromium and biologically formed iron mineral-biochar composites: kinetics, products and mechanisms. Journal of Hazardous Materials, 2021, 405: 124246. DOI:10.1016/j.jhazmat.2020.124246 |
| [45] |
Ma Y, Oliveira R S, Freitas H, Zhang C. Biochemical and molecular mechanisms of plant-microbe-metal interactions: Relevance for phytoremediation. Frontiers in Plant Science, 2016, 7: 918. |
| [46] |
Wu B, Wang Z R, Zhao Y X, Gu Y M, Wang Y, Yu J, Xu H. The performance of biochar-microbe multiple biochemical material on bioremediation and soil micro-ecology in the cadmium aged soil. Science of the Total Environment, 2019, 686: 719-728. DOI:10.1016/j.scitotenv.2019.06.041 |
| [47] |
Bali A S, Sidhu G P S. Arsenic acquisition, toxicity and tolerance in plants-From physiology to remediation: a review. Chemosphere, 2021, 283: 131050. DOI:10.1016/j.chemosphere.2021.131050 |
| [48] |
Lyu C H, Li L, Liu X W, Zhao Z Q. Rape straw application facilitates Se and Cd mobilization in Cd-contaminated seleniferous soils by enhancing microbial iron reduction. Environmental Pollution, 2022, 310: 119818. DOI:10.1016/j.envpol.2022.119818 |
| [49] |
Wu R F, Long M, Tai X S, Wang J L, Lu Y L, Sun X C, Tang D F, Sun L K. Microbiological inoculation with and without biochar reduces the bioavailability of heavy metals by microbial correlation in pig manure composting. Ecotoxicology and Environmental Safety, 2022, 248: 114294. DOI:10.1016/j.ecoenv.2022.114294 |
| [50] |
牛磊, 郑春丽. 生物炭协同微生物矿化技术修复复合重金属污染农田土壤. 有色金属工程, 2023, 13(11): 141-155. DOI:10.3969/j.issn.2095-1744.2023.11.019 |
| [51] |
Wang X H, Dong G Y, Liu X W, Zhang S K, Li C, Lu X Q, Xia T. Poly-γ-glutamic acid-producing bacteria reduced Cd uptake and effected the rhizosphere microbial communities of lettuce. Journal of Hazardous Materials, 2020, 398: 123146. DOI:10.1016/j.jhazmat.2020.123146 |
| [52] |
Wang Y Y, Liu Y D, Zhan W H, Zheng K X, Wang J N, Zhang C S, Chen R H. Stabilization of heavy metal-contaminated soils by biochar: Challenges and recommendations. Science of the Total Environment, 2020, 729: 139060. DOI:10.1016/j.scitotenv.2020.139060 |
| [53] |
Liu X W, Wang X H, Xu T Y, Ma H Z, Xia T. The combined application of γ-PGA-producing bacteria and biochar reduced the content of heavy metals and improved the quality of tomato (Solanum lycopersicum L.). Environmental Science and Pollution Research, 2022, 29(59): 88938-88950. DOI:10.1007/s11356-022-21842-2 |
| [54] |
Khan U, Irfan M, Murad Z, Ahmad I, Khan M O, Mehmood I, Waleed M, Kamal A. Enhancing lettuce growth and cadmium and lead tolerance through biochar and bacteria. Gesunde Pflanzen, 2023, 75(6): 2685-2696. DOI:10.1007/s10343-023-00914-4 |
| [55] |
Wang T, Sun H W, Ren X H, Li B, Mao H J. Evaluation of biochars from different stock materials as carriers of bacterial strain for remediation of heavy metal-contaminated soil. Scientific Reports, 2017, 7(1): 12114. DOI:10.1038/s41598-017-12503-3 |
| [56] |
Wei T, Gao H, An F Q, Ma X L, Hua L, Guo J K. Performance of heavy metal-immobilizing bacteria combined with biochar on remediation of cadmium and lead co-contaminated soil. Environmental Geochemistry and Health, 2023, 45(8): 6009-6026. DOI:10.1007/s10653-023-01605-9 |
| [57] |
Shi A, Hu Y, Zhang X, Zhou D, Xu J L, Rensing C, Zhang L M, Xing S H, Ni W Z, Yang W H. Biochar loaded with bacteria enhanced Cd/Zn phytoextraction by facilitating plant growth and shaping rhizospheric microbial community. Environmental Pollution, 2023, 327: 121559. DOI:10.1016/j.envpol.2023.121559 |
| [58] |
Ji X W, Wan J, Wang X D, Peng C, Wang G H, Liang W Y, Zhang W. Mixed bacteria-loaded biochar for the immobilization of arsenic, lead, and cadmium in a polluted soil system: effects and mechanisms. Science of the Total Environment, 2022, 811: 152112. DOI:10.1016/j.scitotenv.2021.152112 |
| [59] |
张丽, 李如霞, 何玉垒, 姚彦坡, 林大松. 生物炭与氮肥复施对镉污染水稻土修复效应及机制. 环境科学, 2023, 44(8): 4479-4488. |
| [60] |
吴东, 李尚珂, 施维林, 史广宇. 稻壳与稻壳生物炭对镉污染土壤微生物群落结构影响研究. 环境科学学报, 2024, 44(2): 375-385. |
| [61] |
赵淑雯, 张倩茹, 张楚晨, 冀琳宇, OK Yongsik. 土壤污染修复中的生物炭-微生物交互作用研究进展. 农业环境科学学报, 2023, 42(7): 1423-1435. |
| [62] |
孙文君, 袁兴超, 李祖然, 李博, 李元. 三种钝化剂对镉铅污染农田的钝化修复效应研究. 草地学报, 2023, 31(2): 596-608. |
| [63] |
李花, 赵立坤, 包仕钰, 余晓龙, 毛旭辉, 陈超琪. 多环芳烃降解菌及其应用研究进展. 环境工程技术学报, 2023, 13(5): 1663-1676. |
| [64] |
李虹呈, 苏趋, 张武竹, 张耀, 向罗京. 砷胁迫下石油烃降解菌的分离、鉴定及其降解特性. 环境工程, 2023, 41(7): 166-174. |
| [65] |
Iriarte-Velasco U, Sierra I, Zudaire L, Ayastuy J L. Preparation of a porous biochar from the acid activation of pork bones. Food and Bioproducts Processing, 2016, 98: 341-353. DOI:10.1016/j.fbp.2016.03.003 |
| [66] |
Jiao Y Z, Zhang N L, He C, Ma X R, Liu X X, Liu L, Hou T T, Wang Z A, Pan X H. Preparation of sludge-corn stalk biochar and its enhanced anaerobic fermentation. Biochemical Engineering Journal, 2022, 187: 108609. DOI:10.1016/j.bej.2022.108609 |
| [67] |
Mazarji M, Bayero M T, Minkina T, Sushkova S, Mandzhieva S, Bauer T V, Soldatov A, Sillanpää M, Wong M H. Nanomaterials in biochar: Review of their effectiveness in remediating heavy metal-contaminated soils. Science of the Total Environment, 2023, 880: 163330. DOI:10.1016/j.scitotenv.2023.163330 |
| [68] |
He L Z, Zhong H, Liu G X, Dai Z M, Brookes P C, Xu J M. Remediation of heavy metal contaminated soils by biochar: Mechanisms, potential risks and applications in China. Environmental Pollution, 2019, 252: 846-855. DOI:10.1016/j.envpol.2019.05.151 |
| [69] |
Dai S L, Li H, Yang Z L, Dai M W, Dong X J, Ge X, Sun M, Shi L L. Effects of biochar amendments on speciation and bioavailability of heavy metals in coal-mine-contaminated soil. Human and Ecological Risk Assessment, 2018, 24(7): 1887-1900. DOI:10.1080/10807039.2018.1429250 |
| [70] |
Godlewska P, Ok Y S, Oleszczuk P. The dark side of black gold: Ecotoxicological aspects of biochar and biochar-amended soils. Journal of Hazardous Materials, 2021, 403: 123833. DOI:10.1016/j.jhazmat.2020.123833 |
| [71] |
Ibrahim A, Usman A R A, Al-Wabel M I, Nadeem M, Ok Y S, Al-Omran A. Effects of conocarpus biochar on hydraulic properties of calcareous sandy soil: influence of particle size and application depth. Archives of Agronomy and Soil Science, 2017, 63(2): 185-197. DOI:10.1080/03650340.2016.1193785 |