 2024, Vol. 44
2024, Vol. 44文章信息
- 窦艳星, 王宝荣, 廖娇娇, 薛志婧, 侯琳, 安韶山, 张硕新
- DOU Yanxing, WANG Baorong, LIAO Jiaojiao, XUE Zhijing, HOU Lin, AN Shaoshan, ZHANG Shuoxin
- 黄土高原不同人工林型微生物残体碳对土壤有机碳组分的积累贡献及影响因素
- Contributions of microbial necromass carbon to soil organic carbon fractions accumulation and influencing factors under different artificial forest types on the Loess Plateau
- 生态学报. 2024, 44(13): 5494-5507
- Acta Ecologica Sinica. 2024, 44(13): 5494-5507
- http://dx.doi.org/10.20103/j.stxb.202310172260
-
文章历史
- 收稿日期: 2023-10-17
- 网络出版日期: 2024-04-25





2. 西北农林科技大学水土保持科学与工程学院(水土保持研究所), 杨凌 712100;
3. 西北农林科技大学草业与草原学院, 杨凌 712100;
4. 西北农林科技大学资源与环境学院, 杨凌 712100;
5. 陕西师范大学地理科学与旅游学院, 西安 710119;
6. 陕西秦岭森林生态系统国家野外科学观测研究站, 杨凌 712100
2. College of Soil and Water Conservation Science and Engineering, Northwest Agriculture and Forestry University, Yangling 712100, China;
3. College of Grassland Agriculture, Northwest A&F University, Yangling 712100, China;
4. College of Natural Resource and Environment, Northwest A&F University, Yangling 712100, China;
5. School of Geography and Tourism, Shaanxi Normal University, Xi'an 710119, China;
6. Qinling National Forest Ecosystem Research Station, Yangling 712100, China
土壤有机碳(Soil organic carbon, SOC)的来源、形成、转化和稳定机制是深入了解陆地生态系统碳汇功能和应对气候变化的关键[1—2], 微生物在土壤有机质形成与积累过程中发挥着重要作用[3—4]。目前, 越来越多的证据表明微生物残体碳(Microbial necromass carbon, MNC)是土壤稳定性有机碳库的重要组分[5—6]。颗粒态有机碳(Particulate organic carbon, POC)和矿物结合态有机碳(Mineral—associated organic carbon, MAOC)作为土壤有机碳的重要组分, 其形成途径和积累过程能够反应土壤有机碳的不同来源及其对土壤有机碳的贡献[7—8]。微生物死亡、裂解、破碎后与黏土矿物颗粒结合主要形成MAOC[9—10], 而未分解、难分解的植物源碳主要形成POC[11—12], 然而由于植物化学性质及其分解程度的难易不同, 经微生物作用后产生的植物残体碳和微生物残体碳比例也不同[13], 导致微生物源碳对POC和MAOC形成的贡献可能存在差异。已有的研究表明草地中MAOC对土壤有机碳的贡献>50%[14], 而森林土壤中POC在土壤有机碳的积累过程中占主导地位[13], 可见, 微生物源碳对POC和MAOC形成的贡献比例及机制因植被类型而异。森林土壤以凋落物为主要碳源, 影响微生物“碳泵”调控的有机物分解过程, 导致POC和MAOC对总有机碳的贡献比例存在差异[12, 15—16], 而不同森林类型向土壤中输入的有机物不同, 这是否会影响MNC对POC和MAOC的贡献比例还未可知。
黄土高原自退耕还林还草工程实施以来, 人工林地大面积分布且持续贡献土壤有机碳的增加, 目前针对该区土壤有机碳的研究多集中在土壤有机碳储量、土壤有机碳动态、土壤有机碳稳定性变化特征等方面, 并取得了一系列重要成果[17—19], 但该区典型人工林MNC对土壤有机碳组分积累的贡献机制仍不清楚, 而这有助于阐明黄土高原植被恢复中土壤有机碳积累的微生物学机制, 因此有必要针对黄土高原不同人工林型微生物残体碳对土壤POC和MAOC的积累贡献及其影响因素开展研究。鉴于此, 本研究以黄土高原不同人工林型为研究对象, 探究不同人工林型0—60 cm土层土壤有机碳组分(POC和MAOC)、MNC含量变化特征及其对土壤POC和MAOC的积累贡献, 结合土壤理化性质, 辨析影响MNC贡献POC和MAOC积累的主要因素, 揭示黄土高原不同人工林型MNC对POC和MAOC积累的贡献及主要影响因素, 这对深入了解黄土高原土壤有机碳形成与积累的微生物学机制具有重要意义, 同时可为该区“双碳”目标的实现提供理论依据和数据支撑。
1 材料与方法 1.1 研究区概况研究区位于甘肃省庆阳市西峰区杨家沟流域(35°41′28″—35°42′15″ N, 107°33′5″—107°33′30″ E), 该流域位于董志塬腹地, 流域面积约0.66 km2, 以人工林恢复为主, 为典型的黄土高原沟壑区地貌特征, 土壤类型为黄绵土, 土层厚度约250 m, 平均海拔1278.04 m, 气候类型属于温带大陆性半干旱气候, 年日照总时2400—2600 h, 年降水量400—600 mm, 年平均气温10℃, 年无霜期160—180 d, 光照充足, 四季分明, 优势树种包括刺槐(Robinia pseudoacacia)、油松(Pinus tabuliformis)、山杏(Armeniaca sibirica)等[20]。
1.2 试验设计2021年7月在杨家沟流域内选取刺槐(Robinia pseudoacacia)林、山杏(Armeniaca sibirica)林、油松(Pinus tabuliformis)林为研究对象, 开展实验, 每种林型分别设置3个样地, 样地面积为20 m×20 m, 所选人工林种植年限约为67 a, 样地概况见表 1。
| 林型 Forest type |
样地编号 Numbers of plots |
海拔 Altitude/m |
经度 Longitude/(°) |
纬度 Latitude/(°) |
坡向 Aspect |
枯落物厚度 Depth of litter/cm |
主要植物种类 Main plant species |
| CH | 样地1 | 1110 | 107.55 | 35.70 | 阴坡 | 0.5 | 刺槐(Robinia pseudoacacia)、长芒草(Stipa bungeana)、苔草(Carex)、胡枝子(Lespedeza bicolor)、悬钩子(Rubus corchorifolius) |
| 样地2 | 1152 | 107.55 | 35.70 | 阴坡 | 0.7 | 刺槐(Robinia pseudoacacia)、长芒草(Stipa bungeana)、苔草(Carex)、黄蒿(Artemisia scoparia) | |
| 样地3 | 1124 | 107.55 | 35.70 | 阴坡 | 0.8 | 刺槐(Robinia pseudoacacia)、长芒草(Stipa bungeana)、苔草(Carex) | |
| SX | 样地1 | 1128 | 107.57 | 35.69 | 阴坡 | 2.5 | 山杏(Armeniaca sibirica)、胡枝子(Lespedeza bicolor)、苦豆子(Sophora alopecuroides)、早熟禾(Poa annua)、中华隐子草(Cleistogenes chinensis) |
| 样地2 | 1273 | 107.55 | 35.70 | 阴坡 | 1.5 | 山杏(Armeniaca sibirica)、胡枝子(Lespedeza bicolor)、苦豆子(Sophora alopecuroides)、早熟禾(Poa annua)、铁杆蒿(Artemisia gmelinii) | |
| 样地3 | 1303 | 107.55 | 35.70 | 阴坡 | 1.9 | 山杏(Armeniaca sibirica)、胡枝子(Lespedeza bicolor)、苦豆子(Sophora alopecuroides)、早熟禾(Poa annua) | |
| YS | 样地1 | 1303 | 107.55 | 35.71 | 阳坡 | 2.0 | 油松(Pinus tabuliformis)、苔草(Carex)、小蓟(Cirsium setosum)、苦豆子(Sophora alopecuroides)、早熟禾(Poa annua) |
| 样地2 | 1283 | 107.56 | 35.70 | 阳坡 | 1.4 | 油松(Pinus tabuliformis)、苔草(Carex)、小蓟(Cirsium setosum)、苦豆子(Sophora alopecuroides) | |
| 样地3 | 1309 | 107.56 | 35.70 | 阳坡 | 2.1 | 油松(Pinus tabuliformis)、苔草(Carex)、黄蒿(Artemisia scoparia)、早熟禾(Poa annua) | |
| CH: 刺槐林Robinia pseudoacacia;SX: 山杏林Armeniaca sibirica;YS: 油松林Pinus tabuliformis | |||||||
每个样地设置5个灌木样方, 样方面积为2 m×2 m, 然后在每个灌木样方内分别设置1个1 m×1 m的草本样方, 共计5个草本样方(图 1), 先进行植被调查, 再采集土壤样品。

|
| 图 1 试验设计 Fig. 1 Experimental design |
按照“S”型采样法分别采集0—10 cm、10—20 cm、20—40 cm、40—60 cm土壤样品各1 kg, 带回实验室去除植物根系及小石块后过2 mm筛, 自然风干后过0.15 mm筛, 用于测定土壤基本理化性质及POC、MAOC等指标, 同时在测定POC和MAOC的过程中预留出> 0.053 μm和 < 0.053 μm的土样约2—3 g, 用于测定其中氨基糖的含量。
1.3.2 各指标测定方法土壤基本理化性质的测定:土壤容重采用环刀法测定;土壤含水率采用烘干法测定;土壤总有机碳采用重铬酸钾氧化法测定;土壤全氮采用半微量凯氏定氮法测定;土壤全磷采用NaOH—钼锑抗比色法测定[21]。
土壤颗粒态和矿物结合态有机碳的测定:通过粒度分馏方法利用(1.70±0.02) g/cm3的六偏磷酸钠溶液分离, 再用0.5 mol/L的HCl除无机碳后利用碳氮联合分析仪测定[22]。
土壤质地的测定:按照USDA分类系统对粒径的分类要求对黏粒、砂粒及粉粒进行划分, 利用0.5 mol/L的六偏磷酸钠溶液作为分散剂, 通过沉降、比重计法对样品进行测定, 计算黏粒、粉粒及砂粒所占比例[23]。
氨基糖的测定:利用盐酸水解, 经纯化和衍生后利用气相色谱法测定。具体过程:将土壤样品研磨过60目筛, 加入6 mol/L HCl水解、过滤。加入100 μg肌醇后, 调节pH值至6.6—6.8, 离心去除沉淀。上清液用冷冻干燥仪冻干, 残留的固体物质用无水甲醇溶解、离心, 将上清液转移到5 mL衍生瓶中, 用N2吹干溶液, 加入1 mL水, 同时加入100 μg N—甲基氨基葡萄糖(内标2, 用来计算纯化过程的回收率), 摇匀后再次进行冷冻干燥。将氨基糖衍生后, 利用气相色谱进行氨基糖衍生物的分离测定。氨基糖提取液在气相色谱仪(Agilent 7890B GC, Agilent Technologies, Santa Clara, CA, 美国)上进行测定分析。配置HP-5色谱柱(30 m×0.25 mm×0.25 μm)以及火焰离子化检测仪(Flame ionization detector, FID)。1 μL氨基糖提取液进样至色谱柱进行测定, N2作为载气, 流速为0.6 mL/min。气相色谱进样口温度设定为250℃, 分流比为30∶1。参照标准样品中三种氨基糖的出峰时间来分离即可测得土壤氨基糖含量[24—26]。
微生物残体碳(g/kg)含量根据下式进行计算:

|
(1) |

|
(2) |
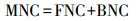
|
(3) |
式中FNC是真菌残体碳(Fungal necromass carbon, FNC), BNC是细菌残体碳(Bacterial necromass carbon, BNC), GluN是氨基葡萄糖含量, MurA是胞壁酸含量, 179.2是氨基葡萄糖的分子质量, 9是真菌氨基葡萄糖转换为真菌残体碳的转换系数, 45是细菌胞壁酸转换为细菌残体碳的转换系数。
不同人工林型土壤基本理化性质如表 2所示。
| 林型 Forest type |
土层深度 Depth of soil layer/cm |
含水率 Water relative content/% |
容重 Bulk density/(g/cm3) |
土壤质地 Soil texture |
pH | 有机碳 Soil organic carbon/(g/kg) |
全氮 Total nitrogen/(g/kg) |
||
| 黏粒占比 Clay/% |
粉粒占比 Silt/% |
砂粒占比 Sand/% |
|||||||
| CH | 0—10 | 11.89±1.38 | 1.09±0.14 | 23.29±3.56 | 37.95±5.54 | 38.76±8.82 | 8.29±0.03 | 13.30±0.57 | 1.36±0.09 |
| 10—20 | 10.83±2.44 | 1.16±0.10 | 20.81±4.59 | 32.39±3.85 | 46.80±8.29 | 8.35±0.03 | 10.10±1.39 | 1.06±0.16 | |
| 20—40 | 9.66±1.18 | 1.14±0.18 | 21.06±2.16 | 35.45±1.64 | 43.49±1.49 | 8.53±0.02 | 6.88±0.87 | 0.71±0.09 | |
| 40—60 | 7.00±3.07 | 1.19±0.15 | 20.00±3.47 | 33.53±0.99 | 46.48±4.36 | 8.55±0.02 | 4.62±0.58 | 0.51±0.06 | |
| SX | 0—10 | 4.65±1.72 | 0.97±0.04 | 22.24±2.44 | 34.12±0.95 | 43.64±1.60 | 8.31±0.09 | 17.39±4.98 | 1.65±0.32 |
| 10—20 | 5.05±0.72 | 1.03±0.07 | 19.19±3.74 | 31.40±1.38 | 49.41±4.89 | 8.39±0.04 | 14.06±5.26 | 1.01±0.12 | |
| 20—40 | 5.00±0.86 | 1.12±0.05 | 15.92±1.97 | 29.71±3.10 | 54.38±5.06 | 8.52±0.02 | 8.38±1.22 | 0.72±0.03 | |
| 40—60 | 8.00±3.49 | 1.16±0.13 | 18.55±3.78 | 33.29±3.56 | 48.16±7.26 | 8.56±0.01 | 5.60±1.95 | 0.56±0.06 | |
| YS | 0—10 | 17.84±0.03 | 1.03±0.03 | 16.99±1.48 | 34.44±1.48 | 48.58±2.96 | 8.88±0.16 | 14.37±1.08 | 1.32±0.12 |
| 10—20 | 11.51±0.04 | 1.11±0.06 | 17.75±2.89 | 32.74±3.84 | 49.51±6.59 | 9.00±0.05 | 8.03±0.69 | 0.83±0.06 | |
| 20—40 | 6.83±0.00 | 1.06±0.06 | 18.42±2.03 | 34.39±1.75 | 47.19±3.14 | 9.00±0.06 | 5.28±1.48 | 0.64±0.14 | |
| 40—60 | 6.23±0.01 | 1.16±0.05 | 18.18±0.63 | 30.13±9.85 | 51.69±10.35 | 9.08±0.04 | 3.89±0.82 | 0.50±0.10 | |
| 表中数据为平均值±标准差(n=3) | |||||||||
采用Excel 2010对原始数据进行统计、计算, 运用Origin2023b对不同人工林型POC和MAOC含量、POC和MAOC中FNC、BNC、MNC含量及其所占比例作图分析, 通过SPSS19.0软件, 采用单因素方差分析方法(One—way ANOVA)对不同人工林型同一土层和同一人工林型不同土层POC和MAOC含量、POC和MAOC中FNC、BNC、MNC含量及其所占比例之间的差异显著性进行分析(P < 0.05), 运用Origin2023b作图软件, 采用Person相关分析法对POC、MAOC、FNC/POC、FNC/MAOC、BNC/POC、BNC/MAOC、MNC/POC、MNC/MAOC与土壤理化性质之间的相关关系进行作图分析(P < 0.05)。
2 结果与分析 2.1 不同人工林型POC、MAOC含量变化特征0—60 cm土层, 三种人工林土壤POC含量均随土层深度的增加而降低, 其中CH林不同土层之间差异均显著(P < 0.05), SX林仅0—10 cm土层与10—60 cm土层之间差异显著(P < 0.05), 而YS林除20—40 cm土层与40—60 cm土层外, 其余土层之间差异均显著(P < 0.05);0—10 cm土层, SX林POC含量最高(10.3 g/kg), YS林(9.5 g/kg)次之, CH林最低(6.8 g/kg), 而10—60 cm土层SX林和CH林均高于YS林。三种人工林土壤MAOC含量均随土层深度的增加而降低, 其中CH林和YS林0—20 cm土层与20—60 cm土层之间差异显著, SX林0—10 cm土层与20—60 cm土层之间差异显著(P < 0.05);0—60 cm土层SX林、CH林MAOC的含量高于YS林, 尤其是0—10 cm土层, CH林和SX林MAOC的含量(6.7 g/kg和6.5 g/kg)是YS林(4.9 g/kg)的1.4倍和1.3倍, 差异显著(P < 0.05)(图 2)。

|
| 图 2 不同人工林型0—60 cm土层POC和MAOC含量变化特征 Fig. 2 Contents of POC and MAOC in 0—60 cm soil layers under different artificial forest types 不同大写字母表示同一土层不同人工林型POC和MAOC含量的差异显著性, 不同小写字母表示同一林型不同土层POC和MAOC含量的差异显著性(P < 0.05)。图中数据为平均值±标准差(n=3) |
三种人工林在0—60 cm土层POC中的FNC含量均随土层深度的增加而降低, 其中SX林不同土层之间差异均显著(P < 0.05), CH林除20—40cm土层与40—60 cm土层外, 其余土层之间差异均显著(P < 0.05), YS林除20—40 cm土层外, 其余土层之间差异均显著(P < 0.05);0—10 cm土层, SX林POC中FNC含量(10.3 g/kg)为CH林(8.3 g/kg)和YS林(7.0 g/kg)的1.2倍和1.5倍, 差异显著(P < 0.05), 10—60 cm土层, CH林和SX林显著高于YS林(P < 0.05)。0—60 cm土层, 三种人工林MAOC中的FNC含量均随土层深度的增加而降低, 其中CH林0—20 cm土层与20—60 cm土层之间差异显著, SX林0—10 cm土层与20—60 cm土层之间差异显著, YS林除20—40 cm与40—60 cm土层外, 其余土层之间差异均显著(P < 0.05);CH林和SX林MAOC中的FNC含量均高于YS林, 尤其是CH林(2.0 g/kg—4.7 g/kg), 与YS林之间(0.7 g/kg—2.4 g/kg)的差异达到显著水平(P < 0.05)(图 3)。

|
| 图 3 不同人工林型0—60 cm土层POC和MAOC中FNC含量的变化特征 Fig. 3 Contents of FNC in POC and MAOC at 0—60 cm soil layers under different artificial forest types 不同大写字母表示同一土层不同人工林型土壤POC和MAOC中FNC含量的差异显著性, 不同小写字母表示同一林型不同土层POC和MAOC中FNC含量的差异显著性(P < 0.05); 图中数据为平均值±标准差(n=3) |
CH林在0—60 cm土层FNC/POC随土层深度的增加而增加, 而YS林则降低, 两种林型0—10 cm土层与40—60 cm土层差异均显著(P < 0.05), SX林先降低后增加;0—20 cm土层, SX林和YS林FNC/POC(34.8%—59.1%和25.5%—57.4%)高于CH林(22.5%—24.0%), 而20—40 cm土层SX林和CH林(25.3%—26.7%和26.1%—28.0%)高于YS林(14.5%—21.1%)。SX林0—10 cm土层FNC/MAOC和20—40 cm土层之间差异显著(P < 0.05), YS林0—10 cm土层与10—40 cm土层之间差异显著(P < 0.05);0—60 cm土层, CH林、SX林土壤FNC/MAOC(52.1%—60.2%和26.3%—56.1%)均高于YS林(20.0%—32.7%), 尤其是0—10 cm土层, 达到显著水平(P < 0.05)(图 4)。

|
| 图 4 不同人工林型0—60 cm土层FNC占POC和MAOC含量比例的变化特征 Fig. 4 Ratios of FNC to POC and MAOC at 0—60 cm soil layers under different artificial forest types 不同大写字母表示同一土层不同人工林型土壤FNC占POC和MAOC含量比例的差异显著性, 不同小写字母表示同一林型不同土层FNC占POC和MAOC含量比例的差异显著性(P < 0.05); 图中数据为平均值±标准差(n=3) |
在0—60 cm土层随土层深度的增加, 三种人工林POC中的BNC含量均显著降低(P < 0.05), 且CH林、SX林POC中BNC含量(1.0 g/kg—3.2 g/kg和0.8 g/kg—3.8 g/kg)显著高于YS林(0.2 g/kg—1.9 g/kg)(P < 0.05)。0—60 cm土层三种人工林MAOC中BNC含量均随土层深度的增加而降低, 其中CH林0—20 cm土层显著高于20—60 cm土层, SX林0—10 cm显著高于10—60 cm土层, YS林除20—40 cm土层与40—60 cm土层外, 其余土层之间差异均显著(P < 0.05);0—10 cm土层SX林MAOC中BNC含量(1.5 g/kg)是CH林(1.1 g/kg)的1.4倍, 达到显著水平(P < 0.05);10—60 cm土层CH林和SX林MAOC中BNC的含量(0.5 g/kg—0.9 g/kg和0.5 g/kg—1.0 g/kg)高于YS林(0.2 g/kg—0.5 g/kg)(图 5)。

|
| 图 5 不同人工林型0—60 cm土层POC和MAOC中BNC含量变化特征 Fig. 5 Contents of BNC in POC and MAOC at 0—60 cm soil layers under different artificial forest types 不同大写字母表示同一土层不同人工林型土壤POC和MAOC中BNC含量的差异显著性, 不同小写字母表示同一林型不同土层POC和MAOC中BNC含量的差异显著性(P < 0.05); 图中数据为平均值±标准差(n=3) |
在0—60 cm土层随土层深度的增加, CH林BNC/POC增加, SX林和YS林降低, 且两种林型0—10 cm土层与20—60 cm土层之间差异均显著(P < 0.05);0—10 cm土层, SX林和YS林BNC/POC(21.5%和16.4%)均高于CH林(9.3%), 10—60 cm土层, CH林和SX林土壤BNC/POC(8.4%—8.9%和8.6%—11.1%)高于YS林(2.1%—6.0%)。SX林BNC/MAOC在0—10 cm土层、40—60 cm土层与10—40 cm土层之间差异显著(P < 0.05), YS林除20—40 cm土层外, 其余土层之间差异均显著(P < 0.05);0—10 cm土层SX林和YS林BNC/MAOC(23.7%和15.5%)高于CH林(13.0%), 尤其是SX林, 显著高于CH林(P < 0.05), 10—60 cm土层, CH林和SX林(12.0%—13.5%和9.7%—13.1%)均高于YS林(4.5%—9.5%), 在40—60 cm土层达到显著水平(P < 0.05)(图 6)。

|
| 图 6 不同人工林型0—60 cm土层BNC占POC和MAOC含量比例的变化特征 Fig. 6 Ratioss of BNC to POC and MAOC at 0—60 cm soil layers under different artificial forest types 不同大写字母表示同一土层不同人工林型土壤BNC占POC和MAOC含量比例的差异显著性, 不同小写字母表示同一林型不同土层BNC占POC和MAOC含量比例的差异显著性(P < 0.05); 图中数据为平均值±标准差(n=3) |
CH林POC中MNC含量在不同土层之间差异均显著(P < 0.05), SX林和YS林除20—40 cm土层外, 其余土层之间差异均显著(P < 0.05);0—60 cm土层, CH林和SX林POC中MNC平均含量(7.5 g/kg和7.9 g/kg)是YS林(4.0 g/kg)的1.9倍和2.0倍, 达到显著水平(P < 0.05)。CH林MAOC中MNC含量在0—20 cm土层和20—40 cm土层之间差异显著(P < 0.05), SX林0—10 cm土层与20—60 cm土层之间差异显著(P < 0.05), YS林除20—40 cm土层, 其余土层之间差异均显著(P < 0.05);0—60 cm土层CH林和SX林MAOC中MNC平均含量(4.1 g/kg和3.2 g/kg)高于YS林(2.0 g/kg), 尤其是CH林, 达到显著水平(P < 0.05)(图 7)。

|
| 图 7 不同人工林型0—60 cm土层POC和MAOC中MNC含量变化特征 Fig. 7 Conetents of MNC in POC and MAOC at 0—60 cm soil layers under different artificial forest types 不同大写字母表示同一土层不同人工林型POC和MAOC中MNC含量的差异显著性, 不同小写字母表示同一林型不同土层POC和MAOC中MNC含量的差异显著性(P < 0.05); 图中数据为平均值±标准差(n=3) |
0—60 cm土层随土层深度的增加, CH林土壤MNC/POC从31.1%增加到36.1%, 且0—10 cm土层与40—60 cm土层之间差异显著(P < 0.05), 而SX林和YS林土壤MNC/POC分别从80.6%和73.8%降低至36.8%和16.8%, 且SX林和YS林0—10 cm土层与40—60 cm土层之间差异均显著(P < 0.05);在0—10 cm土层中, SX林和YS林土壤MNC/POC高于CH林, 而10—60 cm土层SX林和CH林土壤MNC/POC高于YS林。SX林0—10 cm土层MNC/MAOC与20—40 cm土层之间差异显著(P < 0.05), YS林除10—20 cm土层与20—40 cm土层, 其余土层之间差异均显著(P < 0.05);不同土层CH林、SX林土壤MNC/MAOC(64.0%—73.7%和36.0%—79.8%)高于YS林(24.5%—48.2%)(图 8)。

|
| 图 8 不同人工林型0—60 cm土层MNC占POC和MAOC比例 Fig. 8 Ratios of MNC to POC and MAOC at 0—60 cm soil layers under different artificial forest types 不同大写字母表示同一土层不同人工林型MNC占POC和MAOC含量比例的差异显著性, 不同小写字母表示同一林型不同土层MNC占POC和MAOC含量比例的差异显著性(P < 0.05); 图中数据为平均值±标准差(n=3) |
整体上, 0—60 cm土层CH林和SX林土壤BNC/MAOC、FNC/MAOC、MNC/MAOC的平均值高于POC, 且FNC/MAOC的平均比例(56.2%和37.3%)高于BNC(12.8%和14.8%)(表 3)。YS林在0—10 cm土层FNC/POC、MNC/POC(57.4%和73.8%)高于MAOC(32.7%和48.2%)(图 4和8), 因此0—60 cm土层YS林FNC/POC、MNC/POC的平均值高于MAOC(表 3), 而在10—60 cm土层BNC/MAOC、FNC/MAOC、MNC/MAOC的平均值(7.0%, 23.9%, 30.9%)高于POC(4.1%, 20.4%, 24.4%)(图 4, 6和8)。
| 所占比例Ratio | CH/% | SX/% | YS/% | 所占比例Ratio | CH/% | SX/% | YS/% | |
| FNC/POC | 23.9 | 36.9 | 29.6 | BNC/MAOC | 12.8 | 14.8 | 9.2 | |
| BNC/POC | 8.8 | 12.5 | 7.1 | MNC/POC | 33.5 | 49.5 | 36.7 | |
| FNC/MAOC | 56.2 | 37.3 | 26.1 | MNC/MAOC | 69.0 | 52.0 | 35.2 | |
| FNC: 真菌残体碳Fungal necromass carbon;BNC: 细菌残体碳Bacterial necromass carbon;MNC: 微生物残体碳Microbial necromass carbon;POC: 颗粒态有机碳Particulate organic carbon;MAOC: 矿物结合态有机碳Mineral-associated organic carbon;表中所占比例的平均值是4个土层所占比例的平均值(n=4) | ||||||||
相关分析表明, 三种人工林土壤FNC/POC与总有机碳和全氮之间为显著正相关关系(P < 0.05), FNC/MAOC与全氮含量呈现显著正相关关系, 与pH值、砂粒含量呈现显著负相关关系(P < 0.05), BNC/POC与总有机碳和全氮含量之间为显著正相关关系, 与pH值之间为显著负相关关系(P < 0.05), BNC/MAOC与总有机碳和全氮含量之间为显著正相关关系, 与pH值、容重呈现显著负相关关系(P < 0.05), MNC/POC与总有机碳和全氮含量之间呈现显著正相关关系(P < 0.05), MNC/MAOC与黏粒、总有机碳和全氮含量之间为显著正相关关系, 与pH值、砂粒含量呈显著负相关关系(P < 0.05)(图 9)。

|
| 图 9 FNC、BNC、MNC占POC和MAOC含量比例与土壤理化性质之间的相关关系 Fig. 9 Correlations between ratios of FNC, BNC, MNC to POC, MAOC and soil physical and chemical properties (P < 0.05) |
MNC含量占POC和MAOC含量的比例可反映MNC对POC和MAOC的积累贡献[27], 所占比例越大, 积累贡献越大, 反之则积累贡献越小。本研究结果表明, 刺槐林和山杏林0—60 cm土层FNC、BNC、MNC主要贡献MAOC的积累, 积累贡献分别56.2%, 12.8%, 69.0%和37.3%, 14.8%, 52.0%, 其中FNC对MAOC的积累贡献是BNC的4.4倍和2.5倍(表 3), 而油松林0—10 cm土层FNC、MNC主要贡献POC的积累, 积累贡献为57.4%和73.8%, 10—60 cm土层BNC、FNC、MNC主要贡献MAOC的积累, 积累贡献分别为7.0%, 23.9%, 30.9%(图 4, 6和8), 说明与BNC相比, 三种人工林FNC对MAOC的积累起主要作用, 这与现有的研究结果相似[27—28], 可能是由于在MAOC的积累过程中, 微生物形成途径可能是更有效的一种形成途径, 即MNC的续埋效应更有利于MAOC的积累[29—30], 而MNC的续埋主要来源于FNC的积累, 鉴于真菌细胞壁的主要成分是几丁质等, 降解速率较慢, 又容易与黏土矿物颗粒结合, 导致FNC稳定性较高, 有利于稳定性有机碳(MAOC)的积累, 且以氨基葡萄糖为主的FNC易与土壤中的矿物结合, 可形成较为稳定的有机大分子化合物, 进而贡献MAOC的积累[3], 因此, 刺槐林和山杏林FNC主要贡献MAOC的积累;但真菌主要以分解复杂有机物为主[3, 31], 与刺槐林和山杏林相比, 油松林凋落物由于木质素、纤维素含量较高而更难分解[32], 因此油松林凋落物更易被真菌分解、利用, 使表层土壤中的真菌生物量高于细菌生物量, 当养分受限时, 真菌残体可能被POC中的细菌分解而贡献POC的积累[33], 因此, 油松林0—10 cm土层FNC主要贡献POC的积累;而下层微生物群落以细菌为主, 细菌细胞壁主要是胞壁酸, 由肽聚糖组成, 容易被降解, 导致细菌残体可作为真菌的碳源, 被真菌利用且以真菌残体的形式积累[34], 且本研究中BNC/POC、BNC/MAOC与FNC/POC、FNC/MAOC显著正相关(P < 0.05), 因此, 10—60 cm土层FNC主要贡献MAOC的积累。
3.2 影响微生物残体碳贡献土壤有机碳组分积累的主要因素土壤理化性质作为环境因子, 不仅能通过影响微生物的群落结构、多样性及碳利用效率, 进而影响MNC的形成与积累, 还能作用于POC和MAOC的形成过程, 对POC、MAOC含量产生影响, 导致MNC对POC和MAOC的积累贡献存在差异, 如在表层和深层土壤中, 由于土壤理化性质及微生物丰度不同, 微生物源碳对MAOC的贡献比例也不同[29]。本研究中总有机碳、全氮含量与FNC/POC、FNC/MAOC、BNC/POC、BNC/MAOC、MNC/POC、MNC/MAOC均呈现显著正相关关系(P < 0.05), 这与先前的研究结果相似[28], 表明总有机碳和全氮含量能显著影响MNC对土壤有机碳组分的积累贡献, 鉴于MNC主要通过微生物生物标志物-氨基糖表征, 且氨基糖的积累与微生物对碳源和氮源的利用紧密相关, 而土壤中的有机碳、氮为微生物提供了必要的碳源和氮源, 因此有利于氨基糖在土壤中积累, 促进MNC在土壤有机碳组分中的积累[3, 35]。
土壤pH值可通过影响微生物的群落结构及多样性, 进而影响MNC中FNC和BNC的分配比例, 使MNC对POC和MAOC积累的贡献发生变化, 较低的pH值可促进真菌和细菌残体的积累[26]。本研究中pH值与FNC/MAOC、BNC/POC、BNC/MAOC、MNC/MAOC呈显著负相关关系(P < 0.05), 这与现有结果一致[26—27], 表明土壤pH值的增加不利于MNC在土壤有机碳组分中的积累, 尤其是不利于在稳定性碳组分(MAOC)中的积累, 这可能是由于在中性或碱性土壤中微生物生物量的周转速率变慢, 微生物碳利用效率可能降低, 使得微生物残体的产量降低, 导致MNC的积累受到影响[36—38]。
黏粒作为土壤质地中的重要组成部分, 对土壤有机碳及其组分的形成具有重要作用[3], 本研究中黏粒含量与FNC/MAOC、MNC/MAOC、MAOC含量呈显著正相关关系(P < 0.05), 这与现有的研究结果[39]相似, 表明黏粒对MNC、FNC在POC、MAOC中积累具有促进作用, 同时黏粒含量的增加有助于MAOC含量的增加, 这是由于黏粒中次生矿物质含量较高, 是微生物残体积累的主要贮存库, 有利于吸附更多的有机物质, 如微生物残体碳, 提高微生物残体碳的稳定性, 进而形成MAOC[40]。砂粒含量与FNC/MAOC、MNC/MAOC、MAOC含量呈现显著负相关关系(P < 0.05), 表明砂粒含量的增加不利于MNC在MAOC中的积累, 与已有的研究结果[39]相似, 这是因为砂粒中原生矿物含量较高, 而原生矿物粒径较大, 黏着性和吸附性较差, 无法与微生物残体较好地结合, 因此, 不利于MNC在MAOC中积累[41]。土壤容重作为土壤物理性质的重要指标之一, 可反映土壤结构及其透气性的变化, 本研究中土壤容重与BNC/MAOC、POC和MAOC含量之间为显著负相关关系(P < 0.05), 这可能是由于土壤容重越低, 土壤的透气、透水性越好, 有利于微生物生物量的周转和微生物碳利用效率的提高[42], 促进微生物残体的积累, 潜在地影响MNC在土壤有机碳组分中的积累, 尤其是在MAOC中的积累。
4 结论除油松林0—10 cm土层微生物残体碳更有利于颗粒态有机碳的积累外, 整体上, 黄土高原不同人工林在0—60 cm土层微生物残体碳更有利于矿物结合态有机碳的积累, 即随植被的恢复, 黄土高原不同人工林微生物残体碳的积累可促进土壤有机碳稳定性组分的增加, 与细菌残体碳相比, 真菌残体碳对土壤颗粒态和矿物结合态有机碳积累的贡献更大, 土壤总有机碳、全氮、黏粒、砂粒含量、容重及pH值是该区影响微生物残体碳贡献土壤有机碳组分积累的主要因素, 研究结果有助于深入了解黄土高原植被恢复中土壤有机碳的固持机制。
| [1] |
梁超, 朱雪峰. 土壤微生物碳泵储碳机制概论. 中国科学: 地球科学, 2021, 51(5): 680-695. |
| [2] |
Basile-Doelsch I, Balesdent J, Pellerin S. Reviews and syntheses: The mechanisms underlying carbon storage in soil. Biogeosciences, 2020, 17(21): 5223-5242. DOI:10.5194/bg-17-5223-2020 |
| [3] |
张彬, 陈奇, 丁雪丽, 何红波, 张旭东. 微生物残体在土壤中的积累转化过程与稳定机理研究进展. 土壤学报, 2022, 59(6): 1479-1491. |
| [4] |
Schimel J P, Schaeffer S M. Microbial control over carbon cycling in soil. Frontiers in Microbiology, 2012, 3: 348. |
| [5] |
Schweigert M, Herrmann S, Miltner A, Fester T, Kästner M. Fate of ectomycorrhizal fungal biomass in a soil bioreactor system and its contribution to soil organic matter formation. Soil Biology and Biochemistry, 2015, 88: 120-127. DOI:10.1016/j.soilbio.2015.05.012 |
| [6] |
Kindler R, Miltner A, Richnow H H, Kästner M. Fate of gram-negative bacterial biomass in soil-mineralization and contribution to SOM. Soil Biology and Biochemistry, 2006, 38(9): 2860-2870. DOI:10.1016/j.soilbio.2006.04.047 |
| [7] |
Cambardella C A, Elliott E T. Particulate soil organic-matter changes across a grassland cultivation sequence. Soil Science Society of America Journal, 1992, 56(3): 777-783. DOI:10.2136/sssaj1992.03615995005600030017x |
| [8] |
Christensen B T. Physical fractionation of soil and structural and functional complexity in organic matter turnover. European Journal of Soil Science, 2001, 52(3): 345-353. DOI:10.1046/j.1365-2389.2001.00417.x |
| [9] |
Joergensen R G, Wichern F. Alive and kicking: Why dormant soil microorganisms matter. Soil Biology and Biochemistry, 2018, 116: 419-430. DOI:10.1016/j.soilbio.2017.10.022 |
| [10] |
Liang C, Amelung W, Lehmann J, Kästner M. Quantitative assessment of microbial necromass contribution to soil organic matter. Global Change Biology, 2019, 25(11): 3578-3590. DOI:10.1111/gcb.14781 |
| [11] |
Six J, Guggenberger G, Paustian K, Haumaier L, Elliott E T, Zech W. Sources and composition of soil organic matter fractions between and within soil aggregates. European Journal of Soil Science, 2001, 52(4): 607-618. DOI:10.1046/j.1365-2389.2001.00406.x |
| [12] |
Lavallee J M, Soong J L, Cotrufo M F. Conceptualizing soil organic matter into particulate and mineral-associated forms to address global change in the 21st century. Global Change Biology, 2020, 26(1): 261-273. DOI:10.1111/gcb.14859 |
| [13] |
Liang C, Schimel J P, Jastrow J D. The importance of anabolism in microbial control over soil carbon storage. Nature Microbiology, 2017, 2: 17105. DOI:10.1038/nmicrobiol.2017.105 |
| [14] |
Cotrufo M F, Ranalli M G, Haddix M L, Six J, Lugato E. Soil carbon storage informed by particulate and mineral-associated organic matter. Nature Geoscience, 2019, 12: 989-994. DOI:10.1038/s41561-019-0484-6 |
| [15] |
Cotrufo M F, Soong J L, Horton A J, Campbell E E, Haddix M L, Wall D H, Parton W J. Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nature Geoscience, 2015, 8: 776-779. DOI:10.1038/ngeo2520 |
| [16] |
Craig M E, Geyer K M, Beidler K V, Brzostek E R, Frey S D, Stuart Grandy A, Liang C, Phillips R P. Fast-decaying plant litter enhances soil carbon in temperate forests but not through microbial physiological traits. Nature Communications, 2022, 13: 1229. DOI:10.1038/s41467-022-28715-9 |
| [17] |
Xu H W, Qu Q, Lu B B, Zhang Y, Liu G B, Xue S. Variation in soil organic carbon stability and driving factors after vegetation restoration in different vegetation zones on the Loess Plateau, China. Soil and Tillage Research, 2020, 204: 104727. DOI:10.1016/j.still.2020.104727 |
| [18] |
杨阳, 窦艳星, 王宝荣, 薛志婧, 朱兆龙, 黄懿梅, 王云强, 安韶山. 黄土高原土壤有机碳固存机制研究进展. 第四纪研究, 2023, 43(2): 509-522. |
| [19] |
杨阳, 张萍萍, 吴凡, 周媛媛, 宋怡, 王云强, 安韶山. 黄土高原植被建设及其对碳中和的意义与对策. 生态学报, 2023, 43(21): 9071-9081. |
| [20] |
Jin Z, Dong Y S, Wang Y Q, Wei X R, Wang Y F, Cui B L, Zhou W J. Natural vegetation restoration is more beneficial to soil surface organic and inorganic carbon sequestration than tree plantation on the Loess Plateau of China. Science of The Total Environment, 2014, 485/486: 615-623. DOI:10.1016/j.scitotenv.2014.03.105 |
| [21] |
鲍士旦. 土壤农化分析 (3版). 北京: 中国农业出版社, 2000: 22-97.
|
| [22] |
Six J, Elliott E T, Paustian K, Doran J W. Aggregation and soil organic matter accumulation in cultivated and native grassland soils. Soil Science Society of America Journal, 1998, 62(5): 1367-1377. DOI:10.2136/sssaj1998.03615995006200050032x |
| [23] |
杨娥女, 王宝荣, 姚宏佳, 黄懿梅, 安韶山. 黄土高原生物土壤结皮发育过程中颗粒态和矿物结合态有机碳变化特征. 水土保持研究, 2023, 30(1): 25-33, 40. |
| [24] |
Zhang X D, Amelung W. Gas chromatographic determination of muramic acid, glucosamine, mannosamine, and galactosamine in soils. Soil Biology and Biochemistry, 1996, 28(9): 1201-1206. DOI:10.1016/0038-0717(96)00117-4 |
| [25] |
Joergensen R G. Amino sugars as specific indices for fungal and bacterial residues in soil. Biology and Fertility of Soils, 2018, 54(5): 559-568. DOI:10.1007/s00374-018-1288-3 |
| [26] |
Wang B R, An S S, Liang C, Liu Y, Kuzyakov Y. Microbial necromass as the source of soil organic carbon in global ecosystems. Soil Biology and Biochemistry, 2021, 162: 108422. DOI:10.1016/j.soilbio.2021.108422 |
| [27] |
Hu P L, Zhang W, Chen H S, Xu L, Xiao J, Luo Y Q, Wang K L. Lithologic control of microbial-derived carbon in forest soils. Soil Biology and Biochemistry, 2022, 167: 108600. DOI:10.1016/j.soilbio.2022.108600 |
| [28] |
王杉杉, 徐秋芳, 范博, 郑旭理, 王中乾, 梁辰飞, 陈俊辉, 秦华, 王懿祥, 李甄, 王利芝, 邵帅. 毛竹扩张对杉木林土壤微生物残体碳积累的影响. 生态学报, 2023, 43(5): 1902-1912. |
| [29] |
Sokol N W, Sanderman J, Bradford M A. Pathways of mineral-associated soil organic matter formation: Integrating the role of plant carbon source, chemistry, and point of entry. Global Change Biology, 2019, 25(1): 12-24. DOI:10.1111/gcb.14482 |
| [30] |
Sokol N W, Slessarev E, Marschmann G L, Nicolas A, Blazewicz S J, Brodie E L, Firestone M K, Foley M M, Hestrin R, Hungate B A, Koch B J, Benjamin J, Stone B W, Sullivan M B, Zablocki O, Pett-Ridge J. Life and death in the soil microbiome: how ecological processes influence biogeochemistry. Nature reviews microbiology, 2022, 20(7): 415-430. DOI:10.1038/s41579-022-00695-z |
| [31] |
López-Mondéjar R, Brabcová V, Štursová M, Davidová A, Jansa J, Cajthaml T, Baldrian P. Decomposer food web in a deciduous forest shows high share of generalist microorganisms and importance of microbial biomass recycling. ISME Journal, 2018, 12: 1768-1778. DOI:10.1038/s41396-018-0084-2 |
| [32] |
于颖超, 张心昱, 戴晓琴, 吕斯丹, 杨洋, 史丽娟. 亚热带红壤区森林土壤剖面微生物残体碳分布及影响因素. 生态学报, 2022, 42(3): 1108-1117. |
| [33] |
Kennedy P G, Maillard F. Knowns and unknowns of the soil fungal necrobiome. Trends in Microbiology, 2023, 31(2): 173-180. DOI:10.1016/j.tim.2022.08.011 |
| [34] |
Zheng T T, Miltner A, Liang C, Nowak K M, Kästner M. Turnover of bacterial biomass to soil organic matter via fungal biomass and its metabolic implications. Soil Biology and Biochemistry, 2023, 180: 108995. DOI:10.1016/j.soilbio.2023.108995 |
| [35] |
Sun T, Mao X L, Han K F, Wang X J, Cheng Q, Liu X, Zhou J J, Ma Q X, Ni Z H, Wu L H. Nitrogen addition increased soil particulate organic carbon via plant carbon input whereas reduced mineral-associated organic carbon through attenuating mineral protection in agroecosystem. Science of The Total Environment, 2023, 899: 165705. DOI:10.1016/j.scitotenv.2023.165705 |
| [36] |
Malik A A, Puissant J, Buckeridge K M, Goodall T, Jehmlich N, Chowdhury S, Gweon H S, Peyton J M, Mason K E, van Agtmaal M, Blaud A, Clark I M, Whitaker J, Pywell R F, Ostle N, Gleixner G, Griffiths R I, 2018. Land use driven change in soil pH affects microbial carbon cycling processes. Nature Communications, 2018, 9: 3591.
|
| [37] |
Tao F, Huang Y Y, Hungate B A, Hungate B A, Manzoni S, Frey S D, Schmidt M W I, Reichstein M, Carvalhais N, Ciais P, Jiang L F, Lehmann J, Wang Y P, Houlton B Z, Ahrens B, Mishra U, Hugelius G, Hocking T D, Lu X J, Shi Z, Viatkin K, Vargas R, Yigini Y, Omuto C, Malik A A, Peralta G, Cuevas-Corona R, Di Paolo L E, Luotto I, Liao C J, Liang Y S, Saynes V S, Huang X M, Luo Y Q. Microbial carbon use efficiency promotes global soil carbon storage. Nature, 2023, 618(7967): 981-985. DOI:10.1038/s41586-023-06042-3 |
| [38] |
Wang C, Qu L R, Yang L M, Liu D W, Morrissey E, Miao R H, Liu Z P, Wang Q K, Fang Y T, Bai E. Large-scale importance of microbial carbon use efficiency and necromass to soil organic carbon. Global Change Biology, 2021, 27(10): 2039-2048. DOI:10.1111/gcb.15550 |
| [39] |
Wang B R, Huang Y M, Li N, Yao H J, Yang E N, Soromotin A V, Kuzyakov Y, Cheptsov V, Yang Y, An S S. Initial soil formation by biocrusts: Nitrogen demand and clay protection control microbial necromass accrual and recycling. Soil Biology and Biochemistry, 2022, 167: 108607. DOI:10.1016/j.soilbio.2022.108607 |
| [40] |
Aumtong S, de Neergaard A, Magid J. Formation and remobilisation of soil microbial residue. Effect of clay content and repeated additions of cellulose and sucrose. Biology and Fertility of Soils, 2011, 47(8): 863-874. DOI:10.1007/s00374-011-0592-y |
| [41] |
Kubiak-Ossowska K, Tokarczyk K, Jachimska B, Mulheran P A. Bovine serum albumin adsorption at a silica surface explored by simulation and experiment. Journal of Physical Chemistry B, 2017, 121(16): 3975-3986. DOI:10.1021/acs.jpcb.7b01637 |
| [42] |
Spohn M, Klaus K, Wanek W, Richter A. Microbial carbon use efficiency and biomass turnover times depending on soil depth-Implications for carbon cycling. Soil Biology and Biochemistry, 2016, 96: 74-81. DOI:10.1016/j.soilbio.2016.01.016 |