 2024, Vol. 44
2024, Vol. 44文章信息
- 徐嘉艺, 王小玲, 宋坤, 张桂莲, 仲启铖, 韩玉洁, 达良俊
- XU Jiayi, WANG Xiaoling, SONG Kun, ZHANG Guilian, ZHONG Qicheng, HAN Yujie, DA Liangjun
- 上海四种常见树种单株固碳能力差异及其影响因子
- Individual carbon sequestration capacity of four common tree species and their impact factors in Shanghai
- 生态学报. 2024, 44(13): 5532-5541
- Acta Ecologica Sinica. 2024, 44(13): 5532-5541
- http://dx.doi.org/10.20103/j.stxb.202309262094
-
文章历史
- 收稿日期: 2023-09-26
- 网络出版日期: 2024-04-25





2. 崇明生态研究院, 上海 202162;
3. 自然资源部大都市区国土空间生态修复工程技术创新中心, 上海 200062;
4. 上海市园林科学规划研究院, 城市困难立地生态园林国家林业和草原局重点实验室, 上海 200232;
5. 上海市林业总站, 上海 200072;
6. 上海城市森林生态系统国家定位观测研究站, 上海 200240;
7. 西安建筑科技大学交叉创新研究院, 干旱半干旱区生态科学与工程研究院, 西安 710055
2. Institute of Eco-Chongming, Shanghai 202162, China;
3. Technology Innovation Center for Land Spatial Eco-restoration in Metropolitan Area, Ministry of Natural Resources, Shanghai 200062, China;
4. Shanghai Academy of Landscape Architecture Science and Planning, Key Laboratory of National Forestry and Grassland Administration on Ecological Landscaping of Challenging Urban Sites, Shanghai 200232, China;
5. Shanghai Forest Station, Shanghai 200072, China;
6. Shanghai Urban Forest Ecosystem Research Station, Shanghai 200240, China;
7. Institute of Science and Engineering of Ecology in Arid and Semi-arid Areas, Institute for Interdisciplinary and Innovate Research, Xi'an University of Architecture and Technology, Xi'an 710055, China
随着全球经济的快速发展和城市化进程的加快, 人口增长、能源过度使用、土地不合理利用和大规模的森林砍伐等引发的气候变化问题被视为全球主要的环境挑战之一, 也是全球发展中最重要的问题之一[1]。政府间气候变化专门委员会(IPCC)在其第六次评估中明确指出, 人类活动产生的温室气体排放是导致全球变暖的主要原因。近年来, 为应对此类气候问题, 各国政府均提出碳达峰及碳中和的目标[2]。城市森林的固碳能力作为生态系统调节服务中的重要一环, 对实现碳达峰及碳中和的目标起到了重要作用, 因而受到了越来越多的关注。城市树木通过光合作用固定碳并将多余的碳储存为生物质来充当二氧化碳(CO2)的汇, 可以作为抵消CO2排放的有效策略, 在维持和改善城市生态环境中发挥着核心作用[3]。因此, 探究城市树木的固碳能力对城市可持续发展具有重要意义。
目前, 对城市树木固碳能力的研究主要从碳储量、收支或碳库动态变化等方面展开, 对不同城市的林地或绿色空间进行了固碳量的估算[4—6]。对树木固碳量的估算最常见的是基于生物量的方法, 即通过直接或间接调查获取林分数据(如树种、树木胸径、树木高度等), 建立树种生物量异速生长方程, 以此估算树木生物量, 再将此估算值与树种的碳质量分数相乘得到树木碳储量[4—8], 进而计算碳储量年增量并将其作为树木年固碳量[9—10]。也有研究通过测量树木的光合作用效率或是利用遥感的方法推算森林的固碳量[11—13]。但这些研究多在区域尺度上估算城市森林固碳量, 对于在树种尺度以及单木尺度固碳能力的研究, 目前报道较少。然而, 不同树种的固碳能力及其对环境的响应可能存在显著差异[14—15]。Wang等[13]对北京多个城市绿地的植物群落固碳能力进行调查, 发现树木的固碳能力会随着其个体大小发生变化, 较大的树木通常表现出较高的固碳能力。然而, 这种正向的变化趋势是非线性的, 随着树木个体大小的增加, 其固碳能力的提高速率将逐渐下降, 若树木进入到成熟和衰老期, 其固碳能力甚至呈现下降趋势[16]。此外, 树木的固碳能力也受到植株密度的制约, 它可以通过个体间的竞争影响树木的生长和碳积累[16]。在热带半干旱区的森林草地系统中的研究也表明, 不同的植株密度对树木单株固碳量和单位面积固碳量都具有显著影响[17]。
在城市中, 影响树木生长的环境因子在空间上高度异质化。例如, 中心城区与城郊地区相比, 表现出更高的温度、CO2浓度以及气溶胶浓度等[18]。Chen等[19]对沈阳市油松生长情况的研究表明, 树木的径向生长受局地气候和区域气候的显著影响, 尤其是春冬季的温度及降水情况;David等[20]的研究表明, 巴黎行道树的生长速率对春秋季降水情况和气候变化非常敏感;Dale等[21]的研究表明, 水的可用性在树木生长过程中起到主要作用, 同时也是树木对城市压力适应程度的主要决定因素。另外, 过多的夜间照明会产生光污染[22], 使树木在夜间被迫进行光合作用, 导致其生长节律发生改变。因而, 树木在城市异质环境中可能会呈现出不同的固碳能力。
近年来上海以生态之城建设为指引, 城市森林的面积快速增加, 截至2022年底上海市森林覆盖率达到18.51%, 是城市碳汇的主要贡献者。目前, 对上海城市森林碳汇的研究主要集中在评估城市森林或绿地等的碳储量、群落动态和碳库等的大尺度变化上[23—25], 或是针对土壤有机碳进行研究[26]。虽然王丽勉[27]等通过测定光合生理指标, 评估了上海多种绿化树种的固碳能力, 但尚未有研究通过长期监测来评估上海常见树种的固碳能力。此外, 上海的热环境、降水和夜间灯光等影响树木生长的环境因子空间异质性较高[28—29], 其对树木生长的综合影响尚不清楚。因而, 亟需探明上海主要树种的固碳能力及其影响因子。
本研究利用上海城市森林固定样地多年的监测数据, 以上海市常见的四种绿化树种为研究对象, 通过估算年固碳量来表征树木固碳能力, 进而比较不同树种和径组之间固碳能力的差异, 探究不同树种固碳能力与个体大小、植株密度、温度、降水量和夜间灯光强度的关系, 以期能为上海异质城市环境中绿地碳汇功能提升提供科学的树种选择依据。
1 研究方法 1.1 研究区概况上海市位于中国东部沿海地区中部(30°40′N—31°53′N, 120°51′E—122°12′E), 属于北亚热带季风气候类型, 降水充足, 光照充沛, 年均气温15.8 ℃, 年均降水量1200 mm[30]。上海作为我国特大型城市之一和长三角大都市圈的中心, 其现代化和城市化程度高。上海境内自然植被稀少, 大部分为人工植被。人工林种中常绿阔叶树种以香樟(Cinnamomum camphora)、女贞(Ligustrum lucidum)等为主, 落叶针叶树种以水杉(Metasequoia glyptostroboides)等为主, 落叶阔叶树种以银杏(Ginkgo biloba)、复羽叶栾(Koelreuteria bipinnata)等为主, 常绿针叶树种以雪松(Cedrus deodara)等为主。
1.2 数据来源 1.2.1 固定样地数据依据不同生活型, 选取香樟、水杉、银杏和雪松四种上海常见树种作为研究对象, 分别代表常绿阔叶树种、落叶针叶树种、落叶阔叶树种和常绿针叶树种。四种树种的生长动态数据提取于174个长期固定观测样地。样地大小介于20 m × 20 m—50 m × 50 m之间, 主要为人工中幼龄纯林, 涵盖了上海市的大部分行政区(图 1)。固定样地观测最早始于2004年, 对样地内的所有胸径大于5 cm的木本植物进行挂牌, 测量胸径等参数。本研究使用2004年至2022年间的重复测量数据, 95%的重复测量数据其间隔为2—5 a, 5%的重复测量数据其间隔为6—12 a。其中, 样本量(重复测量次数)最多的是香樟, 为3838条;水杉次之, 为1517条;银杏为568条;雪松样本数最少, 为126条。本研究以树木胸径表征个体大小, 以相邻两次测量数据中的前一次数据为基准, 计算植株密度(株/hm2), 作为相邻两次测量间隔期树木固碳能力的影响因子之一。

|
| 图 1 样地位置分布情况 Fig. 1 Location of sample sites |
(1) 温度及降水
从国家青藏高原数据中心(http://data.tpdc.ac.cn/)下载中国1 km空间分辨率的逐月平均气温以及逐月平均降水量数据集, 数据单位分别为0.1 ℃和0.1 mm。该数据集是根据CRU以及WorldClim发布的全球高分辨率气候数据集, 通过Delta空间降尺度方案在中国地区降尺度生成的, 并且经过地面496个独立气象观测点数据验证。利用此数据集计算各样地在每两次测量间隔期的年平均温度(单位:℃)和年平均降水量(单位:mm)。
(2) 夜间灯光强度
夜间灯光数据参考Wu等[31]发表的城市夜间灯光数据集。该数据集是通过整合DMSP-OLS和SNPP-VIIRS夜间灯光遥感数据得到的矫正后的逐年中国范围的类DMSP-OLS数据, 数据空间分辨率为1 km。利用此数据集计算各样地在每两次测量间隔期的年平均夜间灯光强度(无量纲单位)。
1.3 数据处理与统计分析 1.3.1 单株树木固碳能力估算根据已发表文献中上海地区不同树种的生物量异速生长方程(表 1), 计算单株树木的生物量, 再根据相应树种的碳质量分数换算成碳储量(Carbon Storage, CS)。利用相邻两次测量数据, 计算单株树木的年固碳量, 以此表征其在相邻两次测量间隔期的固碳能力(Carbon Sequestration Capacity, CSC), 计算公式为:

|
(1) |
| 生活型 Life form |
树种 Tree species |
生物量异速生长方程 Allometric equation for biomass |
碳质量分数 Mass fraction of carbon |
|
| 方程Equation | R2 | |||
| 常绿阔叶树种Evergreen broadleaved tree | 香樟Cinnamomum camphora | BM=0.10387D2.535 | 0.99 | 0.4433 |
| 落叶针叶树种Deciduous conifer | 水杉Metasequoia glyptostroboides | BM=0.06291D2.4841 | 0.97 | 0.4340 |
| 落叶阔叶树种Deciduous broadleaved tree | 银杏Ginkgo biloba | BM=0.133137D2.3357 | 0.98 | 0.4437 |
| 常绿针叶树种Evergreen conifer | 雪松Cedrus deodara | BM=0.1606D2.2134 | 0.99 | 0.4542 |
| BM为全株生物量/kg, D为胸径/cm, 引自文献[32—33] | ||||
式中, CSt表示前一次调查得到的单株树木碳储量, CSt+1表示后一次调查得到的单株树木碳储量, Δt表示相邻两次调查的年份之差。
1.3.2 统计分析由于树木的固碳能力随着个体的增大而发生变化, 因此本研究根据《森林资源连续清查技术规程》(GB/T 38590—2020)中的径组及径级划分标准, 依据胸径将树木划分成不同的径组(表 2)。分树种和径组采用单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)分析, 检验同一径组不同树种和同一树种不同径组间的固碳能力是否存在显著差异。
| 径组 DBH group |
划分标准(包含的径级) Classification standard (DBH class included) |
径组 DBH group |
划分标准(包含的径级) Classification standard (DBH class included) |
|
| (Ⅰ)小Small | 平均胸径小于13.0 cm(6, 8, 10, 12 cm) | (Ⅳ)大Large | 平均胸径29.0—37.0 cm(30, 32, 34, 36 cm) | |
| (Ⅱ)中Medium | 平均胸径13.0—21.0 cm(14, 16, 18, 20 cm) | (Ⅴ)特大Extra large | 平均胸径≥37.0 cm(38 cm以上) | |
| (Ⅲ)较大Slightly larger | 平均胸径21.0—29.0 cm(22, 24, 26, 28 cm) |
为探究每一树种单株固碳能力与影响因子的关系, 本研究以多种影响因子作为自变量(个体大小、植株密度、温度、降水量和夜间灯光强度), 以树种单株固碳能力作为因变量建立多元线性回归模型。首先, 对于不符合正态分布的参数(单株固碳能力和个体大小), 采用对数转换法将数据转换为正态分布。设y为因变量, x1, x2, …, xk为自变量, 并且自变量与因变量之间为线性关系时, 多元线性回归模型为:
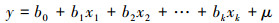
|
(2) |
式中, b0为常数项, b1, b2, …, bk为回归系数, μ为随机误差。
其次根据AIC值采用双向剔除法选择最优解释模型。最后, 采用“平均排序(Averaging over orderings)”(也称LMG指数)的方法, 评估模型中解释变量的相对贡献[34];该分析应用R程序包“relaimpo”实现。所有统计分析均在R Version 4.1.2中执行。
2 结果与分析 2.1 不同树种单株固碳能力及其差异本研究中, 香樟、水杉、银杏和雪松的单株固碳能力分别介于4.01—51.58 kg/a、1.04—13.47 kg/a、0.62—18.56 kg/a和1.48—8.47 kg/a之间(表 3)。在各个径组中, 香樟的单株固碳能力均显著高于其他3个树种(P < 0.05), 而水杉、银杏和雪松的单株固碳能力在不同径组中略有差异。在小径组中, 雪松的单株固碳能力显著高于银杏;在中径组中, 雪松的单株固碳能力显著高于水杉;在较大径组中, 水杉和雪松的单株固碳能力显著高于银杏;在大径组中水杉的单株固碳能力显著高于银杏。
| 树种 Tree species |
样本量/条 Number of samples |
固碳能力/(kg/a) Carbon sequestration capacity |
径组 DBH group |
| 香樟Cinnamomum camphora | 1533 | 4.01±0.14Da | 小 |
| 水杉Metasequoia glyptostroboides | 672 | 1.04±0.10Cbc | 小 |
| 银杏Ginkgo biloba | 486 | 0.62±0.03Cc | 小 |
| 雪松Cedrus deodara | 35 | 1.48±0.31Cb | 小 |
| 香樟Cinnamomum camphora | 1691 | 10.67±0.19Ca | 中 |
| 水杉Metasequoia glyptostroboides | 602 | 3.91±0.24Bc | 中 |
| 银杏Ginkgo biloba | 17 | 5.15±1.35Bbc | 中 |
| 雪松Cedrus deodara | 75 | 4.95±0.36Bb | 中 |
| 香樟Cinnamomum camphora | 548 | 18.49±0.55Ba | 较大 |
| 水杉Metasequoia glyptostroboides | 204 | 7.46±0.43Bb | 较大 |
| 银杏Ginkgo biloba | 16 | 6.49±3.11Bc | 较大 |
| 雪松Cedrus deodara | 16 | 8.47±0.86Ab | 较大 |
| 香樟Cinnamomum camphora | 52 | 34.15±2.69Aa | 大 |
| 水杉Metasequoia glyptostroboides | 36 | 13.47±1.32ABb | 大 |
| 银杏Ginkgo biloba | 33 | 7.06±1.15Bc | 大 |
| 香樟Cinnamomum camphora | 14 | 51.58±8.82Aa | 特大 |
| 水杉Metasequoia glyptostroboides | 3 | 12.40±7.20Ab | 特大 |
| 银杏Ginkgo biloba | 16 | 18.56±3.59Ab | 特大 |
| 均值±标准误;不同大写字母表示径组间差异的显著性, 不同小写字母表示树种间差异的显著性(P<0.05) | |||
总体而言, 四种树种的单株固碳能力随径级的增加呈现增高的趋势(图 2)。然而, 随着径级的增加, 四种树种的单株固碳能力差异也随之增大。胸径超过30 cm的个体, 其单株固碳能力波动范围较大, 尤其是样本数量较多的香樟(图 2), 如在径级40 cm处, 香樟的单株固碳能力达到最大为107.88 kg/a;在径级44 cm处, 香樟的单株固碳能力介于30—100 kg/a;在径级50 cm处, 香樟的单株固碳能力介于19—70 kg/a。

|
| 图 2 上海常见四种树种单株固碳能力随径级增加的变化格局 Fig. 2 Variation of the individual carbon sequestration capacity with increasing DBH class for four common tree species in Shanghai |
个体大小对四种树种的单株固碳能力均有显著正影响, 而植株密度和环境因子对四种树种单株固碳能力的影响不同(表 4)。植株密度对香樟和水杉的单株固碳能力有显著负影响, 而对银杏的单株固碳能力为显著正影响, 对雪松的单株固碳能力无显著影响。在环境因子中, 温度与香樟、水杉和银杏单株固碳能力呈现显著负相关, 而与雪松单株固碳能力无显著相关性。降水量与香樟单株固碳能力呈显著负相关, 与水杉单株固碳能力呈显著正相关, 而与雪松和银杏的单株固碳能力无显著相关性。夜间灯光强度与香樟和水杉的单株固碳能力呈现显著正相关, 而与雪松和银杏的固碳能力无显著相关性。上述影响因子对各树种单株固碳能力变异的总解释率, 即模型R2, 为0.559—0.607。
| 树种 Tree species |
样本量/株 Number of samples |
自变量 Independent variables |
系数 Coefficients |
标准误差 Standard error |
t | Pr(>|t|) | 调整R2 Adjusted R2 |
| 香樟 | 3883 | 个体大小 | 2.110 | 0.045 | 47.242 | <0.001 | 0.607 |
| Cinnamomum camphora | 植株密度 | -0.002 | 0.000 | -17.218 | <0.001 | ||
| 温度 | -0.714 | 0.048 | -14.735 | <0.001 | |||
| 降水量 | -0.004 | 0.000 | -14.014 | <0.001 | |||
| 夜间灯光强度 | 0.008 | 0.001 | 6.441 | <0.001 | |||
| 水杉 | 1517 | 个体大小 | 2.214 | 0.084 | 26.322 | <0.001 | 0.579 |
| Metasequoia | 植株密度 | -0.000 | 0.000 | -9.501 | <0.001 | ||
| glyptostroboides | 温度 | -0.567 | 0.134 | -4.232 | <0.001 | ||
| 降水量 | 0.001 | 0.000 | 2.837 | 0.004 | |||
| 夜间灯光强度 | 0.014 | 0.005 | 2.841 | 0.004 | |||
| 银杏 | 568 | 个体大小 | 2.028 | 0.096 | 21.041 | <0.001 | 0.569 |
| Ginkgo biloba | 植株密度 | 0.001 | 0.000 | 5.617 | <0.001 | ||
| 温度 | -1.488 | 0.147 | -10.144 | <0.001 | |||
| 雪松 | 126 | 个体大小 | 2.501 | 0.237 | 10.549 | <0.001 | 0.559 |
| Cedrus deodara | 温度 | -0.721 | 0.419 | -1.721 | 0.088 | ||
| 夜间灯光强度 | -0.018 | 0.011 | -1.699 | 0.092 |
各影响因子对单株树木固碳能力变异的解释相对贡献如图 3所示。在四个树种中, 个体大小所占贡献均最大, 介于61.4%—78.4%之间。对于香樟和水杉, 植株密度的贡献仅次于个体大小, 分别为31.1%和23.9%。温度、降水量和夜间灯光强度对香樟的贡献分别为3.6%、2.6%和1.4%;温度、降水量和夜间灯光强度对水杉的贡献分别为5.6%、0.6%和5.9%。对于银杏, 贡献排第二的是温度, 为18.0%;植株密度的贡献为13.9%。对于雪松, 贡献排第二的是夜间灯光强度, 为15.4%;温度的贡献为6.1%。

|
| 图 3 各影响因子对上海四种常见树种单株固碳能力变异的相对贡献 Fig. 3 The relative importance of impact factors on variations of individual carbon sequestration capacity of four common tree species in Shanghai ** 表示P < 0.01, *** 表示P < 0.001 |
本研究利用长期固定观测数据, 估算了上海四种常见树种的单株年固碳量, 以表征其固碳能力, 反映了单株个体在现生长体征下的固碳速率[35]。结果表明, 香樟在各径组中均具有最高的固碳能力(4.01—51.58 kg/a), 这与兰延等[36]的研究一致。他们通过对中国不同区域香樟的单位面积日固碳量和释氧量进行分析得出, 香樟是各地区中具有较高固碳释氧能力的绿化树种。香樟作为常绿树种, 在上海乃至亚热带地区可全年生长, 且其光合速率和叶面积指数较大[36], 因此具有更高的年固碳量。在相同径组中, 水杉、银杏和雪松的单株固碳能力较低且基本无显著差异, 这与马陆航[35]的研究相似。他们将城市居住区绿地的乔木树种固碳能力进行聚类后得出, 香樟为高固碳能力树种, 水杉、银杏和雪松为中低固碳能力树种。此外, 水杉、银杏和雪松均为裸子植物, 以往的研究表明, 被子植物占主导地位的林分群落其生物量积累速率高于裸子植物占主导地位的林分群落[37]。基于解剖学数据的理论建模和实验测量结果表明, 裸子植物表现出比被子植物更慢的韧皮部运输速度[38], 而韧皮部中碳水化合物从叶到根的运输速度是影响树木碳封存动力的重要因素[39]。
3.2 个体大小和植株密度对单株树木固碳能力的影响本研究中, 四种树种的单株固碳能力均随个体大小(胸径级)的增加而增加, 是众多解释因子中最为重要的固碳能力预测因子。以往研究表明, 个体大小对木本植物固碳能力起主导作用, 较大的树木通常表现出较高的固碳能力[11]。然而, 在达到一定径级后, 树木的固碳能力会出现随径级的增加而下降的趋势, 这与自然衰老导致的生理功能下降等有关[40]。因而, 人工林不能长期保持较高的固碳水平, 随着林分从幼龄林生长至成熟林, 林分的固碳量显著增加, 但随着年龄和胸径的增长, 固碳量逐渐趋于稳定甚至下降[16]。当前上海绿林地中的树木主要为中小径级个体[32], 因而上海绿林地的固碳能力仍有很大的提升潜力。
植株密度对香樟和水杉单株固碳能力的影响较大, 植株密度越大、单珠固碳能力越小;对银杏的影响较小, 植株密度越大、单株固碳能力越大, 而对雪松的单株固碳能力无显著影响(表 4, 图 3)。植株密度过高能引起生长资源的强制分配, 进而导致种内竞争强度增加, 影响种群中单株植物的生长量和生物量[41]。在上海, 香樟和水杉的生长相对较快, 植株密度偏高、林冠郁闭度高, 树木个体对光照、土壤养分以及水肥等资源的争夺加剧[42], 抑制了单株个体的碳积累。对于银杏而言, 由于研究样地主要为中幼龄林, 个体偏小且植株密度低, 导致林冠郁闭度低, 个体间竞争的影响尚未体现;此外, 植株密度过低时, 易滋生杂草, 影响了银杏对光照和养分资源的获取, 对生长产生负面影响[43]。因此, 在城市森林营造过程中应正确规划并合理控制植株密度, 以发挥树木的最大固碳效益。
3.3 城市环境因子对单株树木固碳能力的影响温度是影响树木固碳能力的重要环境因子, 它对树木固碳能力的影响既可以表现为正影响[44], 也可以表现为负影响[45]。本研究中温度与香樟、水杉和银杏的单株固碳能力均呈现显著的负相关关系。在我国东北林区的研究表明, 在控制降水量的影响后, 树木固碳量与年均温度之间存在显著的负相关关系[46]。在热带森林中的研究表明, 树木的年生长变异性是由温度和降水的变化共同决定的, 随着气温的上升, 温度与生长之间存在负相关关系[47]。这种温度与树木生长之间的负相关机制可能是由于更高的温度导致碳同化率降低, 限制光合作用速率和增加呼吸速率, 植物组织的维护成本随着环境温度的升高而增加, 从而导致整体较低的碳吸收[48]。此外, 在缺水干旱的条件下, 温度升高会增加蒸散需求, 引发干旱胁迫, 导致树木生长减缓[49]。
降水量对树木固碳能力的影响取决于树种的生态习性[50]。本研究中降水量与香樟的单株固碳能力呈显著负相关, 与水杉的单株固碳能力呈显著正相关, 而对银杏和雪松的单株固碳能力无影响。不同树种固碳能力对降水的响应差异可能受根和树干的解剖学特征驱动[50]。香樟属于深根性树种, 主根发达, 其根系蔓延能力相对较差, 不耐积水, 对水分供应反应敏感。有研究表明, 雨季增雨能显著降低香樟细根的根长和比根长, 同时降低粗根的比根长[51]。而水杉个体常分布于河流沿岸, 表现出对潮湿环境的偏好性, 在一定程度上能耐受淹水环境。
夜间灯光强度也是影响植物固碳能力的关键因素[52]。本研究中夜间灯光强度对香樟和水杉的单株固碳能力均有显著正向影响。随着夜间灯光强度的增加, 香樟和水杉的单株固碳能力增加, 这可能与两种树种拥有更低的光补偿点有关, 它们能够利用夜间的低光强度进行碳的吸收和固定[53]。这与李小婷等[54]的研究一致, 即夜间灯光的补充照明促进了香樟的生长, 在一定程度上增加了香樟的生物量。
本研究中影响因子对树木单株固碳能力的作用还需要更多的研究加以验证。首先, 本研究中仅涉及四个树种, 其他树种的响应尚不清晰;树种的生物学和生态学习性如何作用于它们对各影响因子的响应也需进一步探索。其次, 处于不同生态区的城市树木是否具有相似的“树种—环境响应关系”也需要更多的城市研究加以检验。此外, 本研究大多是基于纯林的研究结果, 只考虑了同种邻体密度的影响, 对于混交种植的城市森林, 还需要进一步探究异种邻体密度的影响。
4 结论本研究探究了上海城市森林四种常见树种的单株固碳能力及其影响因子, 发现香樟、水杉、银杏和雪松的单株固碳能力分别介于4.01—51.58 kg/a、1.04—13.47 kg/a、0.62—18.56 kg/a和1.48—8.47 kg/a之间, 且在个体大小相近的植株中, 香樟相较于水杉、银杏和雪松具有更高的单株固碳能力。个体大小对单株树木固碳能力有显著且重要的正向影响, 而植株密度的影响因树种而异。城市环境因子中, 温度对单株树木固碳能力有显著负向影响, 而降水量、夜间灯光强度的影响因树种而异。在未来城市森林建设和管理中, 应根据不同树种单株固碳能力的环境敏感性, 适地植树造林, 进而提升城市森林的固碳效益。
| [1] |
Xu C, Wang B J, Chen J D. Forest carbon sink in China: linked drivers and long short-term memory network-based prediction. Journal of Cleaner Production, 2022, 359: 132085. DOI:10.1016/j.jclepro.2022.132085 |
| [2] |
Wei Y M, Chen K Y, Kang J N, Chen W M, Wang X Y, Zhang X Y. Policy and management of carbon peaking and carbon neutrality: a literature review. Engineering, 2022, 14: 52-63. DOI:10.1016/j.eng.2021.12.018 |
| [3] |
周健, 肖荣波, 庄长伟, 邓一荣. 城市森林碳汇及其抵消能源碳排放效果——以广州为例. 生态学报, 2013, 33(18): 5865-5873. |
| [4] |
de Castro Neto M, Sarmento P. Assessing Lisbon trees' carbon storage quantity, density, and value using open data and allometric equations. Information, 2019, 10(4): 133. DOI:10.3390/info10040133 |
| [5] |
Zhao J F, Ai J L, Zhu Y J, Huang R X, Peng H W, Xie H F. Carbon budget of different forests in China estimated by an individual-based model and remote sensing. PLoS One, 2023, 18(10): e0285790. DOI:10.1371/journal.pone.0285790 |
| [6] |
Gratani L, Varone L, Bonito A. Carbon sequestration of four urban parks in Rome. Urban Forestry & Urban Greening, 2016, 19: 184-193. |
| [7] |
Timilsina N, Staudhammer C L, Escobedo F J, Lawrence A. Tree biomass, wood waste yield, and carbon storage changes in an urban forest. Landscape and Urban Planning, 2014, 127: 18-27. DOI:10.1016/j.landurbplan.2014.04.003 |
| [8] |
Davies Z G, Edmondson J L, Heinemeyer A, Leake J R, Gaston K J. Mapping an urban ecosystem service: quantifying above-ground carbon storage at a city-wide scale. Journal of Applied Ecology, 2011, 48(5): 1125-1134. DOI:10.1111/j.1365-2664.2011.02021.x |
| [9] |
耿燕, 赵秀海, 安黎哲. 阔叶红松林乔木地上碳储量和碳增量对采伐强度的响应. 北京林业大学学报, 2022, 44(10): 23-32. |
| [10] |
庄红蕾, Xavier Becuwe, 肖春波, 王月华, 王海, 殷杉, 刘春江. 上海崇明岛水杉人工林生物量方程构建及固碳潜力研究. 上海交通大学学报: 农业科学版, 2012, 30(2): 48-55. |
| [11] |
Wang Y N, Chang Q, Li X Y. Promoting sustainable carbon sequestration of plants in urban greenspace by planting design: a case study in parks of Beijing. Urban Forestry & Urban Greening, 2021, 64: 127291. |
| [12] |
陆君, 刘亚风, 齐珂, 樊正球. 福州市森林碳储量定量估算及其对土地利用变化的响应. 生态学报, 2016, 36(17): 5411-5420. |
| [13] |
张桂莲. 基于遥感估算的上海城市森林碳储量空间分布特征. 生态环境学报, 2021, 30(9): 1777-1786. |
| [14] |
Behera S K, Mishra S, Sahu N Y, Manika N, Singh S N, Anto S, Kumar R, Husain R, Verma A K, Pandey N. Assessment of carbon sequestration potential of tropical tree species for urban forestry in India. Ecological Engineering, 2022, 181: 106692. DOI:10.1016/j.ecoleng.2022.106692 |
| [15] |
Schütt A, Becker J N, Reisdorff C, Eschenbach A. Growth response of nine tree species to water supply in planting soils representative for urban street tree sites. Forests, 2022, 13(6): 936. DOI:10.3390/f13060936 |
| [16] |
Xie Y Y, Li J T, Liu Q M, Gong J, Wulan T Y, Zhou M, Zheng Y, Shen Z H. Determinants of growth and carbon accumulation of common plantation tree species in the three northern regions, China: responses to climate and management strategies. The Science of the Total Environment, 2023, 900: 165831. DOI:10.1016/j.scitotenv.2023.165831 |
| [17] |
Gupta D K, Bhatt R K, Keerthika A, Noor Mohamed M B, Shukla A K, Jangid B L. Carbon sequestration potential of Hardwickia Binata roxb. based agroforestry in hot semi-arid environment of India: an assessment of tree density impact. Current Science, 2019, 116(1): 112. DOI:10.18520/cs/v116/i1/112-116 |
| [18] |
耿树丰, 任嘉义, 杨俊, 国安东, 席建超. 局地气候区视角下的城市热环境研究. 生态学报, 2022, 42(6): 2221-2227. |
| [19] |
Chen Z J, He X Y, Cui M X, Davi N, Zhang X L, Chen W, Sun Y. The effect of anthropogenic activities on the reduction of urban tree sensitivity to climatic change: dendrochronological evidence from Chinese pine in Shenyang city. Trees, 2011, 25(3): 393-405. DOI:10.1007/s00468-010-0514-x |
| [20] |
David A A J, Boura A, Lata J C, Rankovic A, Kraepiel Y, Charlot C, Barot S, Abbadie L, Ngao J. Street trees in Paris are sensitive to spring and autumn precipitation and recent climate changes. Urban Ecosystems, 2018, 21(1): 133-145. DOI:10.1007/s11252-017-0704-z |
| [21] |
Dale Adam G, Frank Steven D. Water availability determines tree growth and physiological response to biotic and abiotic stress in a temperate North American urban forest. Forests, 2022, 13(7): 1012. DOI:10.3390/f13071012 |
| [22] |
Saraiji R, Oommen M S. Light pollution index (LPI): an integrated approach to study light pollution with street lighting and façade lighting. LEUKOS, 2012, 9(2): 127-145. DOI:10.1582/LEUKOS.2012.09.02.004 |
| [23] |
张彪, 谢紫霞, 高吉喜. 上海城市森林植被固碳功能及其抵消能源碳排放效果评估. 生态学报, 2021, 41(22): 8906-8920. |
| [24] |
凌焕然, 王伟, 樊正球, 王祥荣. 近二十年来上海不同城市空间尺度绿地的生态效益. 生态学报, 2011, 31(19): 5607-5615. |
| [25] |
Shen G R, Wang Z J, Liu C J, Han Y J. Mapping aboveground biomass and carbon in Shanghai's urban forest using Landsat ETM+ and inventory data. Urban Forestry & Urban Greening, 2020, 51: 126655. |
| [26] |
张青青, 伍海兵, 梁晶. 上海市绿地表层土壤有机碳储量的估算. 土壤, 2020, 52(4): 819-824. |
| [27] |
王丽勉, 胡永红, 秦俊, 高凯, 黄娟. 上海地区151种绿化植物固碳释氧能力的研究. 华中农业大学学报, 2007, 26(3): 399-401. |
| [28] |
史军, 崔林丽, 杨涵洧, 孙兰东. 上海气候空间格局和时间变化研究. 地球信息科学学报, 2015, 17(11): 1348-1354. |
| [29] |
戴晓燕, 张利权, 过仲阳, 吴健平, 栗小东, 朱燕玲. 上海城市热岛效应形成机制及空间格局. 生态学报, 2009, 29(7): 3995-4004. |
| [30] |
宋坤, 郭雪艳, 王泽英, 黄莎莎, 严佳瑜, 叶建华, 乐莺, 严明, 吴梅, 达良俊. 上海城市近自然森林的重建动态. 生态学杂志, 2020, 39(4): 1075-1081. |
| [31] |
Wu Y Z, Shi K F, Chen Z Q, Liu S R, Chang Z J. Developing improved time-series DMSP-OLS-like data (1992-2019) in China by integrating DMSP-OLS and SNPP-VⅡRS. IEEE Transactions on Geoscience and Remote Sensing, 2021, 60: 4407714. |
| [32] |
郭雪艳. 上海城市森林多尺度生态质量评价研究[D]. 上海: 华东师范大学, 2017.
|
| [33] |
王哲, 韩玉洁, 康宏樟, 黄丹, 薛春燕, 殷杉, 刘春江. 黄浦江上游主要树种水源涵养林生态系统碳储量. 生态学杂志, 2012, 31(8): 1930-1935. |
| [34] |
刘尧, 于馨, 于洋, 胡文浩, 赖江山. R程序包"rdacca.hp" 在生态学数据分析中的应用: 案例与进展. 植物生态学报, 2023, 47(1): 134-144. |
| [35] |
马陆航. 居住区绿地固碳植物配置模式及设计优化研究——以西安市长庆坊为例[D]. 西安: 西安建筑科技大学, 2023.
|
| [36] |
兰延, 刘顺. 香樟在城市园林绿化中的应用及其生态效益评价. 林业科技通讯, 2019(5): 73-77. |
| [37] |
Anderson K J, Allen A P, Gillooly J F, Brown J H. Temperature-dependence of biomass accumulation rates during secondary succession. Ecology Letters, 2006, 9(6): 673-682. DOI:10.1111/j.1461-0248.2006.00914.x |
| [38] |
Liesche J, Windt C, Bohr T, Schulz A, Jensen K H. Slower phloem transport in gymnosperm trees can be attributed to higher sieve element resistance. Tree Physiology, 2015, 35(4): 376-386. DOI:10.1093/treephys/tpv020 |
| [39] |
Mildner M, Bader M K F, Leuzinger S, Siegwolf R T W, Körner C. Long-term 13C labeling provides evidence for temporal and spatial carbon allocation patterns in mature Picea abies. Oecologia, 2014, 175(3): 747-762. DOI:10.1007/s00442-014-2935-5 |
| [40] |
Sun S J, He C X, Qiu L F, Li C Y, Zhang J S, Meng P. Stable isotope analysis reveals prolonged drought stress in poplar plantation mortality of the Three-North Shelter Forest in Northern China. Agricultural and Forest Meteorology, 2018, 252: 39-48. DOI:10.1016/j.agrformet.2017.12.264 |
| [41] |
李博. 生态学. 北京: 高等教育出版社, 2000.
|
| [42] |
代林利, 周丽丽, 伍丽华, 刘丽, 黄樱, 彭婷婷, 邱静雯, 何宗明, 曹光球. 不同林分密度杉木林生态系统碳密度及其垂直空间分配特征. 生态学报, 2022, 42(2): 710-719. |
| [43] |
Farooq T H, Yan W D, Chen X Y, Shakoor A, Rashid M H U, Gilani M M, He Z M, Wu P F. Dynamics of canopy development of Cunninghamia lanceolata mid-age plantation in relation to foliar nitrogen and soil quality influenced by stand density. Global Ecology and Conservation, 2020, 24: e01209. DOI:10.1016/j.gecco.2020.e01209 |
| [44] |
Li C Y, Chen B, Zhang X D, Wang X P, Zha T S, Wang X W. Prediction of the carbon pool for lacebark pine under future temperature changes. The Forestry Chronicle, 2015, 91(5): 519-523. DOI:10.5558/tfc2015-090 |
| [45] |
Babst F, Bouriaud O, Poulter B, Trouet V, Girardin M P, Frank D C. Twentieth century redistribution in climatic drivers of global tree growth. Science Advances, 2019, 5(1): eaat4313. DOI:10.1126/sciadv.aat4313 |
| [46] |
Yan G Y, Wang Q G, Han S J, Guo Z L, Yu J H, Wang W J, Fan C N, Cao W, Wang L H, Xing Y J, Zhang Z. Beneficial effects of warming on temperate tree carbon storage depend on precipitation and mycorrhizal types. The Science of the Total Environment, 2022, 819: 153086. DOI:10.1016/j.scitotenv.2022.153086 |
| [47] |
Vlam M, Baker P J, Bunyavejchewin S, Zuidema P A. Temperature and rainfall strongly drive temporal growth variation in Asian tropical forest trees. Oecologia, 2014, 174(4): 1449-1461. DOI:10.1007/s00442-013-2846-x |
| [48] |
Awal M A, Ohta T, Matsumoto K, Toba T, Daikoku K, Hattori S, Hiyama T, Park H. Comparing the carbon sequestration capacity of temperate deciduous forests between urban and rural landscapes in central Japan. Urban Forestry & Urban Greening, 2010, 9(3): 261-270. |
| [49] |
Williams A P, Cook E R, Smerdon J E, Cook B I, Abatzoglou J T, Bolles K, Baek S H, Badger A M, Livneh B. Large contribution from anthropogenic warming to an emerging North American megadrought. Science, 2020, 368(6488): 314-318. DOI:10.1126/science.aaz9600 |
| [50] |
Rodriguez-Robles U, Arredondo T, Smart D. Wood anatomical and physiological differences between semiarid pine and oak, stand up for their tree ring sensibility to precipitation variability. Agricultural and Forest Meteorology, 2023, 338: 109530. DOI:10.1016/j.agrformet.2023.109530 |
| [51] |
何至杭, 刘悦, 陶玉柱, 吴妙兰, 高畅, 王艺颖, 于耀泓, 周庆, 莫其锋. 降雨季节分配改变对热带次生林不同功能型幼苗根系形态特征的影响. 生态学杂志, 2024, 43(1): 96-105. |
| [52] |
Bennie J, Davies T W, Cruse D, Gaston K J. Ecological effects of artificial light at night on wild plants. Journal of Ecology, 2016, 104(3): 611-620. DOI:10.1111/1365-2745.12551 |
| [53] |
Falster D S, Duursma R A, FitzJohn R G. How functional traits influence plant growth and shade tolerance across the life cycle. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(29): E6789-E6798. |
| [54] |
李小婷, 陈斌, 王海军, 郑光, 杨丹, 苗馨予, 徐驰. 城市夜间灯光对香樟生长的影响. 应用生态学报, 2019, 30(7): 2284-2290. |