 2024, Vol. 44
2024, Vol. 44文章信息
- 佀海翔, 王瑞燕, 王秋红
- SI Haixiang, WANG Ruiyan, WANG Qiuhong
- 黄河流域植被光合动态时空演变及归因分析——基于日光诱导叶绿素荧光数据
- Temporal and attribution analysis of vegetation photosynthesis dynamics in the Yellow River Basin based on satellite-derived SIF
- 生态学报. 2024, 44(13): 5695-5705
- Acta Ecologica Sinica. 2024, 44(13): 5695-5705
- http://dx.doi.org/10.20103/j.stxb.202309212050
-
文章历史
- 收稿日期: 2023-09-21
- 网络出版日期: 2024-04-25





2. 肥城市现代农业发展服务中心, 泰安 271600
2. Feicheng City Modern Agriculture Development Service Center, Tai'an 271600, China
根据联合国政府间气候变化专门委员会(IPCC)第六次报告, 碳排放会导致气候变暖并可能对生态系统产生不可逆的影响[1]。植被作为陆地生态系统的重要组成部分之一, 联结土壤、大气、水文等生态要素, 在碳平衡调节、水土保持以及稳定气候和生态系统方面具有重要作用[2—3]。植被光合作用作为植被的核心生理过程, 通过将大气中的CO2转化为有机碳并储存于植物中, 不仅维持了生态系统的平衡, 还发挥了重要的碳汇功能, 有助于减缓气候变暖的进程[4—5], 在生态系统中具有突出的地位。
黄河流域作为中国第二大流域, 是中国北方重要的生态安全屏障, 也是中国经济发展以及人类活动非常重要的地区, 在中国经济社会发展和生态安全方面具有十分重要的地位[6]。我国政府在2019年明确提出了黄河流域生态保护和高质量发展的重大国家战略。然而, 近几十年来流域内人类活动不断增加, 导致黄河流域面临着生态环境退化、水土流失严重、碳减排压力大等严重问题[7]。植被作为陆地区域生态系统维持者和碳汇主体, 其光合时空分布的信息获取对于维护和恢复区域生态环境、应对气候变化、确保区域生态安全以及实现黄河流域的高质量发展具有重要意义。
当前, 许多学者已经基于归一化植被指数(normalized difference vegetation index, NDVI)、增强植被指数(enhanced vegetation index, EVI)等植被指数对黄河流域的植被变化以及其推动因素进行了广泛的研究, 并取得了丰富的研究成果[8—11]。然而传统的植被指数仅能反映出植被潜在的光合作用能力, 并不与光合作用过程直接相关[12—13], 在进行植被光合动态研究中可能会存在一些局限性, 因此迫切需要一种新的指标来进行相关研究。
日光诱导叶绿素荧光(solar-induced chlorophyll fluorescence, SIF)是植物叶绿素分子在吸收光量子之后, 在激发态的叶绿素分子跃迁回基态的过程中发射的一种光谱信号(650—800nm), 能够准确地反映出植被光合作用能力以及生理状况等信息, 是植被健康状况的“探针”[14—15], 可以对植被与环境之间的物候关系以及植被光合效率进行评估[16]。SIF与植被光合作用紧密耦合[17], 在植被生理状态刚开始发生变化时就有所体现, 比基于反射率的植被指数更加及时。例如Song等人研究发现SIF对热胁迫的响应显著降低且较早, 而NDVI和EVI在一个月后才能捕捉到这些信号[18]。因此SIF作为一种新兴的遥感数据, 可以为植被光合动态研究提供新的数据源和视角。然而, 卫星SIF在监测黄河流域植被光合动态变化方面的能力仍在研究中。SIF在监测黄河流域植被光合中对于各影响因素的响应也并不明确, 各因素对于植被光合动态的贡献尚难以区分。因此, 本研究利用2001—2021年GOSIF数据对黄河流域植被光合的时空变化特征进行分析, 探究过去21年中植被光合动态规律, 并对气候变化和人类活动对植被光合的影响进行深入剖析。以上研究将为推动黄河流域生态保护和高质量发展提供重要的参考依据, 并为大范围生态环境管理和深入研究气候变化提供新的见解和方向。
1 数据与方法 1.1 研究区概况黄河流域(图 1)位于北纬36.16°—41.86°与东经95.88°—119.07°之间, 东西长约1900km, 南北宽约1100 km, 流域面积约为79.45万km2。黄河流域地处中纬度地带, 位于中国干旱和半干旱地区, 属于温带大陆性气候, 同时它还受到季风环流的影响, 气候状况复杂。气温自西向东由冷变暖, 降水分布不均匀。黄河流域地势西高东低, 自西向东地跨青藏高寒、西北干旱半干旱和华北湿润半湿润等若干自然分区, 穿越青藏高原、内蒙古高原、黄土高原和黄淮海平原四个地貌单元, 涉及青海省、四川省、甘肃省、宁夏回族自治区、内蒙古自治区、陕西省、山西省、河南省和山东省9个省区[19]。

|
| 图 1 研究区概况 Fig. 1 Overview of the study area |
GOSIF(https://data.globalecology.unh.edu/data/GOSIF_v2/)是利用离散的OCO-2 SIF探测数据、MODIS遥感数据以及气象再分析数据开发得出的具有高时空分辨率(0.05°, 8d)的叶绿素荧光数据集[20]。本研究使用来自GOSIF的2001—2021年的年尺度数据。
温度、降水以及太阳总辐射数据产品均来自ECMWF的ERA5—Land月尺度再分析数据产品(https://cds.climate.copernicus.eu/), 空间分辨率为0.1°, 时间范围为2001—2021年。
土地利用数据来自MCD12C1年尺度数据产品(https://lpdaac.usgs.gov/products/mcd12c1v061/), 空间分辨率为0.05°。通过对其进行重分类, 最终确定各土地利用类型为林地、灌木、稀树草原、草地、湿地、农田、水体、裸地以及建设用地9种类型。
本研究使用Google Earth Engine(https://code.earthengine.google.com/)云平台进行气象数据的年尺度合成及下载工作。所有数据在进行分析之前都被重采样至0.1°的空间分辨率。
1.3 方法 1.3.1 SIF变化趋势显著性分析利用Sen+Mann-Kendall趋势分析方法来估算2001—2021年黄河流域SIF的变化趋势。在显著水平α=0.05下判断变量变化的显著性, 使用正态统计量Z来表示趋势的强度, 当|Z|>1.96时变化显著, |Z|≤1.96时变化不显著。分类标准如表 1所示。
| 斜率 Slope/(W m-2μm-1sr-1a-1) |
正态统计量Z Normal statistics |
变化趋势 Variation trends |
斜率 Slope/(W m-2μm-1sr-1a-1) |
正态统计量Z Normal statistics |
变化趋势 Variation trends |
|
| ≥0.0005 | >1.96 | 显著退化 | ≤-0.0005 | -1.96—1.96 | 轻微改善 | |
| ≥0.0005 | -1.96—1.96 | 轻微退化 | ≤-0.0005 | < -1.96 | 显著改善 | |
| -0.0005—0.0005 | -1.96—1.96 | 相对稳定 | ||||
| SIF:日光诱导叶绿素荧光 | ||||||
采用偏相关分析方法研究温度、降水和太阳辐射与SIF之间的偏相关程度。在计算过程中控制其中两个气候因子, 只分析一个气候因子与SIF的相关程度。偏相关系数的正负表示温度、降水和太阳辐射与SIF之间的正负相关性[21]。
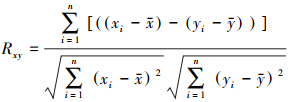
|
(1) |
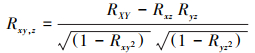
|
(2) |
其中, Rxy为两个变量之间的相关系数;x与y分别表示两个变量n年平均值;n表示样本数。最后采用t检验方法完成偏相关系数的显著性检验。
此外, 利用残差分析来区分由气候变化导致的SIF变化和由人类活动导致的SIF变化[22]。基于预测SIF和实际SIF来计算残差, 同时对残差进行时空变化分析, 当残差可以忽略不计时, 可以用气候要素来解释SIF的变化。当SIF的残差变化很大时, SIF的变化则是由人类活动引起的。SIF残差的上升趋势表明人类活动改善了植被光合活动, 下降趋势表明人类活动导致了植被光合作用的减弱[23]。
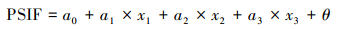
|
(3) |
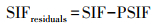
|
(4) |
其中, PSIF为预测SIF, x1、x2和x3分别为各像元的气候数据, a0为回归模型的截距, a1、a2和a3分别表示自变量的回归系数, θ表示随机误差。
2 研究结果 2.1 SIF植被时空变化特征黄河流域SIF总体上呈现出波动上升的趋势(图 2), 2001—2021年SIF平均值由0.04 W m-2μm-1sr-1增长至0.07 W m-2μm-1sr-1, 多年平均值为0.06 W m-2μm-1sr-1, 高峰值出现在2021年。

|
| 图 2 2001—2021年SIF年际变化 Fig. 2 Interannual variation in SIF from 2001 to 2021 SIF:日光诱导叶绿素荧光 |
黄河流域SIF的空间分布格局相对稳定, 总体上呈现出西部和东南部高、北部低的分布格局(图 3)。低SIF区(0.000—0.025)主要分布在黄河流域的西北地区以及青海省的西部地区, 较低SIF区(0.025—0.064)主要分布在甘肃省的东部以及陕西省的北部地区, 而中SIF区(0.064—0.104)主要分布在青海省的东南部、甘肃省的西南部、陕西省和山西省的南部地区、河南省的北部地区以及山东地区。较高SIF区(0.104—0.149)和高SIF区(0.149—0.234)则分布在青海省的南部和黄河中下游地区。这一分布格局可能受到地理、气候和土壤条件等的影响, 反映了不同地区的植被类型和生态系统的差异。

|
| 图 3 2001—2021年黄河流域年平均SIF分布及占比 Fig. 3 Annual average SIF distribution and percentage in the Yellow River Basin from 2001 to 2021 |
黄河流域SIF年动态变化趋势总体以改善为主, 但存在明显的空间异质性(图 4)。显著改善区占黄河流域总面积的75.46%, 主要集中分布在黄河流域的中部和东南部地区, 这种改善趋势反映了生态系统恢复或气候条件的改善。SIF变化趋势不明显的区域主要分布在青海省东南部和黄河流域的西北地区, 这些地区的植被类型主要为草地。1.01%流域面积的SIF呈现出退化趋势, 这些地区主要位于青海省南部、甘肃省与四川省交界地区以及河南省的中部地区, 特别是关中平原、阿尼玛卿山、宁夏平原以及黄河三角洲地区, 说明这些地区正在面临严峻的生态挑战。

|
| 图 4 2001—2021年黄河流域SIF变化趋势及占比 Fig. 4 SIF Change trends and percentage in the Yellow River Basin from 2001 to 2021 |
2001—2021年黄河流域温度呈增加趋势, 降水和太阳辐射呈减少趋势。温度增加速度为0.0361 ℃/a, 降水和太阳辐射的减少速度分别为1.4413 mm/a和0.1028 J m-2a-1, 说明黄河流域存在变暖变干的现象。
黄河流域SIF对温度、降水和太阳辐射的响应如图 5所示。温度与SIF正相关的面积占比为26.42%。这些区域主要分布在陇上高原、关中平原、毛乌素沙地以及黄土高原的北部地区, 其中主要的植被类型为草地(82.09%)和农田(9.47%), 说明温度的增加有利于这些地区的植被生长。温度与SIF负相关的面积占比为73.58%, 显著负相关的面积占0.64%, 主要分布于四川省西北部地区, 其中植被类型主要为草地(80.77%)和农田(19.23%)。

|
| 图 5 2001—2021年黄河流域温度、降水和太阳辐射变化趋势及与SIF的偏相关空间特征 Fig. 5 Trends in Temperature, Precipitation, and Solar Radiation in the Yellow River Basin from 2001 to 2021, along with Spatial Characteristics of Partial Correlation with SIF |
降水与SIF正相关的面积占比为66.13%, 显著正相关的面积占5.27%, 集中分布在陕西省中部以及河南省西部的山区, 这些地区的主要植被类型为农田(44.50%)和林地(29.28%)。降水与SIF负相关的面积占比为39.87%, 显著负相关的面积占2.46%, 集中分布于青海省的南部高海拔地区, 这些地区的主要植被类型为草地(96.00%)。总体上, 降水对黄河流域西南部及东北部为负相关, 其他地区为正相关。
太阳辐射与SIF正相关的面积占比为69.89%, 显著正相关的区域面积占21.65%, 主要分布于黄土高原、关中盆地以及运城盆地等地区, 植被类型主要为草地(36.15%)、农田(33.98%)和林地(14.53%)。太阳辐射与SIF负相关的面积占比为30.11%, 呈显著负相关的面积占比为0.16%, 集中分布于阿尼玛卿的主峰玛卿冈日以及巴颜喀拉山的最高峰年保玉则峰附近, 海拔较高, 植被类型主要为草地(92.31%)。太阳辐射和SIF的偏相关空间分布特征与降水类似。
2.3 植被光合动态变化的归因分析本研究利用温度、降水和太阳辐射这三个气候要素估算了2001—2021年黄河流域预测SIF与SIF残差变化趋势(图 6)。在流域的东部地区, 包括山东省西部、河南省、山西省、陕西省中部以及甘肃省东部, 预测SIF明显增加, 这些区域占流域总面积的25.00%。预测SIF明显下降的区域主要分布在陕西省南部和河南省中部, 占流域总面积的0.17%。此外, SIF残差明显上升的区域占比最大, 为66.29%, 主要分布在黄土高原以及流域的下游区域, 表明在这些地区的人类活动对植被光合动态产生了积极影响。29.74%区域面积的SIF残差基本保持稳定, 主要分布在青海省和内蒙古自治区, 这意味着在这些地区, 人类活动对SIF的影响相对较小。而SIF残差显著下降的区域占0.52%, 主要分布在阿尼玛卿山南部、秦岭以及河南中部地区, 意味着人类活动对SIF产生了负面影响, 导致其减少。

|
| 图 6 黄河流域预测SIF与SIF残差变化趋势及占比 Fig. 6 Predicted SIF and SIF Residual Trends and Percentage in the Yellow River Basin |
本研究进一步根据原始SIF、预测SIF以及SIF残差的Sen斜率, 对黄河流域植被光合动态驱动因素的空间分布进行了分析(图 7), 黄河流域植被光合动态受气候变化和人类活动共同驱动改善的面积最大, 占比为85.14%, 主要集中于甘肃省、陕西省、宁夏回族自治区、山西省、河南省、山东省以及青海省的北部。仅由气候变化驱动改善的区域面积占比为9.98%, 分布于内蒙古自治区的西北部。仅由人类活动驱动改善的区域面积占比为1.14%, 主要分布于青海省的南部、陕西省南部以及黄河三角洲。同时, 流域内受气候变化和人类活动共同驱动退化的区域面积占比为2.35%, 主要分布于内蒙古自治区和宁夏回族自治区的北部、陕西省的关中平原、河南省的中部以及青海省和甘肃省的南部地区。而仅由气候变化驱动退化和仅由人类活动驱动退化的区域面积占比分别为1.08%和0.31%, 这两种类型主要分布于青海省和内蒙古自治区。

|
| 图 7 黄河流域植被光合动态驱动因素空间分布及占比 Fig. 7 Spatial Distribution and Percentage of Vegetation Photosynthetic Dynamic Driving Factors in the Yellow River Basin |
气候变化和人类活动对植被光合动态有不同程度的影响。总体上, 人类活动对植被光合动态的整体贡献率为76.24%, 而气候变化对植被光合动态的贡献率为23.76%, 这表明植被光合动态变化主要受人类活动的影响。
为了更深入地了解黄河流域植被光合动态的驱动机制, 本研究将研究区划分为改善区域和退化区域(图 8)。在改善区域中, 人类活动和气候变化的平均相对贡献率分别为76.67%和23.33%。受气候变化主导的植被光合动态改善区域面积占比约为7.53%, 主要分布在青海省与河南省。受人类活动主导的植被光合动态改善区域面积占比约为92.47%, 主要分布在黄土高原以及鄂尔多斯高原等流域中部地区。在退化区域中, 人类活动和气候变化的平均相对贡献率分别为68.29%和31.71%。受气候变化主导的植被光合动态退化区域面积占比为21.94%, 主要分布在甘肃省西南部、陕西省南部以及山东省。受人类活动主导的植被光合动态退化区域面积占比为78.03%, 主要分布在青海省的东部、内蒙古自治区的北部以及四川省北部地区。

|
| 图 8 改善区域气候变化贡献率、退化区域气候变化贡献率、改善区域人类活动贡献率和退化区域人类活动贡献率 Fig. 8 Contribution Rates of Climate Change in Improved Regions, Degraded Regions, Human Activities in Improved Regions, and Human Activities in Degraded Regions |
不同植被类型在气候变化和人类活动对植被光合的贡献方面也存在差异(图 9)。在植被改善区域中, 人类活动对植被光合的贡献远大于气候变化, 气候变化对林地SIF的贡献在各植被类型中最大(32.95%), 人类活动对草地SIF的贡献最大(79.36%)。在植被退化区域中, 气候变化对稀树草原和农田的贡献大于人类活动, 而林地中的气候变化的贡献与人类活动相当, 人类活动对草地、灌木和湿地的贡献大于气候变化。

|
| 图 9 不同植被类型下气候变化和人类活动相对贡献率 Fig. 9 Relative Contributions of Climate Change and Human Activities to Different Vegetation Types |
在本研究中, 本研究利用趋势分析方法揭示了黄河流域2001—2021年SIF的时空动态特征, 与前人基于NDVI的研究结果进行对照发现[24—26], 两者在黄河流域植被光合作用的整体趋势相同, 整体上均呈现出波动上升的改善趋势, 在黄河流域中下游地区的时空特征也表现出了相似的改善特征。两者退化区均处于关中平原、宁夏平原、黄河三角洲以及黄河源区等地。但在黄河源区, 两者退化区域和退化程度有所差别, SIF存在的零星显著退化的区域集中分布在青海省的南部地区和甘肃省与四川省交界地区的阿尼玛卿山附近的草地。而该区域的NDVI不存在显著退化, 其轻微退化区域主要分布在海南藏族自治州东部以及黄南藏族自治州附近, 植被类型也为草地。这两个区域的SIF与NDVI所展示的光合作用结果的差异可能与地形和土壤背景等有关。黄河源区地形起伏大、景观破碎度高, 干扰了NDVI的对植被光合作用的指示。而黄河流域中下游地区地势较为平坦, 空间景观分布相对均质, 下垫面对NDVI的影响小, 表现出与SIF的一致性。
3.2 SIF对气候要素变化的响应分析SIF对气候要素变化的响应结果发现, 黄河流域内不同地区SIF对气候变化的响应特征并不一致, 这可能与地表形态、气候系统以及植被类型等多种因素的差异有关。就黄河流域温度而言, 在温带大陆性气候区和高原山地气候区的海拔较高地区, 如陇上高原、关中平原、毛乌素沙地以及黄土高原的北部地区, 温度的上升促进了草地和农田的光合作用, 这可能是由于温度是限制这些区域植物生长的主要因素, 适宜的温度上升会提高植被的代谢活动和光合效率[27]。然而, 在四川西北部的高原山地气候区, 温度的轻微增加可能会改变周围环境的变化, 加速土壤水分蒸发和水分亏缺[28], 限制了植物的生长和光合作用, 导致SIF降低。同时, 处于温带季风区的陕西省中部和河南省西部的山区降水及其与植被光合作用关系与其他地区不同, 该区降水与SIF呈显著负相关关系, 即降水显示出轻微减少的趋势, 而SIF不降反增, 这可能与该地区植被的耐旱性有关, 植被可能已经适应了当地的水分条件并具有完善的生理调节机制, 当水分轻微减少时, 植被能通过改善根系吸水结构以适应较干燥的条件, 从而使光合保持稳定甚至呈现改善趋势, 这一发现强调了在季风影响区域中植被类型在应对气象干旱的重要意义。此外, 太阳辐射对草地、农田和林地植被光合作用产生不同的影响, 在阿尼玛卿的主峰玛卿冈日以及巴颜喀拉山的最高峰年保玉则峰附近, 太阳辐射与草地SIF呈现出显著负相关的现象。在黄土高原、关中盆地以及运城盆地等地区, 太阳辐射与农田SIF呈显著正相关。太阳辐射的影响与降水具有相似的空间分布特征, 因此这两者可能相互作用, 共同影响植被的生长。尽管该地区的降水显示出轻微增加的趋势, 但过强的光照可能触发光抑制现象或加剧水分胁迫, 这可能是导致该地区SIF退化的原因。
3.3 气候变化和人类活动对植被光合的相对贡献本研究发现人类活动促进了大多数地区的植被光合作用, 但在一些干旱半干旱地区, 如青海省东部、内蒙古自治区北部以及四川省北部等地, 却产生了负面影响, 这与前人的研究结果一致[9]。此外, 本研究发现人类活动对黄河流域植被光合动态的贡献(76.24%)远大于气候变化(23.76%), 且人类活动多为正向贡献, 这与Dai等人[29]的研究结果一致。
在植被改善区域中, 不同植被类型中人类活动的积极影响均高于气候变化, 说明近些年来, 随着农耕技术的进步和退耕还林还草等广泛实施的林业生态工程显著地促进了流域内植被的改善, 极大的推动了黄河流域生态环境质量建设[30], 这也说明了政府和社会在生态保护方面的积极努力。但是, 在青海省的南部地区、甘肃省与四川省交界地区以及河南省的中部地区, 特别是关中平原、阿尼玛卿山、宁夏平原以及黄河三角洲地区, 本研究观察到了植被光合作用显著退化的现象, 这些地区可能受到了来自气候变化或人类活动的负面影响, 例如交通设施工程的建设、矿产资源的不合理开采以及水资源分配不均等造成植被破坏[31], 需要对这些地区进行重点研究, 并制定合理的政策及措施来恢复自然植被, 维持生态系统的稳定。如在工程施工过程中, 实施严格水土资源保护措施, 确保植被生态系统稳定;通过实施调水调沙等措施, 实现区域内科学的水资源调配和管理, 确保植被所需的水源得以充分供应;持续开展生态恢复工程, 因地制宜, 选择适宜树种草种, 提高植被覆盖率;同时建立完善的生态环境监测系统, 定期监测植被光合活性及相关环境因素, 增加对植被光合退化机制的理解, 以实现植被光合脆弱区的可持续性改善。
此外, 在植被退化区域中, 气候变化对稀树草原和农田的贡献高于人类活动, 林地中的气候变化的贡献与人类活动相当, 但人类活动对草地、灌木和湿地的贡献又高于气候变化, 尤其是人类活动对草地的贡献最大。人类活动对草地类型的负面影响最为显著, 这可能由于过度放牧、城市化扩张等因素导致草地的退化, 从而造成植被光合作用的下降。因此, 在城市发展过程中, 需要加强黄河全流域城镇化的“大协同”, 并建立与生态环境相协调的城镇空间体系。同时需要完善相关政策, 以在更高层面统筹推动黄河流域城镇化, 在不同的区域因地制宜, 实现黄河流域生态保护与高质量发展。
本研究中还存在不足之处, 由于当前CO2数据集的时间不连续等问题, 本研究重点关注了气候变化及人类活动对黄河流域植被光合时空动态的影响, 并未充分考虑到CO2在这一过程中的作用。本研究将在后续的研究中深入探讨CO2在黄河流域植被光合动态变化中的具体作用机制。
4 结论本研究深入分析了黄河流域植被光合作用的时空变化特征, 揭示了SIF过去21年中的动态趋势, 并对气候变化和人类活动对植被光合的影响进行了深入剖析, 得到的结论如下:
(1) 2001—2021年黄河流域SIF总体上呈波动上升趋势, 在空间上呈现出稳定的西部和东南部高、北部低的分布格局。青海省的南部地区、甘肃省与四川省交界地区以及河南省的中部地区, 特别是关中平原、阿尼玛卿山、宁夏平原以及黄河三角洲地区的SIF年变化趋势呈现出显著退化的趋势, 这些地区是黄河流域植被光合脆弱区。
(2) 总体上温度与SIF呈负相关, 而降水和太阳辐射对SIF呈正相关。在四川省西北部等地区, 温度与植被光合呈显著负相关关系。陕西省中部和河南省西部的山区的植被光合与降水呈现正相关关系。黄土高原、关中盆地以及运城盆地等地的植被光合与太阳辐射具有正相关关系。黄河流域SIF对气候变化的响应因地区而异, 这可能与地形地貌、气候类型和植被类型有关。
(3) 气候变化和人类活动对黄河流域植被光合动态的影响具有双重效应, 整体上以改善为主。受气候变化和人类活动双重驱动改善的区域面积占比为85.14%。在植被改善区域中, 人类活动对植被光合作用的贡献远大于气候变化。但在植被退化区域中, 稀树草原和农田中的气候变化贡献大于人类活动, 林地中的气候变化贡献与人类活动相当, 草地、灌木和湿地中的人类活动贡献大于气候变化。
| [1] |
Misal H, Hoare V C, Miles V. Responding to the climate crisis-taking action on the IPCC 6th Assessment Report. Weather, 2022, 77(4): 149-150. DOI:10.1002/wea.4162 |
| [2] |
姜田亮, 粟晓玲, 郭盛明, 吴海江. 西北地区植被耗水量的时空变化规律及其对气象干旱的响应. 水利学报, 2021, 52(2): 229-240. |
| [3] |
王浩, 胡鹏. 水循环视角下的黄河流域生态保护关键问题. 水利学报, 2020, 51(9): 1009-1014. |
| [4] |
Bai X Y, Zhang S R, Li C J, Xiong L, Song F J, Du C C, Li M H, Luo Q, Xue Y Y, Wang S J. A carbon-neutrality-capacity index for evaluating carbon sink contributions. Environmental Science and Ecotechnology, 2023, 15: 100237. DOI:10.1016/j.ese.2023.100237 |
| [5] |
Piao S L, Yue C, Ding J Z, Guo Z T. Perspectives on the role of terrestrial ecosystems in the 'carbon neutrality' strategy. Science China Earth Sciences, 2022, 65(6): 1178-1186. DOI:10.1007/s11430-022-9926-6 |
| [6] |
Ni Y X, Yu Z B, Lv X Z, Qin T L, Yan D H, Zhang Q F, Ma L. Spatial difference analysis of the runoff evolution attribution in the Yellow River Basin. Journal of Hydrology, 2022, 612: 128149. DOI:10.1016/j.jhydrol.2022.128149 |
| [7] |
Wu Q J, Zhu J F, Zhao X D. Effects of human social-economic activities on vegetation suitability in the Yellow River Basin, China. Forests, 2023, 14(2): 234. DOI:10.3390/f14020234 |
| [8] |
Ren Y Q, Liu J P, Liu S X, Wang Z G, Liu T, Shalamzari M J. Effects of climate change on vegetation growth in the Yellow River Basin from 2000 to 2019. Remote Sensing, 2022, 14(3): 687. DOI:10.3390/rs14030687 |
| [9] |
陈晨, 王义民, 黎云云, 周帅. 黄河流域1982—2015年不同气候区植被时空变化特征及其影响因素. 长江科学院院报, 2022, 39(2): 56-62, 81. |
| [10] |
Han J X, Zhang X Y, Wang J H, Zhai J Q. Geographic exploration of the driving forces of the NDVI spatial differentiation in the upper Yellow River Basin from 2000 to 2020. Sustainability, 2023, 15(3): 1922. DOI:10.3390/su15031922 |
| [11] |
田智慧, 任祖光, 魏海涛. 2000—2020年黄河流域植被时空演化驱动机制. 环境科学, 2022, 43(2): 743-751. |
| [12] |
Meroni M, Rossini M, Picchi V, Panigada C, Cogliati S, Nali C, Colombo R. Assessing steady-state fluorescence and PRI from hyperspectral proximal sensing as early indicators of plant stress: the case of ozone exposure. Sensors, 2008, 8(3): 1740-1754. DOI:10.3390/s8031740 |
| [13] |
van Leeuwen W J D, Orr B J, Marsh S E, Herrmann S M. Multi-sensor NDVI data continuity: uncertainties and implications for vegetation monitoring applications. Remote Sensing of Environment, 2006, 100(1): 67-81. DOI:10.1016/j.rse.2005.10.002 |
| [14] |
Gupana R S, Odermatt D, Cesana I, Giardino C, Nedbal L, Damm A. Remote sensing of Sun-induced chlorophyll-a fluorescence in inland and coastal waters: current state and future prospects. Remote Sensing of Environment, 2021, 262: 112482. DOI:10.1016/j.rse.2021.112482 |
| [15] |
Si H X, Wang R Y, Wang R H, He Z X. Can the MODIS data achieve the downscaling of GOME-2 SIF? validation of data from China. Sustainability, 2023, 15(7): 5920. DOI:10.3390/su15075920 |
| [16] |
Umuhoza J, Jiapaer G, Tao Y, Jiang L L, Zhang L C, Gasirabo A, Umwali E D, Umugwaneza A. Analysis of fluctuations in vegetation dynamic over Africa using satellite data of solar-induced chlorophyll fluorescence. Ecological Indicators, 2023, 146: 109846. DOI:10.1016/j.ecolind.2022.109846 |
| [17] |
Lu X L, Liu Z Q, Zhou Y Y, Liu Y L, An S Q, Tang J W. Comparison of phenology estimated from reflectance-based indices and solar-induced chlorophyll fluorescence (SIF) observations in a temperate forest using GPP-based phenology as the standard. Remote Sensing, 2018, 10(6): 932. DOI:10.3390/rs10060932 |
| [18] |
Song L, Guanter L, Guan K Y, You L Z, Huete A, Ju W M, Zhang Y G. Satellite Sun-induced chlorophyll fluorescence detects early response of winter wheat to heat stress in the Indian Indo-Gangetic Plains. Global Change Biology, 2018, 24(9): 4023-4037. DOI:10.1111/gcb.14302 |
| [19] |
Yuan Z, Yan D H, Yang Z Y, Yin J, Yuan Y. Temporal and spatial variability of drought in Huang-Huai-Hai River Basin, China. Theoretical and Applied Climatology, 2015, 122(3): 755-769. |
| [20] |
Li X, Xiao J F. A global, 0.05-degree product of solar-induced chlorophyll fluorescence derived from OCO-2, MODIS, and reanalysis data. Remote Sensing, 2019, 11(5): 517. DOI:10.3390/rs11050517 |
| [21] |
Zhong R, Wang P F, Mao G Q, Chen A F, Liu J G. Spatiotemporal variation of enhanced vegetation index in the Amazon Basin and its response to climate change. Physics and Chemistry of the Earth, Parts A/B/C, 2021, 123: 103024. DOI:10.1016/j.pce.2021.103024 |
| [22] |
Evans J, Geerken R. Discrimination between climate and human-induced dryland degradation. Journal of Arid Environments, 2004, 57(4): 535-554. DOI:10.1016/S0140-1963(03)00121-6 |
| [23] |
Li A, Wu J G, Huang J H. Distinguishing between human-induced and climate-driven vegetation changes: a critical application of RESTREND in Inner Mongolia. Landscape Ecology, 2012, 27(7): 969-982. DOI:10.1007/s10980-012-9751-2 |
| [24] |
Lu C P, Hou M C, Liu Z L, Li H J, Lu C Y. Variation characteristic of NDVI and its response to climate change in the middle and upper reaches of Yellow River Basin, China. IEEE Journal of Selected Topics in Applied Earth Observations and Remote Sensing, 2021, 14: 8484-8496. DOI:10.1109/JSTARS.2021.3105897 |
| [25] |
Tian F, Liu L Z, Yang J H, Wu J J. Vegetation greening in more than 94% of the Yellow River Basin (YRB) region in China during the 21st century caused jointly by warming and anthropogenic activities. Ecological Indicators, 2021, 125: 107479. |
| [26] |
孙瑞, 张方敏, 翁升恒, 刘倩. 2001—2021年中国NDVI时空格局变化及对气候的响应. 中国环境科学, 2023, 43(10): 5519-5528. |
| [27] |
Peng S S, Piao S L, Ciais P, Myneni R B, Chen A P, Chevallier F, Dolman A J, Janssens I A, Peñuelas J, Zhang G X, Vicca S, Wan S Q, Wang S P, Zeng H. Asymmetric effects of daytime and night-time warming on Northern Hemisphere vegetation. Nature, 2013, 501: 88-92. |
| [28] |
Liu Y C, Li Z, Chen Y N. Continuous warming shift greening towards browning in the Southeast and Northwest High Mountain Asia. Scientific Reports, 2021, 11: 17920. |
| [29] |
Dai Q, Cui C F, Wang S. Spatiotemporal variation and sustainability of NDVI in the Yellow River Basin. Irrigation and Drainage, 2022, 71(5): 1304-1318. |
| [30] |
程昌武. 黄河流域1982—2015年植被变化及其影响因素分析[D]. 西安: 长安大学, 2020.
|
| [31] |
郑子彦, 吕美霞, 马柱国. 黄河源区气候水文和植被覆盖变化及面临问题的对策建议. 中国科学院院刊, 2020, 35(1): 61-72. |